Polymorphism of the Transcription Factor 7-Like 2 Gene (TCF7L2) Interacts with Obesity on Type-2 Diabetes in the PREDIMED Study Emphasizing the Heterogeneity of Genetic Variants in Type-2 Diabetes Risk Prediction: Time for Obesity-Specific Genetic Risk Scores

 ,
,  ,
,  ,
,  , ,
, ,  , , ,
, , ,  , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Demographic, Clinical, Anthropometric, and Dietary Measurements
2.2. Biochemical Determinations, DNA Extraction and Genotyping
2.3. Outcomes and Follow-Up
2.4. Statistical Analyses
2.4.1. Baseline Association and Interaction Analyses in Determining T2D Prevalence and Obesity-Related Variables
2.4.2. Interaction Analysis between the TCF7L2-rs7903146 Polymorphism and Obesity in Determining T2D Incidence
2.4.3. Predictive Value Calculations for the TCF72-rs7903146 Polymorphism on T2D Incidence and Prevalence in the Whole PREDIMED Participants
2.4.4. Construction of Obesity-Specific GRS with the TCF7L2 and Other T2D-SNPs; Association and Evaluation of the PV for T2D Prevalence
2.4.5. Longitudinal Association and Interaction Analysis between the TCF7L2-rs7903146 Polymorphism and T2D in Determining BMI
2.4.6. Power Calculations
3. Results
3.1. Interaction between the TCF7L2-rs7903146 Polymorphism and Obesity in Determining T2D at Baseline
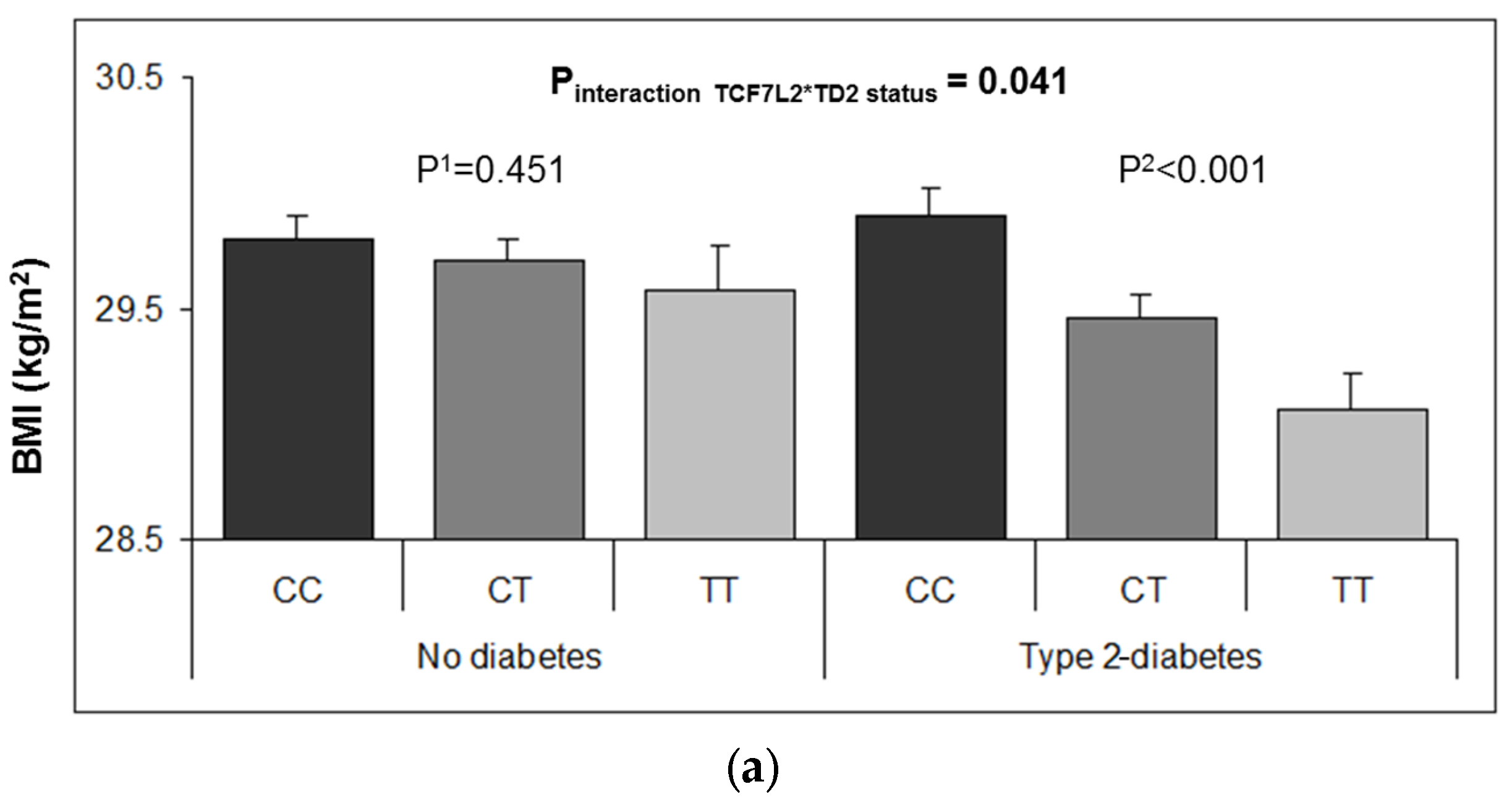
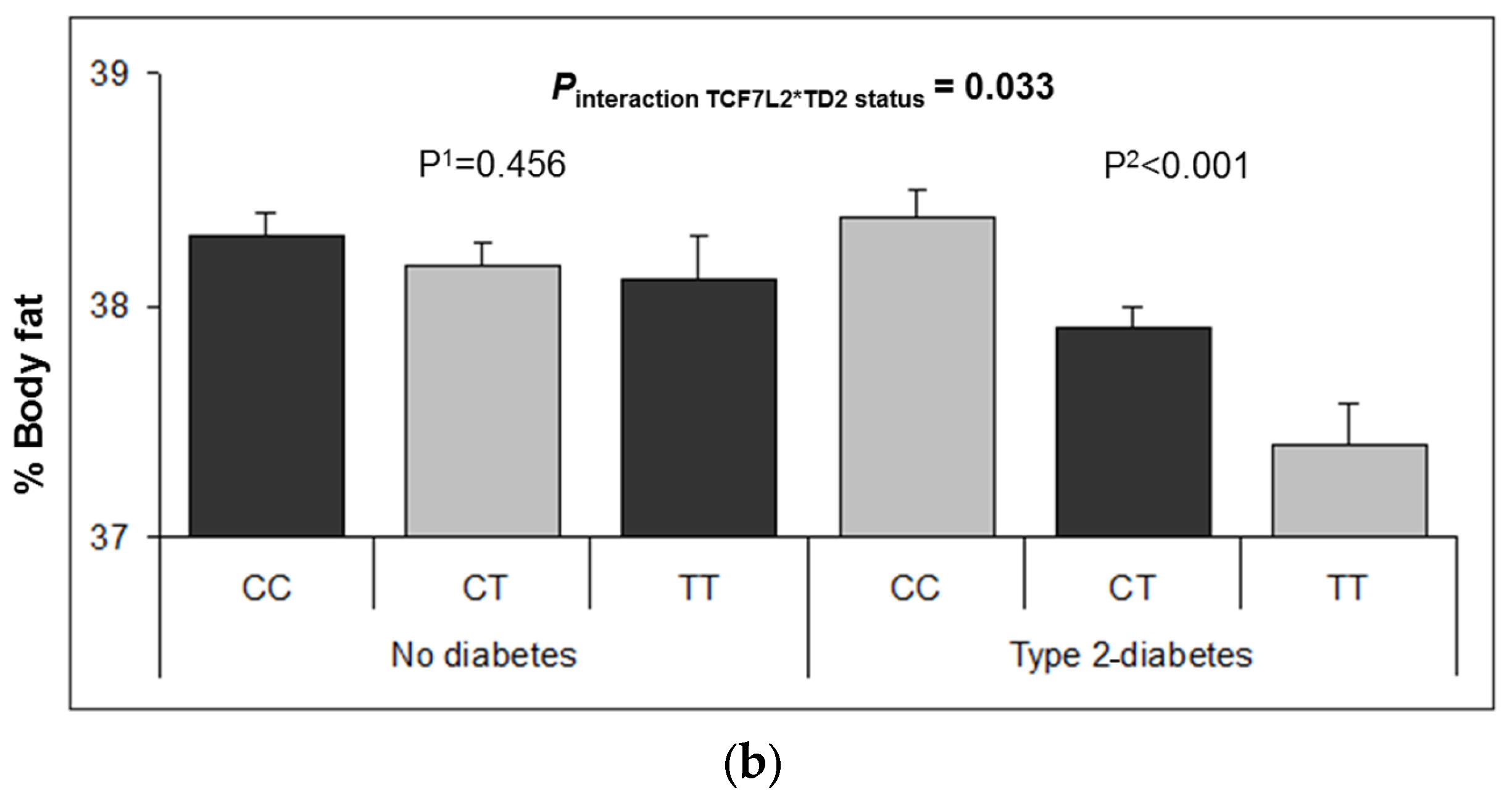
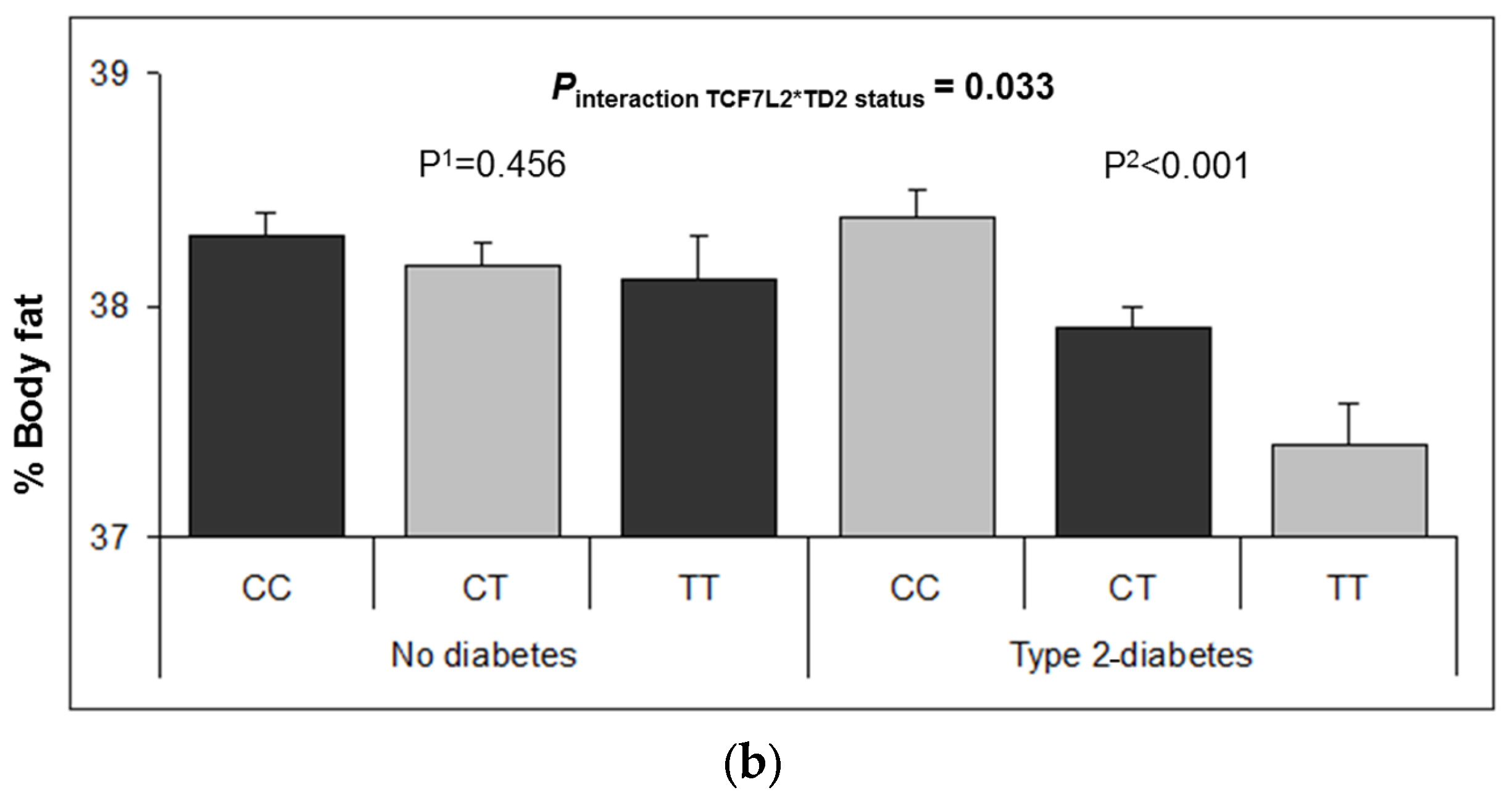
3.2. Interaction between the TCF7L2-rs7903146 Polymorphism and T2D in Determining Obesity-Related Measures at Baseline
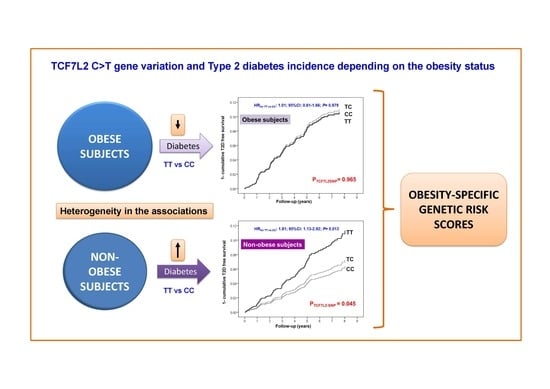
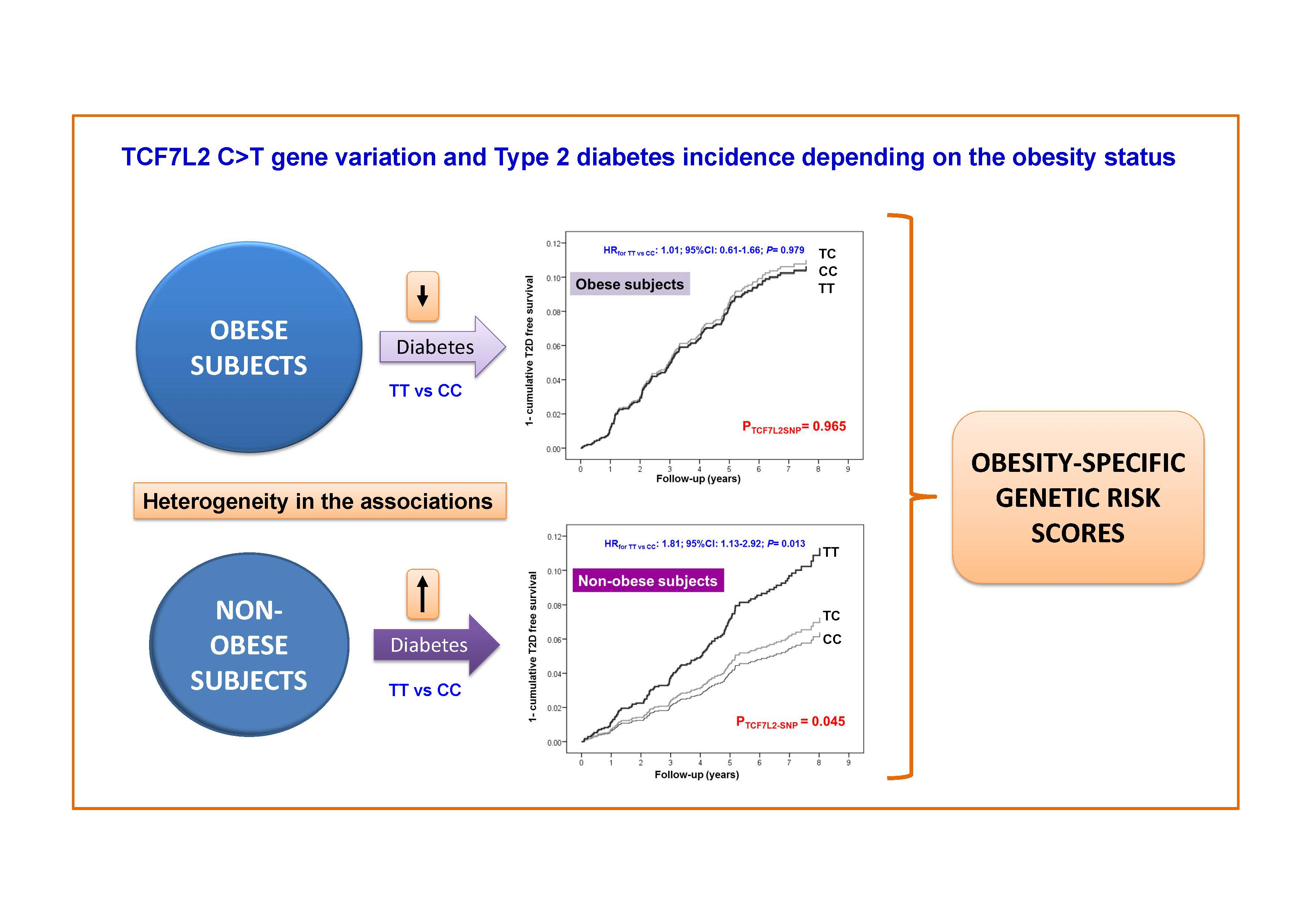
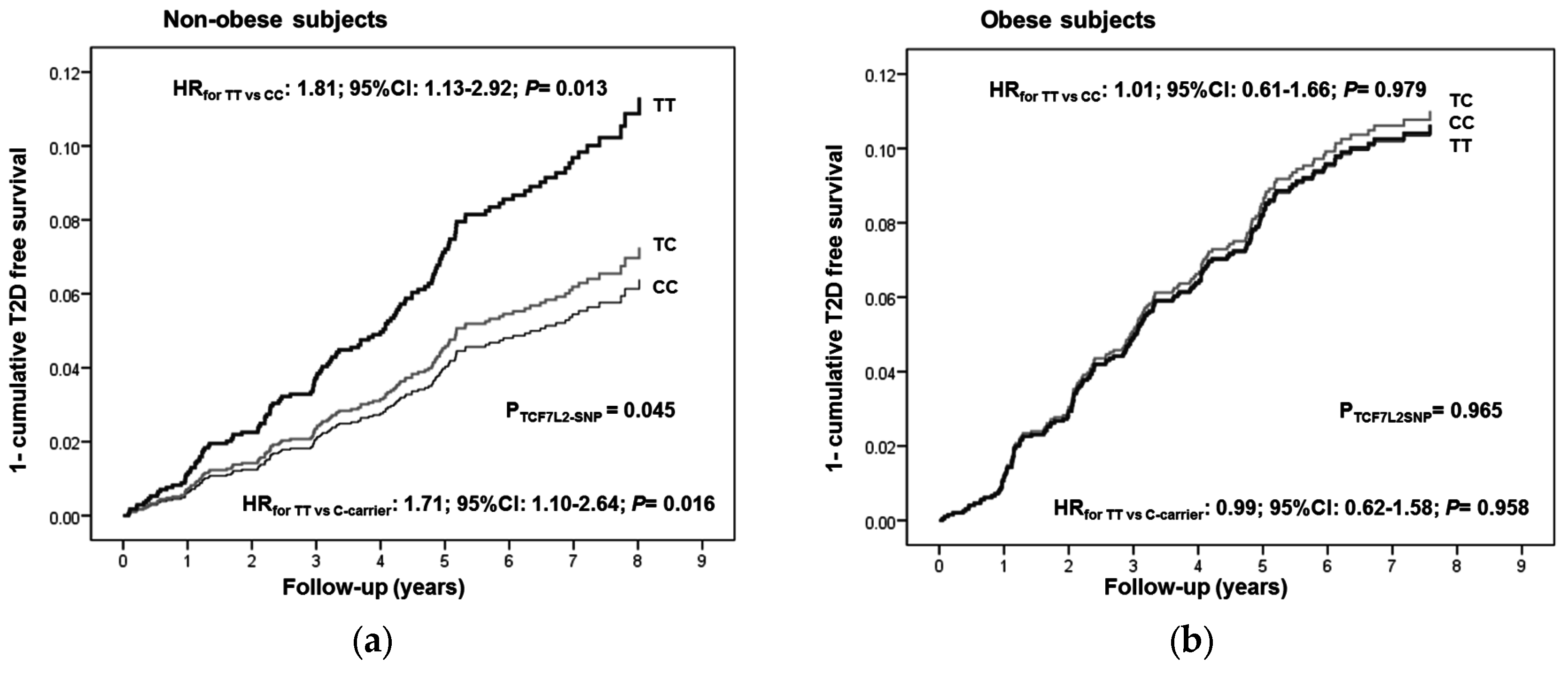
3.3. Interaction between the TCF7L2-rs7903146 Polymorphism and Obesity in the Incidence of T2D
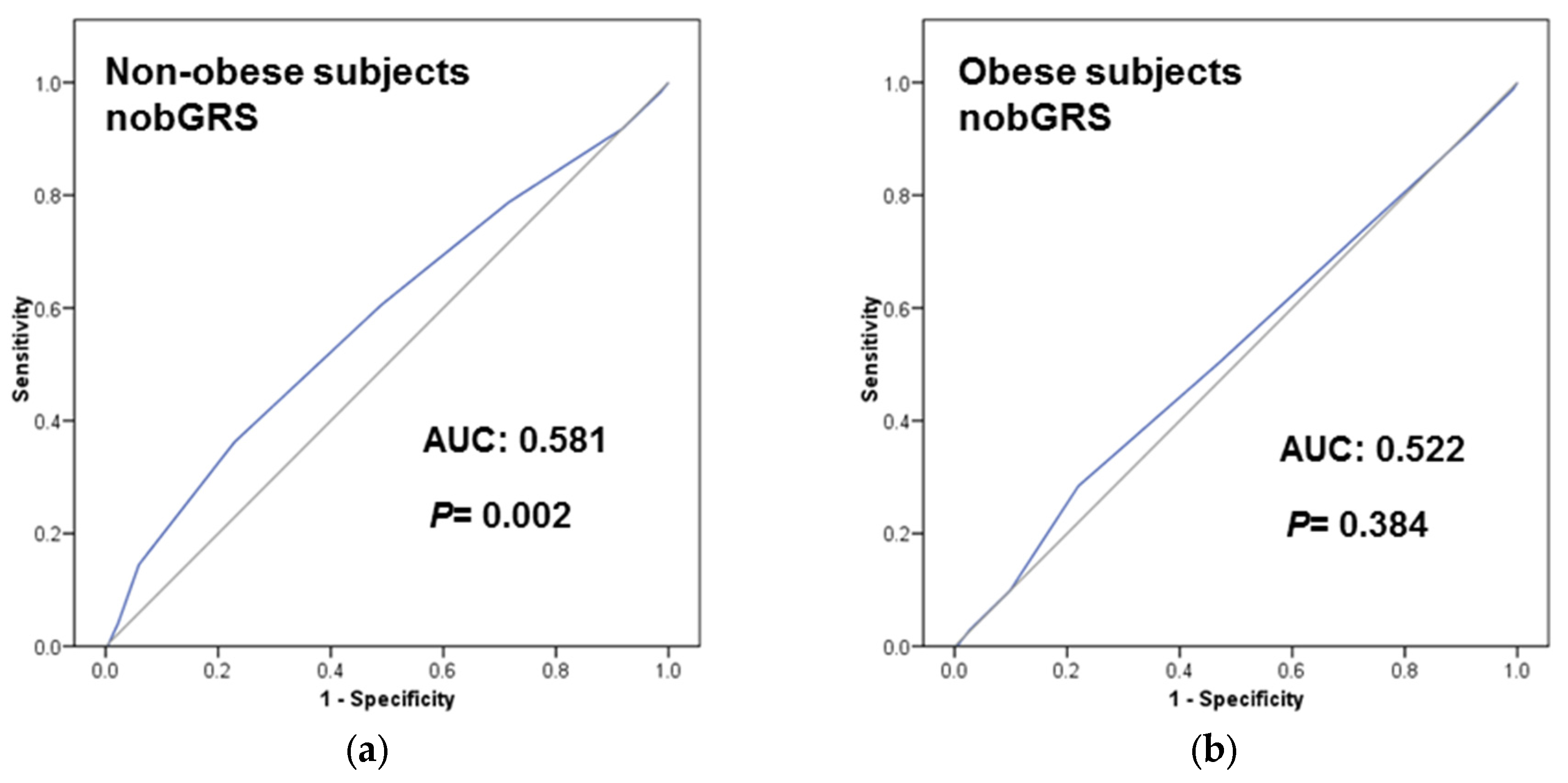
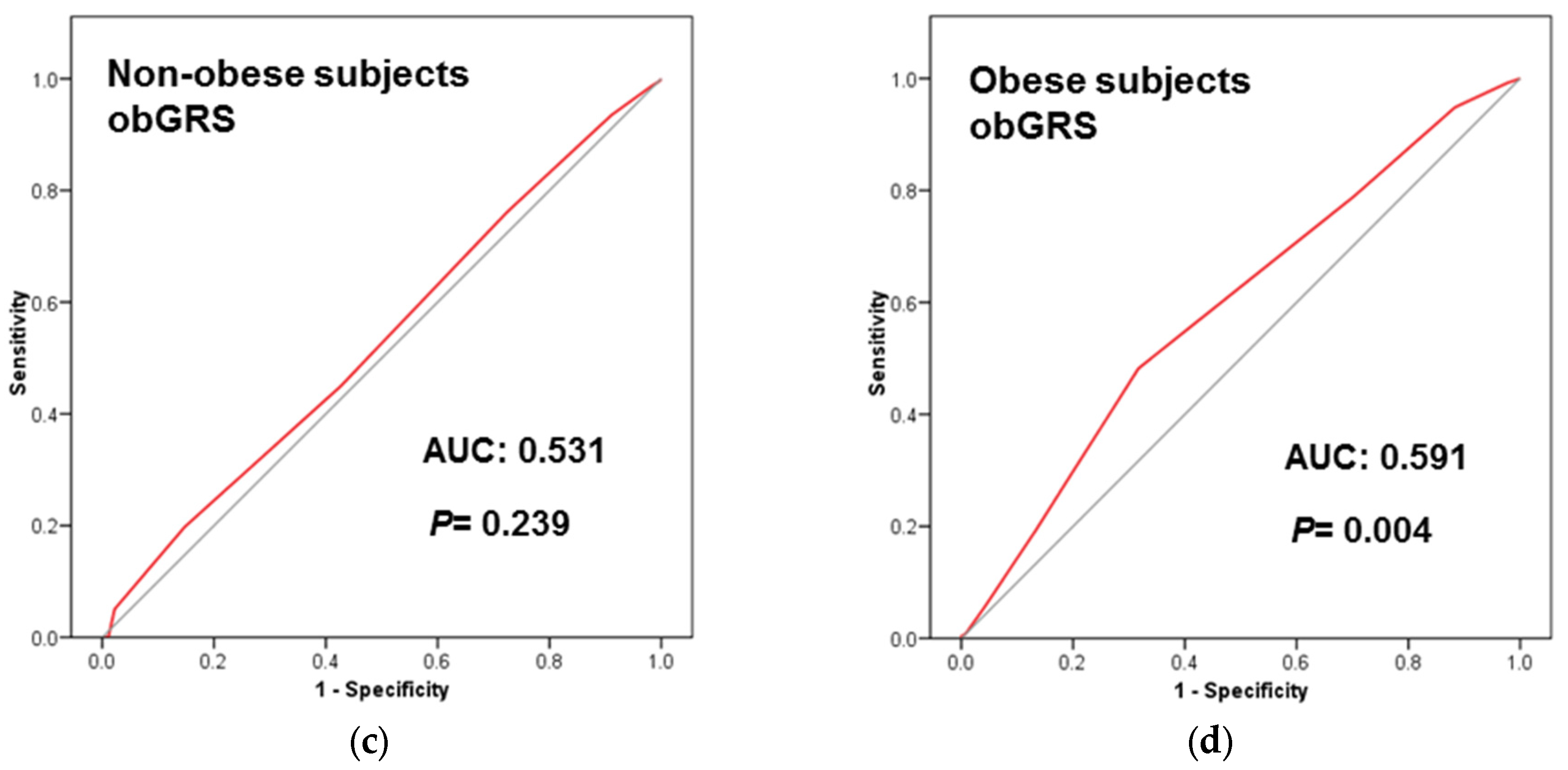
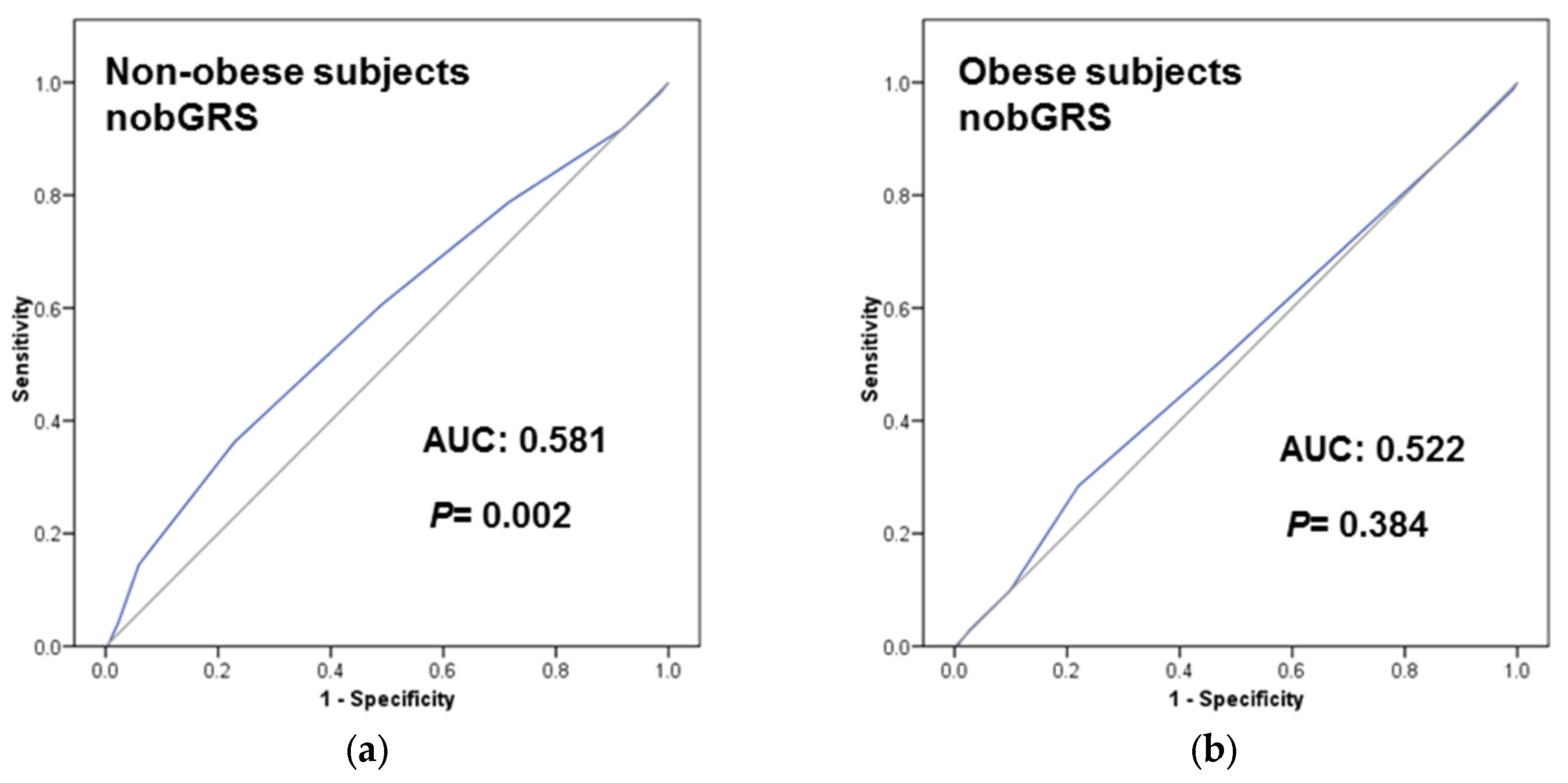
3.4. Predictive Ability of the TCF7L2-rs7903146 on T2D Incidence and Prevalence Depending on Obesity Status
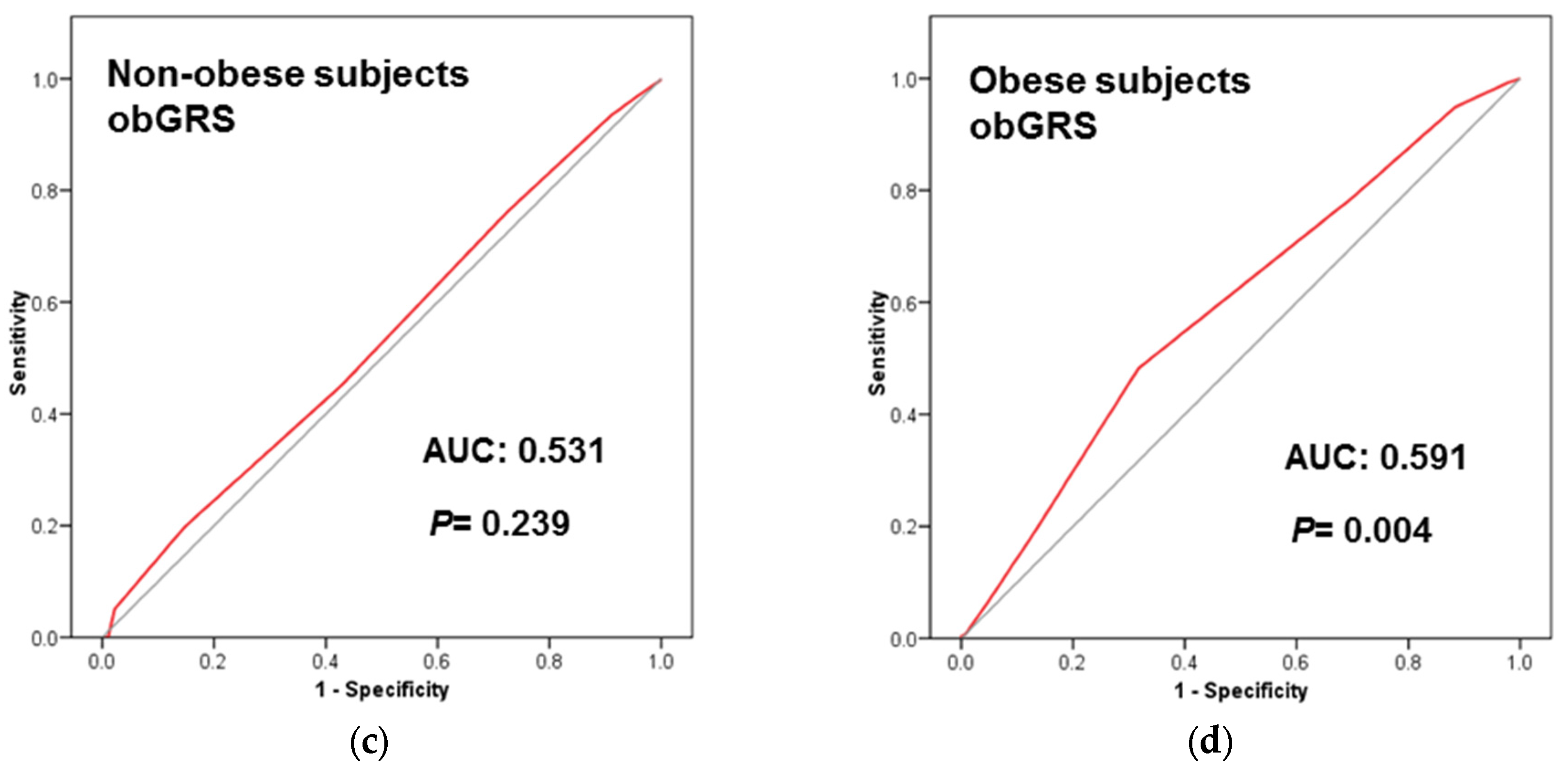
3.5. Obesity Specific-GRS Construction, Association with T2D Prevalence and Estimations of the Predictive Value of These GRS in a Subsample of Participants
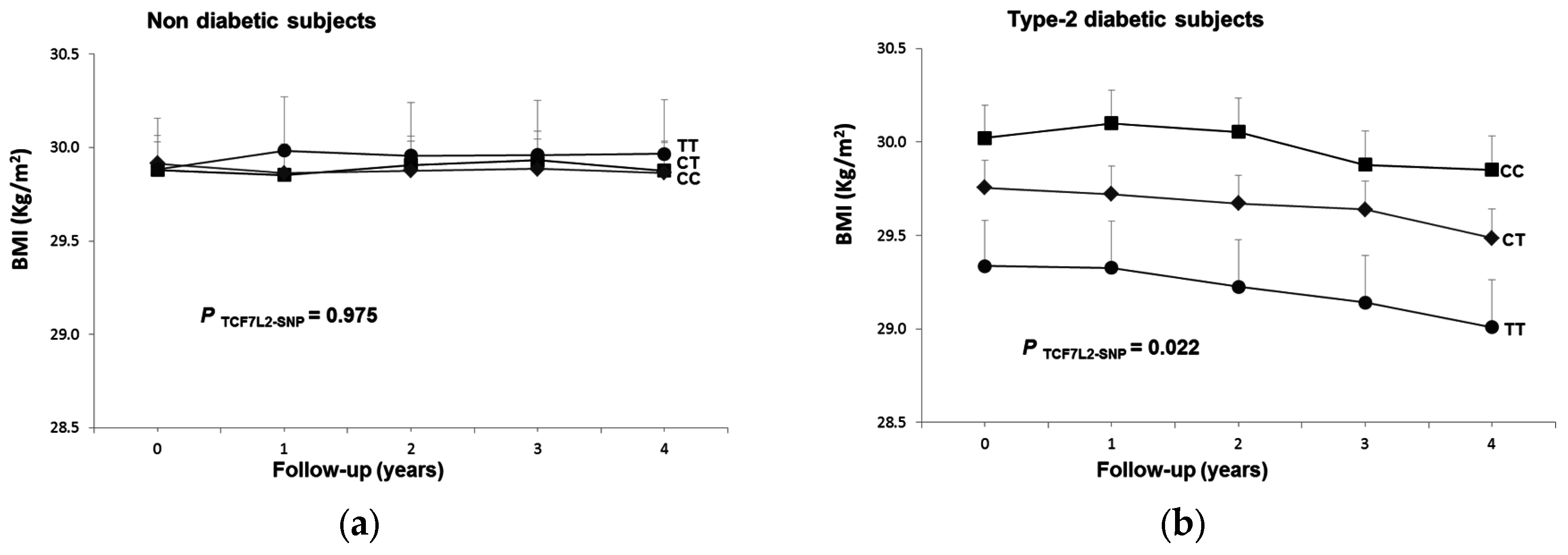
3.6. Longitudinal Interaction between the TCF7L2-rs7903146 Polymorphism and T2D in BMI
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jaacks, L.M.; Siegel, K.R.; Gujral, U.P.; Narayan, K.M. Type 2 diabetes: A 21st century epidemic. Best Pract. Res. Clin. Endocrinol. Metab. 2016, 30, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Kahn, S.E.; Cooper, M.E.; Del Prato, S. Pathophysiology and treatment of type 2 diabetes: Perspectives on the past, present, and future. Lancet 2014, 383, 1068–1083. [Google Scholar] [CrossRef]
- Huang, T.; Qi, Q.; Zheng, Y.; Ley, S.H.; Manson, J.E.; Hu, F.B.; Qi, L. Genetic predisposition to central obesity and risk of type 2 diabetes: Two independent cohort studies. Diabetes Care 2015, 38, 1306–1311. [Google Scholar] [CrossRef] [PubMed]
- Tuomi, T.; Santoro, N.; Caprio, S.; Cai, M.; Weng, J.; Groop, L. The many faces of diabetes: A disease with increasing heterogeneity. Lancet 2014, 383, 1084–1094. [Google Scholar] [CrossRef]
- Timpson, N.J.; Lindgren, C.M.; Weedon, M.N.; Randall, J.; Ouwehand, W.H.; Strachan, D.P.; Rayner, N.W.; Walker, M.; Hitman, G.A.; Doney, A.S.; et al. Adiposity-related heterogeneity in patterns of type 2 diabetes susceptibility observed in genome-wide association data. Diabetes 2009, 58, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Cauchi, S.; Nead, K.T.; Choquet, H.; Horber, F.; Potoczna, N.; Balkau, B.; Marre, M.; Charpentier, G.; Froguel, P.; Meyre, D. The genetic susceptibility to type 2 diabetes may be modulated by obesity status: Implications for association studies. BMC. Med. Genet. 2008, 9, 45. [Google Scholar] [CrossRef] [PubMed]
- Manning, A.K.; Hivert, M.F.; Scott, R.A.; Grimsby, J.L.; Bouatia-Naji, N.; Chen, H.; Rybin, D.; Liu, C.T.; Bielak, L.F.; Prokopenko, I.; et al. A genome-wide approach accounting for body mass index identifies genetic variants influencing fasting glycemic traits and insulin resistance. Nat. Genet. 2012, 44, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Perry, J.R.; Voight, B.F.; Yengo, L.; Amin, N.; Dupuis, J.; Ganser, M.; Grallert, H.; Navarro, P.; Li, M.; Qi, L.; et al. Stratifying type 2 diabetes cases by BMI identifies genetic risk variants in LAMA1 and enrichment for risk variants in lean compared to obese cases. PLoS Genet. 2012, 8, e1002741. [Google Scholar] [CrossRef] [PubMed]
- Grant, S.F.; Thorleifsson, G.; Reynisdottir, I.; Benediktsson, R.; Manolescu, A.; Sainz, J.; Helgason, A.; Stefansson, H.; Emilsson, V.; Helgadottir, A.; et al. Variant of transcription factor 7-like 2 (TCF7L2) gene confers risk of type 2 diabetes. Nat. Genet. 2006, 38, 320–323. [Google Scholar] [CrossRef] [PubMed]
- Voight, B.F.; Scott, L.J.; Steinthorsdottir, V.; Morris, A.P.; Dina, C.; Welch, R.P.; Zeggini, E.; Huth, C.; Aulchenko, Y.S.; Thorleifsson, G.; et al. Twelve type 2 diabetes susceptibility loci identified through large-scale association analysis. Nat. Genet. 2010, 42, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Zhu, Y.; Lü, B.; Xu, F.; Li, X.; Lai, M. TCF7L2 gene polymorphisms and type 2 diabetes risk: A comprehensive and updated meta-analysis involving 121,174 subjects. Mutagenesis 2013, 28, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Bouhaha, R.; Choquet, H.; Meyre, D.; Abid Kamoun, H.; Ennafaa, H.; Baroudi, T.; Sassi, R.; Vaxillaire, M.; Elgaaied, A.; Froguel, P.; et al. TCF7L2 is associated with type 2 diabetes in nonobese individuals from Tunisia. Pathol. Biol. (Paris) 2010, 58, 426–429. [Google Scholar] [CrossRef] [PubMed]
- Lukacs, K.; Hosszufalusi, N.; Dinya, E.; Bakacs, M.; Madacsy, L.; Panczel, P. The type 2 diabetes-associated variant in TCF7L2 is associated with latent autoimmune diabetes in adult Europeans and the gene effect is modified by obesity: A meta-analysis and an individual study. Diabetologia 2012, 55, 689–693. [Google Scholar] [CrossRef] [PubMed]
- Kalnina, I.; Geldnere, K.; Tarasova, L.; Nikitina-Zake, L.; Peculis, R.; Fridmanis, D.; Pirags, V.; Klovins, J. Stronger association of common variants in TCF7L2 gene with nonobese type 2 diabetes in the Latvian population. Exp. Clin. Endocrinol. Diabetes 2012, 120, 466–468. [Google Scholar] [CrossRef] [PubMed]
- Salpea, K.D.; Gable, D.R.; Cooper, J.A.; Stephens, J.W.; Hurel, S.J.; Ireland, H.A.; Feher, M.D.; Godsland, I.F.; Humphries, S.E. The effect of WNT5B IVS3C > G on the susceptibility to type 2 diabetes in UK Caucasian subjects. Nutr. Metab. Cardiovasc. Dis. 2009, 19, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Langenberg, C.; Sharp, S.J.; Franks, P.W.; Scott, R.A.; Deloukas, P.; Forouhi, N.G.; Froguel, P.; Groop, L.C.; Hansen, T.; Palla, L.; et al. Gene-lifestyle interaction and type 2 diabetes: The EPIC interact case-cohort study. PLoS Med. 2014, 11, e1001647. [Google Scholar] [CrossRef] [PubMed]
- Vassy, J.L.; Hivert, M.F.; Porneala, B.; Dauriz, M.; Florez, J.C.; Dupuis, J.; Siscovick, D.S.; Fornage, M.; Rasmussen-Torvik, L.J.; Bouchard, C.; et al. Polygenic type 2 diabetes prediction at the limit of common variant detection. Diabetes 2014, 63, 2172–2182. [Google Scholar] [CrossRef] [PubMed]
- Scott, R.A.; Fall, T.; Pasko, D.; Barker, A.; Sharp, S.J.; Arriola, L.; Balkau, B.; Barricarte, A.; Barroso, I.; Boeing, H.; et al. Common genetic variants highlight the role of insulin resistance and body fat distribution in type 2 diabetes, independent of obesity. Diabetes 2014, 63, 4378–4387. [Google Scholar] [CrossRef] [PubMed]
- Talmud, P.J.; Cooper, J.A.; Morris, R.W.; Dudbridge, F.; Shah, T.; Engmann, J.; Dale, C.; White, J.; McLachlan, S.; Zabaneh, D.; et al. Sixty-five common genetic variants and prediction of type 2 diabetes. Diabetes 2015, 64, 1830–1840. [Google Scholar] [CrossRef] [PubMed]
- Florez, J.C.; Jablonski, K.A.; Bayley, N.; Pollin, T.I.; de Bakker, P.I.; Shuldiner, A.R.; Knowler, W.C.; Nathan, D.M.; Altshuler, D. Diabetes Prevention Program Research Group. TCF7L2 polymorphisms and progression to diabetes in the Diabetes Prevention Program. N. Engl. J. Med. 2006, 355, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Cauchi, S.; Choquet, H.; Gutiérrez-Aguilar, R.; Capel, F.; Grau, K.; Proença, C.; Dina, C.; Duval, A.; Balkau, B.; Marre, M.; et al. Effects of TCF7L2 polymorphisms on obesity in European populations. Obesity (Silver Spring) 2008, 16, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Peter, I.; McCaffery, J.M.; Kelley-Hedgepeth, A.; Hakonarson, H.; Reis, S.; Wagenknecht, L.E.; Kopin, A.S.; Huggins, G.S.; Genetics Subgroup of the Look AHEAD Study. Association of type 2 diabetes susceptibility loci with one-year weight loss in the look AHEAD clinical trial. Obesity (Silver Spring) 2012, 20, 1675–1682. [Google Scholar] [CrossRef] [PubMed]
- Lyssenko, V.; Lupi, R.; Marchetti, P.; Del Guerra, S.; Orho-Melander, M.; Almgren, P.; Sjögren, M.; Ling, C.; Eriksson, K.F.; Lethagen, A.L.; et al. Mechanisms by which common variants in the TCF7L2 gene increase risk of type 2 diabetes. J. Clin. Investig. 2007, 117, 2155–2163. [Google Scholar] [CrossRef] [PubMed]
- Pecioska, S.; Zillikens, M.C.; Henneman, P.; Snijders, P.J.; Oostra, B.A.; van Duijn, C.M.; Aulchenko, Y.S. Association between type 2 diabetes loci and measures of fatness. PLoS ONE 2010, 5, e8541. [Google Scholar] [CrossRef] [PubMed]
- Stolerman, E.S.; Manning, A.K.; McAteer, J.B.; Fox, C.S.; Dupuis, J.; Meigs, J.B.; Florez, J.C. TCF7L2 variants are associated with increased proinsulin/insulin ratios but not obesity traits in the Framingham Heart Study. Diabetologia 2009, 52, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.; Vinay, D.G.; Sovio, U.; Rafiq, S.; Kranthi Kumar, M.V.; Janipalli, C.S.; Evans, D.; Mani, K.R.; Sandeep, M.N.; Taylor, A.; et al. Association study of 25 type 2 diabetes related Loci with measures of obesity in Indian sib pairs. PLoS ONE 2013, 8, e53944. [Google Scholar] [CrossRef] [PubMed]
- Helgason, A.; Pálsson, S.; Thorleifsson, G.; Grant, S.F.; Emilsson, V.; Gunnarsdottir, S.; Adeyemo, A.; Chen, Y.; Chen, G.; Reynisdottir, I.; et al. Refining the impact of TCF7L2 gene variants on type 2 diabetes and adaptive evolution. Nat. Genet. 2007, 39, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Locke, A.E.; Kahali, B.; Berndt, S.I.; Justice, A.E.; Pers, T.H.; Day, F.R.; Powell, C.; Vedantam, S.; Buchkovich, M.L.; Yang, J.; et al. Genetic studies of body mass index yield new insights for obesity biology. Nature 2015, 518, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Martínez-González, M.A.; Corella, D.; Salas-Salvadó, J.; Ros, E.; Covas, M.I.; Fiol, M.; Wärnberg, J.; Arós, F.; Ruíz-Gutiérrez, V.; Lamuela-Raventós, R.M.; et al. Cohort profile: Design and methods of the PREDIMED study. Int. J. Epidemiol. 2012, 41, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Estruch, R.; Ros, E.; Salas-Salvadó, J.; Covas, M.I.; Corella, D.; Arós, F.; Gómez-Gracia, E.; Ruiz-Gutiérrez, V.; Fiol, M.; Lapetra, J.; et al. Primary prevention of cardiovascular disease with a Mediterranean diet. N. Engl. J. Med. 2013, 368, 1279–1290. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Ambrosi, J.; Silva, C.; Catalán, V.; Rodríguez, A.; Galofré, J.C.; Escalada, J.; Valentí, V.; Rotellar, F.; Romero, S.; Ramírez, B.; et al. Clinical usefulness of a new equation for estimating body fat. Diabetes Care 2012, 35, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Salas-Salvadó, J.; Bulló, M.; Estruch, R.; Ros, E.; Covas, M.I.; Ibarrola-Jurado, N.; Corella, D.; Arós, F.; Gómez-Gracia, E.; Ruiz-Gutiérrez, V.; et al. Prevention of diabetes with Mediterranean diets: A subgroup analysis of a randomized trial. Ann. Intern. Med. 2014, 160, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Corella, D.; Carrasco, P.; Sorlí, J.V.; Estruch, R.; Rico-Sanz, J.; Martínez-González, M.Á.; Salas-Salvadó, J.; Covas, M.I.; Coltell, O.; Arós, F.; et al. Mediterranean diet reduces the adverse effect of the TCF7L2-rs7903146 polymorphism on cardiovascular risk factors and stroke incidence: A randomized controlled trial in a high-cardiovascular-risk population. Diabetes Care 2013, 36, 3803–3811. [Google Scholar] [CrossRef] [PubMed]
- Florez, J.C. Leveraging genetics to advance type 2 diabetes prevention. PLoS Med. 2016, 13, e1002102. [Google Scholar] [CrossRef] [PubMed]
- Xia, Q.; Chesi, A.; Manduchi, E.; Johnston, B.T.; Lu, S.; Leonard, M.E.; Parlin, U.W.; Rappaport, E.F.; Huang, P.; Wells, A.D.; et al. The type 2 diabetes presumed causal variant within TCF7L2 resides in an element that controls the expression of ACSL5. Diabetologia 2016, 59, 2360–2368. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Park, S.Y.; Su, J.; Bailey, K.; Ottosson-Laakso, E.; Shcherbina, L.; Oskolkov, N.; Zhang, E.; Thevenin, T.; Fadista, J.; et al. TCF7L2 is a master regulator of insulin production and processing. Hum. Mol. Genet. 2014, 23, 6419–6431. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, R.K.; Mondragon, A.; Chen, L.; Mcginty, J.A.; French, P.M.; Ferrer, J.; Thorens, B.; Hodson, D.J.; Rutter, G.A.; Da Silva Xavier, G. Selective disruption of TCF7L2 in the pancreatic β cell impairs secretory function and lowers β cell mass. Hum. Mol. Genet. 2015, 24, 1390–1399. [Google Scholar] [CrossRef] [PubMed]
- Dimas, A.S.; Lagou, V.; Barker, A.; Knowles, J.W.; Mägi, R.; Hivert, M.F.; Benazzo, A.; Rybin, D.; Jackson, A.U.; Stringham, H.M.; et al. Impact of type 2 diabetes susceptibility variants on quantitative glycemic traits reveals mechanistic heterogeneity. Diabetes 2014, 63, 2158–2171. [Google Scholar] [CrossRef] [PubMed]
- Bailey, K.A.; Savic, D.; Zielinski, M.; Park, S.Y.; Wang, L.J.; Witkowski, P.; Brady, M.; Hara, M.; Bell, G.I.; Nobrega, M.A. Evidence of non-pancreatic beta cell-dependent roles of TCF7L2 in the regulation of glucose metabolism in mice. Hum. Mol. Genet. 2015, 24, 1646–1654. [Google Scholar] [CrossRef] [PubMed]
- Läll, K.; Mägi, R.; Morris, A.; Metspalu, A.; Fischer, K. Personalized risk prediction for type 2 diabetes: The potential of genetic risk scores. Genet. Med. 2016. [Google Scholar] [CrossRef] [PubMed]
- Gamboa-Meléndez, M.A.; Huerta-Chagoya, A.; Moreno-Macías, H.; Vázquez-Cárdenas, P.; Ordóñez-Sánchez, M.L.; Rodríguez-Guillén, R.; Riba, L.; Rodríguez-Torres, M.; Guerra-García, M.T.; Guillén-Pineda, L.E.; et al. Contribution of common genetic variation to the risk of type 2 diabetes in the Mexican Mestizo population. Diabetes 2012, 61, 3314–3321. [Google Scholar] [CrossRef] [PubMed]
- Brito, E.C.; Lyssenko, V.; Renström, F.; Berglund, G.; Nilsson, P.M.; Groop, L.; Franks, P.W. Previously associated type 2 diabetes variants may interact with physical activity to modify the risk of impaired glucose regulation and type 2 diabetes: A study of 16,003 Swedish adults. Diabetes 2009, 58, 1411–1418. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Xing, X.; Hong, J.; Zhang, X.; Yang, W. Genetic variants associated with lean and obese type 2 diabetes in a Han Chinese population: A case-control study. Medicine (Baltimore) 2016, 95, e3841. [Google Scholar] [CrossRef] [PubMed]
- Palomaki, G.E.; Melillo, S.; Marrone, M.; Douglas, M.P. Use of genomic panels to determine risk of developing type 2 diabetes in the general population: A targeted evidence-based review. Genet. Med. 2013, 15, 600–601. [Google Scholar] [CrossRef] [PubMed]
- Evaluation of Genomic Applications in Practice and Prevention (EGAPP) Working Group. Recommendations from the EGAPP Working Group: Does genomic profiling to assess type 2 diabetes risk improve health outcomes? Genet. Med. 2013, 15, 612–617. [Google Scholar]
- Fox, C.S.; Hall, J.L.; Arnett, D.K.; Ashley, E.A.; Delles, C.; Engler, M.B.; Freeman, M.W.; Johnson, J.A.; Lanfear, D.E.; Liggett, S.B.; et al. Future translational applications from the contemporary genomics era: A scientific statement from the American Heart Association. Circulation 2015, 131, 1715–1736. [Google Scholar] [CrossRef] [PubMed]
- Keating, B.J. Advances in risk prediction of type 2 diabetes: Integrating genetic scores with Framingham risk models. Diabetes 2015, 64, 1495–1497. [Google Scholar] [CrossRef] [PubMed]
- Roswall, N.; Ängquist, L.; Ahluwalia, T.S.; Romaguera, D.; Larsen, S.C.; Østergaard, J.N.; Halkjaer, J.; Vimaleswaran, K.S.; Wareham, N.J.; Bendinelli, B.; et al. Association between Mediterranean and Nordic diet scores and changes in weight and waist circumference: Influence of FTO and TCF7L2 loci. Am. J. Clin. Nutr. 2014, 100, 1188–1197. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total (n = 7018) | Non-Diabetic Subjects (n = 3607) | T2D Subjects (n = 3411) | p | ||||
|---|---|---|---|---|---|---|---|
| Age (years) | 67.0 | ±6.2 | 66.6 | ±6.1 | 67.4 | ±6.3 | <0.001 |
| Weight (Kg) | 76.8 | ±11.9 | 76.7 | ±11.7 | 76.9 | ±12.2 | 0.476 |
| BMI (Kg/m2) | 30.0 | ±3.8 | 30.1 | ±3.7 | 29.9 | ±4.0 | 0.042 |
| Waist circumference (cm) | 100.4 | ±10.6 | 99.7 | ±10.6 | 101.2 | ±10.5 | <0.001 |
| Body fat (%) | 39.3 | ±7.4 | 39.9 | ±7.2 | 38.7 | ±7.7 | <0.001 |
| Female sex: n, % | 4025 | (57.4) | 2232 | (61.9) | 1793 | (52.6) | <0.001 |
| Current smokers: n, % | 989 | (14.1) | 581 | (16.1) | 408 | (12.0) | <0.001 |
| TCF7L2-rs7903146: n, % | <0.001 | ||||||
| CC | 2770 | (39.5) | 1612 | (44.7) | 1158 | (33.9) | |
| CT | 3249 | (46.3) | 1569 | (43.5) | 1680 | (49.3) | |
| TT | 999 | (14.2) | 426 | (11.8) | 573 | (16.8) | |
| Intervention groups: n, % | 0.059 | ||||||
| MedDiet + EVOO | 2411 | (34.4) | 1204 | (33.4) | 1207 | (35.4) | |
| MedDiet + nuts | 2316 | (33.0) | 1235 | (34.2) | 1081 | (31.7) | |
| Control group | 2291 | (32.6) | 1168 | (32.4) | 1123 | (32.9) | |
| Energy intake (kcal/day) | 2276 | ±607 | 2322 | ±603 | 2228 | ±607 | <0.001 |
| Total fat (% energy) | 39.2 | ±6.8 | 38.5 | ±6.5 | 39.9 | ±7.0 | <0.001 |
| Saturated fat (% energy) | 10.0 | ±2.3 | 9.7 | ±2.2 | 10.2 | ±2.3 | <0.001 |
| MUFA (% energy) | 19.5 | ±4.6 | 19.2 | ±4.3 | 19.7 | ±4.8 | <0.001 |
| Carbohydrates (% energy) | 41.9 | ±7.2 | 42.8 | ±6.9 | 40.9 | ±7.3 | <0.001 |
| Adherence to the MedDiet | 8.7 | ±2.0 | 8.7 | ±2.0 | 8.6 | ±2.0 | 0.003 |
| Alcohol consumption (g/day) | 8.4 | ±14.2 | 9.1 | ±14.8 | 7.6 | ±13.5 | <0.001 |
| Physical activity (MET.min/day) | 231.6 | ±240.4 | 225.5 | ±226.8 | 238.0 | ±253.8 | 0.030 |
| SBP (mm·Hg) | 149.3 | ±20.8 | 149.0 | ±20.6 | 149.7 | ±21.0 | 0.187 |
| DBP (mm·Hg) | 83.4 | ±11.0 | 84.5 | ±11.0 | 82.2 | ±10.9 | <0.001 |
| Total cholesterol (mg/dL) | 211.0 | ±39.4 | 220.0 | ±39.8 | 201.4 | ±36.6 | <0.001 |
| LDL-C (mg/dL) | 130.3 | ±35.1 | 137.9 | ±36.2 | 122.1 | ±31.8 | <0.001 |
| HDL-C (mg/dL) | 53.8 | ±14.1 | 55.8 | ±14.6 | 51.7 | ±13.2 | <0.001 |
| Triglycerides (mg/dL) | 137.4 | ±79.7 | 132.6 | ±73.9 | 142.4 | ±85.2 | <0.001 |
| Fasting glucose (mg/dL) | 122.2 | ±41.6 | 98.2 | ±16.4 | 147.4 | ±45.1 | <0.001 |
| Non-Obese | p 3 for Interaction | Obese | |||||||
|---|---|---|---|---|---|---|---|---|---|
| n | OR | 95% CI | p-Value | Genotype × Obesity | n | OR | 95% CI | p-Value | |
| Model 1 1 | |||||||||
| TCF7L2 | 0.002 | ||||||||
| CC | 1424 | 1.00 | (reference) | 1346 | 1.00 | (reference) | |||
| CT | 1742 | 1.79 | (1.55–2.08) | 5.5 × 10−15 | 1507 | 1.28 | (1.10–1.49) | 0.0012 | |
| TT | 573 | 2.32 | (1.90–2.85) | 5.4 × 10−16 | 426 | 1.51 | (1.21–1.89) | 0.0003 | |
| p 4: 2.5 × 10−20 | p 4: 0.0002 | ||||||||
| Model 2 2 | |||||||||
| TCF7L2 | |||||||||
| CC | 1.00 | (reference) | 1.00 | (reference) | |||||
| CT | 1.78 | (1.54–2.09) | 1.4 × 10−14 | 0.003 | 1.27 | (1.09–1.48) | 0.0020 | ||
| TT | 2.26 | (1.84–2.78) | 1.6 × 10−14 | 1.53 | (1.22–1.92) | 0.0002 | |||
| p 4: 4.3 × 10−19 | p 4: 0.00017 | ||||||||
| Non-Diabetic | p 3 for Interaction | T2D Subjects | |||||||
|---|---|---|---|---|---|---|---|---|---|
| n | OR | 95% CI | p-Value | Genotype × T2D | n | OR | 95% CI | p-Value | |
| Model 1 1 | |||||||||
| TCF7L2 | 0.008 | ||||||||
| CC | 1612 | 1.00 | (reference) | 1158 | 1.00 | (reference) | |||
| CT | 1569 | 1.05 | (0.91–1.20) | 0.544 | 1680 | 0.78 | (0.67–0.91) | 0.001 | |
| TT | 426 | 0.96 | (0.77–1.19) | 0.694 | 573 | 0.64 | (0.52–0.78) | 2.0 × 10−5 | |
| p 4: 0.685 | p 4: 3.90 × 10−5 | ||||||||
| Model 2 2 | |||||||||
| TCF7L2 | |||||||||
| CC | 1.00 | (reference) | 1.00 | (reference) | |||||
| CT | 1.05 | (0.91–1.21) | 0.529 | 0.014 | 0.77 | (0.66–0.90) | 0.001 | ||
| TT | 0.93 | (0.74–1.16) | 0.493 | 0.63 | (0.51–0.78) | 1.6 × 10−5 | |||
| p 4: 0.528 | p 4: 3.02 × 10−5 | ||||||||
| Obese Subjects 1 (n = 1693) | ||||||||||
| Cases | Non-Cases | Person-Years | Incidence Rate 4 | Model 1 2 | Model 2 3 | |||||
| HR | 95% CI | p-Value | HR | 95% CI | p-Value | |||||
| TCF7L2 genotypes | 0.960 | 0.965 | ||||||||
| CC | 73 | 677 | 4117.5 | 17.7 | 1.00 | (reference) | 1.00 | (reference) | ||
| CT | 79 | 669 | 4203.8 | 18.8 | 1.04 | (0.76–1.44) | 0.777 | 1.05 | (0.76–1.46) | 0.750 |
| TT | 21 | 174 | 1094.0 | 19.2 | 1.01 | (0.62–1.65) | 0.957 | 1.01 | (0.61–1.66) | 0.979 |
| Non-Obese Subjects (n = 1904) | ||||||||||
| Model 1 2 | Model 2 3 | |||||||||
| TCF7L2 genotypes | 0.035 | 0.045 | ||||||||
| CC | 56 | 803 | 4819.0 | 11.6 | 1.00 | (reference) | 1.00 | (reference) | ||
| CT | 57 | 758 | 4661.8 | 12.2 | 1.08 | (0.75–1.57) | 0.671 | 1.14 | (0.79–1.66) | 0.531 |
| TT | 26 | 204 | 1285.7 | 20.2 | 1.82 | (1.14–2.92) | 0.012 | 1.81 | (1.13–2.92) | 0.013 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corella, D.; Coltell, O.; Sorlí, J.V.; Estruch, R.; Quiles, L.; Martínez-González, M.Á.; Salas-Salvadó, J.; Castañer, O.; Arós, F.; Ortega-Calvo, M.; et al. Polymorphism of the Transcription Factor 7-Like 2 Gene (TCF7L2) Interacts with Obesity on Type-2 Diabetes in the PREDIMED Study Emphasizing the Heterogeneity of Genetic Variants in Type-2 Diabetes Risk Prediction: Time for Obesity-Specific Genetic Risk Scores. Nutrients 2016, 8, 793. https://doi.org/10.3390/nu8120793
Corella D, Coltell O, Sorlí JV, Estruch R, Quiles L, Martínez-González MÁ, Salas-Salvadó J, Castañer O, Arós F, Ortega-Calvo M, et al. Polymorphism of the Transcription Factor 7-Like 2 Gene (TCF7L2) Interacts with Obesity on Type-2 Diabetes in the PREDIMED Study Emphasizing the Heterogeneity of Genetic Variants in Type-2 Diabetes Risk Prediction: Time for Obesity-Specific Genetic Risk Scores. Nutrients. 2016; 8(12):793. https://doi.org/10.3390/nu8120793
Chicago/Turabian StyleCorella, Dolores, Oscar Coltell, Jose V. Sorlí, Ramón Estruch, Laura Quiles, Miguel Ángel Martínez-González, Jordi Salas-Salvadó, Olga Castañer, Fernando Arós, Manuel Ortega-Calvo, and et al. 2016. "Polymorphism of the Transcription Factor 7-Like 2 Gene (TCF7L2) Interacts with Obesity on Type-2 Diabetes in the PREDIMED Study Emphasizing the Heterogeneity of Genetic Variants in Type-2 Diabetes Risk Prediction: Time for Obesity-Specific Genetic Risk Scores" Nutrients 8, no. 12: 793. https://doi.org/10.3390/nu8120793
APA StyleCorella, D., Coltell, O., Sorlí, J. V., Estruch, R., Quiles, L., Martínez-González, M. Á., Salas-Salvadó, J., Castañer, O., Arós, F., Ortega-Calvo, M., Serra-Majem, L., Gómez-Gracia, E., Portolés, O., Fiol, M., Díez Espino, J., Basora, J., Fitó, M., Ros, E., & Ordovás, J. M. (2016). Polymorphism of the Transcription Factor 7-Like 2 Gene (TCF7L2) Interacts with Obesity on Type-2 Diabetes in the PREDIMED Study Emphasizing the Heterogeneity of Genetic Variants in Type-2 Diabetes Risk Prediction: Time for Obesity-Specific Genetic Risk Scores. Nutrients, 8(12), 793. https://doi.org/10.3390/nu8120793