1. Introduction
The developmental origins of health and disease (DOHaD) hypothesis proposes that in utero and early life exposures can lead to altered programming of offspring. Such programming events can cause permanent changes in organ development, physiology and metabolism leading to altered disease risk in later life. Suboptimal nutrition, both under- and over-nutrition, during fetal and neonatal development increases susceptibility to a wide range of diseases [
1]. These observations indicate a degree of plasticity during development, in which the fetal phenotype may be altered due to environment cues [
2] to prepare it for the anticipated post-natal environment [
3]. The biological mechanisms underlying this “programming” effect of nutrition during early life are poorly understood, but are likely to involve changes in gene expression.
The protective effect of adequate dietary folate intake and of folic acid supplementation during pregnancy on risk of neural tube defects (NTDs) is well established [
4,
5]. Further, epidemiological evidence suggests that adequate dietary folate intake or supplementation with folic acid during pregnancy may reduce the risk of other congenital defects [
6] and adverse pregnancy outcomes [
7], as well enhancing neurodevelopment [
8] and reducing the risk of severe language delay [
9], autism [
10,
11] and some cancers (leukaemia [
12,
13,
14], brain tumours [
15,
16] and neuroblastoma [
17]) [
18] in children. In rodent models, folate deficiency during pregnancy can cause spontaneous abortion, teratogenic effects in offspring, reduced litter number, and altered offspring body weight [
19,
20]. Sufficient maternal folate intake during pregnancy is essential for successful pregnancy outcomes/normal fetal development and for the long-term health of the offspring.
Folate, a B vitamin, is central to one-carbon metabolism which, in addition to formation of the universal methyl donor,
S-adenosyl-methionine (SAM), interacts with several other cellular pathways including amino acid metabolism and biosynthesis of purine and pyrimidines [
21]. SAM is critical for the methylation of biological molecules including DNA, lipids and proteins. Epigenetic mechanisms (including methylation of DNA and of histones) which regulate gene expression are susceptible to modification via altered SAM availability in response to changes in folate intake (reviewed in [
22]). Therefore, we hypothesised that inadequate folate supply during pregnancy alters programming of the offspring via changes in gene expression and that this is responsible for the observed adverse effects of this maternal nutritional insult on pregnancy outcomes and offspring health in later life. Mammals respond to inadequate nutrient supply by prioritising allocation of nutrients to specific purposes [
23,
24] which may result in cell, tissue and organ differences in gene expression. Little is known about such inter-organ differences in fetal gene expression in response to restricted folate supply. We hypothesised that the fetal liver and placenta, which represent organs with long and short-term consequences for the developing animal, would be subject to distinctly different responses to folate depletion. To test these hypotheses, we quantified genome-wide gene expression patterns in fetal liver and placenta in response to maternal folate depletion before, and throughout, pregnancy.
2. Experimental Section
2.1. Animal Husbandry and Experimental Diets
All animal procedures were approved by the Newcastle University Ethics Review Committee and the UK Home Office (Project Licence number 60/3979) and have been described previously [
25]. Animals were housed in the Comparative Biology Centre, Newcastle University at 20–22 °C and with 12 h light and dark cycles. Fresh water was available ad libitum. Female C57BL/6J mice were allocated at random to either a low folate (0.4 mg folic acid/kg diet) or normal folate diet (2 mg folic acid/kg diet) (6 g of allocated diet was offered to each mouse per day), and maintained on this diet for 4 weeks prior to mating. Diet compositions were modified from AIN-93G [
26] and have been described previously [
25].
l-amino acids were used as a protein substitute. All ingredients, other than folic acid, were included in both diets at the same concentrations to avoid potential confounding through other dietary factors. The degree of folate depletion induced by feeding the diet containing 0.4 mg folic acid/kg was sufficient to impose a nutritional stress (evidenced by reduced circulating concentrations of folate) but not so severe as to limit reproduction. The normal folate diet contained 2 mg folic acid/kg diet which is considered sufficient to support breeding and maintenance in this species. Mice were time mated i.e., a male was added to a cage containing two females overnight and removed the following morning. Pregnant females, identified by the presence of a vaginal plug, were re-caged and offered 10 g/day of allocated diet throughout pregnancy. At 17.5 days gestation, dams were killed for collection of blood and organs.
2.2. Sample Collection
Animals were anesthetised using gaseous isoflurane, blood was removed by cardiac puncture and animals were killed by cervical dislocation. Blood was collected and stored in EDTA tubes. Whole blood 5-methyltetrahydrofolate (THF) and 5–10-methylTHF concentrations were measured by liquid chromatography-tandem mass spectrometry (LC-MS/MS) and data reported previously [
25]. The uterus, containing all fetuses and placentas, was removed and placed immediately in ice cold PBS. The liver and placenta of each fetus were removed, weighed and snap frozen in liquid nitrogen and stored at −80 °C until required for RNA extraction.
2.3. RNA Extraction
To avoid any potential influence of sex on resultant data, only male tissues were analysed. Fetal sex was determined by polymerase chain reaction (PCR) of the sex determining region-Y (
SRY) gene using DNA extracted from embryonic tail tissue [
25,
27]. RNA was extracted from whole fetal livers and placentas of males only using Tri-reagent (Sigma-Aldrich, Gillingham, Dorset, SP8 4XT, UK) and following the manufacturer’s instructions. Briefly, 50 mg tissue was homogenised in 500 μL Tri-reagent on ice. A further 500 μL Tri-reagent and 200 μL chloroform were added, then the sample was mixed by inversion and incubated on ice for 5 min. Samples were then centrifuged at 13,500 rpm for 15 min at 4 °C. The upper aqueous phase was removed, RNA was precipitated by incubation on ice with 500 μL isopropanol for 10 min, after which it was collected by centrifugation at 13,500 rpm for 10 min at 4 °C. The supernatant fluid was removed and the pellet was washed with 1 mL 75% ethanol for 10 min before centrifuging at 6000 rpm for 5 min at 4 °C. The supernatant fluid was removed, and the pellet was allowed to dry for 10 min before resuspending the RNA in 40 μL water. Contaminating DNA was removed using RQ1 DNase (Promega, Southampton, Hampshire, SO16 7NS, UK). RNA integrity was checked on an agarose gel and concentration and purity were measured using a Nanodrop spectrophotometer (Thermo Fisher, Waltham, MA, USA).
2.4. Gene Expression Arrays
RNA from each tissue was pooled for three male fetuses per litter (5 μg/fetus with a final concentration of 500 ng/μL) and hybridised to a single array for a total of 12 litters (n = 6 per dietary group) for each organ. Where a litter had more than three male fetuses, the three fetuses with weights closest to the mean litter weight were analysed.
Genome-wide transcript abundance was determined by ServiceXS (Plesmanlaan 1/D, 2333 BZ Leiden, The Netherlands) on the Affymetrix GeneChip platform with the NuGO mouse array (NuGO_Mm1a520177). This array comprises over 24,000 probe sets, covering the majority of established genes. Before the labelling process, the integrity of all RNA samples (RNA Integrity Number (RIN) > 8) was confirmed using the Agilent 2100 Bioanalyser (Agilent Technologies, Stockport, Cheshire, SK8 3GY, UK). Output data were supplied as Affymetrix CEL files and imported into R (version 2.15.3) using the Affy package [
28]. Data from liver and placenta were pre-processed separately using gcRMA background correction and quantile normalisation [
29] to correct for batch effects and other technical confounders. To maximize sensitivity and specificity, updated Entrez gene probe-set annotation was used from the BrainArray project [
30] (version 14.1.0) resulting in 16,270 re-annotated probesets mapping to unique transcripts. Statistical analysis comparing the two diet groups was performed separately for each organ using the empirical Bayes approach of the Limma package [
31] which performs a moderated
t-test. On filtering genes for significant changes using False Discovery Rate (FDR) values calculated using the Benjamini and Hochberg method (FDR < 0.05), no genes remained statistically significant. Pathway analysis can in part replace FDR corrections by testing for the regulation of related genes, which suffers less from oversampling. Genes were, therefore, considered to show a differential expression relevant for further analysis in enrichment analysis in response to maternal folate depletion if there was a significant (
p < 0.05) increase or reduction of at least 1.2 fold. DAVID [
32] was used to carry out Gene Ontology enrichment analysis and to investigate KEGG pathways affected by maternal folate depletion. The threshold for significance for Gene Ontology enrichment analysis was set at
p < 0.05 (corrected for multiple testing), and at
p < 0.05 (uncorrected) for KEGG pathway enrichment analysis. Additional pathway analysis was carried out using PathVisio 3.2.0 and the curated pathway collection of WikiPathways (download date: 1 September 2015), applying the same parameters for significant fold-change as stated above, imposing a Z score of 1.9 for significance to filter for probable changed pathways. All raw and processed microarray data have been deposited in the ArrayExpress database (accession ID E-MTAB-3940).
2.5. Validation of Gene Expression Changes Using Real-Time PCR
To confirm the gene expression changes observed in the microarray analysis, real-time PCR was performed on each individual (i.e., not pooled) fetal RNA sample that was analysed by microarray hybridisation, focusing on 13 gene targets. Target genes were selected on the basis of being the most up or down regulated in the liver and placenta. Gene transcripts analysed in placental RNA were Cyp21a1, Hbb-y, Slco1b2, Ptgs2, Vcan, Lrrn4, and Mettl7b and in fetal liver RNA were Asgr1, Hamp, Actc1, Ckm, Tnnc2 and Smpx.
RNA (1 μg) was reverse transcribed using Quantitect Reverse Transcription kit (Qiagen, Cat No. 205313) according to the manufacturer’s instructions. Briefly, 1 μg of RNA was incubated with 2 μL genomic DNA Wipeout buffer in a reaction volume of 14 μL at 42 °C for 2 min to remove any genomic DNA. The mixture was then placed immediately on ice. RNA was incubated with 1 μL Quantiscript reverse transcriptase, 1 μL RT primer mix and 4 μL Quantiscript RT buffer for 15 min at 42 °C, after which time it was incubated at 95 °C for 3 min. The cDNA samples generated were then diluted 1:9 with water for use in real time PCR.
Prior to sample analysis, expression values, linearity and efficiency of each assay were determined through cDNA standard curves completed for each transcript measured. Transcript levels of the genes of interest were measured on a Roche Lightcycler 480 (Roche Applied Science, Burgess Hill, West Sussex, RH15 9RY, UK) in a total reaction volume of 25 μL using 2.5 μL Quantitect sybr green transcript specific primers (Qiagen, see
Supplementary Materials Table S1 for details of manufacturer’s individual catalogue numbers), 12.5 μL Quantitect SYBR green mix (Qiagen Cat. No. 204145), 1 μL diluted cDNA and 9 μL water and cycling parameters 95 °C for 5 min (1 cycle); 95 °C for 10 s followed by 60 °C for 30 s (40 cycles); followed by a final melt curve analysis and cooling to 40 °C. Using the delta CT method, transcript levels of the genes of interest were normalised to
GAPDH transcript levels which were determined using the same procedure and parameters described.
2.6. In Silico Analysis of Gene Promoter Regions for Transcription Factor (TF) Binding Sites
Genomatix software (
https://www.genomatix.de/v3.5 24 July 2015) was employed to obtain promoter sequences for genes of interest using the Gene2Promoter function. Promoter sequences were analysed for common transcription factor binding sites using the Common TFs function. Lists of TFs belonging to the transcription factor binding site families identified were downloaded from Genomatix and compared with fetal liver gene expression data.
2.7. Statistical Analysis
Statistical analysis of array data is described above. For all other datasets, data distributions were examined by the Kolmogorov-Smirnov test and all datasets were normally distributed. Analysis of variance (Statistical Package for the Social Sciences (SPSS) version 21, IBM, Armonk, New York 10504, NY, USA) was used to examine the effects of diet on placental weight, placental efficiency and gene expression analysis on data obtained from RT PCR analysis. p < 0.05 was considered statistically significant.
4. Discussion
Here we describe gene expression changes at the transcriptome level in the placenta and fetal liver in response to maternal folate depletion before mating and throughout pregnancy. Ames’ triage theory posits that if availability of a specific nutrient is inadequate, Darwinian processes ensure that essential functions, i.e., those required for short-term survival and/or reproduction, dependent on that nutrient that are protected at the expense of those functions that are less essential i.e., where reduced function does not have short term negative consequences. However, such prioritisation of some most critical (short-term) functions may impact adversely on other functions which may have long-term insidious effects that increase risk of diseases associated with ageing [
23,
24]. In response to inadequate maternal folate intake during pregnancy, we hypothesised that, due to different organ-specific functions, individual fetal organs would implement differential prioritisation hierarchies and thus display differential gene expression changes in response to folate depletion. We chose to investigate gene expression in the liver and placenta as examples of organs with very different roles in mammalian function across the lifecourse. In addition, the liver represents a major site for folate storage and metabolism, whilst the placenta (containing both maternal and fetal cells) is the route for delivery of nutrients (including folate) from the dam to the fetus during development. Our observations support our hypothesis that specific fetal organs exhibit specific transcriptional responses to maternal folate depletion. It is plausible that these organ-specific changes in gene expression are due to differential pathway prioritisation in each organ which reflect their contrasting functions.
The primary function of the placenta is the transfer of nutrients to the fetus, so it is essential that this function is protected to support fetal growth and development. In response to maternal folate depletion, we saw no changes in pathways associated with nutrient transport, suggesting that placental transfer of nutrients to the fetus was protected despite the reduced maternal folate intake. We observed no teratogenic effects of this reduced nutrient supply on the offspring [
25]. It is important to note that we analysed whole placental samples containing both maternal and fetal cell populations and so we are unable to determine the cellular origin of the observed expression changes.
The liver is a highly metabolic organ responsible for a wide range of functions that are important at all life stages and include defence of the body against xenobiotics, assisting with digestion and with metabolism of absorbed nutrients, regulating blood lipids, synthesis of a wide range of secreted proteins and hormonal regulation. As such, during periods of undernutrition, it may not be possible for the liver to maintain all important functions, so that those functions essential for short-term survival and/or reproduction may be prioritised. The sequela is that some functions not critical for short-term survival but which are important for long term health may be de-prioritised leading to impaired function which may have significant long-term adverse effects for the organism. We observed that maternal folate depletion altered several processes and pathways, each of which could impact adversely on long-term liver function and organismal health. These included “adipogenesis genes”, “fatty acid biosynthesis” and “iron homeostasis” pathways. Whilst only one time point was investigated here (so that we are unable to comment on long-term gene expression changes or phenotypic outcomes), these observations are consistent with the fact that diets deficient in folate and choline can induce non-alcoholic fatty liver disease (NAFLD) in rodent models [
36]. We observed up-regulation of genes involved in “fatty acid biosynthesis” (5/5 differentially expressed genes present on the pathway), and an abundance of up-regulated genes involved in the “adipogenesis genes” pathway (10/16 differentially expressed genes present on the pathway). Up-regulation of these genes may result in overall up-regulation of the pathways that potentially could result in altered hepatic fatty acid metabolism in offspring. Indeed, we observed that when the offspring of folate depleted dams were fed a high fat diet from weaning [
37], they had significantly increased plasma triacylglycerol (TAG) concentrations whilst plasma TAG concentrations were unaffected in the offspring of folate replete dams [
37]. Taken together, these data suggest that maternal folate depletion results in an altered response to high fat feeding which may be via programmed transcriptional changes in pathways associated with fat metabolism in the liver.
This analysis has focussed on the genes which were differentially expressed in the liver and in the placenta in response to maternal folate depletion. However, it is important to recognise that there are alternative explanations for genes which did not change in response to this nutritional insult. Whilst such unaltered genes may be components of pathways which are especially critical to that particular organ, it is also possible that these genes are simply unresponsive to folate supply. The present study design is unable to distinguish between these alternatives.
It is important to highlight that most of the gene expression changes observed in this study were relatively small and not statistically significant when applying the more stringent FDR value so that these data should be interpreted with caution. However, our observations are consistent with findings from other studies [
33,
34] and confirms the subtle, but pervasive, effects on gene expression of folate/methyl donor depletion. Furthermore, it is pertinent to point out that our gene expression studies were undertaken in male offspring only and it is possible, if theoretically unlikely, that effects could be different in females.
Whilst we show clearly that folate responsive gene expression changes are organ specific, the mechanisms behind the observed organ specific expression changes are less clear. A plausible mechanistic explanation could be based on inter-organ differences in expression of transcription factors (TFs) when exposed to inadequate folate intake. We tested this hypothesis by identifying all TFs which were differentially expressed in the liver and in the placenta in response to maternal folate depletion. In the fetal liver, we found that expression of 73 TFs differed in folate replete vs. folate depleted offspring, which accounted for 7.4% of the altered genes. In contrast, in the placenta only 22 TFs (4.8% of altered genes) displayed altered expression. Consistent with the idea that organ-specific changes in gene expression profiles are driven by organ-specific changes in TF expression, there was very little overlap between the TFs that were altered in liver and placenta—only one TF, Med6, was common between the organs.
It is plausible that changes in epigenetic processes are responsible for the organ-specific patterns in gene expression in response to folate depletion. Epigenetic marks, including DNA methylation, are modifiable by diet and other environmental factors and constitute an important and flexible system for regulating gene expression [
38,
39]. Folate is central to one-carbon metabolism and the formation of the universal methyl donor SAM, which is critical for the methylation of biological molecules including DNA. Indeed the influence of dietary folate intake on DNA methylation patterns has been reported widely [
25,
34,
40,
41,
42] and may be responsible for the organ specific folate responsive gene expression changes observed here.
Although previous studies have reported gene specific expression changes in the rat placenta in response to increased levels of folate in the maternal diet [
42,
43], to the best of our knowledge this is the first report describing gene expression changes in response to low maternal folate (or methyl donor) intake in this organ, and the first study to investigate this at the transcriptomic level. However, previous studies have used transcriptomic approaches to investigate gene expression changes in the liver in response to low folate-containing diets [
33,
34,
35,
36,
44,
45] so we compared our data with outcomes from these studies. Whilst there were differences in gene expression profiles between studies, likely to be due to differences in rodent models, diet composition, duration and timing of the nutrient insult and the particular transcriptomic approaches employed (
Table 10), we uncovered a key set of genes which were differentially expressed in the liver in response to low dietary folate/methyl donor intake across multiple studies (
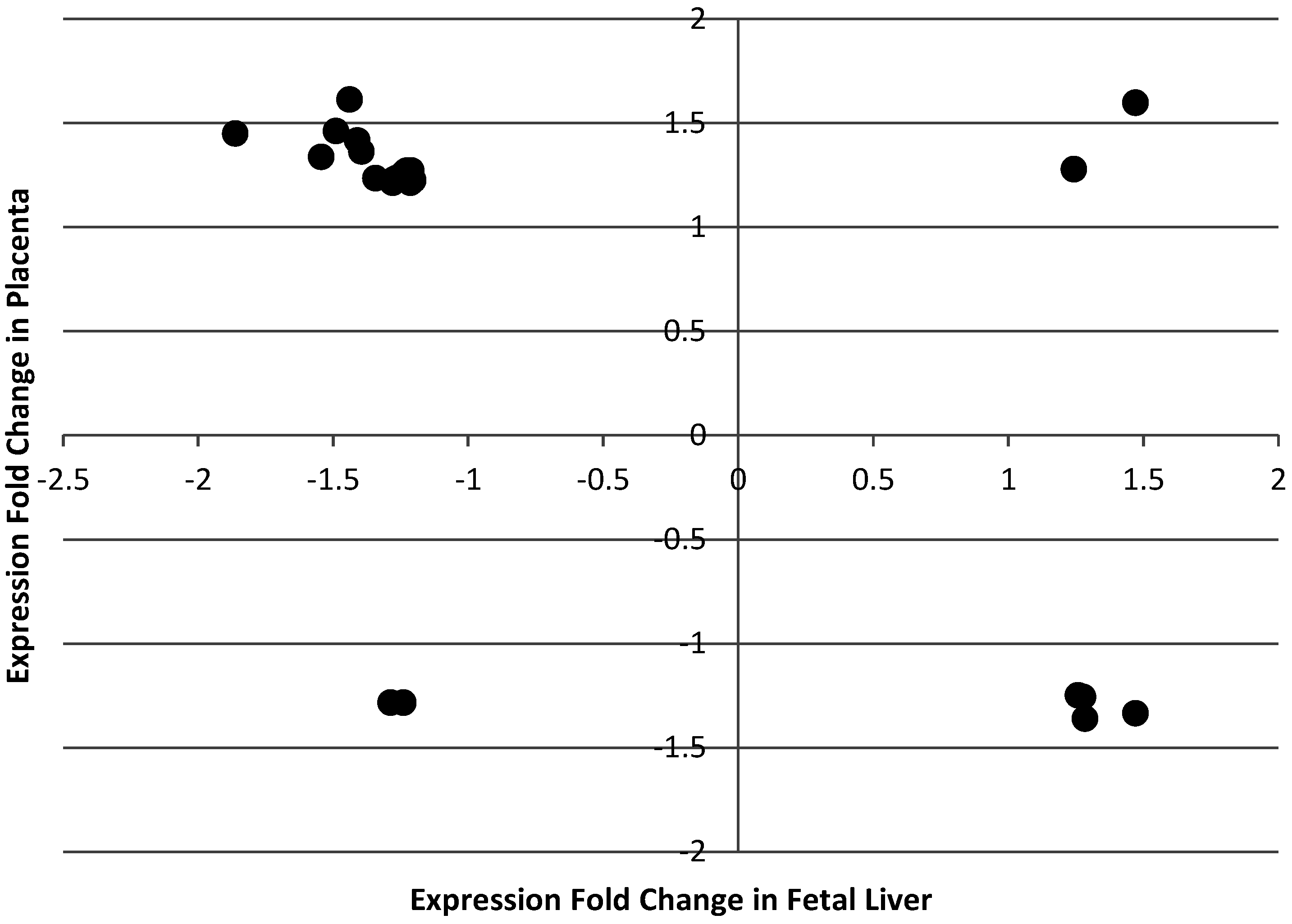
Table 11). Importantly, expression of none of these genes was altered in placenta by maternal folate depletion, indicating that these changes reflect a liver-specific response rather than being genes that are generally malleable in response to perturbation in folate supply. Moreover, this observation substantiates our suggestion that the transcriptome responses of fetal liver and placenta to folate restriction are distinct and physiologically-appropriate. Of the key set of 21 genes differentially expressed across multiple studies (
Table 11), the use of DAVID [
32] to carry out Gene Ontology enrichment analysis suggested that three genes are involved in wound healing i.e.,
JUB,
ENTPD2 and
SERPINE1 and, thus, altered transcription of genes associated with wound healing may help explain the observed association between low methyl donor intake and liver damage [
36]. Furthermore, the discovery of these key folate responsive genes may provide candidate biomarkers for future studies of folate adequacy. Such studies should test the effects of different doses of folate without potential confounding from other dietary factors which may have been an issue in some of the studies included in
Table 10.
To investigate potential mechanisms through which expression of these key genes in the liver are altered in response to methyl donor intake, the promoter regions were interrogated for the presence of common transcription factor binding sites. Whilst binding sites for five families of transcription factors were found to be highly represented in the promoters of these 21 genes, similar occurrence rates of these same transcription factor binding sites were also observed when seven randomly selected sets of genes were analysed. This suggests that this specific set of TFs is not responsible for co-ordinating transcriptional responses to maternal folate/methyl-donor supply in the liver. However, it is possible that there are regulatory regions outside the promoters of the genes investigated here which would merit investigation in future studies.


 ,
, 

{kind=link}