The Potential Protective Effects of Polyphenols in Asbestos-Mediated Inflammation and Carcinogenesis of Mesothelium


 ,
, 

Abstract
:1. Introduction
2. Polyphenols
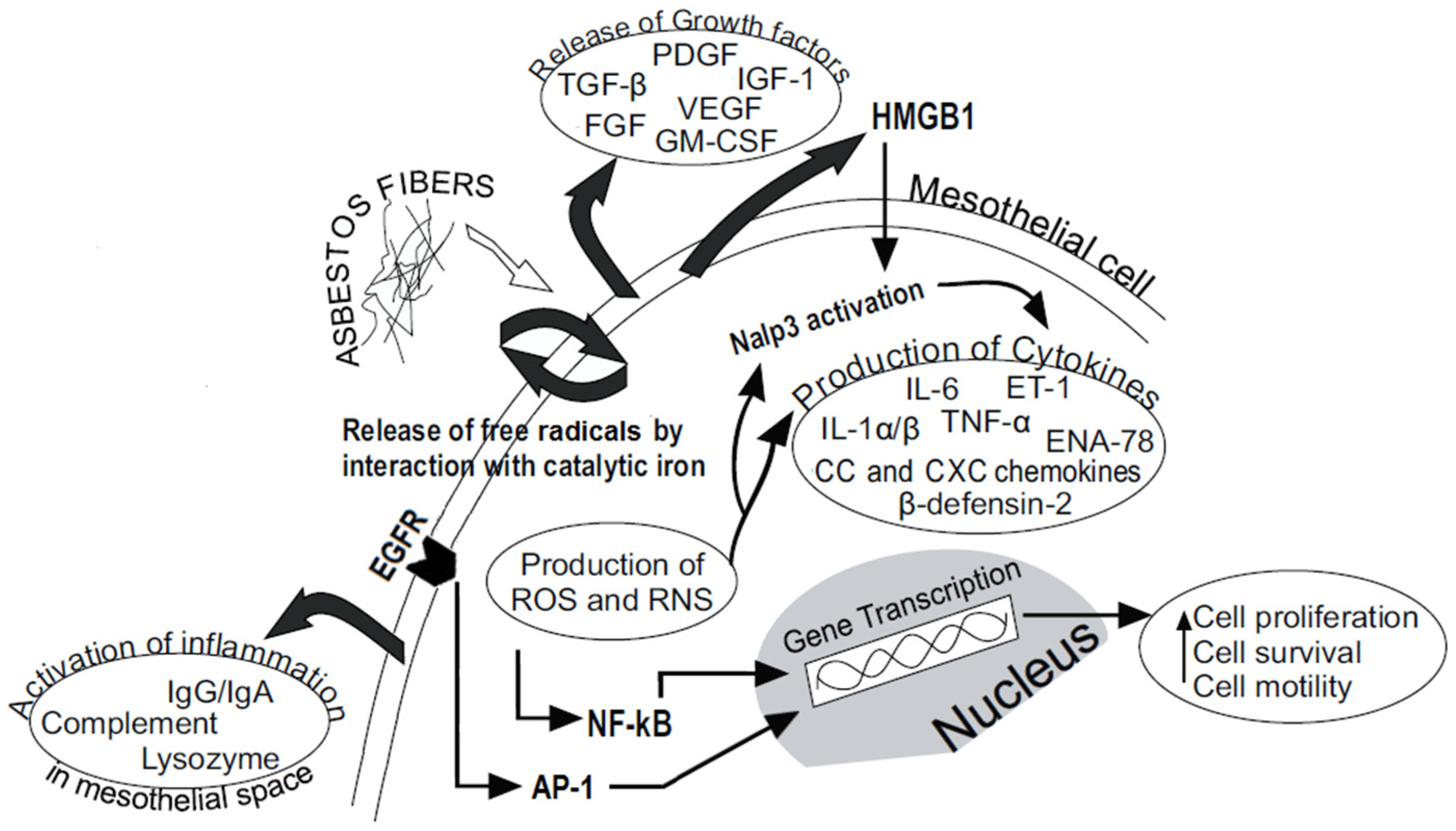
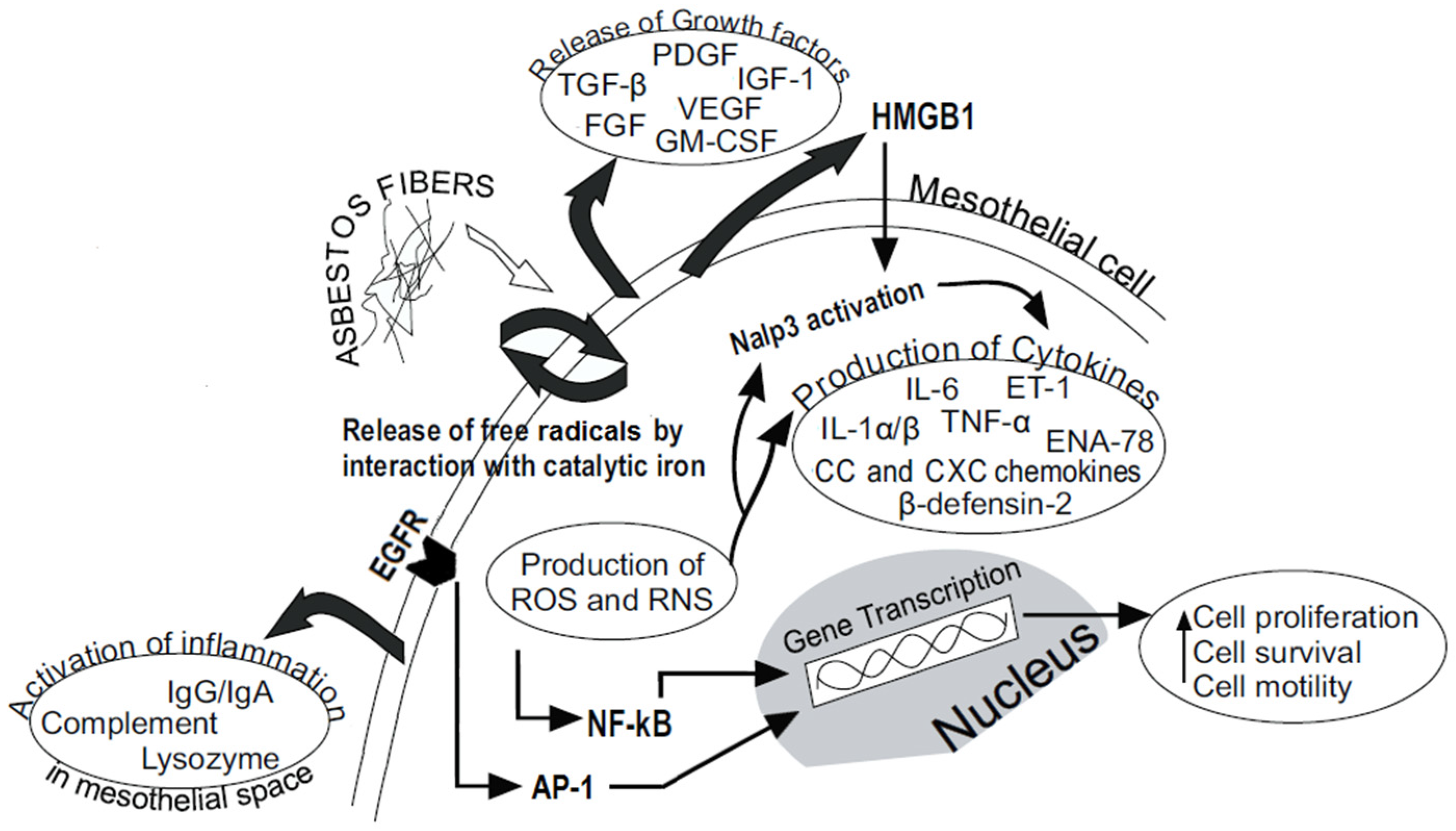
3. Asbestos Fibers and MM
4. Chronic Inflammation Affects MM Development
4.1. Overproduction of ROS and RNS
4.2. Inflammasome Activation and Cytokines Secretion
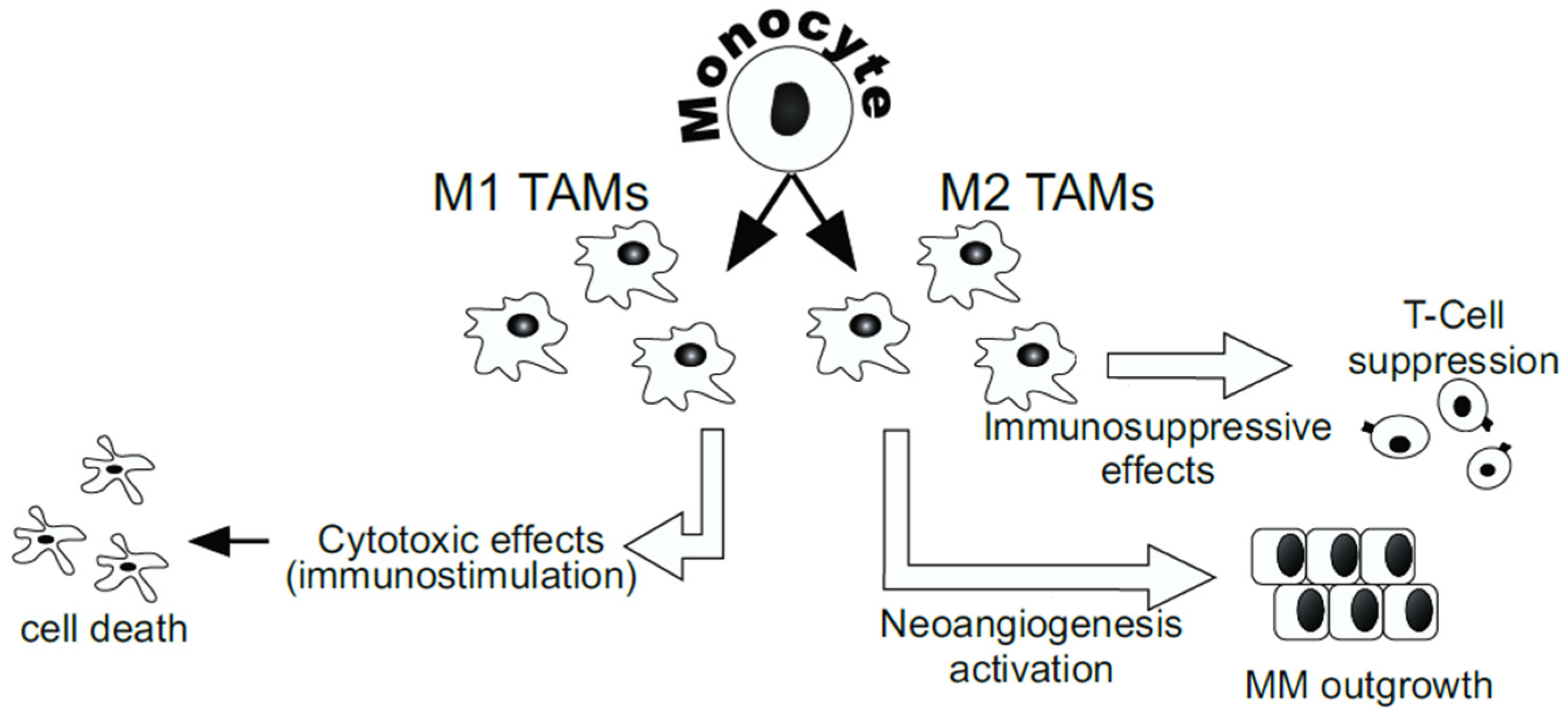
4.3. Innate Immunity and Cytokines in the Development of MM: MM-Driven Immunoediting
5. Effects of Polyphenols in MM
5.1. Effects of Polyphenols on ROS in MM
5.2. Effects of Polyphenols on Mediators of Inflammation in MM
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Carbone, M.; Ly, B.H.; Dodson, R.F.; Pagano, I.; Morris, P.T.; Dogan, U.A.; Gazdar, A.F.; Pass, H.I.; Yang, H. Malignant mesothelioma: Facts, myths, and hypotheses. J. Cell Physiol. 2012, 227, 44–58. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Rivera, Z.; Jube, S.; Nasu, M.; Bertino, P.; Goparaju, C.; Franzoso, G.; Lotze, M.T.; Krausz, T.; Pass, H.I.; et al. Programmed necrosis induced by asbestos in human mesothelial cells causes high-mobility group box 1 protein release and resultant inflammation. Proc. Nat. Acad. Sci. USA 2010, 107, 12611–12616. [Google Scholar] [CrossRef] [PubMed]
- Jube, S.; Rivera, Z.S.; Bianchi, M.E.; Powers, A.; Wang, E.; Pagano, I.; Pass, H.I.; Gaudino, G.; Carbone, M.; Yang, H. Cancer cell secretion of the DAMP protein HMGB1 supports progression in malignant mesothelioma. Cancer Res. 2012, 72, 3290–3301. [Google Scholar] [CrossRef] [PubMed]
- Napolitano, A.; Antoine, D.J.; Pellegrini, L.; Baumann, F.; Pagano, I.; Pastorino, S.; Goparaju, C.M.; Prokrym, K.; Canino, C.; Pass, H.I.; et al. HMGB1 and its hyperacetylated isoform are sensitive and specific serum biomarkers to detect asbestos exposure and to identify mesothelioma patients. Clin. Cancer Res. 2016. [Google Scholar] [CrossRef] [PubMed]
- Remon, J.; Lianes, P.; Martinez, S.; Velasco, M.; Querol, R.; Zanui, M. Malignant mesothelioma: New insights into a rare disease. Cancer Treat. Rev. 2013, 39, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Faig, J.; Howard, S.; Levine, E.A.; Casselman, G.; Hesdorffer, M.; Ohar, J.A. Changing Pattern in Malignant Mesothelioma Survival. Transl. Oncol. 2015, 8, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Testa, J.R.; Cheung, M.; Pei, J.; Below, J.E.; Tan, Y.; Sementino, E.; Cox, N.J.; Dogan, A.U.; Pass, H.I.; Trusa, S.; et al. Germline BAP1 mutations predispose to malignant mesothelioma. Nat. Genet. 2011, 43, 1022–1025. [Google Scholar] [CrossRef] [PubMed]
- Astoul, P.; Roca, E.; Galateau-Salle, F.; Scherpereel, A. Malignant pleural mesothelioma: From the bench to the bedside. Respiration 2012, 83, 481–493. [Google Scholar] [CrossRef] [PubMed]
- Izzi, V.; Masuelli, L.; Tresoldi, I.; Foti, C.; Modesti, A.; Bei, R. Immunity and malignant mesothelioma: From mesothelial cell damage to tumor development and immune response-based therapies. Cancer Lett. 2012, 322, 18–34. [Google Scholar] [CrossRef] [PubMed]
- Menges, C.W.; Chen, Y.; Mossman, B.T.; Chernoff, J.; Yeung, A.T.; Testa, J.R. A phosphotyrosine proteomic screen identifies multiple tyrosine kinase signaling pathways aberrantly activated in malignant mesothelioma. Genes Cancer 2010, 1, 493–505. [Google Scholar] [CrossRef] [PubMed]
- Albonici, L.; Palumbo, C.; Manzari, V. Role of inflammation and angiogenic growth factors in malignant mesothelioma. In Malignant Mesothelioma; Belli, C., Ed.; InTech: Rijeka, Croatia, 2012. [Google Scholar]
- Bajaj, G.; Yeo, Y. Drug delivery systems for intraperitoneal therapy. Pharm. Res. 2010, 27, 735–738. [Google Scholar] [CrossRef] [PubMed]
- Benvenuto, M.; Fantini, M.; Masuelli, L.; de Smaele, E.; Zazzeroni, F.; Tresoldi, I.; Calabrese, G.; Galvano, F.; Modesti, A.; Bei, R. Inhibition of ErbB receptors, Hedgehog and NF-kappaB signaling by polyphenols in cancer. Front. Biosci. 2013, 18, 1290–1310. [Google Scholar]
- Ghiringhelli, F.; Rebe, C.; Hichami, A.; Delmas, D. Immunomodulation and anti-inflammatory roles of polyphenols as anticancer agents. Anticancer Agents Med. Chem. 2012, 12, 852–873. [Google Scholar] [CrossRef] [PubMed]
- Gonzáles, R.; Ballester, I.; López-Posadas, R.; Suárez, M.D.; Zarzuelo, A.; Martínez-Augustin, O.; de Medina, F.S. Effects of flavonoids and other polyphenols on inflammation. Crit. Rev. Food Sci. Nutr. 2011, 51, 331–362. [Google Scholar] [CrossRef] [PubMed]
- Santangelo, C.; Vari, R.; Scazzocchio, B.; di Benedetto, R.; Filesi, C.; Masella, R. Polyphenols, intracellular signalling and inflammation. Ann. Ist. Super. Sanità 2007, 43, 394–405. [Google Scholar] [PubMed]
- Cuevas, A.; Saavedra, N.; Salazar, L.A.; Abdalla, D.S. Modulation of immune function by polyphenols: Possible contribution of epigenetic factors. Nutrients 2013, 5, 2314–2332. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [PubMed]
- Scalbert, A.; Manach, C.; Morand, C.; Rémésy, C.; Jiménez, L. Dietary polyphenols and the prevention of diseases. Crit. Rev. Food Sci. Nutr. 2005, 45, 287–306. [Google Scholar] [CrossRef] [PubMed]
- Marzocchella, L.; Fantini, M.; Benvenuto, M.; Masuelli, L.; Tresoldi, I.; Modest, A.; Bei, R. Dietary flavonoids: Molecular mechanisms of action as anti- inflammatory agents. Recent Patent Inflamm. Allergy Drug Disc. 2011, 5, 200–220. [Google Scholar] [CrossRef]
- Crozier, A.; Jaganath, I.B.; Clifford, M.N. Dietary phenolics: Chemistry, bioavailability and effects on health. Nat. Prod. Rep. 2009, 26, 1001–1043. [Google Scholar] [CrossRef] [PubMed]
- Fantini, M.; Benvenuto, M.; Masuelli, L.; Frajese, G.V.; Tresoldi, I.; Modesti, A.; Bei, R. In vitro and in vivo antitumoral effects of combinations of polyphenols, or polyphenols and anticancer drugs: Perspectives on cancer treatment. Int. J. Mol. Sci. 2015, 16, 9236–9282. [Google Scholar] [CrossRef] [PubMed]
- Bei, R.; Masuelli, L.; Turriziani, M.; Li Volti, G.; Malaguarnera, M.; Galvano, F. Impaired expression and function of signaling pathway enzymes by anthocyanins: Role on cancer prevention and progression. Curr. Enzym. Inhib. 2009, 5, 184–197. [Google Scholar] [CrossRef]
- Tomás-Barberán, F.A.; Clifford, M.N. Flavanones, chalcones and dihydrochalcones-nature, occurrence and dietary burden. J. Sci. Food Agric. 2000, 80, 1073–1080. [Google Scholar] [CrossRef]
- Cassidy, A.; Hanley, B.; Lamuela-Raventos, R.M. Isoflavones, lignans and stilbenes-origins, metabolism and potential importance to human health. J. Sci. Food Agric. 2000, 80, 1044–1062. [Google Scholar] [CrossRef]
- Prasad, S.; Tyagi, A.K.; Aggarwal, B.B. Recent developments in delivery, bioavailability, absorption and metabolism of curcumin: The golden pigment from golden spice. Cancer Res. Treat. 2014, 46, 2–18. [Google Scholar] [CrossRef] [PubMed]
- García-Lafuente, A.; Guillamón, E.; Villares, A.; Rostagno, M.A.; Martínez, J.A. Flavonoids as anti-inflammatory agents: Implications in cancer and cardiovascular disease. Inflamm. Res. 2009, 58, 537–552. [Google Scholar] [CrossRef] [PubMed]
- Izzi, V.; Masuelli, L.; Tresoldi, I.; Sacchetti, P.; Modesti, A.; Galvano, F.; Bei, R. The effects of dietary flavonoids on the regulation of redox inflammatory networks. Front. Biosci. 2012, 17, 2396–2418. [Google Scholar] [CrossRef]
- Edwards, S.W. The O-2 Generating NADPH Oxidase of Phagocytes: Structure and Methods of Detection. Methods 1996, 9, 563–577. [Google Scholar] [CrossRef] [PubMed]
- Cotelle, N. Role of flavonoids in oxidative stress. Curr. Top. Med. Chem. 2001, 1, 569–590. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [PubMed]
- Ginsburg, I.; Kohen, R.; Koren, E. Microbial and host cells acquire enhanced oxidant-scavenging abilities by binding polyphenols. Arch. Biochem. Biophys. 2011, 506, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Bohn, T. Dietary factors affecting polyphenol bioavailability. Nutr. Rev. 2014, 72, 429–452. [Google Scholar] [CrossRef] [PubMed]
- Masuelli, L.; Fantini, M.; Benvenuto, M.; Sacchetti, P.; Giganti, M.G.; Tresoldi, I.; Lido, P.; Lista, F.; Cavallo, F.; Nanni, P.; et al. Intratumoral delivery of recombinant vaccinia virus encoding for ErbB2/Neu inhibits the growth of salivary gland carcinoma cells. J. Transl. Med. 2014, 12. [Google Scholar] [CrossRef] [PubMed]
- Masuelli, L.; Marzocchella, L.; Focaccetti, C.; Lista, F.; Nardi, A.; Scardino, A.; Mattei, M.; Turriziani, M.; Modesti, M.; Forni, G.; et al. Local delivery of recombinant vaccinia virus encoding for neu counteracts growth of mammary tumors more efficiently than systemic delivery in neu transgenic mice. Cancer Immunol. Immunother. 2010, 59, 1247–1258. [Google Scholar] [CrossRef] [PubMed]
- Galanis, E.; Russell, S. Cancer gene therapy clinical trials: Lessons for the future. Br. J. Cancer 2001, 85, 1432–1436. [Google Scholar] [CrossRef] [PubMed]
- Forsyth, P.; Roldán, G.; George, D.; Wallace, C.; Palmer, C.A.; Morris, D.; Cairncross, G.; Matthews, M.V.; Markert, J.; Gillespie, Y.; et al. A phase I trial of intratumoral administration of reovirus in patients with histologically confirmed recurrent malignant gliomas. Mol. Ther. 2008, 16, 627–632. [Google Scholar] [CrossRef] [PubMed]
- Roberts, N.J.; Zhang, L.; Janku, F.; Collins, A.; Bai, R.Y.; Staedtke, V.; Rusk, A.W.; Tung, D.; Miller, M.; Roix, J.; et al. Intratumoral injection of Clostridium novyi-NT spores induces antitumor responses. Sci. Transl. Med. 2014, 6. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, S.; Wada, H.; Miyata, H.; Kawada, J.; Kawabata, R.; Nishikawa, H.; Gnjatic, S.; Sedrak, C.; Sato, E.; Nakamura, Y.; et al. Clinical trial of the intratumoral administration of labeled DC combined with systemic chemotherapy for esophageal cancer. J. Immunother. 2012, 35, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Masuelli, L.; Pantanella, F.; la Regina, G.; Benvenuto, M.; Fantini, M.; Mattera, R.; Di Stefano, E.; Mattei, M.; Silvestri, R.; Schippa, S.; et al. Violacein, an indole-derived purple-colored natural pigment produced by Janthinobacterium lividum, inhibits the growth of head and neck carcinoma cell lines both in vitro and in vivo. Tumour Biol. 2016, 37, 3705–3717. [Google Scholar] [CrossRef] [PubMed]
- Price, B.; Ware, A. Time trend of mesothelioma incidence in the United States and projection of future cases: An update based on SEER data for 1973 through 2005. Crit. Rev. Toxicol. 2009, 39, 576–588. [Google Scholar] [CrossRef] [PubMed]
- Pietrofesa, R.A.; Velalopoulou, A.; Arguiri, E.; Menges, C.W.; Testa, J.R.; Hwang, W.T.; Albelda, S.M.; Christofidou-Solomidou, M. Flaxseed lignans enriched in secoisolariciresinol diglucoside prevent acute asbestos-induced peritoneal inflammation in mice. Carcinogenesis 2016, 37, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Chew, S.H.; Toyokuni, S. Malignant mesothelioma as an oxidative stress-induced cancer: An update. Free Radic. Biol. Med. 2015, 86, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Otsuki, T.; Maeda, M.; Murakami, S.; Hayashi, H.; Miura, Y.; Kusaka, M.; Nakano, T.; Fukuoka, K.; Kishimoto, T.; Hyodoh, F.; et al. Immunological effects of silica and asbestos. Cell. Mol. Immunol. 2007, 4, 261–268. [Google Scholar] [PubMed]
- Miserocchi, G.; Sancini, G.; Mantegazza, F.; Chiappino, G. Translocation pathways for inhaled asbestos fibers. Environ. Health 2008, 7. [Google Scholar] [CrossRef] [PubMed]
- Tunesi, S.; Ferrante, D.; Mirabelli, D.; Andorno, S.; Betti, M.; Fiorito, G.; Guarrera, S.; Casalone, E.; Neri, M.; Ugolini, D.; et al. Gene-asbestos interaction in malignant pleural mesothelioma susceptibility. Carcinogenesis 2015, 36, 1129–1135. [Google Scholar] [CrossRef] [PubMed]
- Jantz, M.A.; Antony, V.B. Pathophysiology of the pleura. Respiration 2008, 75, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Carbone, M.; Bedrossian, C.W. The pathogenesis of mesothelioma. Semin. Diagn. Pathol. 2006, 23, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Acencio, M.M.; Soares, B.; Marchi, E.; Silva, C.S.; Teixeira, L.R.; Broaddus, V.C. Inflammatory cytokines contribute to asbestos-induced injury of mesothelial cells. Lung 2015, 193, 831–837. [Google Scholar] [CrossRef] [PubMed]
- Carbone, M.; Yang, H. Molecular pathways: Targeting mechanisms of asbestos and erionite carcinogenesis in mesothelioma. Clin. Cancer Res. 2012, 18, 598–604. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, S.; Nuvoli, B.; Catalani, S.; Galati, R. Reactive oxygen species a double-edged sword for mesothelioma. Oncotarget 2015, 6, 16848–16865. [Google Scholar] [CrossRef] [PubMed]
- Kamp, D.W.; Weitzman, S.A. The molecular basis of asbestos induced lung injury. Thorax 1999, 54, 638–652. [Google Scholar] [CrossRef] [PubMed]
- Kinnula, V.L.; Aalto, K.; Raivio, K.O.; Walles, S.; Linnainmaa, K. Cytotoxicity of oxidants and asbestos fibers in cultured human mesothelial cells. Free Radic. Biol. Med. 1994, 16, 169–176. [Google Scholar] [CrossRef]
- Kinnula, V.L.; Raivio, K.O.; Linnainmaa, K.; Ekman, A.; Klockars, M. Neutrophil and asbestos fiber-induced cytotoxicity in cultured human mesothelial and bronchial epithelial cells. Free Radic. Biol. Med. 1995, 18, 391–399. [Google Scholar] [CrossRef]
- Moslen, M.T. Reactive oxygen species in normal physiology, cell injury and phagocytosis. Adv. Exp. Med. Biol. 1994, 366, 17–27. [Google Scholar] [PubMed]
- Hansen, K.; Mossman, B.T. Generation of superoxide (O2-.) from alveolar macrophages exposed to asbestiform and nonfibrous particles. Cancer Res. 1987, 47, 1681–1686. [Google Scholar] [PubMed]
- Shukla, A.; Gulumian, M.; Hei, T.K.; Kamp, D.; Rahman, Q.; Mossman, B.T. Multiple roles of oxidants in the pathogenesis of asbestos-induced diseases. Free Radic. Biol. Med. 2003, 34, 1117–1129. [Google Scholar] [CrossRef]
- Funahashi, S.; Okazaki, Y.; Ito, D.; Asakawa, A.; Nagai, H.; Tajima, M.; Toyokuni, S. Asbestos and multi-walled carbon nanotubes generate distinct oxidative responses in inflammatory cells. J. Clin. Biochem. Nutr. 2015, 56, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Choe, N.; Hemenway, D.R.; Zhu, S.; Matalon, S.; Kagan, E. Asbestos inhalation induces reactive nitrogen species and nitrotyrosine formation in the lungs and pleura of the rat. J. Clin. Investig. 1998, 102, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Soini, Y.; Kahlos, K.; Puhakka, A.; Lakari, E.; Säily, M.; Pääkkö, P.; Kinnula, V. Expression of inducible nitric oxide synthase in healthy pleura and in malignant mesothelioma. Br. J. Cancer 2000, 83, 880–886. [Google Scholar] [CrossRef] [PubMed]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Liu, W.; Koenig, K.; Idell, S.; Broaddus, V.C. Vitronectin adsorption to chrysotile asbestos increases fiber phagocytosis and toxicity for mesothelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 279, L916–L923. [Google Scholar] [PubMed]
- Hillegass, J.M.; Miller, J.M.; MacPherson, M.B.; Westbom, C.M.; Sayan, M.; Thompson, J.K.; Macura, S.L.; Perkins, T.N.; Beuschel, S.L.; Alexeeva, V.; et al. Asbestos and erionite prime and activate the NLRP3 inflammasome that stimulates autocrine cytokine release in human mesothelial cells. Part. Fibre Toxicol. 2013, 10, 39. [Google Scholar] [CrossRef] [PubMed]
- Chow, M.T.; Tschopp, J.; Möller, A.; Smyth, M.J. NLRP3 promotes inflammation-induced skin cancer but is dispensable for asbestos-induced mesothelioma. Immunol. Cell Biol. 2012, 90, 983–986. [Google Scholar] [CrossRef] [PubMed]
- Petrilli, V.; Dostert, C.; Muruve, D.A.; Tschopp, J. The inflammasome: A danger sensing complex triggering innate immunity. Curr. Opin. Immunol. 2007, 19, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Westbom, C.; Thompson, J.K.; Leggett, A.; MacPherson, M.; Beuschel, S.; Pass, H.; Vacek, P.; Shukla, A. Inflammasome modulation by chemotherapeutics in malignant mesothelioma. PLoS ONE 2015, 10, e0145404. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Faux, S.P.; Hallden, G.; Kirn, D.H.; Houghton, C.E.; Lemoine, N.R.; Patrick, G. Interleukin-1beta and tumour necrosis factor-alpha promote the transformation of human immortalised mesothelial cells by erionite. Int. J. Oncol. 2004, 25, 173–178. [Google Scholar] [PubMed]
- Antony, V.B. Immunological mechanisms in pleural disease. Eur. Respir. J. 2003, 21, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Driscoll, K.E.; Maurer, J.K.; Higgins, J.; Poynter, J. Alveolar macrophage cytokine and growth factor production in a rat model of crocidolite-induced pulmonary inflammation and fibrosis. J. Toxicol. Environ. Health 1995, 46, 155–169. [Google Scholar] [CrossRef] [PubMed]
- Driscoll, K.E.; Carter, J.M.; Howard, B.W.; Hassenbein, D.; Janssen, Y.M.; Mossman, B.T. Crocidolite activates NF-kappa B and MIP-2 gene expression in rat alveolar epithelial cells. Role of mitochondrial-derived oxidants. Environ. Health Perspect. 1998, 106, 1171–1174. [Google Scholar] [CrossRef] [PubMed]
- Bielefeldt-Ohmann, H.; Fitzpatrick, D.R.; Marzo, A.L.; Jarnicki, A.G.; Himbeck, R.P.; Davis, M.R.; Manning, L.S.; Robinson, B.W. Patho- and immunobiology of malignant mesothelioma: Characterisation of tumour infiltrating leucocytes and cytokine production in a murine model. Cancer Immunol. Immunother. 1994, 39, 347–359. [Google Scholar] [CrossRef] [PubMed]
- Landskron, G.; de la Fuente, M.; Thuwajit, P.; Thuwajit, C.; Hermoso, M.A. Chronic inflammation and cytokines in the tumor microenvironment. J. Immunol. Res. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Caja, F.; Vannucci, L. TGFβ: A player on multiple fronts in the tumor microenvironment. J. Immunotoxicol. 2015, 12, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Zitvogel, L.; Kepp, O.; Galluzzi, L.; Kroemer, G. Inflammasomes in carcinogenesis and anticancer immune responses. Nat. Immunol. 2012, 13, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Naldini, A.; Carraro, F. Role of inflammatory mediators in angiogenesis. Curr. Drug Targets Inflamm. Allergy 2005, 4, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Fox, S.A.; Loh, S.S.; Mahendran, S.K.; Garlepp, M.J. Regulated chemokine gene expression in mouse mesothelioma and mesothelial cells: TNF-α upregulates both CC and CXC chemokine genes. Oncol. Rep. 2012, 28, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Rollins, B.J. Inflammatory chemokines in cancer growth and progression. Eur. J. Cancer 2006, 42, 760–767. [Google Scholar] [CrossRef] [PubMed]
- Vlaeminck-Guillem, V.; Bienvenu, J.; Isaac, S.; Grangier, B.; Golfier, F.; Passot, G.; Bakrin, N.; RodriguezLafrasse, C.; Gilly, F.N.; Glehen, O. Intraperitoneal cytokine level in patients with peritoneal surface malignancies. A study of the RENAPE (French Network for Rare Peritoneal Malignancies). Ann. Surg. Oncol. 2013, 20, 2655–2662. [Google Scholar] [CrossRef] [PubMed]
- Comar, M.; Zanotta, N.; Bonotti, A.; Tognon, M.; Negro, C.; Cristaudo, A.; Bovenzi, M. Increased levels of C-C chemokine RANTES in asbestos exposed workers and in malignant mesothelioma patients from an hyperendemic area. PLoS ONE 2014, 9, e104848. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Alexander, D.B.; Iigo, M.; Hamano, H.; Takahashi, S.; Yokoyama, T.; Kato, M.; Usami, I.; Tokuyama, T.; Tsutsumi, M. Chemokine (C-C motif) ligand 3 detection in the serum of persons exposed to asbestos: A patient-based study. Cancer Sci. 2015, 106, 825–832. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Terlizzi, M.; Casolaro, V.; Pinto, A.; Sorrentino, R. Inflammasome: Cancer’s friend or foe? Pharmacol. Therap. 2014, 143, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Why not treat human cancer with interleukin-1 blockade? Cancer Metastasis Rev. 2010, 29, 317–329. [Google Scholar] [CrossRef] [PubMed]
- Izzi, V.; Chiurchiù, V.; Doldo, E.; Palumbo, C.; Tesoldi, I.; Bei, R.; Albonici, L.; Modesti, A. Interleukin-17 produced by malignant mesothelioma-polarized immune cells promotes tumor growth and invasiveness. Eur. J. Inflamm. 2013, 11, 203–214. [Google Scholar]
- Miselis, N.R.; Lau, B.W.; Wu, Z.; Kane, A.B. Kinetics of host cell recruitment during dissemination of diffuse malignant peritoneal mesothelioma. Cancer Microenviron. 2010, 4, 39–50. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Veltman, J.D.; Lambers, M.E.; van Nimwegen, M.; Hendriks, R.W.; Hoogsteden, H.C.; Hegmans, J.P.; Aerts, J.G. Zoledronic acid impairs myeloid differentiation to tumour-associated macrophages in mesothelioma. Br. J. Cancer 2010, 103, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Veltman, J.D.; Lambers, M.E.; van Nimwegen, M.; Hendriks, R.W.; Hoogsteden, H.C.; Aerts, J.G.; Hegmans, J.P. COX-2 inhibition improves immunotherapy and is associated with decreased numbers of myeloid-derived suppressor cells in mesothelioma. Celecoxib influences MDSC function. BMC Cancer 2010, 10, 464. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Pellegrini, L.; Napolitano, A.; Giorgi, C.; Jube, S.; Preti, A.; Jennings, C.J.; de Marchis, F.; Flores, E.G.; Larson, D.; et al. Aspirin delays mesothelioma growth by inhibiting HMGB1-mediated tumor progression. Cell Death Dis. 2015, 6, e1786. [Google Scholar] [CrossRef] [PubMed]
- Sica, A.; Allavena, P.; Mantovani, A. Cancer related inflammation: The macrophage connection. Cancer Lett. 2008, 267, 204–215. [Google Scholar] [CrossRef] [PubMed]
- Burt, B.M.; Rodig, S.J.; Tilleman, T.R.; Elbardissi, A.W.; Bueno, R.; Sugarbaker, D.J. Circulating and tumor-infiltrating myeloid cells predict survival in human pleural mesothelioma. Cancer 2011, 117, 5234–5244. [Google Scholar] [CrossRef] [PubMed]
- Hegmans, J.P.; Hemmes, A.; Hammad, H.; Boon, L.; Hoogsteden, H.C.; Lambrecht, B.N. Mesothelioma environment comprises cytokines and T-regulatory cells that suppress immune responses. Eur. Respir. J. 2006, 27, 1086–1095. [Google Scholar] [CrossRef] [PubMed]
- Miselis, N.R.; Wu, Z.J.; van Rooijen, N.; Kane, A.B. Targeting tumor-associated macrophages in an orthotopic murine model of diffuse malignant mesothelioma. Mol. Cancer Ther. 2008, 7, 788–799. [Google Scholar] [CrossRef] [PubMed]
- Napolitano, A.; Pellegrini, L.; Dey, A.; Larson, D.; Tanji, M.; Flores, E.G.; Kendrick, B.; Lapid, D.; Powers, A.; Kanodia, S.; et al. Minimal asbestos exposure in germline BAP1 heterozygous mice is associated with deregulated inflammatory response and increased risk of mesothelioma. Oncogene 2016, 35, 1996–2002. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, Y.; Kumagai-Takei, N.; Matsuzaki, H.; Lee, S.; Maeda, M.; Kishimoto, T.; Fukuoka, K.; Nakano, T.; Otsuki, T. Functional alteration of natural killer cells and cytotoxic T limphocytes upon asbestos exposure and in malignant mesothelioma patients. BioMed. Res. Int. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Maeda, M.; Nishimura, Y.; Hayashi, H.; Kumagai, N.; Chen, Y.; Murakami, S.; Miura, Y.; Hiratsuka, J.; Kishimoto, T.; Otsuki, T. Reduction of CXC chemokine receptor 3 in an in vitro model of continuous exposure to asbestos in a human T-cell line, MT-2. Am. J. Respira. Cell Mol. Biol. 2011, 45, 470–479. [Google Scholar] [CrossRef] [PubMed]
- Maeda, M.; Nishimura, Y.; Hayashi, H.; Kumagai, N.; Chen, Y.; Murakami, S.; Miura, Y.; Hiratsuka, J.; Kishimoto, T.; Otsuki, T. Decreased CXCR3 expression in CD4+ T cells exposed to asbestos or derived from asbestos-exposed patients. Am. J. Respira. Cell Mol. Biol. 2011, 45, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Maeda, M.; Chen, Y.; Hayashi, H.; Kumagai-Takei, N.; Matsuzaki, H.; Lee, S.; Nishimura, Y.; Otsuki, T. Chronic exposure to asbestos enhances TGF-β1 production in the human adult T cell leukemia virus-immortalized T cell line MT-2. Int. J. Oncol. 2014, 45, 2522–2532. [Google Scholar] [CrossRef] [PubMed]
- Miura, Y.; Nishimura, Y.; Katsuyama, H.; Maeda, M.; Hayashi, H.; Dong, M.; Hyodoh, F.; Tomita, M.; Matsuo, Y.; Uesaka, A.; et al. Involvement of IL-10 and Bcl-2 in resistance against an asbestos-induced apoptosis of T cells. Apoptosis 2006, 11, 1825–1835. [Google Scholar] [CrossRef] [PubMed]
- Feskanich, D.; Ziegler, R.G.; Michaud, D.S.; Giovannucci, E.L.; Speizer, F.E.; Willett, W.C.; Colditz, G.A. Prospective study of fruit and vegetable consumption and risk of lung cancer among men and women. J. Natl. Cancer Inst. 2000, 92, 1812–1823. [Google Scholar] [CrossRef] [PubMed]
- Bazzano, L.A.; He, J.; Ogden, L.G.; Loria, C.M.; Vupputuri, S.; Myers, L.; Whelton, P.K. Fruit and vegetable intake and risk of cardiovascular disease in US adults: The first National Health and Nutrition Examination Survey Epidemiologic follow-up study. Am. J. Clin. Nutr. 2002, 76, 93–99. [Google Scholar] [PubMed]
- Mennen, L.I.; Sapinho, D.; de Bree, A.; Arnault, N.; Bertrais, S.; Galan, P.; Hercberg, S. Consumption of foods rich in flavonoids is related to a decreased cardiovascular risk in apparently healthy French women. J. Nutr. 2004, 134, 923–926. [Google Scholar] [PubMed]
- Chang, H.W.; Baek, S.H.; Chung, K.W.; Son, K.H.; Kim, H.P.; Kang, S.S. Inactivation of phospholipase A2 by naturally occurring biflavonoid, ochnaflavone. Biochem. Biophys. Res. Commun. 1994, 205, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.K.; Ecklund, D.; Liu, M.; Datta, S.K. Apigenin, a non-mutagenic dietary flavonoid, suppresses lupus by inhibiting autoantigen presentation for expansion of autoreactive Th1 and Th17 cells. Arthritis Res. Ther. 2009, 11, R59. [Google Scholar] [CrossRef] [PubMed]
- Chi, Y.S.; Kim, H.P. Suppression of cyclooxigenase-2 expression of skin fibroblasts by wogonin, a plant flavones from Scutellaria radix. Prostaglandins Leukot. Essent. Fat. Acids 2005, 72, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.C.; Huang, Y.T.; Tsai, S.H.; Lin-Shiau, S.Y.; Chen, C.F.; Lin, J.K. Suppression of inducible cyclooxygenase and inducible nitric oxide synthase by apigenin and related flavonoids in mouse macrophages. Carcinogenesis 1999, 20, 1945–1952. [Google Scholar] [CrossRef] [PubMed]
- Hamalainen, M.; Nieminen, R.; Vuorela, P.; Heinonen, M.; Moilanen, E. Anti-inflammatory effects of flavonoids: Genistein, kaempferol, quercetin, and daidzein inhibit STAT-1 and NF-kappaB activations, whereas flavone, isorhamnetin, naringenin, and pelargonidin inhibit only NF-kappaB activation along with their inhibitory effect on iNOS expression and NO production in activated macrophages. Mediat. Inflamm. 2007, 2007. [Google Scholar] [CrossRef]
- Halliwell, B. Are polyphenols antioxidants or pro-oxidants? What do we learn from cell culture and in vivo studies? Arch. Biochem. Biophys. 2008, 476, 107–112. [Google Scholar] [CrossRef] [PubMed]
- León-Gonzáles, A.J.; Auger, C.; Schini-Kerth, V.B. Pro-oxidant activity of polyphenols and its implication on cancer chemoprevention and chemotherapy. Biochem. Pharmacol. 2015, 98, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.M.; Thompson, J.K.; MacPherson, M.B.; Beuschel, S.L.; Westbom, C.M.; Sayan, M.; Shukla, A. Curcumin: A double hit on malignant mesothelioma. Cancer Prev. Res. 2014, 7, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Satoh, M.; Takemura, Y.; Hamada, H.; Sekido, Y.; Kubota, S. EGCG indices human mesothelioma cell death by inducing reactive oxygen species and autophagy. Cancer Cell Int. 2013, 13, 19. [Google Scholar] [CrossRef] [PubMed]
- Ranzato, E.; Martinotti, S.; Magnelli, V.; Murer, B.; Biffo, S.; Mutti, L.; Burlando, B. Epigallocathechin-3-gallate induces mesothelioma cell death via H2 O2-dependent T-type Ca2+ channel opening. J. Cell. Mol. Med. 2012, 16, 2667–2678. [Google Scholar] [CrossRef] [PubMed]
- Valenti, D.; de Bari, L.; Manente, G.A.; Rossi, L.; Mutti, L.; Moro, L.; Vacca, R.A. Negative modulation of mitochondrial oxidative phosphorylation by epigallocatechin-3 gallate leads to growth arrest and apoptosis in human malignant pleural mesothelioma cells. Biochim. Biophys. Acta 2013, 1382, 2085–2096. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Im, J.H.; Lee, D.M.; Park, J.S.; Won, S.Y.; Cho, M.K.; Nam, H.S.; Lee, Y.J.; Lee, S.H. Synergistic inhibition of mesothelioma cell growth by the combination of clofarabine and resveratrol involves Nrf2 downregulation. BMB Rep. 2012, 45, 647–652. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Park, I.S.; Lee, Y.J.; Shim, J.H.; Cho, M.K.; Nam, H.S.; Park, J.W.; Oh, M.H.; Lee, S.H. Resveratrol contributes to chemosensivity of malignant mesothelioma cells with activation of p53. Food Chem. Toxicol. 2014, 63, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Faraonio, R.; Vergara, P.; Di Marzo, D.; Pierantoni, M.G.; Napolitano, M.; Russo, T.; Cimino, F. p53 suppresses the Nrf2-dependent transcription of antioxidant response genes. J. Biol. Chem. 2006, 281, 39776–39784. [Google Scholar] [CrossRef] [PubMed]
- Pietrofesa, R.A.; Velalopoulou, A.; Albelda, S.M.; Christofidou-Solomidou, M. Asbestos Induces Oxidative Stress and Activation of Nrf2 Signaling in Murine Macrophages: Chemopreventive Role of the Synthetic Lignan Secoisolariciresinol Diglucoside (LGM2605). Int. J. Mol. Sci. 2016, 17, 322. [Google Scholar] [CrossRef] [PubMed]
- Kostyuk, V.A.; Potapovich, A.I.; Speransky, S.D.; Maslova, G.T. Protective effect of natural flavonoids on rat peritoneal macrophages injury caused by asbestos fibers. Free Radic. Biol. Med. 1996, 21, 487–493. [Google Scholar] [CrossRef]
- Kostyuk, V.A.; Potapovich, A.I.; Vladykovskaya, E.N.; Hiramatsu, M. Protective effects of green tea catechins against asbestos-induced cell injury. Planta Med. 2000, 66, 762–764. [Google Scholar] [CrossRef] [PubMed]
- Potapovich, A.I.; Kostyuk, V.A. Comparative study of antioxidant properties and cytoprotective activity of flavonoids. Biochemistry 2003, 68, 514–519. [Google Scholar] [PubMed]
- Grivennikov, S.I.; Greten, F.R.; Karin, M. Immunity, inflammation, and cancer. Cell 2010, 140, 883–899. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.N.; Veena, M.S.; Srivatsan, E.S.; Wang, M.B. Suppression of interleukin 6 and 8 production in head and neck cancer cells with curcumin via inhibition of Ikappa beta kinase. Arch. Otolaryngol. Head Neck Surg. 2009, 135, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yu, C.; Pan, Y.; Yang, X.; Huang, Y.; Feng, Z.; Li, X.; Yang, S.; Liang, G. A novel synthetic mono-carbonyl analogue of curcumin, A13, exhibits anti-inflammatory effect in vivo by inhibition of inflammatory mediators. Inflammation 2011, 35, 594–604. [Google Scholar] [CrossRef] [PubMed]
- Serafini, M.; Peluso, I.; Raguzzini, A. Flavonoids as anti-inflammatory agents. Proc. Nutr. Soc. 2010, 69, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T.; Higa, S.; Arimitsu, J.; Naka, T.; Shima, Y.; Ohshima, S.; Fujimoto, M.; Yamadori, T.; Kawase, I.; Tanaka, T. Flavonoids such as luteolin, fisetin and apigenin are inhibitors of interleukin-4 and interleukin-13 production by activated human basophils. Int. Arch. Allergy Immunol. 2004, 134, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.C.; Wheeler, D.S.; Malhotra, V.; Odoms, K.; Denenberg, A.G.; Wong, H.R. A green tea-derived polyphenol, epigallocatechin-3-gallate, inhibits IKappaB kinase activation and IL-8 gene expression in respiratory epithelium. Inflammation 2002, 26, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.Y.; Kim, S.H.; Jeong, H.J.; Kim, S.Y.; Shin, T.Y.; Um, J.Y.; Hong, S.H.; Kim, H.M. Epigallocatechin-3- gallate inhibits secretion of TNF-alpha, IL-6 and IL-8 through the attenuation of ERK and NF-kappaB in HMC- 1 cells. Int. Arch. Allergy Immunol. 2007, 142, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Marotte, H.; Kwan, K.; Ruth, J.H.; Campbell, P.L.; Rabquer, B.J.; Pakozdi, A.; Koch, A.E. Epigallocatechin-3-gallate inhibits IL-6 synthesis and suppresses transsignaling by enhancing soluble gp130 production. Proc. Natl. Acad. Sci. USA 2008, 105, 14692–14697. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, Z.; Anbazhagan, A.N.; Akhtar, N.; Ramamurthy, S.; Voss, F.R.; Haqqi, T.M. Green tea polyphenol epigallocatechin-3-gallate inhibits advanced glycation end product-induced expression of tumor necrosis factor-alpha and matrix metalloproteinase-13 in human chondrocytes. Arthritis Res. Ther. 2009, 11, R71. [Google Scholar] [CrossRef] [PubMed]
- Park, H.H.; Lee, S.; Son, H.Y.; Park, S.B.; Kim, M.S.; Choi, E.J.; Singh, T.S.; Ha, J.H.; Lee, M.G.; Kim, J.E. Flavonoids inhibit histamine release and expression of proinflammatory cytokines in mast cells. Arch. Pharm. Res. 2008, 31, 1303–1311. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Hu, Y.; Liu, X.; Wu, T.; Wang, Y.; He, W.; Wei, W. Resveratrol downregulates the constitutional activation of nuclear factor-kappaB in multiple myeloma cells, leading to suppression of proliferation and invasion, arrest of cell cycle, and induction of apoptosis. Cancer Genet. Cytogenet. 2006, 165, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Manna, S.K.; Mukhopadhyay, A.; Aggarwal, B.B. Resveratrol suppresses TNF-induced activation of nuclear transcription factors NF-kappaB, activator protein-1, and apoptosis: Potential role of reactive oxygen intermediates and lipid peroxidation. J. Immunol. 2000, 164, 6509–6519. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.Y.; Kim, K.H.; Lee, S.H.; Yoon, M.S.; Lee, H.J.; Moon, D.O.; Lee, C.M.; Ahn, S.C.; Park, Y.C.; Park, Y.M. Curcumin inhibits immunostimulatory function of dendritic cells: MAPKs and translocation of NF-kappaB as potential targets. J. Immunol. 2005, 174, 8116–8124. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Mediavilla, M.V.; Crespo, I.; Collado, P.S.; Esteller, A.; Sanchez-Campos, S.; Tunon, M.J. Anti-inflammatory effect of the flavones quercetin and kaempferol in Chang Liver cells involves inhibition of inducible nitric oxide synthase, cyclooxygenase-2 and reactive C-protein, and down-regulation of the nuclear factor kappaB pathway. Eur. J. Pharmacol. 2007, 557, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, C.; Batra, S.; Vargo, M.A.; Voss, O.H.; Gavrilin, M.A.; Wewers, M.D.; Guttridge, D.C.; Grotewold, E.; Doseff, A.I. Apigenin blocks lipopolysaccharide-induced lethality in vivo and proinflammatory cytokines expression by inactivating NF-kappa B through the suppression of p65 phosphorylation. J. Immunol. 2007, 179, 7121–7127. [Google Scholar] [CrossRef] [PubMed]
- Romier, B.; van de Walle, J.; During, A.; Larondelle, Y.; Schneider, Y.J. Modulation of signaling nuclear factor-kappaB activation pathway by polyphenols in human intestinal Caco-2 cells. Br. J. Nutr. 2008, 100, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.W.; Chen, C.H.; Wang, Y.H.; Ho, M.L.; Hung, S.H.; Chen, I.S.; Wang, G.J. (−)-Epigallocatechin gallate inhibition of osteoclastic differentiation via NF-kappaB Biochem. Biophys. Res. Commun. 2009, 379, 1033–1037. [Google Scholar] [CrossRef] [PubMed]
- Laua, F.C.; Josepha, J.A.; McDonald, J.E.; Kalt, A.W. Attenuation of iNOS and COX2 by blueberry polyphenols is mediated through the suppression of NF-κB activation. J. Funct. Foods 2009, 1, 274–283. [Google Scholar] [CrossRef]
- Kim, J.W.; Jin, Y.C.; Kim, Y.M.; Rhie, S.; Kim, H.J.; Seo, H.G.; Lee, J.H.; Ha, Y.L.; Chang, K.C. Daidzein administration in vivo reduces myocardial injury in a rat ischemia/reperfusion model by inhibiting NF-κB activation. Life Sci. 2009, 84, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Bharti, A.C.; Donato, N.; Aggarwal, B.B. Curcumin (diferuloylmethane) inhibits constitutive and IL-6-inducible STAT3 phosphorylation in human multiple myeloma cells. J. Immunol. 2003, 171, 3863–3871. [Google Scholar] [CrossRef] [PubMed]
- Chakravarti, N.; Myers, J.N.; Aggarwal, B.B. Targeting constitutive and interleukin-6-inducible signal transducers and activators of transcription 3 pathway in head and neck squamous cell carcinoma cells by curcumin (diferuloylmethane). Int. J. Cancer 2006, 119, 1268–1275. [Google Scholar] [CrossRef] [PubMed]
- Wung, B.S.; Hsu, M.C.; Wu, C.C.; Hsieh, C.W. Resveratrol suppresses IL-6- induced ICAM-1 gene expression in endothelial cells: Effects on the inhibition of STAT3 phosphorylation. Life Sci. 2005, 78, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Masuda, M.; Suzui, M.; Weinstein, I.B. Effects of epigallocatechin-3- gallate on growth, epidermal growth factor receptor signaling pathways, gene expression, and chemosensitivity in human head and neck squamous cell carcinoma cell lines. Clin. Cancer Res. 2001, 7, 4220–4229. [Google Scholar] [PubMed]
- Kadariya, Y.; Menges, C.W.; Talarchek, J.; Cai, K.Q.; Klein-Szanto, A.J.; Pietrofesa, R.A.; Christofidou-Solomidou, M.; Cheung, M.; Mossman, B.T.; Shukla, A.; Testa, J.R. Inflammation-Related IL-1β/IL-1R Signaling Promotes the Development of Asbestos-Induced Malignant Mesothelioma. Cancer Prev. Res. 2016, 9, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Miao, E.A.; Rajan, J.V.; Aderem, A. Caspase-1-induced pyroptotic cell death. Immunol. Rev. 2011, 243, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Rishi, A.K.; Wu, W.; Polin, L.; Sharma, S.; Levi, E.; Albelda, S.; Pass, H.I.; Wali, A. Curcumin suppresses growth of mesothelioma cells in vitro and in vivo, in part, by stimulating apoptosis. Mol. Cell. Biochem. 2011, 357, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Cioce, M.; Canino, C.; Pulito, C.; Muti, P.; Strano, S.; Blandino, G. Butein impairs the protumorigenic activity of malignant pleural mesothelioma cells. Cell Cycle 2012, 11, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Dabir, S.; Kluge, A.; Dowlati, A. The association and nuclear translocation of the PIAS3-STAT3 complex is ligand and time dependent. Mol. Cancer Res. 2009, 7, 1854–1860. [Google Scholar] [CrossRef] [PubMed]
- Dabir, S.; Kluge, A.; Kresak, A.; Yang, M.; Fu, P.; Groner, B.; Wildey, G.; Dowlati, A. Low PIAS3 expression in malignant mesothelioma is associated with increased STAT3 activation and poor patient survival. Clin. Cancer Res. 2014, 20, 5124–5132. [Google Scholar] [CrossRef] [PubMed]
- Martinotti, S.; Ranzato, E.; Burlando, B. In vitro screening of synergistic ascorbate-drug combinations for the treatment of malignant mesothelioma. Toxicol. Vitro 2011, 25, 1568–1574. [Google Scholar] [CrossRef] [PubMed]
- Martinotti, S.; Ranzato, E.; Parodi, M.; Vitale, M.; Burlando, B. Combination of ascorbate/epigallocatechin-3-gallate/gemcitabine synergistically induces cell cycle deregulation and apoptosis in mesothelioma cells. Toxicol. Appl. Pharmacol. 2014, 274, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Beishline, K.; Azizkhan-Clifford, J. Sp1 and the ‘hallmarks of cancer’. FEBS J. 2015, 282, 224–258. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.A.; Lee, Y.J.; Ban, J.O.; Lee, Y.J.; Lee, S.H.; Cho, M.K.; Nam, H.S.; Hong, J.T.; Shim, J.H. The flavonoid resveratrol suppresses growth of human malignant pleural mesothelioma cells through direct inhibition of specificity protein 1. Int. J. Mol. Med. 2012, 30, 21–27. [Google Scholar] [PubMed]
- Lee, Y.J.; Lee, Y.J.; Im, J.H.; Won, S.Y.; Kim, Y.B.; Cho, M.K.; Nam, H.S.; Choi, Y.J.; Lee, S.H. Synergistic anti-cancer effects of resveratrol and chemotherapeutic agent clofarabine against human malignant mesothelioma MSTO-211H cells. Food Chem. Toxicol. 2013, 52, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Chae, J.I.; Cho, J.H.; Lee, K.A.; Choi, N.J.; Seo, K.S.; Kim, S.B.; Lee, S.H.; Shim, J.H. Role of transcriptor factor Sp1 in the quercetin-mediated inhibitory effect on human malignant pleural mesothelioma. Int. J. Mol. Med. 2012, 30, 835–841. [Google Scholar] [PubMed]
- Chae, J.I.; Jeon, Y.J.; Shim, J.H. Downregulation of Sp1 is involved in honokiol-induced cell cycle arrest and apoptosis in human malignant pleural mesothelioma cells. Oncol. Rep. 2013, 29, 2318–2324. [Google Scholar] [PubMed]
- Kim, K.H.; Yoon, G.; Cho, J.J.; Cho, J.H.; Cho, Y.S.; Chae, J.I.; Shim, J.H. Licochalcone A induces apoptosis in malignant pleural mesothelioma through downregulation of Sp1 and subsequent activation of mitochondria-related apoptotic pathway. Int. J. Oncol. 2015, 46, 1385–1392. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.A.; Lee, S.H.; Lee, Y.J.; Baeg, S.M.; Shim, J.H. Hesperidin induces apoptosis by inhibiting Sp1 and its regulatory protein in MSTO-211H cells. Biomol. Ther. 2012, 20, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.A.; Chae, J.I.; Shim, J.H. Natural diterpenes from coffee, cafestol and kahweol induce apoptosis through regulation of specificity protein 1 expression in human malignant pleural mesothelioma. J. Biomed. Sci. 2012, 19, 60. [Google Scholar] [CrossRef] [PubMed]
- Roomi, M.W.; Ivanov, V.; Kalinovsky, T.; Niedzwiecki, A.; Rath, M. Inhibition of malignant mesothelioma cell matrix metalloproteinase production and invasion by a novel nutrient mixture. Exp. Lung Res. 2006, 32, 69–79. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Polyphenols | Cell Type | Effects on ROS | Ref. |
|---|---|---|---|
| Curcumin | H-MESO cells | ↑ ROS | [108] |
| ↑ Caspase-1 | |||
| ↑ Pyroptotic cell death | |||
| EGCG | ACC-meso 1, Y-meso 8A, EHMES-10, EHMES-1, MSTO-211H, REN, MM98, BR95, E198 cells | ↑ ROS | [109,110,111] |
| ↑ H2O2 outside of cells | |||
| ↑ Apoptosis and necrosis | |||
| ↓ Cell proliferation | |||
| Resveratrol (+Clofarabine) | MSTO-211H cells | ↓ Nrf2 pathway | [112,113] |
| ↑ p53 phosphorylation | |||
| ↑ Pro-apoptotic proteins | |||
| LGM2605 (a synthetic lignan) | Murine peritoneal macrophages | ↓ ROS | [115] |
| ↓ Cytotoxicity | |||
| ↑ Phase II anti-oxidant enzymes | |||
| Quercetin + Rutin | Peritoneal macrophages of Wistar rats | ↓ ROS | [116] |
| ↓ Peritoneal macrophages injury by asbestos | |||
| EGCG + ECG | Peritoneal macrophages of Wistar rats | ↓ ROS | [117] |
| ↓ Peritoneal macrophages injury by asbestos |
| Polyphenols | Cell Type or Animal Model | Effects on Inflammation | Ref. |
|---|---|---|---|
| Curcumin | H-MESO, NCI-2052, NCI-H2452, MSTO-211H, and NCI-H28 cells | ↑ Caspase-1 | [108,148] |
| ↑ pro-IL-1β and block of maturation of IL-1 β | |||
| ↑ NLRP3 | |||
| ↓ NF-κB, TRL, and IL-1 pathways | |||
| ↑ PIAS-3 | |||
| ↓ p-STAT-3 | |||
| Butein | MSTO-211H, NCI-H28, NCI-H2052 | ↓ NF-κB, p-STAT-3 | [146] |
| ↓VEGF | |||
| ↓ IL-6, IL-8 | |||
| Flaxseed Lignans | MM-prone Nf2+/mut mice | ↓ IL-1β, IL-6, HMGB1, TNF-α, TGF-β1 | [42] |
| ↑ Nrf2-regulated anti-oxidant enzymes | |||
| EGCG + Ascorbate + Gemcitabine (AND) | REN cells | ↑ DAPK2 | [150] |
| ↑ TNSFR11B | |||
| ↓ TNFAIP3 | |||
| ↓ NF-κB pathway | |||
| Resveratrol | MSTO-211H cells | ↓ Sp1, p21, p27, cyclin D1, Mcl-1 | [152] |
| ↓ survivin | |||
| ↑ Apoptosis | |||
| Resveratrol + Clofarabine | MSTO-211H cells | ↓ Sp1, p-Akt | [153] |
| ↓ c-Met, cyclin D1, p21 | |||
| Quercetin | MSTO-211H cells | ↓ Sp1, cyclin D1, Mcl-1, survivin | [154] |
| ↑ Apoptosis | |||
| Honokiol | MSTO-211H cells | ↓ Sp1 | [155] |
| ↓ cyclin D1, Mcl-1, survivin | |||
| ↑ Apoptosis | |||
| Licochalcone A | MSTO-211H and H28 cells | ↓ Sp1 | [156] |
| ↓ cyclin D1, Mcl-1, survivin | |||
| ↑ Apoptosis | |||
| Hesperidin | MSTO-211H cells | ↓ Sp1 | [157] |
| ↓ p27, p21, cyclin D1, Mcl-1, survivin | |||
| ↑ Apoptosis | |||
| Cafestol and kahweol | MSTO-211H cells | ↓ Sp1, cyclin D1, Mcl-1, survivin | [158] |
| ↑ Apoptosis |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benvenuto, M.; Mattera, R.; Taffera, G.; Giganti, M.G.; Lido, P.; Masuelli, L.; Modesti, A.; Bei, R. The Potential Protective Effects of Polyphenols in Asbestos-Mediated Inflammation and Carcinogenesis of Mesothelium. Nutrients 2016, 8, 275. https://doi.org/10.3390/nu8050275
Benvenuto M, Mattera R, Taffera G, Giganti MG, Lido P, Masuelli L, Modesti A, Bei R. The Potential Protective Effects of Polyphenols in Asbestos-Mediated Inflammation and Carcinogenesis of Mesothelium. Nutrients. 2016; 8(5):275. https://doi.org/10.3390/nu8050275
Chicago/Turabian StyleBenvenuto, Monica, Rosanna Mattera, Gloria Taffera, Maria Gabriella Giganti, Paolo Lido, Laura Masuelli, Andrea Modesti, and Roberto Bei. 2016. "The Potential Protective Effects of Polyphenols in Asbestos-Mediated Inflammation and Carcinogenesis of Mesothelium" Nutrients 8, no. 5: 275. https://doi.org/10.3390/nu8050275
APA StyleBenvenuto, M., Mattera, R., Taffera, G., Giganti, M. G., Lido, P., Masuelli, L., Modesti, A., & Bei, R. (2016). The Potential Protective Effects of Polyphenols in Asbestos-Mediated Inflammation and Carcinogenesis of Mesothelium. Nutrients, 8(5), 275. https://doi.org/10.3390/nu8050275