Capsaicin Supplementation Reduces Physical Fatigue and Improves Exercise Performance in Mice


Abstract
:1. Introduction
2. Materials and Methods
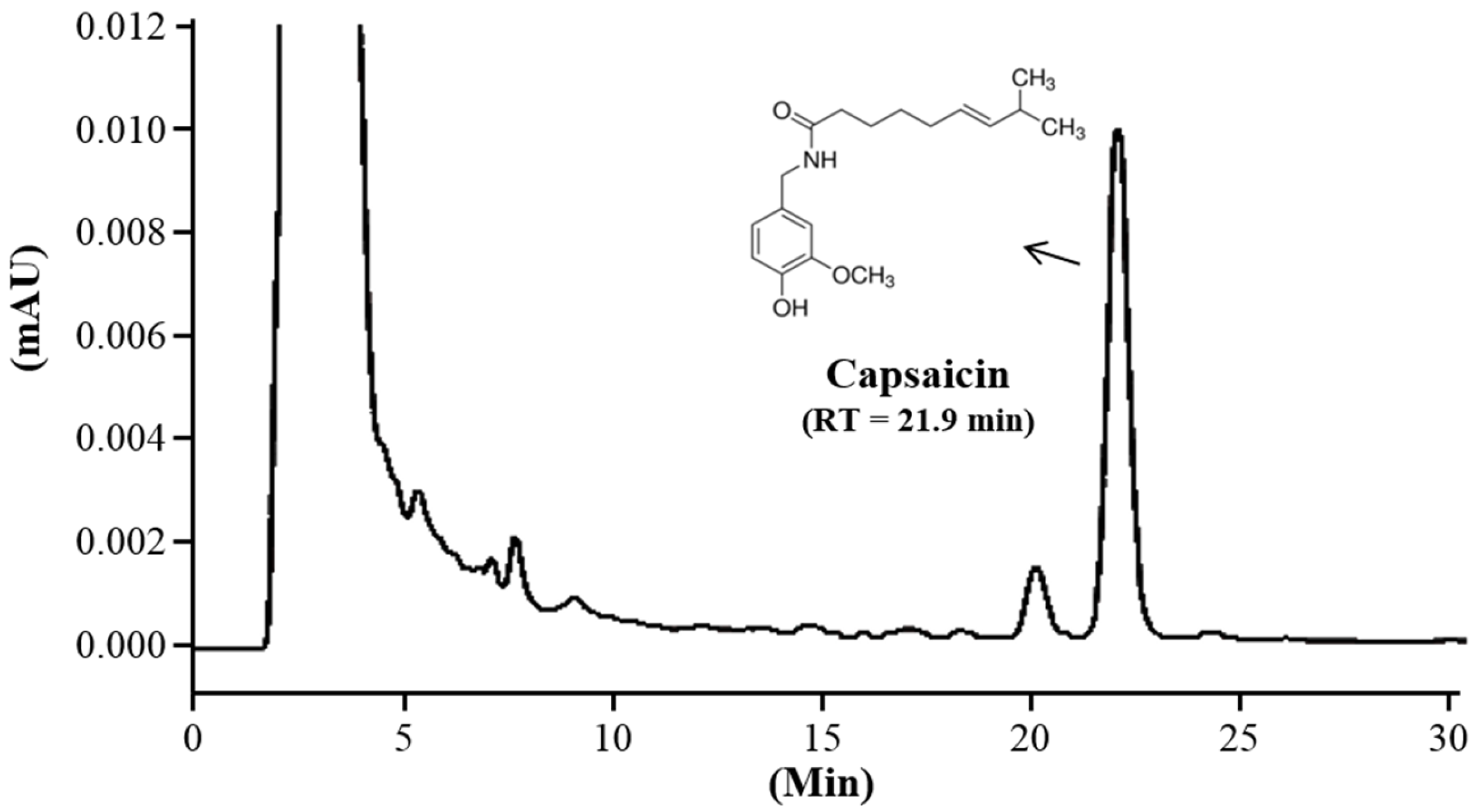
2.1. High-Performance Liquid Chromatography (HPLC) Analysis
2.2. Animals and Experiment Design
2.3. CAP Preparation and Supplementation
2.4. Sample Collection
2.5. Forelimb Grip Strength Test
2.6. Swimming Exercise Performance Test
2.7. Determination of Fatigue-Associated Biochemical Variables
2.8. Clinical Biochemical Profiles
2.9. Tissue Glycogen Determination
2.10. Histological Staining of Tissue
2.11. Statistical Analysis
3. Results
3.1. Content of Capsaicin in CAP
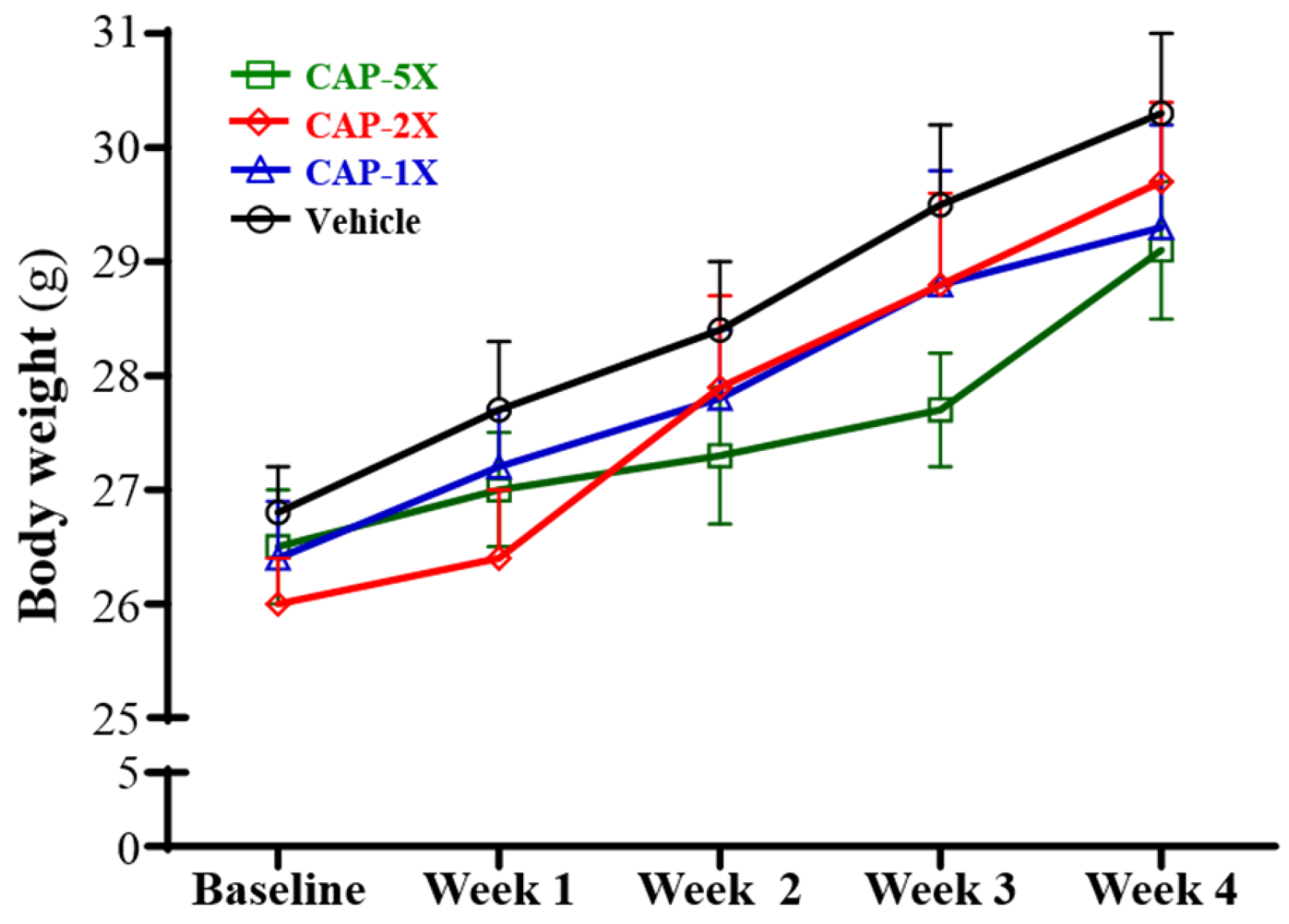
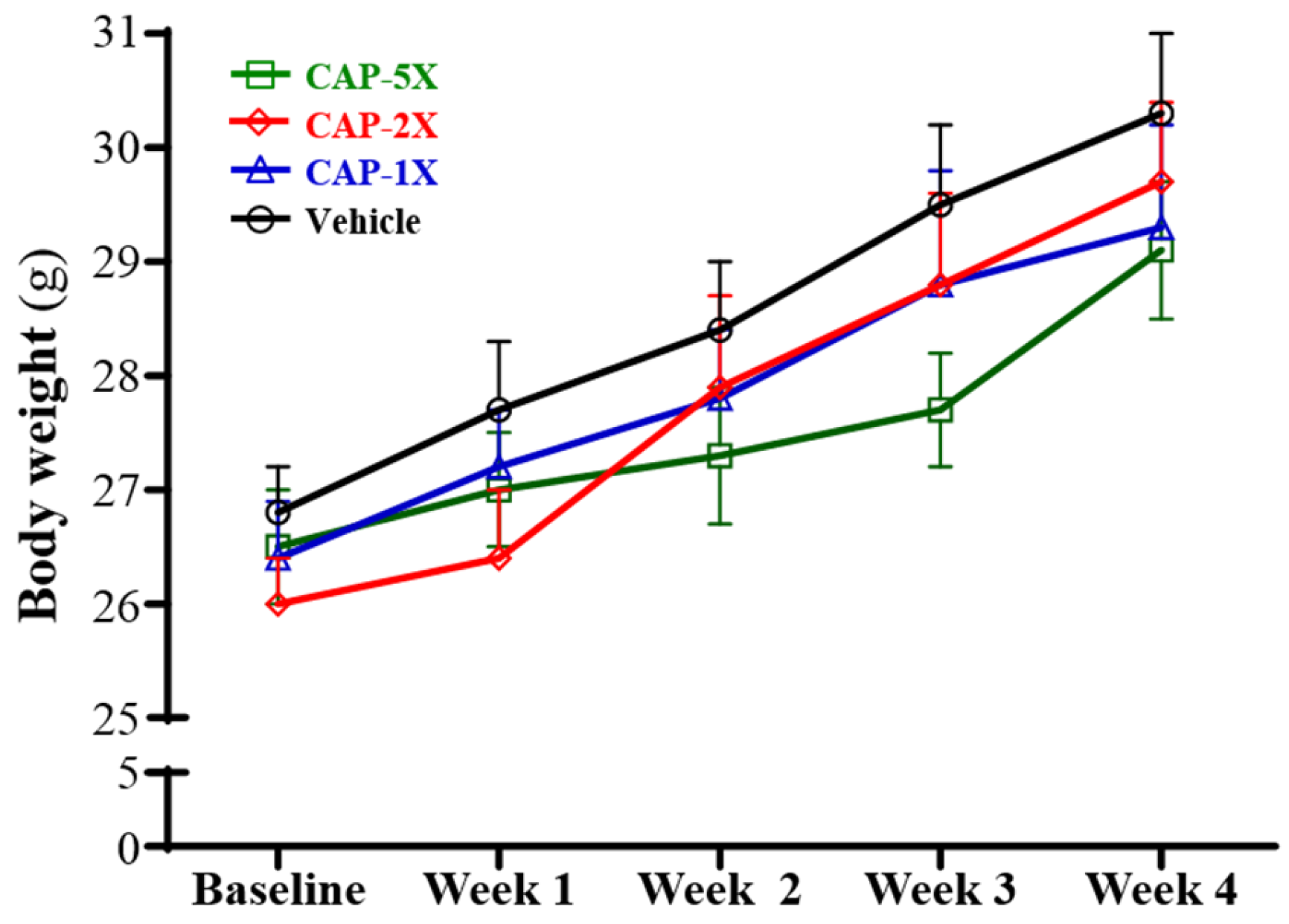
3.2. Effect of CAP Supplementation on Body Weight and Organ Weights
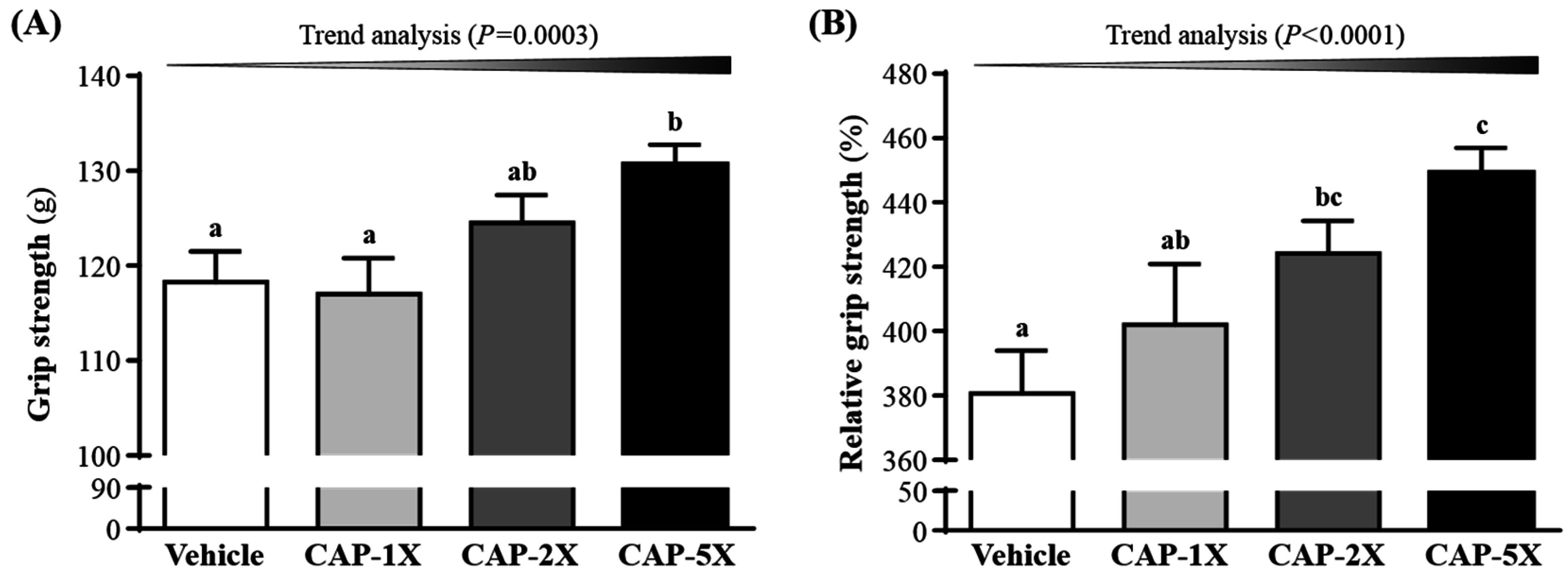
3.3. Effect of CAP on Forelimb Grip Strength
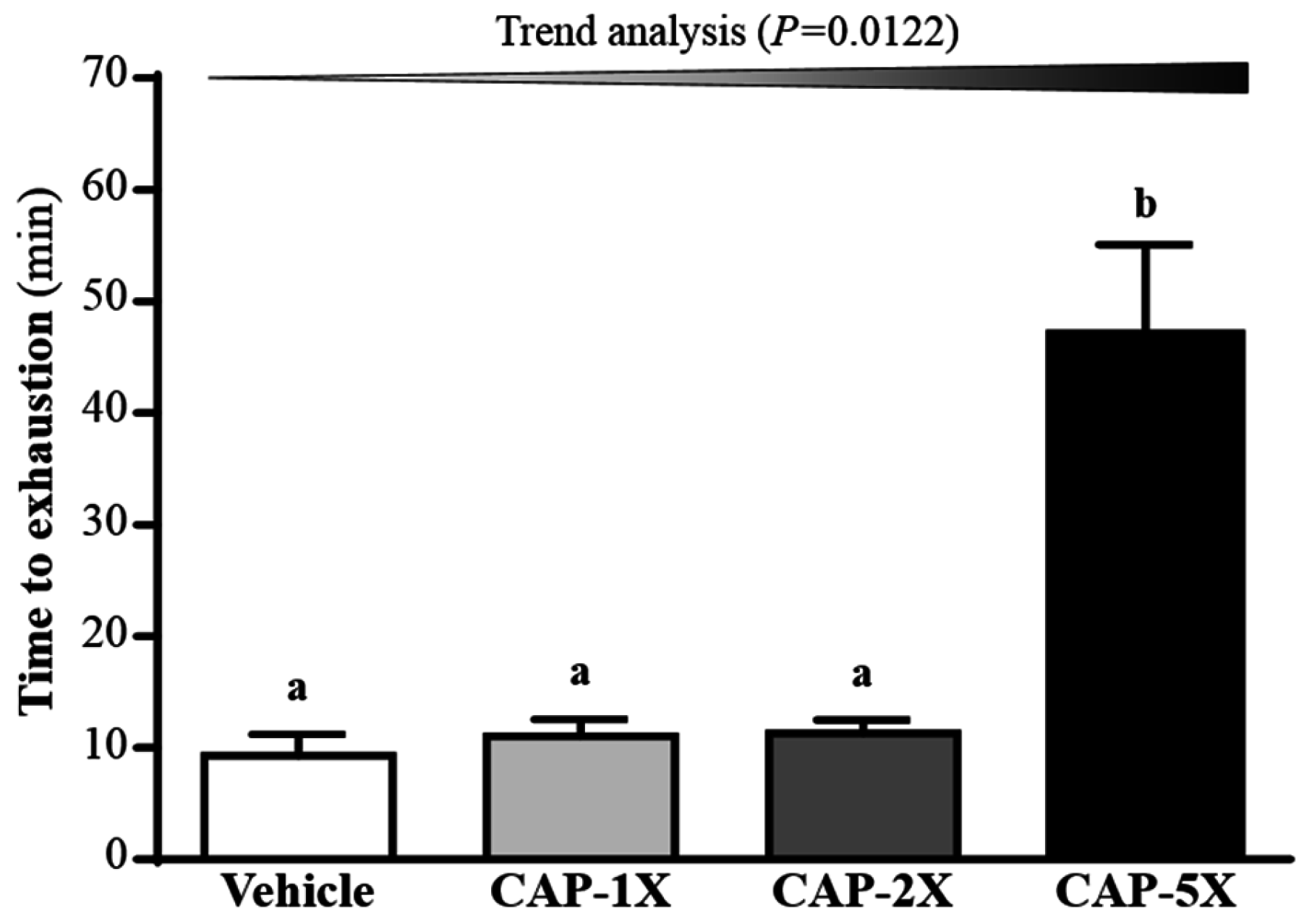
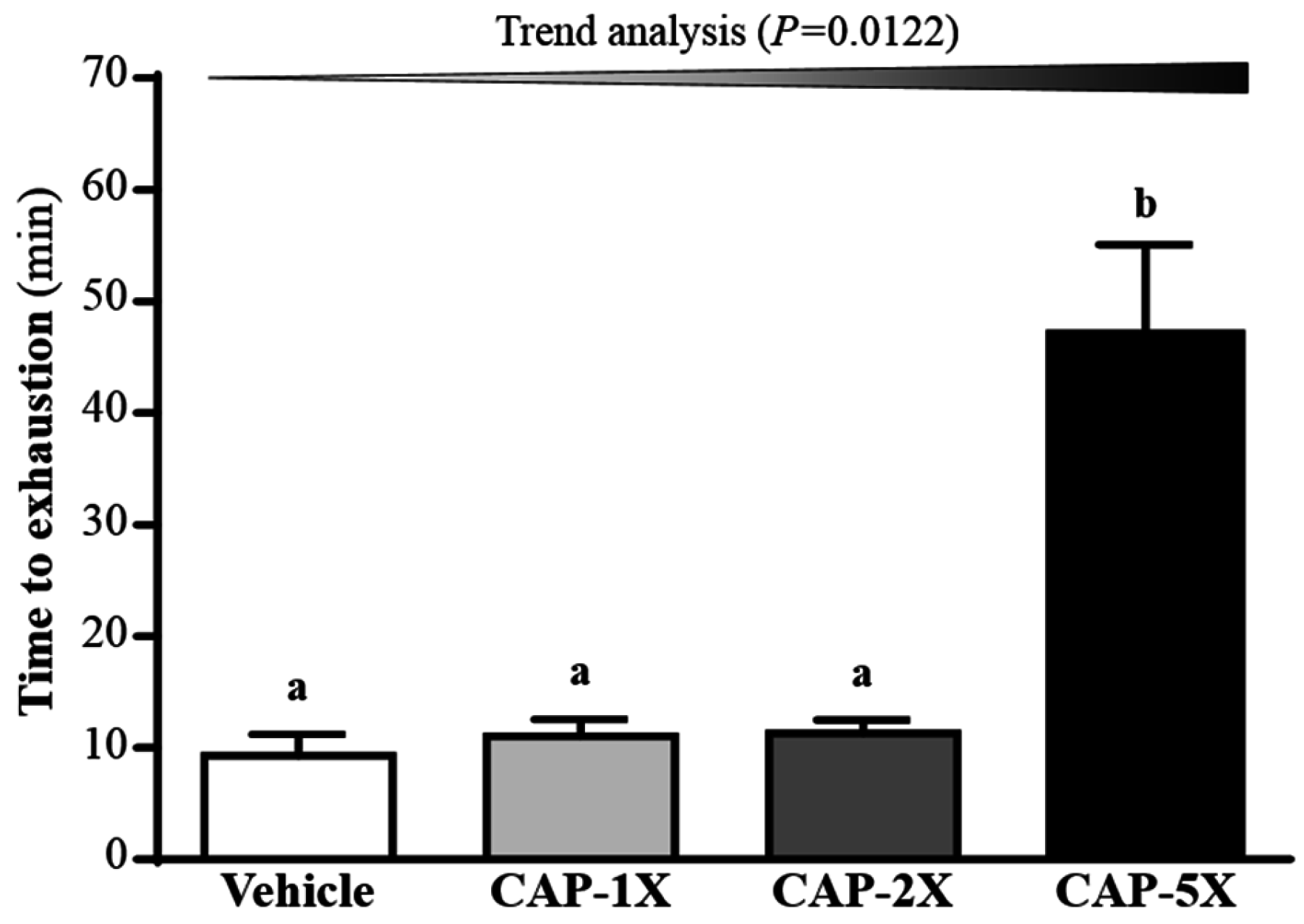
3.4. Effect of CAP Supplementation on Exhaustive Swimming Test
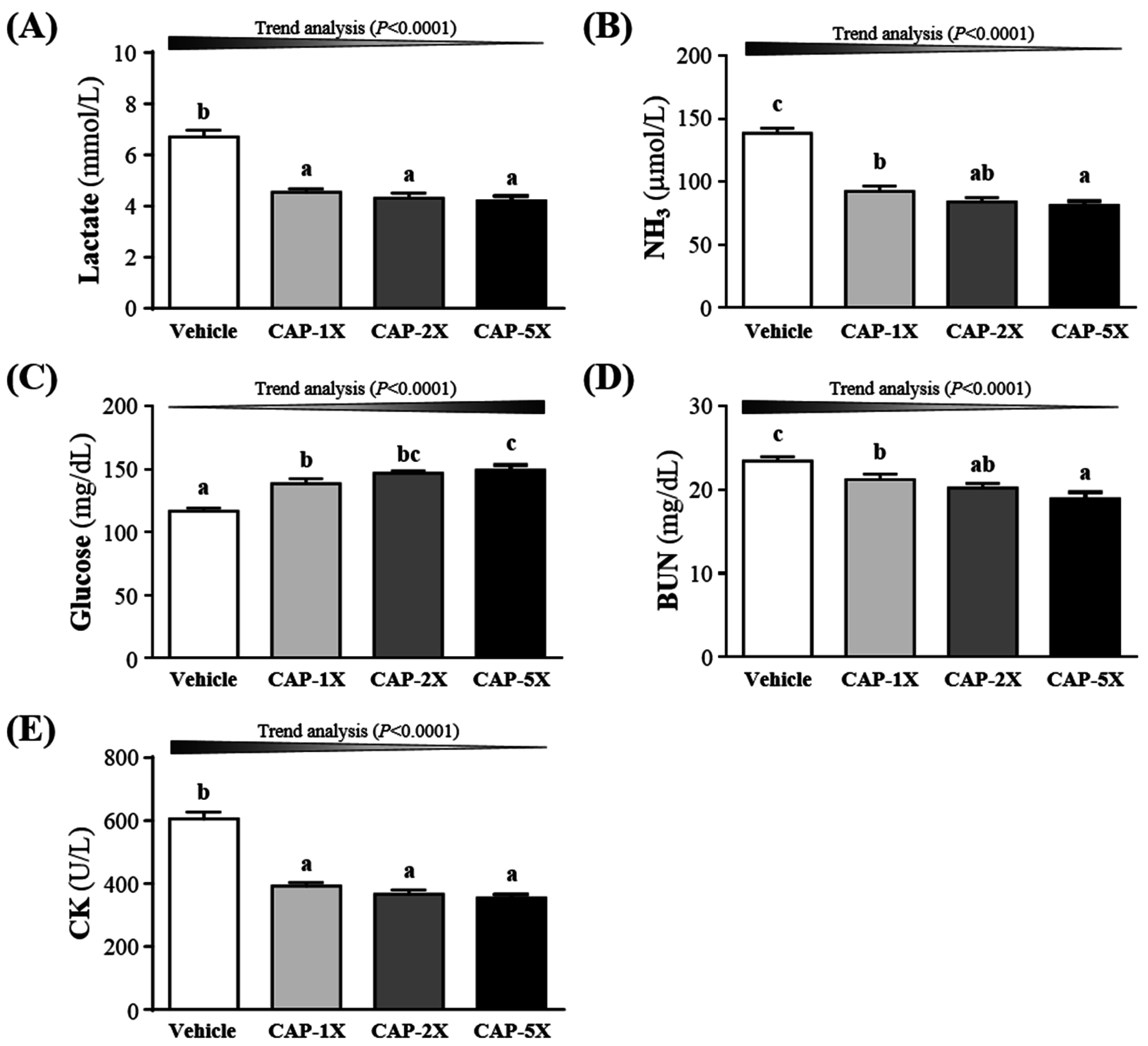
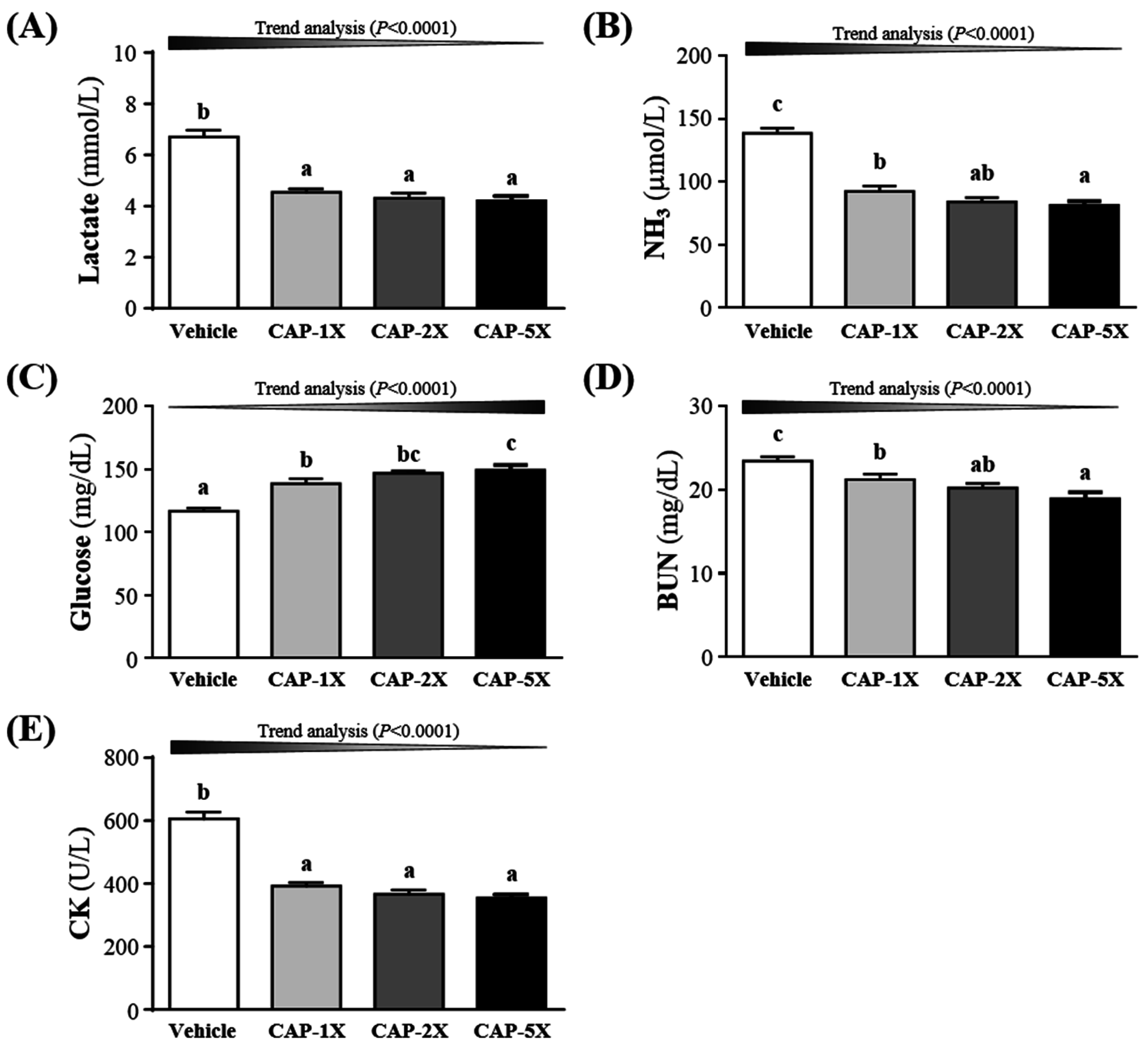
3.5. Effect of CAP Supplementation on Exercise Fatigue-Related Indicators after Acute Exercise
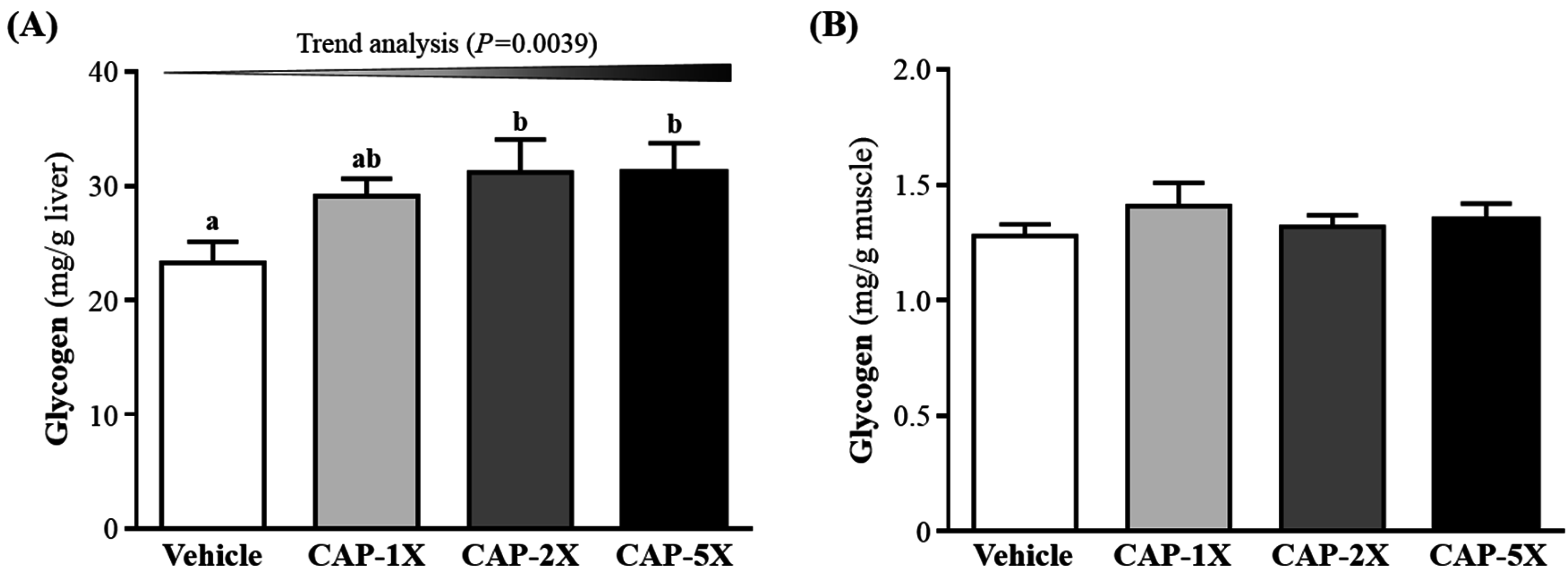
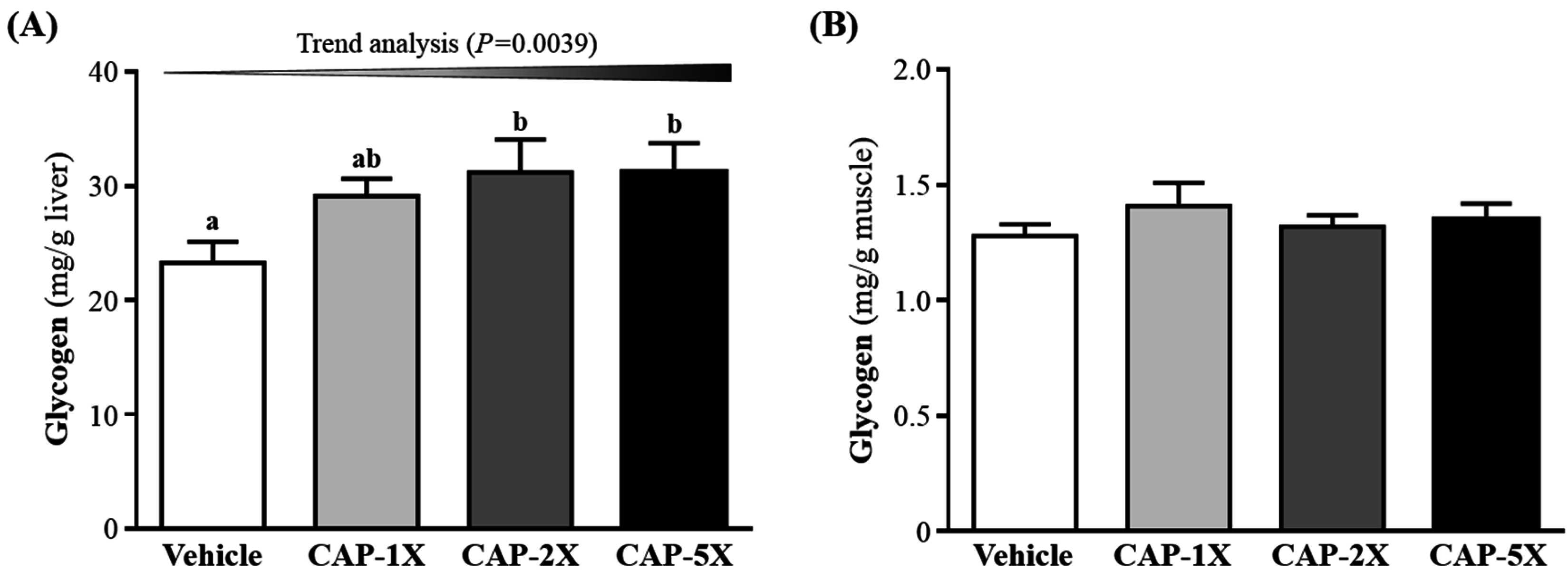
3.6. Effect of CAP Supplementation on Hepatic and Muscular Glycogen Level
3.7. Effect of CAP Supplementation on Biochemical Analyses at the End of the Experiment
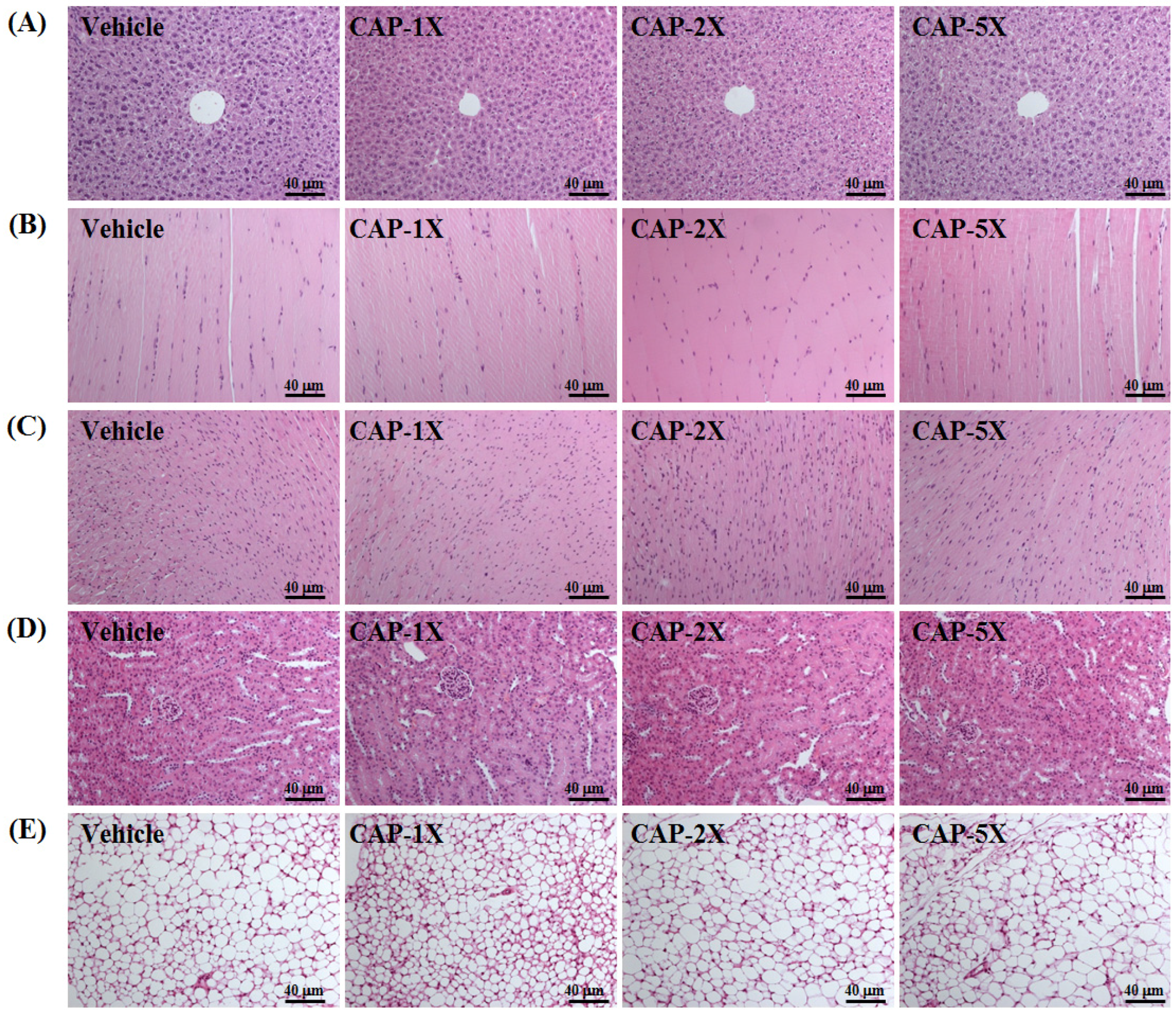
3.8. Effect of CAP Supplementation on Histological Examinations at the End of the Experiment
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Naj, A. Peppers: A Story of Hot Pursuits; Knopf: New York, NY, USA, 1992. [Google Scholar]
- Qiu, J.; Niu, X.; Wang, J.; Xing, Y.; Leng, B.; Dong, J.; Li, H.; Luo, M.; Zhang, Y.; Dai, X.; et al. Capsaicin protects mice from community-associated methicillin-resistant Staphylococcus aureus pneumonia. PLoS ONE 2012, 7, e33032. [Google Scholar] [CrossRef] [PubMed]
- Arora, R.; Gill, N.S.; Chauhan, G.; Rana, A.C. An overview about versatile molecule capsaicin. Int. J. Pharm. Sci. Drug Res. 2011, 4, 280–286. [Google Scholar]
- Purseglove, J.W.; Brown, E.G.; Green, C.L.; Robbins, S.R.J. Spices. Longman 1981, 2, 10–99. [Google Scholar]
- Sung, Y.; Chang, Y.Y.; Ting, N.L. Capsaicin biosynthesis in water-stressed hot pepper fruits. Bot. Bull. Acad. Sin. 2005, 46, 35–42. [Google Scholar]
- Cheema, S.K.; Pant, M.R. Estimation of capsaicin in seven cultivated varieties of Capsicum Annuum L. Res. J. Pharm. Biol. Chem. Sci. 2011, 2, 701–706. [Google Scholar]
- Supalkova, V.; Stavelikova, H.; Krizkova, S.; Adam, V.; Horna, A.; Havel, L.; Ryant, P.; Babula, B.; Kizek, R. Study of capsaicin content in various parts of pepper fruit by liquid chromatography with electrochemical detection. Acta Chim. Slov. 2007, 54, 55–59. [Google Scholar]
- Derry, S.; Sven-Rice, A.; Cole, P.; Tan, T.; Moore, R.A. Topical capsaicin (high concentration) for chronic neuropathic pain in adults. Cochrane Database Syst. Rev. 2013, 28, Cd007393. [Google Scholar]
- Chen, K.S.; Chen, P.N.; Hsieh, Y.S.; Lin, C.Y.; Lee, Y.H.; Chu, S.C. Capsaicin protects endothelial cells and macrophage against oxidized low-density lipoprotein-induced injury by direct antioxidant action. Chem. Biol. Interact. 2015, 228, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.; Lee, S.H. Anticancer Properties of Capsaicin against Human Cancer. Anticancer Res. 2016, 36, 837–843. [Google Scholar] [PubMed]
- Kang, J.H.; Tsuyoshi, G.; Le Ngoc, H.; Kim, H.M.; Tu, T.H.; Noh, H.J.; Kim, C.S.; Choe, S.Y.; Kawada, T.; Yoo, H.; et al. Dietary capsaicin attenuates metabolic dysregulation in genetically obese diabetic mice. J. Med. Food 2011, 14, 310–315. [Google Scholar] [CrossRef] [PubMed]
- Janssens, P.L.; Hursel, R.; Martens, E.A.; Westerterp-Plantenga, M.S. Acute effects of capsaicin on energy expenditure and fat oxidation in negative energy balance. PLoS ONE 2013, 8, e67786. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Ma, L.; Zhao, Z.; He, H.; Yang, D.; Feng, X.; Ma, S.; Chen, X.; Zhu, T.; Cao, T.; et al. TRPV1 activation improves exercise endurance and energy metabolism through PGC-1alpha upregulation in mice. Cell Res. 2012, 22, 551–564. [Google Scholar] [CrossRef] [PubMed]
- Lotteau, S.; Ducreux, S.; Romestaing, C.; Legrand, C.; Van Coppenolle, F. Characterization of functional TRPV1 channels in the sarcoplasmic reticulum of mouse skeletal muscle. PLoS ONE 2013, 8, e58673. [Google Scholar] [CrossRef] [PubMed]
- Hudson, A.S.; Kunstetter, A.C.; Damasceno, W.C.; Wanner, S.P. Involvement of the TRPV1 channel in the modulation of spontaneous locomotor activity, physical performance and physical exercise-induced physiological responses. Braz. J. Med. Biol. Res. 2016, 49, e5183. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Azechi, H.; Board, M. Essential role of excessive tryptophan and its neurometabolites in fatigue. Can. J. Neurol. Sci. 2012, 39, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Fitts, R.H. Cellular mechanisms of muscle fatigue. Physiol. Rev. 1994, 74, 49–94. [Google Scholar] [PubMed]
- You, L.; Zhao, M.; Regenstein, J.M.; Ren, J. In vitro antioxidant activity and in vivo anti-fatigue effect of loach (Misgurnus anguillicaudatus) peptides prepared by papain digestion. Food Chem. 2011, 124, 188–194. [Google Scholar] [CrossRef]
- Wu, R.E.; Huang, W.C.; Liao, C.C.; Chang, Y.K.; Kan, N.W.; Huang, C.C. Resveratrol protects against physical fatigue and improves exercise performance in mice. Molecules 2013, 18, 4689–4702. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C.; Chiu, W.C.; Chuang, H.L.; Tang, D.W.; Lee, Z.M.; Wei, L.; Chen, F.A.; Huang, C.C. Effect of curcumin supplementation on physiological fatigue and physical performance in mice. Nutrients 2015, 7, 905–921. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.C.; Huang, W.C.; Chiu, C.C.; Chang, Y.K.; Huang, C.C. Whey protein improves exercise performance and biochemical profiles in trained mice. Med. Sci. Sports Exerc. 2014, 46, 1517–1524. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Y.; Huang, W.C.; Liu, C.C.; Wang, M.F.; Ho, C.S.; Huang, W.P.; Hou, C.C.; Chuang, H.L.; Huang, C.C. Pumpkin (Cucurbita moschata) fruit extract improves physical fatigue and exercise performance in mice. Molecules 2012, 17, 11864–11876. [Google Scholar] [CrossRef] [PubMed]
- Radley, H.G.; de Luca, A.; Lynch, G.S.; Grounds, M.D. Duchenne muscular dystrophy: Focus on pharmaceutical and nutritional interventions. Int. J. Biochem. Cell Biol. 2007, 39, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Katunuma, N.; Okada, M.; Nishii, Y. Regulation of the urea cycle and TCA cycle by ammonia. Adv. Enzyme Regul. 1996, 4, 317–336. [Google Scholar] [CrossRef]
- Young, A.J.; Castellani, J.W. Exertion-induced fatigue and thermoregulation in the cold. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2001, 128, 769–776. [Google Scholar] [CrossRef]
- Mun, J.M.; Ok, H.M.; Kwon, O. Corn gluten hydrolysate and capsaicin have complimentary actions on body weight reduction and lipid-related genes in diet-induced obese rats. Nutr Res. 2014, 34, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Kazuya, Y.; Tonson, A.; Pecchi, E.; Dalmasso, C.; Vilmen, C.; Fur, Y.L.; Bernard, M.; Bendahan, D.; Giannesini, B. A single intake of capsiate improves mechanical performance and bioenergetics efficiency in contracting mouse skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2014, 15, E1110–E1119. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.P.; Anand, T.; Singsit, D.; Khanum, F.; Anilakumar, K.R. Evaluation of antioxidant and anti-fatigue properties of Trigonella foenum-graecum L. in rats subjected to weight loaded forced swim test. Pharmacogn. J. 2013, 5, 66–71. [Google Scholar] [CrossRef]
- Yue, Z.; Xiuhong, Z.; Shuyan, Y.; Zhonghua, Z. Effect of Inonotus Obliquus Polysaccharides on physical fatigue in mice. J. Tradit. Chin. Med. 2015, 35, 468–472. [Google Scholar] [PubMed]
- Cairns, S.P. Lactic acid and exercise performance: Culprit or friend? Sports Med. 2006, 36, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Haramizu, S.; Mizunoya, W.; Masuda, Y.; Ohnuki, K.; Watanabe, T.; Yazawa, S.; Fushiki, T. Capsiate, a nonpungent capsaicin analog, increases endurance swimming capacity of mice by stimulation of vanilloid receptors. Biosci. Biotechnol. Biochem. 2006, 70, 774–781. [Google Scholar] [CrossRef] [PubMed]
- Bassini-Cameron, A.; Monteiro, A.; Gomes, A.; Werneck-de-Castro, J.P.; Cameron, L. Glutamine protects against increases in blood ammonia in football players in an exercise intensity-dependent way. Br. J. Sports Med. 2008, 42, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Verlander, J.W.; Chu, D.; Lee, H.W.; Handlogten, M.E.; Weiner, I.D. Expression of glutamine synthetase in the mouse kidney: Localization in multiple epithelial cell types and differential regulation by hypokalemia. Am. J. Physiol.-Ren. Physiol. 2013, 305, F701–F713. [Google Scholar] [CrossRef] [PubMed]
- Suh, S.H.; Paik, I.Y.; Jacobs, K. Regulation of blood glucose homeostasis during prolonged. Mol. Cells 2007, 23, 272–279. [Google Scholar] [PubMed]
- Emhoff, C.A.; Messonnier, L.A.; Horning, M.A.; Fattor, J.A.; Carlson, T.J.; Brooks, G.A. Gluconeogenesis and hepatic glycogenolysis during exercise at the lactate threshold. J. Appl. Physiol. 2013, 114, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, M. Exercise, muscle, and CHO metabolism. Scand. J. Med. Sci. Sports 2015, 25, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Oh, T.W.; Ohta, F. Dose-dependent effect of capsaicin on endurance capacity in rats. Br. J. Nutr. 2003, 90, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Hwang, J.T.; Park, H.S.; Kwon, D.Y.; Kim, M.S. Capsaicin stimulates glucose uptake in C2C12 muscle cells via the reactive oxygen species (ROS)/AMPK/p38 MAPK pathway. Biochem. Biophys. Res. Commun. 2013, 13, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Yashiro, K.; Tonson, A.; Pecchi, É.; Vilmen, C.; le Fur, Y.; Bernard, M.; Bendahan, D.; Giannesini, B. Capsiate supplementation reduces oxidative cost of contraction in exercising mouse skeletal muscle in vivo. PLoS ONE 2015, 1, e0128016. [Google Scholar] [CrossRef] [PubMed]
- Langenberg, C.; Wan, L.; Egi, M.; May, C.N.; Bellomo, R. Renal blood flow in experimental septic acute renal failure. Kidney Int. 2006, 69, 1996–2002. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.H.; Kim, H.J.; Oh, G.S.; Shen, A.; Lee, S.; Choe, S.K.; Park, R.; So, H.S. Capsaicin ameliorates cisplatin-induced renal injury through induction of heme oxygenase-1. Mol. Cells 2014, 37, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Baird, M.F.; Graham, S.M.; Baker, J.S.; Bickerstaff, G.F. Creatine-kinase- and exercise-related muscle damage implications for muscle performance and recovery. J. Nutr. Metab. 2012, 2012, 960363. [Google Scholar] [CrossRef] [PubMed]
- Houmard, J.A.; Costill, D.L.; Mitchell, J.B.; Park, S.H.; Fink, W.J.; Burns, J.M. Testosterone, cortisol, and creatine kinase levels in male distance runners during reduced training. Int. J. Sports Med. 1990, 11, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, M. Skeletal muscle metabolism during exercise in humans. Clin. Exp. Pharmacol. Physiol. 2000, 27, 225–228. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.M.; Kawada, T.; Ishihara, K.; Inoue, K.; Fushiki, T. Increase in swimming endurance capacity of mice by capsaicin-induced adrenal catecholamine secretion. Biosci. Biotechnol. Biochem. 1997, 61, 1718–1723. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.Y.; Kim, Y.S.; Ryu, S.Y.; Cha, M.R.; Yon, G.H.; Yang, H.J.; Kim, M.J.; Kang, S.; Park, S. Capsiate improves glucose metabolism by improving insulin sensitivity better than capsaicin in diabetic rats. J. Nutr. Biochem. 2013, 24, 1078–1085. [Google Scholar] [CrossRef] [PubMed]
- Koopmans, S.J.; Leighton, B.; DeFronzo, R.A. Neonatal de-afferentation of capsaicin-sensitive sensory nerves increases in vivo insulin sensitivity in conscious adult rats. Diabetologia 1998, 41, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.H.; Edfawy, M.; Mansour, A.; Hamed, A.A. Antioxidant and antiapoptotic effects of capsaicin against carbon tetrachloride-induced hepatotoxicity in rats. Toxicol. Ind. Health 2012, 28, 428–438. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, H.; Kawada, N.; Kaimori, J.Y.; Kitamura, H.; Moriyama, T.; Rakugi, H.; Takahara, S.; Isaka, Y. Febuxostat suppressed renal ischemia-reperfusion injury via educed oxidative stress. Biochem. Biophys. Res. Commun. 2012, 427, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.W.; Li, Y.J.; Deng, H.W. Early and delayed protection by capsaicin against reperfusion injury in rat hearts. Eur. PMC 1999, 20, 912–916. [Google Scholar]
- Li, L.; Chen, J.; Ni, Y.; Feng, X.; Zhao, Z.; Wang, P.; Sun, J.; Yu, H.; Yan, Z.; Liu, D.; et al. TRPV1 activation prevents nonalcoholic fatty liver through UCP2 upregulation in mice. Pflugers Arch. 2012, 463, 727–732. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Vehicle | CAP-1X | CAP-2X | CAP-5X | Trend Analysis |
|---|---|---|---|---|---|
| Initial BW (g) | 26.8 ± 0.4 | 26.4 ± 0.5 | 26.0 ± 0.4 | 26.5 ± 0.5 | 0.1161 |
| Final BW (g) | 30.3 ± 0.7 | 29.3 ± 0.9 | 29.7 ± 0.7 | 29.1 ± 0.6 | 0.3282 |
| Food intake (g/day) | 5.57 ± 0.02 b | 5.52 ± 0.26 a,b | 5.17± 0.00 a | 5.34 ± 0.03 a,b | 0.6056 |
| Water intake (mL/day) | 6.37± 0.08 b | 6.14 ± 0.22 a,b | 5.86 ± 0.03 a | 6.14 ± 0.20 a,b | 0.0875 |
| Liver (g) | 1.41 ± 0.04 | 1.37 ± 0.04 | 1.37 ± 0.04 | 1.46 ± 0.02 | 0.2536 |
| Muscle (g) | 0.30 ± 0.01 | 0.30 ± 0.00 | 0.30 ± 0.01 | 0.30 ± 0.00 | 0.6938 |
| Kidney (g) | 0.38 ± 0.01 | 0.38 ± 0.00 | 0.38 ± 0.01 | 0.38 ± 0.01 | 0.9420 |
| Heart (g) | 0.18 ± 0.01 | 0.16 ± 0.01 | 0.15 ± 0.00 | 0.16 ± 0.00 | 0.7942 |
| UFP (g) | 0.20 ± 0.01 | 0.19 ± 0.02 | 0.20 ± 0.02 | 0.20 ± 0.01 | 0.7942 |
| BAT (g) | 0.08 ± 0.003 | 0.09 ± 0.002 | 0.08 ± 0.004 | 0.09 ± 0.003 | 0.9519 |
| Relative liver weight (%) | 4.73 ± 0.18 | 4.92 ± 0.10 | 4.73 ± 0.18 | 5.03 ± 0.14 | 0.3336 |
| Relative muscle weight (%) | 1.00 ± 0.01 | 1.02 ± 0.02 | 1.03 ± 0.03 | 1.04 ± 0.02 | 0.3257 |
| Relative kidney weight (%) | 1.28 ± 0.05 | 1.28 ± 0.03 | 1.31 ± 0.03 | 1.30 ± 0.05 | 0.9771 |
| Relative heart weight (%) | 0.59 ± 0.03 b | 0.54 ± 0.03 a,b | 0.51 ± 0.02 a | 0.56 ± 0.02 a,b | 0.6981 |
| Relative UFP weight (%) | 0.68 ± 0.03 | 0.66 ± 0.08 | 0.69 ± 0.08 | 0.67 ± 0.05 | 0.9121 |
| Relative BAT weight (%) | 0.28 ± 0.01 | 0.30 ± 0.01 | 0.29 ± 0.02 | 0.30 ± 0.01 | 0.5119 |
| Parameter | Vehicle | CAP-1X | CAP-2X | CAP-5X | Trend Analysis |
|---|---|---|---|---|---|
| AST (U/L) | 95 ± 5 b | 80 ± 2 a | 80 ± 3 a | 77 ± 3 a | 0.0003 |
| ALT (U/L) | 45 ± 3 d | 37 ± 2 c | 31 ±1 a,b | 30 ± 2 a | <0.0001 |
| Albumin (g/dL) | 3.26 ± 0.04 | 3.28 ± 0.04 | 3.26 ± 0.04 | 3.28 ± 0.03 | 0.8838 |
| TP (g/dL) | 5.04 ± 0.06 | 4.98 ± 0.07 | 5.04 ± 0.05 | 5.05 ± 0.02 | 0.5837 |
| BUN (mg/dL) | 22.3 ± 0.6 c | 18.8 ± 0.3 b | 17.9 ± 0.2 b | 16.6 ± 0.5 a | <0.0001 |
| Creatinine (mg/dL) | 0.29 ± 0.01 c | 0.25 ± 0.01 b | 0.24 ± 0.01 b | 0.20 ± 0.01 a | <0.0001 |
| UA (mg/dL) | 1.75 ± 0.11 b | 1.01 ± 0.09 a | 0.90 ± 0.07 a | 0.79 ± 0.04 a | <0.0001 |
| CK (U/L) | 259 ± 24 d | 173 ± 12 c | 126 ± 10 a,b | 101 ± 7 a | <0.0001 |
| TC (mg/dL) | 111 ± 3 | 103 ± 4 | 104 ± 5 | 102 ± 5 | 0.1701 |
| TG (mg/dL) | 152 ± 5 b | 147 ± 4 b,c | 138 ± 6 a,c | 134 ± 3 a | 0.0014 |
| Glucose (mg/dL) | 154 ± 3 | 153 ± 4 | 158 ± 2 | 158 ± 3 | 0.5448 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsu, Y.-J.; Huang, W.-C.; Chiu, C.-C.; Liu, Y.-L.; Chiu, W.-C.; Chiu, C.-H.; Chiu, Y.-S.; Huang, C.-C. Capsaicin Supplementation Reduces Physical Fatigue and Improves Exercise Performance in Mice. Nutrients 2016, 8, 648. https://doi.org/10.3390/nu8100648
Hsu Y-J, Huang W-C, Chiu C-C, Liu Y-L, Chiu W-C, Chiu C-H, Chiu Y-S, Huang C-C. Capsaicin Supplementation Reduces Physical Fatigue and Improves Exercise Performance in Mice. Nutrients. 2016; 8(10):648. https://doi.org/10.3390/nu8100648
Chicago/Turabian StyleHsu, Yi-Ju, Wen-Ching Huang, Chien-Chao Chiu, Yan-Lin Liu, Wan-Chun Chiu, Chun-Hui Chiu, Yen-Shuo Chiu, and Chi-Chang Huang. 2016. "Capsaicin Supplementation Reduces Physical Fatigue and Improves Exercise Performance in Mice" Nutrients 8, no. 10: 648. https://doi.org/10.3390/nu8100648
APA StyleHsu, Y.-J., Huang, W.-C., Chiu, C.-C., Liu, Y.-L., Chiu, W.-C., Chiu, C.-H., Chiu, Y.-S., & Huang, C.-C. (2016). Capsaicin Supplementation Reduces Physical Fatigue and Improves Exercise Performance in Mice. Nutrients, 8(10), 648. https://doi.org/10.3390/nu8100648