Role of Chlorogenic Acids in Controlling Oxidative and Inflammatory Stress Conditions

Abstract
:1. Introduction
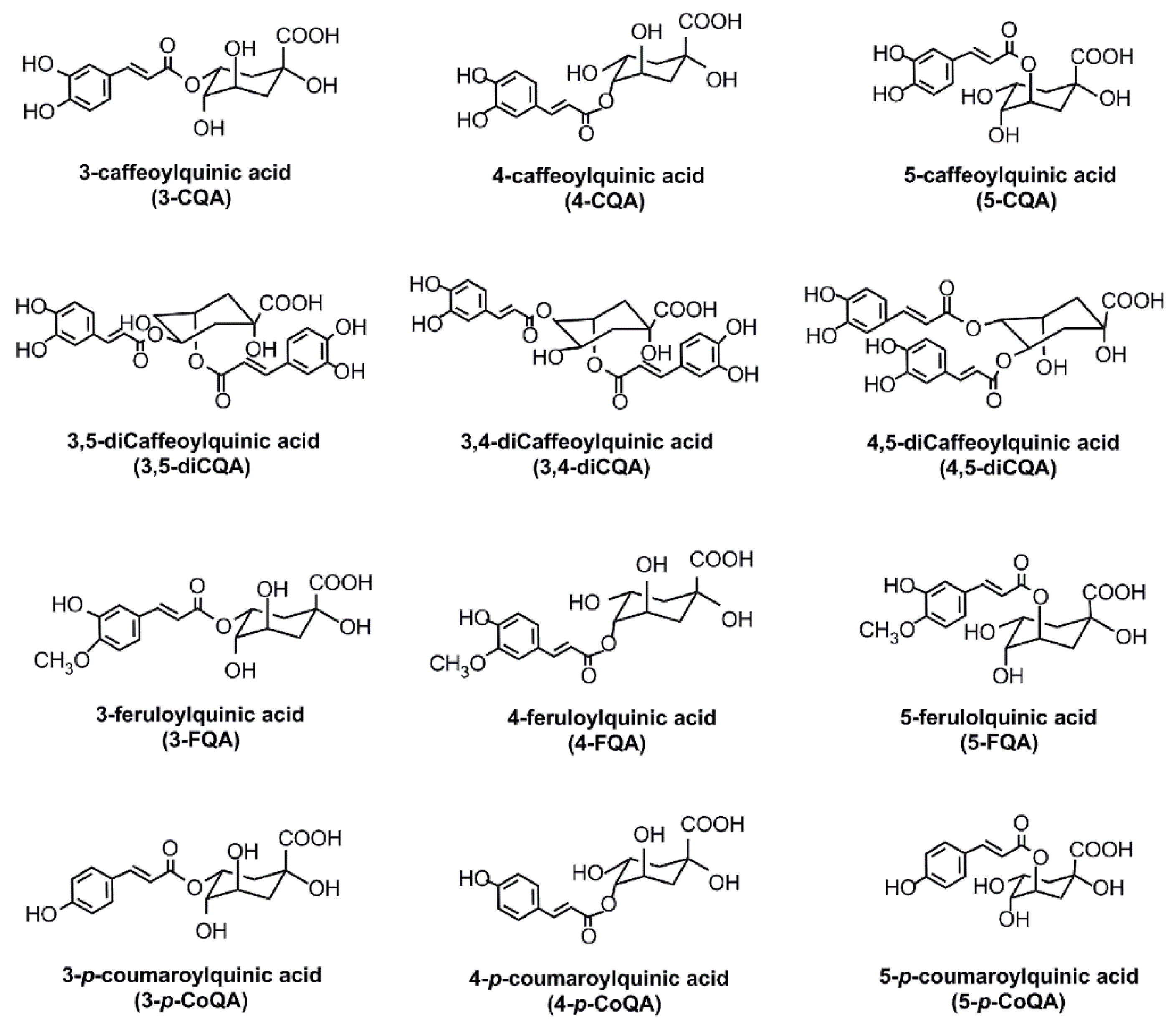
2. CGAs Content in Coffee Beans and Coffee Brew
3. Chlorogenic Acid Content in Other Plant Sources
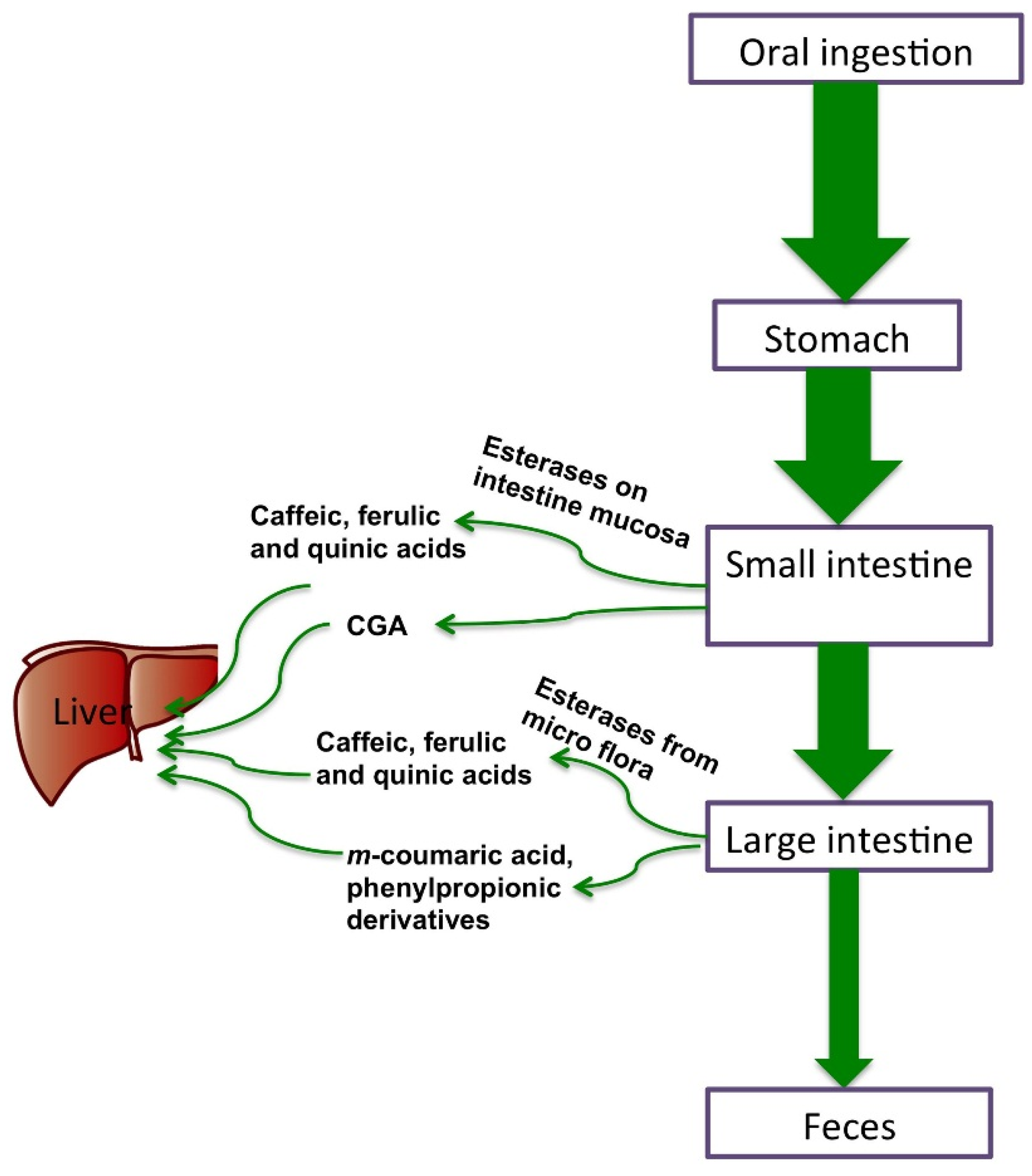
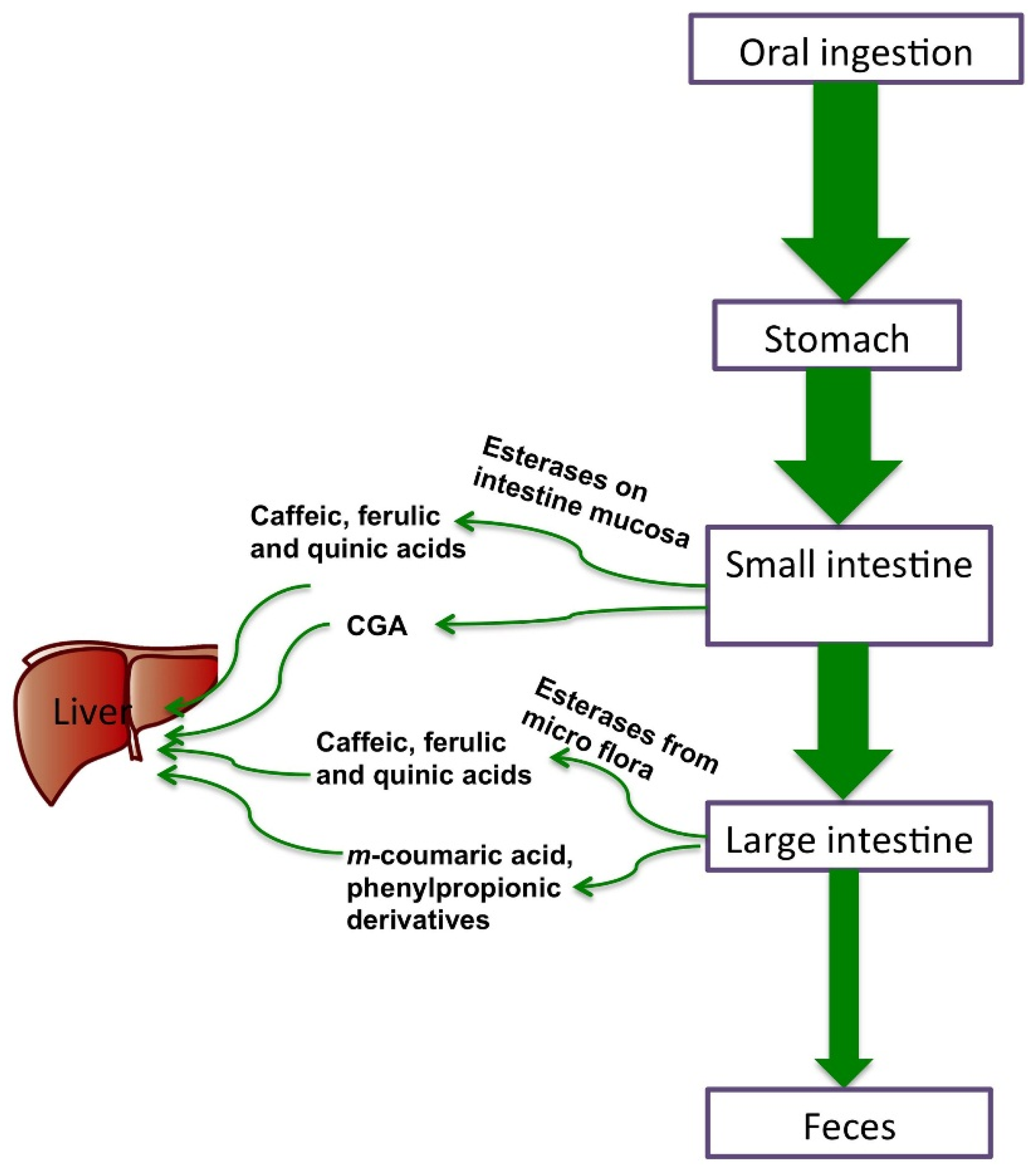
4. Bioavailability and Metabolism of CGAs
5. Antioxidant Activity of CGA Isomers
5.1. Overview of Antioxidant and Prooxidant Mechanisms and Assays for Assessing Antioxidant Activity in Vitro and in Vivo
5.2. Evidence from Chemical-Based Assays
5.3. Evidence from Cell-Based Assays

{kind=link}
{kind=link}
| Chemical Assays | End-Point Measure | CGA Isomer | Concentration/Exposure Time | Results | References |
|---|---|---|---|---|---|
| DPPH assay | DPPH | 5-CQA | 5–80 μM for 3 h | 10%~90% inhibition on DPPH | [51] |
| Xanthine/xanthine oxidase system | DMPO/·OOH adducts | 5-CQA | 20 μM for 2.5 min | ↓ 30% ·OOH | [51] |
| FeSO4 + H2O2 | DMPO/·OH adducts | 5-CQA | 20 μM for 2.5 min | ↓ 51% ·OH | [51] |
| FeSO4 + H2O2 | DMPO/·OH adducts | 5-CQA | 100–400 μM for 1 min | ↓ 50% to 80% ·OH | [52] |
| ABTS assay | ABTS·+ | 5-CQA | Serials concentration for 15 min | The ability of 100 g of CGA in scavenging ABTS·+ is equivalent to 3.7 mmol Trolox | [53] |
| Rat brain homogenates + sodium nitroprusside | MDA | 5-CQA | 1.56–6.25 μg/mL | No significant inhibition of MDA | [53] |
| Liposome system containing AAPH | MDA | 5-CQA | 0.1–0.5 mM | Second order rate of constant of the reactions of LOO· with CGA is 1.28 ± 0.11 × 105 M−1·s−1 | [54] |
| Pulse radiolysis to generate O2- | O2- | 5-CQA | 0.2–0.75 mM | Second order rate of constant of the reactions of O2- with CGA is 0.96 ± 0.01 × 106 M−1·s−1 | [54] |
| Fenton-type reaction to generate ·OH | ·OH | 5-CQA | 0.1–0.75 mM | Second order rate of constant of the reactions of ·OH with CGA is 3.34 ± 0.19 × 109 M−1·s−1 | [54] |
| Potassium phosphate to generate ONOO- | ONOO- | 5-CQA | 80 μM | Second order rate of constant of the reactions of ONOO- with CGA is 1.6 ± 0.7 × 105 M−1·s−1 | [54] |
| DPPH assay | DPPH | 3-CQA, | 5 μg/mL–60 μg/mL | EC50 a 3-CQA:13.4 μg/mL | [55] |
| 4-CQA, | 4-CQA: 13.2 μg/mL | ||||
| 5-CQA, | 5-CQA: 13.8 μg/mL | ||||
| 3,5-diCQA, | 3,5-diCQA: 9.3 μg/mL | ||||
| 3,4-diCQA, | 3,4-diCQA: 9.4 μg/mL | ||||
| 4,5-diCQA | 4,5-diCQA: 7.5 μg/mL | ||||
| ABTS assay | ABTS·+ | 3-CQA | 50 μg/mL–150 μg/mL | EC50 a 3-CQA: 91.4 μg/mL | [55] |
| 4-CQA, | 4-CQA: 87.5 μg/mL | ||||
| 5-CQA, | 5-CQA: 91.5 μg/mL | ||||
| 3,5-diCQA, | 3,5-diCQA: 77.6 μg/mL | ||||
| 3,4-diCQA, | 3,4-diCQA: 77.4 μg/mL | ||||
| 4,5-diCQA | 4,5-diCQA: 67.3 μg/mL | ||||
| FRAP assay | Reducing power | 3-CQA | 25–125 μg/mL | 4,5-diCQA > 3,5-diCQA > 3,4-diCQA > 5-CQA = 4-CQA = 3-CQA | [55] |
| 4-CQA, | |||||
| 5-CQA, | |||||
| 3,5-diCQA, | |||||
| 3,4-diCQA, | |||||
| 4,5-diCQA | |||||
| DNA damage protective effect assay | DNA damage | 3-CQA | 50 μg/mL | ↓ 43.1 to 62.4% DNA damage | [55] |
| 4-CQA, | |||||
| 5-CQA, | |||||
| 3,5-diCQA, | 4,5-diCQA > 3,4-diCQA > 3,5-diCQA > 5-CQA > 4-CQA > 3-CQA | ||||
| 3,4-diCQA, | |||||
| 4,5-diCQA | |||||
| Plasmid pUC18 + NH2Cl | Supercoiled DNA, nicked circular DNA and linear duple | 5-CQA | 0.01 mM–1.23 mM | Prevented a stepwise conversion of plasmid DNA form supercoiled DNA, nicked circular DNA and linear duplex DNA | [50] |
| LDL + copper | Conjugated dienes | 5-CQA | 0.25–1.0 μM | ↑ lag time of LDL oxidation | [56] |
| LDL + metmyoglobin + H2O2 | ROS | 5-CQA | 1 molar ratio to metmyoglobin | Effectively blocked LDL oxidation | [57] |
| Model | End-Point Measure | CGA Isomer | Concentration/Exposure Time | Results Compared to the Control without CGA Treatment | References | |
|---|---|---|---|---|---|---|
| Cell-based assay | ||||||
| HaCaT cell + H2O2 | H2O2 | 5-CQA | 20 μM for 20 h | ↓ ROS | [51] | |
| HaCaT cell + UVB | ROS, DNA damage, cell viability | 5-CQA | 20 μM for 20 h | ↓ ROS | [51] | |
| ↓ DNA damage | ||||||
| ↑ 13% cell viability | ||||||
| Mesenchymal stem cell + H2O2 | Chromosomal condensation, cell apoptosis, ROS | 5-CQA | 10 mM for 12 h | ↓ Chromosomal condensation | [58] | |
| ↓ Cell apoptosis | ||||||
| ↓ ROS | ||||||
| Primary cortical neurons + H2O2 | NADPH: quinine oxido-reductase 1, Cell viability | 5-CQA | 12.5–100 μM for 1 h | ↑ NADPH: quinine oxido-reductase 1 | [59] | |
| ↑ Cell viability | ||||||
| Differentiated neuronal PC12 cells + H2O2 | Cell viability, GSH | 5-CQA | 6.2–25 μM for 2 h | ↑ Cell viability | [60] | |
| Attenuated GSH decrease | ||||||
| Differentiated neuronal PC12 cells + FeSO4 | Cell viability, ROS, MDA | 5-CQA | 6.2–25 μM for 2 h | Did not change cell viability | [60] | |
| ↓ ROS level | ||||||
| ↓ MDA | ||||||
| Differentiated neuronal PC12 cell + t-BHP | Cell viability, GSH | 5-CQA | 6.2–25 μM for 2 h | ↑ Cell viability | [60] | |
| Did not change GSH level | ||||||
| Human breast cancer cell line MCF-7+ | Global methylation status | 5-CQA | 1–20 μM for 8 days | ↓ Global methylation | [61] | |
| Human breast cancer cell line MDA-MB-231+ | Global methylation status | 5-CQA | 0.2–20 μM for 3 days | Did not change the global methylation status | [61] | |
| Human breast cancer cell line T-47D+ | Global methylation status | 5-CQA | 20–50 μM for 2 days | ↓ Global methylation | [61] | |
| Human lymphocyte + X-ray radiation | Genetic damage index | 5-CQA | 0.5–4 μg/mL | ↓ Genetic damge index by 4.49% to 48.15% | [62] | |
| Mouse epidermal cell line JB6 + UVB | GST, NADPH: quinone oxido-reductase, Nrf2 | 5-CQA | 5–160 μM for 1 h | ↑ GST | [64] | |
| ↑ NADPH:quinone oxido-reductase | ||||||
| ↑ Nrf2 nuclear translocation | ||||||
| Differentiated neuronal PC12 cell + H2O2 | Mitochondrial membrane depolarization | 3-CQA, 4-CQA, 5-CQA | 10 μM for 20 min | Protected mitochondrial membrane depolarization through | [65] | |
| ↓ Caspase 9 activation | ||||||
| Human hepatoma HepG2 cell + t-BOOH | ROS, GSH, GPx, GR, MDA | 5-CQA, 3,5-diCQA | 10–20 μM for 20 h | ↓ ROS, ↑ GSH, ↑ GR, ↓ GPx, ↓ MDA | [66] | |
| Animal Models | ||||||
| Type 2 diabetic rat model | Lipid peroxidation, GSH, VC, VE | 5-CQA | Oral administration at 5 mg/kg body weight daily for 45 days | ↓ Plasma lipid hydroperoxides, ↑ GSH, ↑ VC, ↑ VE | [67] | |
| Type 2 diabetic rat model | Lipid peroxidation, GST, SOD, GPx, CAT | 5-CQA | Oral administration at 5 mg/kg body weight daily for 45 days | ↓ Lipid oxidation, ↑ GST, ↑ SOD, ↑ GPx, ↑ CAT | [68] | |
| Methamphetamine induced oxidative stress rat model | NO, MDA, SOD, GPx | 5-CQA | Oral administration at 60 mg/kg body weight, single dose | ↓ NO, ↓ MDA, ↑ SOD, ↑ GPx | [69] | |
| Cd induced brain impairment rat model | SOD, CAT, GPx, GSH, VC, VE, MDA | 5-CQA | Intragastric administration, 60 mg/kg body weight daily for 30 days | ↑ SOD, ↑ CAT, ↑ GPx, ↑ GSH, ↑VC, ↑ VE, ↓ MDA | [70] | |
| Scopolamine induced brain impairment rat model | MDA | 5-CQA | Orally administered at 3–9 mg/kg body weight, single dose | ↓ MDA | [71] | |
| Benzopyrene induced gastrointestinal pathogenesis rat model | GST, Cytochrome P-450 | 5-CQA | Eating 0.2% 5-CQA containing diet for 10 weeks | ↑ GST, | [72] | |
| Did not significantly change cytochrome P-450 | ||||||
| Sodium pentobarbital induced intestinal ischemia-reperfusion rat model | Vascular permeability in the small intestine | 5-CQA | Directly administrate 1 mM into jejunum, single dose | Attenuated the increased vascular permeability | [73] | |
| Azoxymethane induced colon cancer mice model | GSH/GSSG ratio | 5-CQA | Orally administered at 0.1% 5-CQA containing diet for 20 weeks | ↑ Hepatic GSH/GSSG ratio | [44] | |
| UV irradiation induced erythema formation in Guinea pig and Yucatan micropig | Erythema | 5-CQA | Intradermal delivery of 5-CQA at 1.49 μmol/g skin | Prevented erythema formation induced by UV irradiation | [74] | |
| Gamma irradiation induced chromosomal damage in mice model | Frequencies of micro-nucleated polychromatic erythrocytes | 5-CQA | Orally administered at 50–200 mg/kg body weight, single dose | ↓ Frequencies of micro-nucleated polychromatic erythrocytes | [75] | |
5.4. Evidence from Animal-Based Assays
6. The Ability of CGA on Modulating Inflammatory Responses
6.1. Overview of Inflammation and Anti-Inflammatory Mechanisms
6.2. CGA Suppression of Inflammation through Inhibition of Pro-Inflammatory Cytokines via Modulation of Key Transcription Factors
| Models | End-Point Measure | CGA Isomer | Concentration/Exposure Time | Results Compared to the Control without CGA Treatment | References |
|---|---|---|---|---|---|
| Cell Models | |||||
| Caco-2 + TNF-α and H2O2 | IL-8 | 5-CQA | 0.5–2 mM | ↓ IL-8 | [86] |
| Caco-2 + cocktail of inflammatory mediators | IL-6, IL-8 | Mixture of all CGAs | Unknown composition of CGAs for 1 h | ↓ IL-6, ↓ IL-8 | [30] |
| RAW 264.7 + LPS | NO, IL-1β, TNF-α, cyclooxygenase-2, NF-κB, IL-6 | 5-CQA | 2–20 μM for 24 h | ↓ NO, ↓ IL-1β, ↓ TNF-α, ↓ IL-6, ↓ cyclooxygenase-2, ↓ NFκB | [87] |
| RAW 264.7 + LPS | Cyclooxygenase, Prostaglandin E2, NF-κB | 5-CQA | 12.5–37.4 μg/mL for 2 h | ↓ Cyclooxygenase, ↓ Prostaglandin E2, ↓ NF-κB | [89] |
| Animal Models | |||||
| Trinitrobenzenesulfonic acid induced colitis mice model | Myeloperoxidase, H2O2, NF-κB | 5-CQA | Orally administration at 20 mg/kg body weight, twice a day for 6 days | ↓ Myeloperoxidase, ↓ H2O2, ↓ NF-κB | [48] |
| Dextran sulfate sodium induced colitis mice model | IL-1β, TNF-α, macrophage inflammatory protein 2 | 5-CQA | Orally administration at 1 mM for 15 days | ↓ IL-1β, did not significantly change the levels of TNF-α and macrophage inflammatory protein 2 | [86] |
| Trinitrobenzenesulfonic acid induced colitis mice model | Myeloperoxidase, H2O2, NF-κB | 5-CQA | Orally administration at 20 mg/kg body weight twice a day | ↓ Myeloperoxidase, ↓ H2O2, ↓ NF-κB | [90] |
| Rheumatoid Arthritis rat model | IL-1β, TNF-α, T cells count, Th1 cytokines, Th2 cytokines | 5-CQA | Orally administration at 40 mg/kg body weight | ↓ IL-1β, ↓ TNF-α, ↓ T cells count, ↓ Th1 cytokines, ↑ Th2 cytokines | [91] |
| Wounds in diabetic rat | Wound healing speed, NO, MDA, GSH | 5-CQA | Intraperitoneal injection at 50 mg/kg/day for 15 days | ↑ Wound healing speed, ↑ GSH, ↓ NO, ↓ MDA | [92] |
| Liver injury rat model | MDA, GSH, TNF-α, NO, cyclooxygenase-2 protein increase | 5-CQA | Orally administration at 2.5–10 mg/kg body weight, twice a day | ↓ MDA, ↑ GSH, ↓ TNF-α, ↓ NO, ↓ cyclooxygenase-2 protein increase | [93] |
7. Summary and Conclusions
Acknowledgments
Conflicts of Interest
References
- Matei, M.F.; Jaiswal, R.; Kuhnert, N. Investigating the Chemical Changes of Chlorogenic Acids during Coffee Brewing: Conjugate Addition of Water to the Olefinic Moiety of Chlorogenic Acids and Their Quinides. J. Agric. Food Chem. 2012, 60, 12105–12115. [Google Scholar] [CrossRef] [PubMed]
- Farah, A.; de Paulis, T.; Trugo, L.C.; Martin, P.R. Effect of roasting on the formation of chlorogenic acid lactones in coffee. J. Agric. Food Chem. 2005, 53, 1505–1513. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N. Chlorogenic acids and other cinnamates—Nature, occurrence and dietary burden. J. Sci. Food Agric. 1999, 79, 362–372. [Google Scholar] [CrossRef]
- Perrone, D.; Donangelo, R.; Donangelo, C.M.; Farah, A. Modeling Weight Loss and Chlorogenic Acids Content in Coffee during Roasting. J. Agric. Food Chem. 2010, 58, 12238–12243. [Google Scholar] [CrossRef] [PubMed]
- Farah, A.; Donangelo, C.M. Phenolic compounds in coffee. Braz. J. Plant Physiol. 2006, 18. [Google Scholar] [CrossRef]
- Clifford, M.N.; Wight, J. The measurement of feruloylquinic acids and caffeoylquinic acids in coffee beans. Development of the technique and its preliminary application to green coffee beans. J. Sci. Food Agric. 1976, 27, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N.; Ramirez-Martinez, J.R. Phenols and caffeine in wet-processed coffee beans and coffee pulp. Food Chem. 1991, 40, 35–42. [Google Scholar] [CrossRef]
- Perrone, D.; Farah, A.; Donangelo, C.M.; de Paulis, T.; Martin, P.R. Comprehensive analysis of major and minor chlorogenic acids and lactones in economically relevant Brazilian coffee cultivars. Food Chem. 2008, 106, 859–867. [Google Scholar] [CrossRef]
- Farah, A.; Monteiro, M.; Donangelo, C.M.; Lafay, S. Chlorogenic Acids from Green Coffee Extract are Highly Bioavailable in Humans. J. Nutr. 2008, 138, 2309–2315. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, R.; Matei, M.F.; Golon, A.; Witt, M.; Kuhnert, N. Understanding the fate of chlorogenic acids in coffee roasting using mass spectrometry based targeted and non-targeted analytical strategies. Food Funct. 2012, 3, 976–984. [Google Scholar] [CrossRef] [PubMed]
- Schols, H.A.; Boekel, V.T.; Smit, G.; Bekedam, E.K. Incorporation of Chlorogenic Acids in Coffee Brew Melanoidins. J. Agric. Food Chem. 2008, 56, 2055–2063. [Google Scholar]
- Schols, H.A.; Boekel, V.T.; Smit, G.; Bekedam, E.K. High Molecular Weight Melanoidins from Coffee Brew. J. Agric. Food Chem. 2006, 54, 7658–7666. [Google Scholar]
- Adams, A.; Borrelli, R.C.; Fogliano, V.; de Kimpe, N. Thermal degradation studies of food melanoidins. J. Agric. Food Chem. 2005, 53, 4136–4142. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.K.; Yoo, H.S.; Shibamoto, T. Role of Roasting Conditions in the Level of Chlorogenic Acid Content in Coffee Beans: Correlation with Coffee Acidity. J. Agric. Food Chem. 2009, 57, 5365–5369. [Google Scholar] [CrossRef] [PubMed]
- Tfouni, S.A.V.; Carreiro, L.B.; Teles, C.R.A.; Furlani, R.P.Z.; Cipolli, K.M.V.A.B.; Camargo, M.C.R. Caffeine and chlorogenic acids intake from coffee brew: Influence of roasting degree and brewing procedure. Int. J. Food Sci. Technol. 2014, 49, 747–752. [Google Scholar] [CrossRef]
- Ludwig, L.A.; Mena, P.; Calani, L.; Cid, C.; del Rio, D.; Lean, M.E.J.; Crozier, A. Variations in caffeine and chlorogenic acid contents of coffees: What are we drinking? Food Funct. 2014, 5, 1718–1726. [Google Scholar] [CrossRef] [PubMed]
- Crozier, T.W.M.; Stalmach, A.; Lean, M.E.J.; Crozier, A. Espresso coffees, caffeine and chlorogenic acid intake: Potential health implications. Food Funct. 2012, 3, 3–33. [Google Scholar] [CrossRef] [PubMed]
- Mills, C.E.; Oruna-Concha, M.J.; Mottram, D.S.; Gibson, G.R.; Spencer, J.P.E. The effect of processing on chlorogenic acid content of commercially available coffee. Food Chem. 2013, 141, 3335–3340. [Google Scholar] [CrossRef] [PubMed]
- Fujioka, K.; Shibamoto, T. Chlorogenic acid and caffeine contents in various commercial brewed coffees. Food Chem. 2008, 106, 217–221. [Google Scholar] [CrossRef]
- Ludwig, I.A.; Sanchez, L.; Caemmerer, B.; Kroh, L.W.; de Peña, M.P.; Cid, C. Extraction of coffee antioxidants: Impact of brewing time and method. Food Res. Int. 2012, 48, 57–64. [Google Scholar] [CrossRef]
- Dao, L.; Friedman, M. Chlorogenic acid content of fresh and processed potatoes determined by ultraviolet spectrophotometry. J. Agric. Food Chem. 1992, 40, 2152–2156. [Google Scholar] [CrossRef]
- Malmberg, A.G.; Theander, O. Determination of chlorogenic acid in potato tubers. J. Agric. Food Chem. 1985, 33, 549–551. [Google Scholar] [CrossRef]
- Griffiths, D.W.; Bain, H. Photo-induced changes in the concentrations of individual chlorogenic acid isomers in potato (Solanum tuberosum) tubers and their complexation with ferric ions. Potato Res. 1997, 40, 307–315. [Google Scholar] [CrossRef]
- Papetti, A.; Daglia, M.; Aceti, C.; Sordelli, B.; Spini, V.; Carazzone, C.; Gazzani, G. Hydroxycinnamic acid derivatives occurring in Cichorium endivia vegetables. J. Pharm. Biomed. Anal. 2008, 48, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Niggeweg, R.; Martin, C.; Michael, A.J. Engineering plants with increased levels of the antioxidant chlorogenic acid. Nat. Biotechnol. 2004, 22, 746–754. [Google Scholar] [CrossRef] [PubMed]
- Cui, T.; Nakamura, K.; Ma, L.; Li, J.Z.; Kayahara, H. Analyses of arbutin and chlorogenic acid, the major phenolic constituents in oriental pear. J. Agric. Food Chem. 2005, 53, 3882–3887. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, J.Y.; Gao, W.Y.; Wang, Y.; Wang, H.Y.; Cao, J.G.; Huang, L.Q. Chemical Composition and Anti-inflammatory and Antioxidant Activities of Eight Pear Cultivars. J. Agric. Food Chem. 2012, 60, 8738–8744. [Google Scholar] [CrossRef] [PubMed]
- Awad, M.A.; de Jager, A.; van Westing, L.M. Flavonoid and chlorogenic acid levels in apple fruit: Characterisation of variation. Sci. Horticult. 2000, 83, 249–263. [Google Scholar] [CrossRef]
- Wang, Z.; Clifford, M.N.; Sharp, P. Analysis of chlorogenic acids in beverages prepared from Chinese health foods and investigation, in vitro, of effects on glucose absorption in cultured Caco-2 cells. Food Chem. 2008, 108, 369–373. [Google Scholar] [CrossRef]
- De Magalhaes, P.M.; Dupont, I.; Hendrickx, A.; Joly, A.; Raas, T.; Dessy, S.; Sergent, T.; Schneider, Y.J. Anti-inflammatory effect and modulation of cytochrome P450 activities by Artemisia annua tea infusions in human intestinal Caco-2 cells. Food Chem. 2012, 134, 864–871. [Google Scholar] [CrossRef] [PubMed]
- Farrell, T.L.; Dew, T.P.; Poquet, L.; Hanson, P.; Williamson, G. Absorption and Metabolism of Chlorogenic Acids in Cultured Gastric Epithelial Monolayers. Drug Metab. Dispoition 2011, 39, 2338–2346. [Google Scholar] [CrossRef] [PubMed]
- Lafay, S.; Gil-Izquierdo, A.; Manach, C.; Morand, C.; Besson, C.; Scalbert, A. Chlorogenic acid is absorbed in its intact form in the stomach of rats. J. Nutr. 2006, 136, 1192–1197. [Google Scholar] [PubMed]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine; Clarendon Press: Oxford, UK, 2007. [Google Scholar]
- Scherbl, D.; Muentnich, S.; Richling, E. In vitro absorption studies of chlorogenic acids from coffee using the Ussing chamber model. Food Res. Int. 2014, 63, 456–463. [Google Scholar] [CrossRef]
- Konishi, Y.; Kobayashi, S. Transepithelial transport of chlorogenic acid, caffeic acid, and their colonic metabolites in intestinal Caco-2 cell monolayers. J. Agric. Food Chem. 2004, 52, 2518–2526. [Google Scholar] [CrossRef] [PubMed]
- Rechner, A.R.; Kuhnle, G.; Bremner, P.; Hubbard, G.P.; Moore, K.P.; Rice-Evans, C.A. The metabolic fate of dietary polyphenols in humans. Free Radic. Biol. Med. 2002, 33, 220–235. [Google Scholar] [CrossRef]
- Gonthier, M.-P.; Verny, M.-A.; Besson, C.; Rémésy, C.; Scalbert, A. Chlorogenic acid bioavailability largely depends on its metabolism by the gut microflora in rats. J. Nutr. 2003, 133, 1853–1859. [Google Scholar] [PubMed]
- Scalbert, A.; Morand, C.; Manach, C.; Rémésy, C. Absorption and metabolism of polyphenols in the gut and impact on health. Biomed. Pharmacother. 2002, 56, 276–282. [Google Scholar] [CrossRef]
- Choudhury, R.; Srai, S.K.; Debnam, E.; Rice-Evans, C.A. Urinary excretion of hydroxycinnamates and flavonoids after oral and intravenous administration. Free Radic. Biol. Med. 1999, 27, 278–286. [Google Scholar] [CrossRef]
- Katan, M.B.; Hollman, P.C.H.; Olthof, M.R. Chlorogenic acid and caffeic acid are absorbed in humans. J. Nutr. 2001, 131, 66–71. [Google Scholar]
- Renouf, M.; Marmet, C.; Giuffrida, F.; Lepage, M.; Barron, D.; Beaumont, M.; Williamson, G.; Dionisi, F. Dose-response plasma appearance of coffee chlorogenic and phenolic acids in adults. Mol. Nutr. Food Res. 2014, 58, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Tagliazucchi, D.; Helal, A.; Verzelloni, E.; Conte, A. The Type and Concentration of Milk Increase the in Vitro Bioaccessibility of Coffee Chlorogenic Acids. J. Agric. Food Chem. 2012, 60, 11056–11064. [Google Scholar] [CrossRef] [PubMed]
- Stalmach, A.; Williamson, G.; Crozier, A. Impact of dose on the bioavailability of coffee chlorogenic acids in humans. Food Funct. 2014, 5, 1727–1737. [Google Scholar] [CrossRef] [PubMed]
- Davis, S.; Bruno, R.; Rosenberg, D.; Liang, H.-Y.; Park, H. Chlorogenic Acid Differentially Alters Hepatic and Small Intestinal Thiol Redox Status Without Protecting Against Azoxymethane-Induced Colon Carcinogenesis in Mice. Nutr. Cancer 2010, 62, 362–370. [Google Scholar]
- Jena, N.R. DNA damage by reactive species: Mechanisms, mutation and repair. J. Biosci. 2012, 37, 503–517. [Google Scholar] [CrossRef] [PubMed]
- Sakihama, Y.; Cohen, M.F.; Grace, S.C.; Yamasaki, H. Plant phenolic antioxidant and prooxidant activities: Phenolics-induced oxidative damage mediated by metals in plants. Toxicology. 2002, 177, 67–80. [Google Scholar] [CrossRef]
- Sakihama, Y.; Yamasaki, H. Lipid Peroxidation Induced by Phenolics in Conjunction with Aluminum Ions. Biol. Plant. 2002, 45, 249–254. [Google Scholar] [CrossRef]
- Zatorski, H.; Sałaga, M.; Zielińska, M.; Piechota-Polańczyk, A.; Owczarek, K.; Kordek, R.; Lewandowska, U.; Chen, C.; Fichna, J. Experimental colitis in mice is attenuated by topical administration of chlorogenic acid. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2015, 388, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; DesMarais, T.L.; Tong, Z.; Yao, Y.; Costa, M. Oxidative stress alters global histone modification and DNA methylation. Free Radic. Biol. Med. 2015, 82, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Shibata, H.; Sakamoto, Y.; Oka, M.; Kono, Y. Natural antioxidant, chlorogenic acid, protects against DNA breakage caused by monochloramine. Biosci. Biotechnol. Biochem. 1999, 63, 1295–1297. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.W.; Piao, M.J.; Kim, K.C.; Yao, C.W.; Zheng, J.; Kim, S.M.; Hyun, C.L.; Ahn, Y.S.; Hyun, J.W. The Polyphenol Chlorogenic Acid Attenuates UVB-mediated Oxidative Stress in Human HaCaT Keratinocytes. Biomol. Ther. 2014, 22, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.-Y.; Cosma, G.; Gardner, H.; Vallyathan, V.; Castranova, V. Effect of chlorogenic acid on hydroxyl radical. Mol. Cell. Biochem. 2003, 247, 205–210. [Google Scholar] [CrossRef]
- Oboh, G.; Agunloye, O.M.; Akinyemi, A.J.; Ademiluyi, A.O.; Adefegha, S.A. Comparative Study on the Inhibitory Effect of Caffeic and Chlorogenic Acids on Key Enzymes Linked to Alzheimer’s Disease and Some Pro-oxidant Induced Oxidative Stress in Rats’ Brain-in Vitro. Neurochem. Res. 2013, 38, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Kono, Y.; Kobayashi, K.; Tagawa, S.; Adachi, K.; Ueda, A.; Sawa, Y.; Shibata, H. Antioxidant activity of polyphenolics in diets. BBA Gen. Subj. 1997, 1335, 335–342. [Google Scholar] [CrossRef]
- Xu, J.G.; Hu, Q.P.; Liu, Y. Antioxidant and DNA-Protective Activities of Chlorogenic Acid Isomers. J. Agric. Food Chem. 2012, 60, 11625–11630. [Google Scholar] [CrossRef] [PubMed]
- Gordon, M.H.; Wishart, K. Effects of Chlorogenic Acid and Bovine Serum Albumin on the Oxidative Stability of Low Density Lipoproteins in Vitro. J. Agric. Food Chem. 2010, 58, 5828–5833. [Google Scholar] [CrossRef] [PubMed]
- Laranjinha, J.; Vieira, O.; Almeida, L.; Madeira, V. Inhibition of metmyoglobin/H2O2-dependent low density lipoprotein lipid peroxidation by naturally occurring phenolic acids. Biochem. Pharmacol. 1996, 51, 395–402. [Google Scholar] [CrossRef]
- Li, S.Y.; Bian, H.T.; Liu, Z.; Wang, Y.; Dai, J.H.; He, W.F.; Liao, X.G.; Liu, R.R.; Luo, J. Chlorogenic acid protects MSCs against oxidative stress by altering FOXO family genes and activating intrinsic pathway. Eur. J. Pharmacol. 2012, 674, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.W.; Kim, J.; Kim, J.; Lee, K.W.; Lee, H.J.; Lee, S.; Shim, J.; Jang, Y.J.; Yang, H.; Park, J.; et al. Caffeinated coffee, decaffeinated coffee, and the phenolic phytochemical chlorogenic acid up-regulate NQO1 expression and prevent H2O2-induced apoptosis in primary cortical neurons. Neurochem. Int. 2012, 60, 466–474. [Google Scholar] [CrossRef] [PubMed]
- Pavlica, S.; Gebhardt, R. Protective effects of ellagic and chlorogenic acids against oxidative stress in PC12 cells. Free Radic. Res. 2005, 39, 1377–1390. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.J.; Zhu, B.T. Inhibition of DNA methylation by caffeic acid and chlorogenic acid, two common catechol-containing coffee polyphenols. Carcinogenesis 2006, 27, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Cinkilic, N.; Cetintas, S.K.; Zorlu, T.; Vatan, O.; Yilmaz, D.; Cavas, T.; Tunc, S.; Ozkan, L.; Bilaloglu, R. Radioprotection by two phenolic compounds: Chlorogenic and quinic acid, on X-ray induced DNA damage in human blood lymphocytes in vitro. Food Chem. Toxicol. 2013, 53, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Heck, D.E.; Vetrano, A.M.; Mariano, T.M.; Laskin, J.D. UVB light stimulates production of reactive oxygen species—Unexpected role for catalase. J. Biol. Chem. 2003, 278, 22432–22436. [Google Scholar] [CrossRef] [PubMed]
- Feng, R.T.; Lu, Y.J.; Bowman, L.L.; Qian, Y.; Castranova, V.; Ding, M. Inhibition of activator protein-1, NF-kappa B, and MAPKs and induction of phase 2 detoxifying enzyme activity by chlorogenic acid. J. Biol. Chem. 2005, 280, 27888–27895. [Google Scholar] [CrossRef] [PubMed]
- Park, J.B. Isolation and quantification of major chlorogenic acids in three major instant coffee brands and their potential effects on H2O2-induced mitochondrial membrane depolarization and apoptosis in PC-12 cells. Food Funct. 2013, 4, 1632–1638. [Google Scholar] [CrossRef] [PubMed]
- Baeza, G.; Amigo-Benavent, M.; Sarria, B.; Goya, L.; Mateos, R.; Bravo, L. Green coffee hydroxycinnamic acids but not caffeine protect human HepG2 cells against oxidative stress. Food Res. Int. 2014, 62, 1038–1046. [Google Scholar] [CrossRef]
- Karthikesan, K.; Pari, L.; Menon, V.P. Protective effect of tetrahydrocurcumin and chlorogenic acid against streptozotocin–nicotinamide generated oxidative stress induced diabetes. J. Funct. Foods 2010, 2, 134–142. [Google Scholar] [CrossRef]
- Pari, L.; Karthikesan, K.; Menon, V.P. Comparative and combined effect of chlorogenic acid and tetrahydrocurcumin on antioxidant disparities in chemical induced experimental diabetes. Mol. Cell. Biochem. 2010, 341, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Koriem, K.M.M.; Soliman, R.E. Chlorogenic and caftaric acids in liver toxicity and oxidative stress induced by methamphetamine. J. Toxicol. 2014, 2014, 583494. [Google Scholar] [CrossRef] [PubMed]
- Hao, M.-L.; Pan, N.; Zhang, Q.-H.; Wang, X.-H. Therapeutic efficacy of chlorogenic acid on cadmium-induced oxidative neuropathy in a murine model. Exp. Ther. Med. 2015, 9, 1887–1894. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.-H.; Lee, S.-Y.; Lee, C.-K.; Lee, H.-K.; Kim, H.-C.; Kim, Y.-B.; Kim, J.-A.; Hong, S.-I.; Jo, T.-H.; Park, Y.-I.; et al. Neuroprotective effects of chlorogenic acid on scopolamine-induced amnesia via anti-acetylcholinesterase and anti-oxidative activities in mice. Eur. J. Pharmacol. 2010, 649, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Kitts, D.D.; Wijewickreme, A.N. Effect of dietary caffeic and chlorogenic acids on in vivo xenobiotic enzyme-systems. Plant Foods Hum. Nutr. 1994, 45, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Itagaki, S.; Kurokawa, T.; Ogura, J.; Kobayashi, M.; Hirano, T.; Sugawara, M.; Iseki, K. In vitro and in vivo antioxidant properties of chlorogenic acid and caffeic acid. Int. J. Pharm. 2011, 403, 136–138. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, S.; Yoshii, K.; Morita, S.; Teraoka, R. Efficient Topical Delivery of Chlorogenic Acid by an Oil-in-Water Microemulsion to Protect Skin against UV-Induced Damage. Chem. Pharm. Bull. 2011, 59, 793–796. [Google Scholar] [CrossRef] [PubMed]
- Abraham, S.K.; Sarma, L.; Kesavan, P.C. Protective effects of chlorogenic acid, curcumin and β-carotene against γ-radiation-induced in vivo chromosomal damage. Mutat. Res. Lett. 1993, 303, 109–112. [Google Scholar] [CrossRef]
- Bonnefont-Rousselot, D. Glucose and reactive oxygen species. Curr. Opin. Clin. Nutr. Metab. Care 2002, 5, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Kasprzak, K.S. Oxidative DNA and protein damage in metal-induced toxicity and carcinogenesis. Free Radic. Biol. Med. 2002, 32, 958–967. [Google Scholar] [CrossRef]
- Jeong, E.J.; Lee, K.Y.; Kim, S.H.; Kim, Y.C.; Sung, S.H. Cognitive-enhancing and antioxidant activities of iridoid glycosides from Scrophularia buergeriana in scopolamine-treated mice. Eur. J. Pharmacol. 2008, 588, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R.; Janeway, C.A. Innate Immunity: The Virtues of a Nonclonal System of Recognition; Elsevier Inc.: Cambridge, UK, 1997; Volume 91, pp. 295–298. [Google Scholar]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Nathan, C. Neutrophils and immunity: Challenges and opportunities. Nat. Rev. Immunol. 2006, 6, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Elenkov, I.J.; Iezzoni, D.G.; Daly, A.; Harris, A.G.; Chrousos, G.P. Cytokine Dysregulation, Inflammation and Well-Being. Neuroimmunomodulation 2005, 12, 255–269. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1. [Google Scholar] [CrossRef] [PubMed]
- Laine, L. Approaches to Nonsteroidal Anti-inflammatory Drug Use in the High-Risk Patient. Gastroenterology 2001, 120, 594–606. [Google Scholar] [CrossRef] [PubMed]
- Neurath, M.F. Cytokines in inflammatory bowel disease. Nat. Rev. Immunol. 2014, 14, 329–342. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.S.; Satsu, H.; Bae, M.J.; Zhao, Z.H.; Ogiwara, H.; Totsuka, M.; Shimizu, M. Anti-inflammatory effect of chlorogenic acid on the IL-8 production in Caco-2 cells and the dextran sulphate sodium-induced colitis symptoms in C57BL/6 mice. Food Chem. 2015, 168, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.J.; Kim, Y.-W.; Park, Y.; Lee, H.-J.; Kim, K.-W. Anti-inflammatory effects of chlorogenic acid in lipopolysaccharide-stimulated RAW 264.7 cells. Inflamm. Res. 2014, 63, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Mitjavila, M.T.; Moreno, J.J. The effects of polyphenols on oxidative stress and the arachidonic acid cascade. Implications for the prevention/treatment of high prevalence diseases. Biochem. Pharmacol. 2012, 84, 1113–1122. [Google Scholar] [CrossRef] [PubMed]
- Shan, J.; Fu, J.; Zhao, Z.; Kong, X.; Huang, H.; Luo, L.; Yin, Z. Chlorogenic acid inhibits lipopolysaccharide-induced cyclooxygenase-2 expression in RAW264.7 cells through suppressing NF-κB and JNK/AP-1 activation. Int. Immunopharmacol. 2009, 9, 1042–1048. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, P.S.; Satti, N.K.; Sharma, V.K.; Dutt, P.; Avtar, K.; Bani, S. Amelioration of inflammatory responses by chlorogenic acid via suppression of pro-inflammatory mediators. J. Appl. Pharm. Sci. 2011, 1, 67–75. [Google Scholar]
- Chauhan, P.S.; Satti, N.K.; Sharma, P.; Sharma, V.K.; Suri, K.A.; Bani, S. Differential Effects of Chlorogenic Acid on Various Immunological Parameters Relevant to Rheumatoid Arthritis. Phytother. Res. 2012, 26, 1156–1165. [Google Scholar] [CrossRef] [PubMed]
- Bagdas, D.; Etoz, B.C.; Gul, Z.; Ziyanok, S.; Inan, S.; Turacozen, O.; Gul, N.Y.; Topal, A.; Cinkilic, N.; Tas, S.; et al. In vivo systemic chlorogenic acid therapy under diabetic conditions: Wound healing effects and cytotoxicity/genotoxicity profile. Food Chem. Toxicol. 2015, 81, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Yun, N.; Kang, J.W.; Lee, S.M. Protective effects of chlorogenic acid against ischemia/reperfusion injury in rat liver: Molecular evidence of its antioxidant and anti-inflammatory properties. J. Nutr. Biochem. 2012, 23, 1249–1255. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.T.; Dong, L.; Jiang, J.; Zhao, J.H.; Zhao, G.; Dang, X.Y.; Lu, X.L.; Jia, M. Chlorogenic acid reduces liver inflammation and fibrosis through inhibition of toll-like receptor 4 signaling pathway. Toxicology 2013, 303, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Kitts, D.D. Confirmation that the Maillard reaction is the principle contributor to the antioxidant capacity of coffee brews. Food Res. Int. 2011, 44, 2418–2424. [Google Scholar] [CrossRef]
- Du, W.Y.; Chang, C.; Zhang, Y.; Liu, Y.Y.; Sun, K.; Wang, C.S.; Wang, M.X.; Liu, Y.; Wang, F.; Fan, J.Y.; et al. High-dose chlorogenic acid induces inflammation reactions and oxidative stress injury in rats without implication of mast cell degranulation. J. Ethnopharmacol. 2013, 147, 74–83. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, N.; Kitts, D.D. Role of Chlorogenic Acids in Controlling Oxidative and Inflammatory Stress Conditions. Nutrients 2016, 8, 16. https://doi.org/10.3390/nu8010016
Liang N, Kitts DD. Role of Chlorogenic Acids in Controlling Oxidative and Inflammatory Stress Conditions. Nutrients. 2016; 8(1):16. https://doi.org/10.3390/nu8010016
Chicago/Turabian StyleLiang, Ningjian, and David D. Kitts. 2016. "Role of Chlorogenic Acids in Controlling Oxidative and Inflammatory Stress Conditions" Nutrients 8, no. 1: 16. https://doi.org/10.3390/nu8010016
APA StyleLiang, N., & Kitts, D. D. (2016). Role of Chlorogenic Acids in Controlling Oxidative and Inflammatory Stress Conditions. Nutrients, 8(1), 16. https://doi.org/10.3390/nu8010016