Gender Differences in Response to Prolonged Every-Other-Day Feeding on the Proliferation and Apoptosis of Hepatocytes in Mice


Abstract
:1. Introduction
2. Material and Methods
2.1. Animals
2.2. Tissue Preparation
2.3. Histological Analysis
2.4. Immunohistochemical Analysis of Proliferative and Apoptotic Markers
2.5. Hepatocytes Nuclear Size and Nuclear Area
2.6. Real-Time Quantitative Reverse Transcription PCR (RQ-PCR)
2.7. Statistical Analysis
2.8. Statistical Analysis
3. Results
3.1. Body and Liver Mass
3.2. Histology of the Liver
3.3. The Nuclear Area of Hepatocytes
3.4. Immunolocalization of Ki67 and Bax
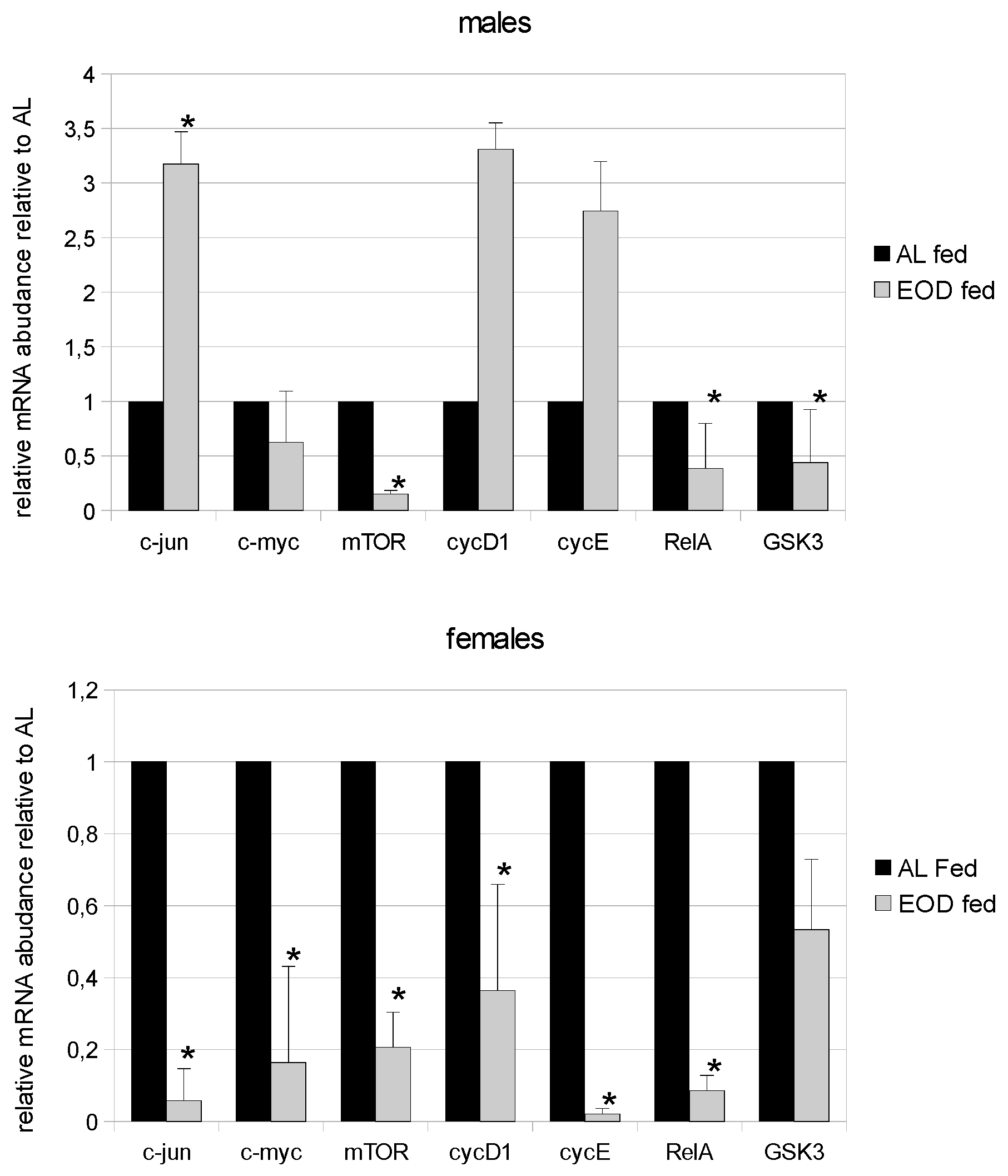
3.5. Expression of c-jun
3.6. Expression of the c-myc Proto-Oncogene
3.7. Expression of mTOR
3.8. Expression of Cyclin-D (cycD) and Cyclin-E (cycE)
3.9. Expression of RelA (p65)
3.10. Expression of GSK3
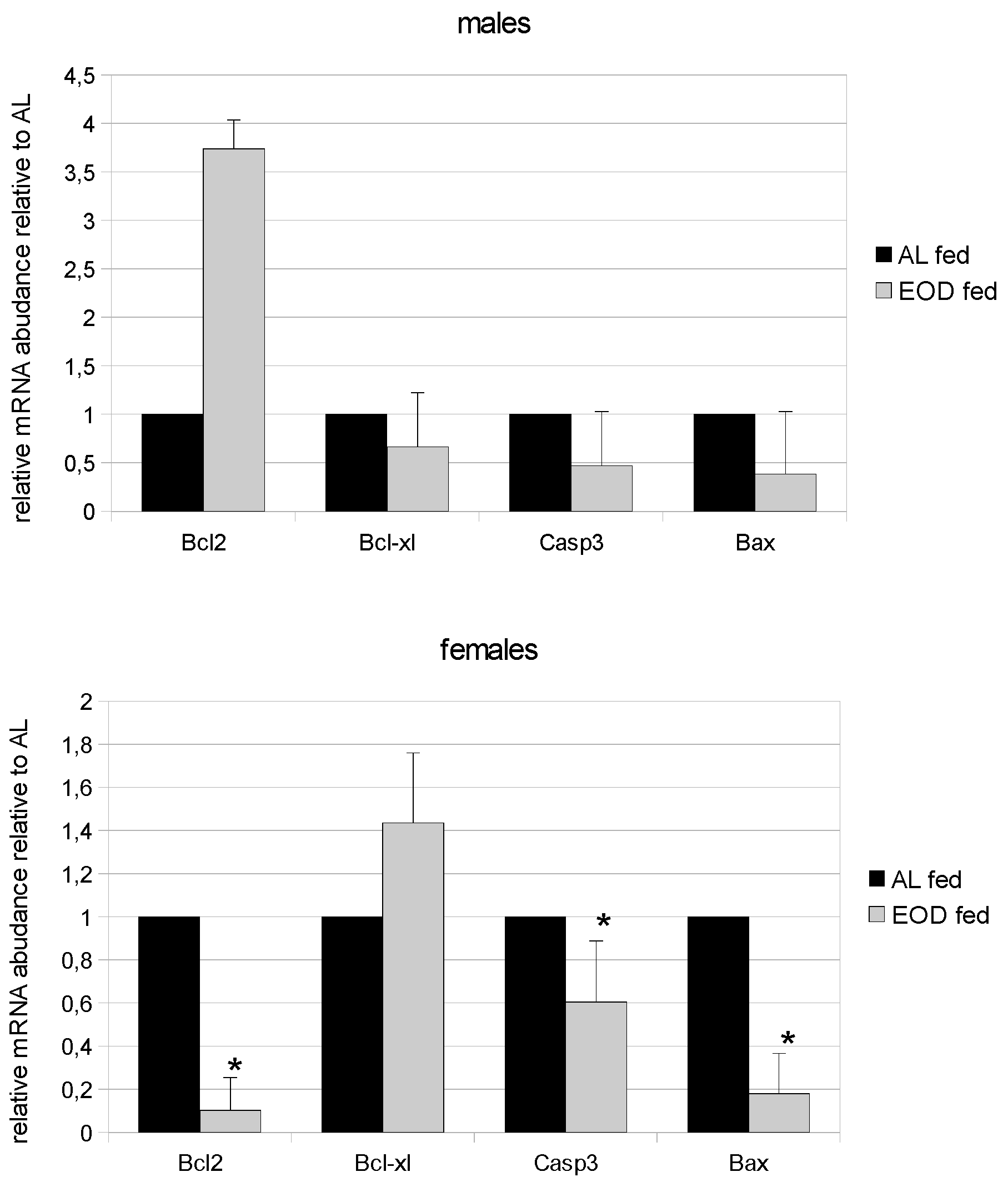
3.11. Expression of Bcl-2 and Bcl-xL
3.12. Expression of Caspase 3 and Bax
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Piper, M.D.W.; Selman, C.; McElwee, J.J.; Partridge, L. Separating cause from effect: How does insulin/IGF signaling control lifespan in worms, flies and mice? J. Intern. Med. 2008, 263, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Marques, F.Z.; Markus, M.A.; Morris, B.J. The molecular basis of longevity, and clinical implications. Maturitas 2010, 65, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Avogaro, A.; de Kreutzenberg, S.V.; Fadini, G.P. Insulin signaling and life span. Eur. J. Physiol. 2010, 459, 301–314. [Google Scholar] [CrossRef] [PubMed]
- Selman, C.; Lingard, S.; Choudhury, A.I.; Batterham, R.L.; Claret, M.; Clements, M.; Ramadani, F.; Okkenhang, K.; Schuster, E.; Blanc, E.; et al. Evidence for lifespan extension and delayed age-related biomarkers in insulin receptor substrate 1 null mice. FASEB J. 2008, 22, 807–818. [Google Scholar] [CrossRef] [PubMed]
- Bartke, A.; Brown-Borg, H.; Mattison, J.; Kinney, B.; Hauck, S.; Wright, C. Prolonged longevity of hopopituitary dwarf mice. Exp. Gerontol. 2001, 36, 21–28. [Google Scholar] [CrossRef]
- Anisimov, V.N. Metformin for aging and cancer prevention. Aging 2010, 2, 760–774. [Google Scholar] [CrossRef] [PubMed]
- Anisimov, V.N.; Zabezhinski, M.A.; Popovich, I.G.; Piskunova, T.S.; Semenchenko, A.V.; Tyndyk, M.L.; Yurova, M.N.; Antoch, M.P.; Blagosklonny, M.V. Rapamycin extends maximal lifespan in cancer-prone mice. Am. J. Pathol. 2010, 176, 2092–2097. [Google Scholar] [CrossRef] [PubMed]
- Stankovic, M.; Mladenovic, D.; Ninkovic, M.; Vucevic, D.; Tomasevic, T.; Radosavljevic, T. Effects of caloric restriction on oxidative stress parameters. Gen. Physiol. Biophys. 2013, 32, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A.E.; Lashinger, L.M.; Otto, G.; Nunez, N.P.; Hursting, S.D. Decreased systemic IGF-1 in response to calorie restriction modulates murine tumor cell growth, nuclear factor-κB activation, and inflammation-related gene expression. Mol. Carcinog. 2013, 52, 997–1006. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; de Cobo, R.; Anson, R.M.; Ingram, D.K.; Lane, M.A. Caloric restriction modulates insulin receptor signaling in liver and skeletal muscle of rat. Nutrition 2005, 21, 378–388. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.N.; Mitchell, S.E.; Hambly, C.; Morgan, D.G.; Clapham, J.C.; Speakman, J.R. Physiological and behavioral responses to intermitted starvation in C57BL/6J mice. Physiol. Behav. 2012, 105, 376–387. [Google Scholar] [CrossRef] [PubMed]
- Descamps, O.; Riondel, J.; Ducros, V.; Roussel, A.M. Mitochondrial production of reactive oxygen species and incidence of age-associated lymphoma in OF1 mice: Effect of alternate-day fasting. Mech. Ageing Dev. 2005, 126, 1185–1191. [Google Scholar] [CrossRef] [PubMed]
- Masternak, M.M.; Al-Regaiey, K.A.; Bonkowski, M.S.; Panici, J.A.; Bartke, A. Effect of every othr day feeding diet on gene expression in normal and long-lived Ames dwarf mice. Exp. Gerontol. 2005, 40, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Soeters, M.R.; Lammers, N.M.; Dubbelhuis, P.F.; Ackermans, M.; Jonkers-Schuitema, C.F.; Fliers, E.; Sauerwein, H.P.; Aerts, J.M.; Serlie, M.J. Intermittent fasting does not affect whole-body glucose. Lipid, or protein metabolism. Am. J. Clin. Nutr. 2009, 90, 1244–1251. [Google Scholar] [CrossRef] [PubMed]
- Harvie, M.N.; Pegington, M.; Mattson, M.P.; Frystyk, J.; Dillon, B.; Evans, G.; Cuzick, J.; Jebb, S.A.; Martin, B.; Cutler, R.G. The effects of intermittent or continuous energy restriction on weoght loss and metabolic disease risk markers: A randomized trial in young overweight women. Int. J. Obes. 2011, 35, 714–727. [Google Scholar] [CrossRef] [PubMed]
- Kouda, K.; Nakamura, H.; Kohno, H.; Ha-Kawa, S.K.; Tokunaga, R.; Sawada, S. Dietary restriction: Effects of short-term fasting on protein uptake and cell death/proliferation in the rat liver. Mech. Ageing Dev. 2004, 125, 375–380. [Google Scholar] [CrossRef] [PubMed]
- Sokolović, A.; van Roomen, C.P.A.A.; Ottenhoff, R.; Scheij, S.; Hiralall, J.K.; Claessen, N.; Aten, J.; Oude Elferink, R.P.; Groen, A.K.; Sokolović, M. Fasting reduces liver fibrosis in a mouse model for chronic cholangiopathies. Biochim. Biphys. Acta 2013, 1832, 1482–1491. [Google Scholar] [CrossRef] [PubMed]
- Chausse, B.; Vieira-Lara, M.A.; Sanchez, A.; Medeiros, M.H.G.; Kowaltowski, A.J. Intermittent fasting results in tissue-specific changes in bioenergetics and redox state. PLoS ONE 2015. [Google Scholar] [CrossRef] [PubMed]
- Martin, B.; Pearson, M.; Kebejian, L.; Golden, E.; Keselman, A.; Bender, M.; Carlson, O.; Egan, J.; Ladenheim, B.; Cadet, J.L. Sex dependent metabolic, neuroendocrine, and cognitive responses to dietary energy restriction and excess. Endocrinology 2007, 148, 4318–4333. [Google Scholar] [CrossRef] [PubMed]
- Guevara, R.; Valle, A.; Gianotti, M.; Roca, P.; Oliver, J. Gender-dependent differences in serum profiles of insulin and leptin in caloric restricted rats. Horm. Metab. Res. 2008, 40, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Anisimov, V.N.; Popovich, I.G.; Zabezhinski, M.A.; Egormin, P.A.; Yurova, M.N.; Semenchenko, A.V.; Tyndyk, M.L.; Panchenko, A.V.; Trashkov, A.P.; Vasiliev, A.G.; et al. Sex differences in aging, life span and spontaneous tumorigenesis in 129/Sv mice neonatally expose to metformin. Cell Cycle 2015, 14, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Grymula, K.; Piotrowska, K.; Słuczanowska-Głąbowska, S.; Mierzejewska, K.; Tarnowski, M.; Tkacz, M.; Poniewierska-Baran, A.; Pedziwiatr, D.; Suszynska, E.; Laszczynska, M.; et al. Positive effects of prolonged caloric restriction on the population of very small embryonic-like stem cells—Hematopoietic and ovarian implications. J. Ovarian Res. 2014, 7, 68. [Google Scholar] [CrossRef] [PubMed]
- Park, I.-S.; Kim, B.S.; Kim, H.S. Effect of starvation on growth and hepatocyte nuclear size of larval haddock, Melanogrammus aeglefinus. Dev. Reprod. 2012, 16, 107–112. [Google Scholar]
- Gregg, S.Q.; Gutierrez, V.; Robinson, A.R.; Woodell, T.; Nakao, A.; Ross, M.A.; Michalopoulos, G.K.; Rigatti, L.; Rothermel, C.E.; Kemileri, J.; et al. A mouse model of accelerated aging caused by a defect in DNA repair. Hepatology 2012, 55, 609–621. [Google Scholar] [CrossRef] [PubMed]
- Youen, M.F.; Wu, P.C.; Lai, V.C.; Lau, J.Y.; Lai, C.L. Expression of c-Myc, c-Fos and c-jun in hepatocellular carcinoma. Cancer 2001, 91, 106–112. [Google Scholar] [CrossRef]
- Lamming, D.W.; Sabatini, D.M. A central role for mTOR in lipid homeostasis. Cell Metab. 2013, 18, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Bhardwaj, A.; Aggarwal, R.S.; Seeram, N.P.; Shishodia, S.; Takada, Y. Role of resveratrol in prevention and therapy of cancer: Preclinical and clinical studies. Anticancer Res. 2004, 24, 2783–2840. [Google Scholar] [PubMed]
- Liu, H.X.; Ly, I.; Hu, Y.; Wan, Y.J. Retinoic acid regulates cell cycle genes and accelerates normal mouse liver regeneration. Biochem. Pharmacol. 2014, 91, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Shevra, C.R.; Ghosh, A.; Kumar, M. Cyclin D1 and Ki-67 expression in normal, hyperplastic and neoplastic endometrium. J. Postgrad. Med. 2015, 61, 15–20. [Google Scholar] [PubMed]
- Zhang, N.-N.; Sun, Q.-S.; Chen, Z.; Liu, F.; Jiang, Y.-Y. Homeostatic regulatory role of Pokemon in NF-κB signaling: Stimulating both p65 and IκBα expression in human hepatocellular carcinoma cells. Mol. Cell. Biochem. 2013, 372, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Miquet, J.G.; Freud, T.; Martinez, C.S.; González, L.; Diaz, M.E.; Micucci, G.P.; Zotta, E.; Boparai, R.K.; Bartke, A.; Turyn, D.; et al. Hepatocellular alternations and dysregulation of oncogenic pathways in the liver of transgenic mice overexpressing groth hormone. Cell Cycle 2013, 12, 1042–1057. [Google Scholar] [CrossRef] [PubMed]
- Jope, R.S.; Yuskaitis, C.J.; Beurel, E. Glycogen synthase kinase -3 (GSK3): Inflammation, diseases and therapeutics. Neurochem. Res. 2007, 32, 577–595. [Google Scholar] [CrossRef] [PubMed]
- Van Delft, M.F.; Huang, D.C. How the Bcl-2 family of proteins interact to regulate apoptosis. Cell Res. 2006, 16, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Kirchner, H.; Hofmann, S.M.; Fischer-Rosinský, A.; Hembree, J.; Abplanalp, W.; Ottaway, N.; Donelan, E.; Krishna, R.; Woods, S.C.; Müller, T.D.; et al. Caloric restriction chronically impairs metabolic programming in mice. Diabetes 2012, 61, 2734–2742. [Google Scholar] [CrossRef] [PubMed]
- Farooq, N.; Priyamvada, S.; Arivarasu, N.A.; Salim, S.; Khan, F.; Yusufi, A.N. Influence of Ramadan-type fasting on enzymes of carbohydrate metabolizm and brush border membrane In small interstine and liver of rat used as a model. Br. J. Nutr. 2006, 96, 1087–1094. [Google Scholar] [CrossRef] [PubMed]
- Duan, W.; Guo, Z.; Jiang, H.; Ware, M.; Mattson, M.P. Reversal of behavioral and metabolic abnormalities, and insulin resistance syndrome, by dietary restriction in mice deficient in brain-derived neutrophic factor. Endocrinology 2003, 144, 2446–2453. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Jiang, W.; McGinley, J.; Wolfe, P.; Thompson, H.J. Effects of dietary energy repletion and IGF-1 infusion on the inhibition of mammary carcinogenesis by dietary energy restriction. Mol. Carcinog. 2005, 42, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Wronska, A.; Sledzinski, T.; Goyke, E.; Lawniczak, A.; Wierzbicki, P.; Kmiec, Z. Short term calorie restriction and refeeding differently affect lipogenic enzymes in major white adipose tissue depot of Young and old rats. J. Physiol. Pharmacol. 2014, 65, 117–126. [Google Scholar] [PubMed]
- Li, L.; Fu, Y.-C.; Xu, J.-J.; Chen, X.-C.; Lin, X.-H.; Luo, L.-L. Caloric restriction promotes the reproductive capacity of female rats via modulating the level of insulin-like growth factor -1 (IGF-1). Gen. Comp. Endocrinol. 2011, 174, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Sluczanowska-Glabowska, S.; Laszczynska, M.; Piotrowska, K.; Grabowska, M.; Grymula, K.; Ratajczak, M.Z. Caloric restriction increases ratio of estrogen to androgen receptors expression in murine ovaries- potential therapeutic implications. J. Ovarian Res. 2015, 8, 57. [Google Scholar] [CrossRef] [PubMed]
- Mauvais-Jarvis, F.; Clegg, D.J.; Hevener, A.L. The role of estrogens in control of energy balance and glucose homeostasis. Endocr. Rev. 2013, 34, 309–338. [Google Scholar] [CrossRef] [PubMed]
- Jelenik, T.; Roden, M. How estrogens prevent from lipid-induced insulin resistance. Endocrinology 2013, 154, 989–992. [Google Scholar] [CrossRef] [PubMed]
- Rogers, N.H.; Perfield, J.W., II; Strissel, K.J.; Obin, M.S.; Greenberg, A.S. Reduced energy expenditure and increased inflammation are early events in the development of ovariectomy-induced obesity. Endocrinology 2009, 150, 2161–2168. [Google Scholar] [CrossRef] [PubMed]
- Stubbins, R.E.; Najjar, K.; Holcomb, V.B.; Hong, J.; Núñez, N.P. Estrogen alters adipocyte biology and protects female mice from adipocyte inflammation and insulin resistance. Diabetes Obes. Metab. 2012, 14, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Ning, B.-F.; Ding, J.; Liu, J.; Yin, C.; Xu, W.P.; Cong, W.M.; Zhang, Q.; Chen, F.; Han, T.; Deng, X.; et al. Hepatocyte Nuclear Factor 4α-Nuclear Factor-κB Feedback Circuit Modulates Liver Cancer Progression. Hepatology 2014, 60, 1607–1619. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.; Nagini, S.; Rana, A. Expression and inactivation of glycogen synthase kinase 3 alpha/beta and their association with the expression of cyclin D1 and p53 in oral squamous cell carcinoma progression. Mol. Cancer 2015, 14, 20. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, K.; Van Bockstaele, D.R.; Berneman, Z.N. The cell cycle: A review of regulation, deregulation and therapeutic targets in cancer. Cell Prolif. 2003, 36, 131–149. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.A.; He, L.F.; Chen, X.M. Effects of cadmium on hepatocellular DNA damage, proto-oncogen expression and apoptosis in rats. Biomed. Environ. Sci. 2007, 20, 146–153. [Google Scholar] [PubMed]
- Lv, M.; Zhu, X.; Wang, H.; Wang, F.; Guan, W. Roles of caloric restriction, ketogenic diet and intermitted fasting during initiation, progression and metastasis of cancer in animal models: A systematic review and meta-analysis. PLoS ONE 2014, 9, e115147. [Google Scholar] [CrossRef] [PubMed]
- Rizza, W.; Veronese, N.; Fontana, L. What are the roles of calorie restriction and diet quality in promoting healthy longevity. Aging Res. Rev. 2014, 13, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Niedernhofer, L.J.; Garinis, G.A.; Raams, A.; Lalai, A.S.; Robinson, A.R.; Appeldoorn, E.; Odijk, H.; Oostendorp, R.; Ahmad, A.; van Leeuwen, W.; et al. A new progeroid syndrome reveals that genotoxic stress suppresses the somatotroph axis. Nature 2006, 444, 1038–1043. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Mayer, J.A.; Mazumdar, A.; Fertuck, K.; Kim, H.; Brown, M.; Brown, P.H. Estrogen induces c-myc gene expression via an upstream enhancer activated by the estrogen receptor and the AP-1 Transcription Factor. Mol. Endocrinol. 2011, 25, 1527–1538. [Google Scholar] [CrossRef] [PubMed]
- Kashima, H.; Shiozawa, T.; Miyamoto, T.; Suzuki, A.; Uchikawa, J.; Kurai, M.; Konishi, J. Autocrine stimulation of IGF-1 in estrogen-induced growth of endometrial carcinoma cells: Involvement of the mitogen-activated protein kinase pathway followed by up-regulation of cyclin D1 and cyclin E. Endocr. Relat. Cancer 2009, 16, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, E.; Dieudonné, M.N.; Leneveu, M.C.; Sérazin, V.; Rincheval, V.; Mignotte, B.; Chouillard, E.; De Mazancourt, P.; Giudicelli, Y.; Pecquery, R. Effects of 17beta-estradiol on preadipocyte proliferation in human adipose tissue: Involvement of IGF-1-R signaling. Horm. Metab. Res. 2010, 42, 514–520. [Google Scholar] [CrossRef] [PubMed]
- Uebi, T.; Umeda, M.; Imai, T. Estrogen induces estrogen receptor alpha expression and hepatocyte proliferation in the livers of male mice. Genes Cells 2015, 20, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Schadt, E.E.; Wang, S.; Wang, H.; Arnold, A.P.; Ingram-Drake, T.A.; Lusis, A.J. Tissue-specific expression and regulation of sexually dimorphic genes in mice. Genome Res. 2006, 16, 995–1004. [Google Scholar] [CrossRef] [PubMed]
- Kosters, A.; Sun, D.; Wu, H.; Tian, F.; Felix, J.C.; Li, W.; Karpen, S.J. Sexually dimorphic genome-wide binding of retinoid X receptor alpha (RXRα) determines male-female differences in the expression of hepatic lipid processing genes in mice. PLoS ONE 2013, 8, e71538. [Google Scholar]
- Conforto, T.L.; Waxman, D.J. Sex-specific mouse liver gene expression: Genome wide analysis of developmental changes from pre-pubertal period to young adulthood. Biol. Sex Differ. 2012, 3, 9. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.; Sheng, Y.; Jiang, P.; Ji, L.; Xia, Y.; Min, Y.; Wang, Z. The gender-dependent difference of liver GSH antioxidant system in mice and its influence on isoline-induced liver injury. Toxicology 2011, 280, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Eghbali-Webb, M. Gender-related differences in basal and hypoxia- induced activation signal transduction pathways controlling cell cycle progression and apoptosis, in cardiac fibroblasts. Endocrine 2002, 18, 137–145. [Google Scholar] [CrossRef]
- Chen, C.; Hu, L.X.; Dong, T.; Wang, G.Q.; Wang, L.H.; Zhou, X.P.; Jiang, Y.; Murao, K.; Lu, S.Q.; Chen, J.W.; et al. Apoptosis and autophagy contribute to gender difference in cardiac ischemia-reperfusion induced injury in rats. Life Sci. 2013, 28, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Tsukahara, S.; Hojo, R.; Kuroda, Y.; Fujimaki, H. Estrogen modulates Bcl-2 family protein expression in the sexually dimorphic nucleus of the preoptic area of postnatal rats. Neurosci. Lett. 2008, 432, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Ren, P.; Li, X.; Guan, Y.; Zhang, Y.A. 17β-estradiol protects dopaminergic neurons in organotypic slice of mesencephalon by MAPK-mediated activation of anti-apoptosis gene BCL2. J. Mol. Neurosci. 2011, 45, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, M.A.; Rakoczy, S.G.; Brown-Borg, H.M. Long-living Ames dwarf Mouse hepatocytes readily undergo apoptosis. Exp. Gerontol. 2003, 38, 997–1008. [Google Scholar] [CrossRef]
- Muskhelishvili, L.; Hart, R.W.; Turturro, A.; James, S.J. Age-related changes in the intristic rate of apoptosis in livers of diet-restricted and Ad Libitum-fed B6C3F1 mice. Am. J. Pathol. 1995, 147, 20–24. [Google Scholar] [PubMed]
- Lee, J.H.; Jung, K.J.; Kim, J.W.; Kim, H.J.; Yu, B.P.; Chung, H.Y. Suppression of apoptosis by calorie restriction in aged kidney. Exp. Gerontol. 2004, 39, 1361–1368. [Google Scholar] [CrossRef] [PubMed]
- López –Domínguez, J.A.; Khraiwesh, H.; González-Reyes, J.A.; López-Lluch, G.; Navos, P.; Ramsey, J.J.; de Cabo, R.; Burón, M.I.; Villalba, J.M. Dietary fat and aging modulate apoptotic signaling in liver of calorie-restricted mice. J. Gerontol. A Biol. Sci. Med. Sci. 2015, 70, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Gesing, A.; Masternak, M.M.; Wang, F.; Lewinski, A.; Karbownik-Lewinska, M.; Bartke, A. Decreased expression level of apoptosis-related genes and/or proteins in skeletal muscles, but not in hearts, of growth hormone receptor knockout mice. Exp. Biol. Med. (Maywood) 2011, 236, 156–168. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| c-myc-forward | 5′-TCACCAGCACAACTACGCCG |
|---|---|
| c-myc-reverse | 5′-TGCTTCAGGACCCTGCCACT |
| c-jun-forward | 5′-CATTGCCGCCTCCAAGTG |
| c-jun-reverse | 5′-CCAGCTCGGAGTTTTGCG |
| mTOR-forward | 5′-GTTTGTGGCTCTGAATGACC |
| mTOR-reverse | 5′-TCAGGATCTGGATGAGCATC |
| cyclD1-forward | 5′-GCGAAGTGGAGACCATCCG |
| cyclD1-reverse | 5′-GGTCTCCTCCGTCTTGAGC |
| RelA-forward | 5′-GCTCCTGTTCGAGTCTCCAT |
| RelA-reverse | 5′-TAGGTCCTTTTGCGCTTCTC |
| cyclE-forward | 5′-GGCGGACACAGCTTCGGGTC |
| cyclE-reverse | 5′-TGGGTCTTGCAAAAACACGGCCA |
| GSK3b-forvard | 5′-CCACCATCCTTATCCCTCCAC |
| GSK3b-reverse | 5′-GTATCTGAGGCTGCTGTGGC |
| Bcl-2-forward | 5′-GTC CCG CCT CTT CAC CTT TCA G |
| Bcl-2-reverse | 5′-GAT TCT GGT GTT TCC CCG TTG G |
| Bcl-xL -forward | 5′-AAC ATC CCA GCT TCA CAT AAC CCC |
| Bcl-xL-reverse | 5′-GCG ACC CCA GTT TAC TCC ATC C |
| Bax -forward | 5′-GCG TGG TTG CCC TCT TCT ACT TTG |
| Bax-reverse | 5′-AGT CCA GTG TCC AGC CCA TGA TG |
| Casp3-forward | 5′-ATGGAGAACAACAAAACCTCAGT |
| Casp3-reverse | 5′-TTGCTCCCATGTATGGTCTTTAC |
| Body Weight (g) | Beginning of the Experiment | End of the Experiment | Gain of Weight (%) |
|---|---|---|---|
| AL males | 28.66 ± 0.75 | 41.79 ± 4.96 * | 45.81% |
| EOD. males | 24.48 ± 1.47 | 29.67 ± 2.54 * | 21.2% |
| AL. Females | 21.24 ± 1.15 | 30.13 ± 3.35 | 41.85% |
| EOD Females | 22.14 ± 1.97 | 29.13 ± 2.46 | 31.57% |
| Body Weight (g) | Liver Weight (g) | Liver/Body Weight | |
|---|---|---|---|
| AL. Males | 41.79 ± 4.96 * | 1.74 ± 0.44 | 4.16% |
| EOD Males | 29.67 ± 2.54 * | 1.42 ± 0.14 | 4.78% |
| AL. Females | 30.13 ± 3.35 | 1.30 ± 0.37 | 4.31% |
| EOD Females | 29.13 ± 2.46 | 1.42 ± 0.17 | 4.87% |
| AL Males | EOD Males | AL Females | EOD Females | |
|---|---|---|---|---|
| Average | 72.66 * | 51.02 * | 54.02 * | 32.63 * |
| SD | ±25.51 | ±14.97 | 9.81 | 8.45 |
| median | 70.10 | 48.60 | 54.11 | 30.30 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piotrowska, K.; Tarnowski, M.; Zgutka, K.; Pawlik, A. Gender Differences in Response to Prolonged Every-Other-Day Feeding on the Proliferation and Apoptosis of Hepatocytes in Mice. Nutrients 2016, 8, 176. https://doi.org/10.3390/nu8030176
Piotrowska K, Tarnowski M, Zgutka K, Pawlik A. Gender Differences in Response to Prolonged Every-Other-Day Feeding on the Proliferation and Apoptosis of Hepatocytes in Mice. Nutrients. 2016; 8(3):176. https://doi.org/10.3390/nu8030176
Chicago/Turabian StylePiotrowska, Katarzyna, Maciej Tarnowski, Katarzyna Zgutka, and Andrzej Pawlik. 2016. "Gender Differences in Response to Prolonged Every-Other-Day Feeding on the Proliferation and Apoptosis of Hepatocytes in Mice" Nutrients 8, no. 3: 176. https://doi.org/10.3390/nu8030176
APA StylePiotrowska, K., Tarnowski, M., Zgutka, K., & Pawlik, A. (2016). Gender Differences in Response to Prolonged Every-Other-Day Feeding on the Proliferation and Apoptosis of Hepatocytes in Mice. Nutrients, 8(3), 176. https://doi.org/10.3390/nu8030176