The Impaired Function of Macrophages Induced by Strenuous Exercise Could Not Be Ameliorated by BCAA Supplementation

Abstract
:1. Introduction
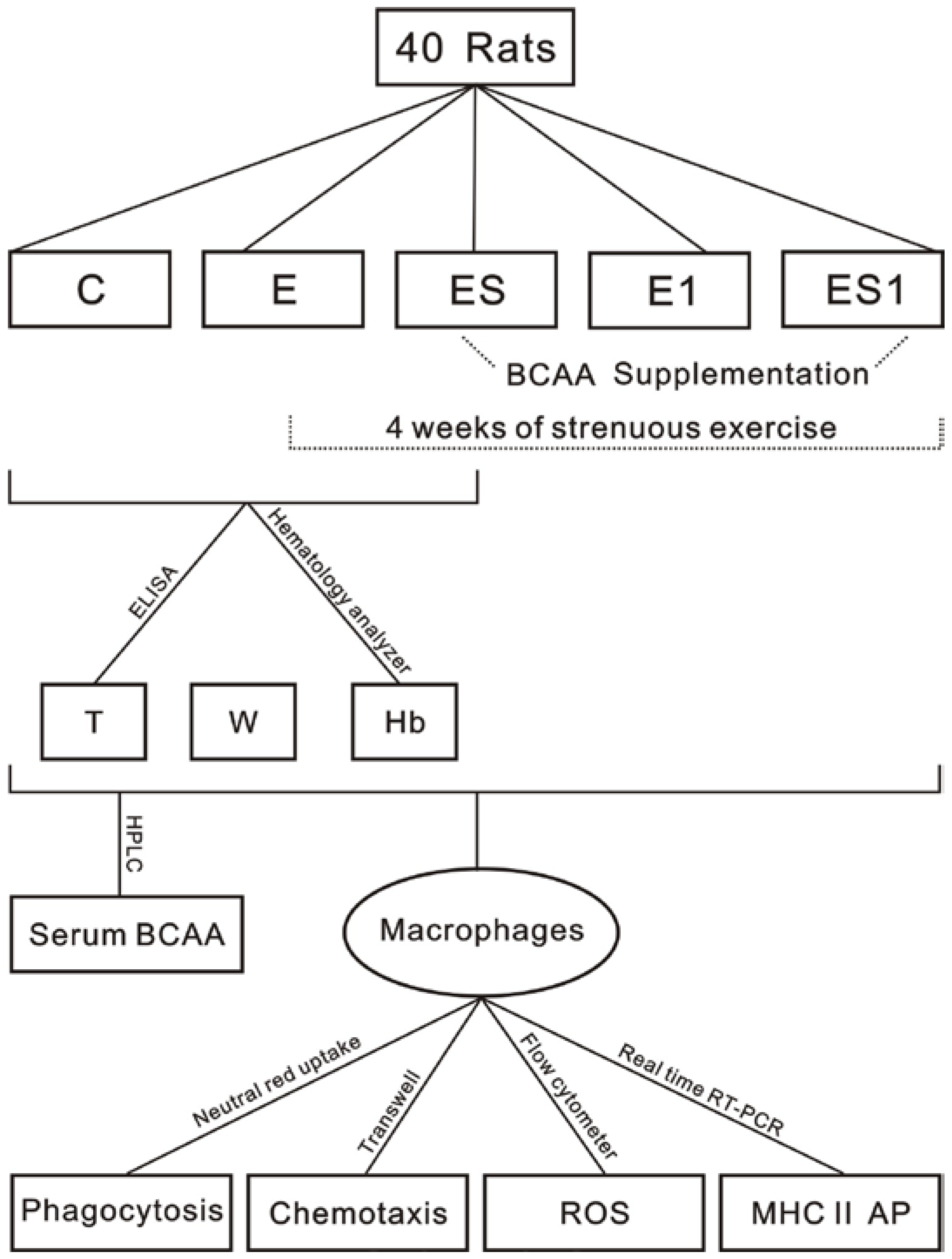
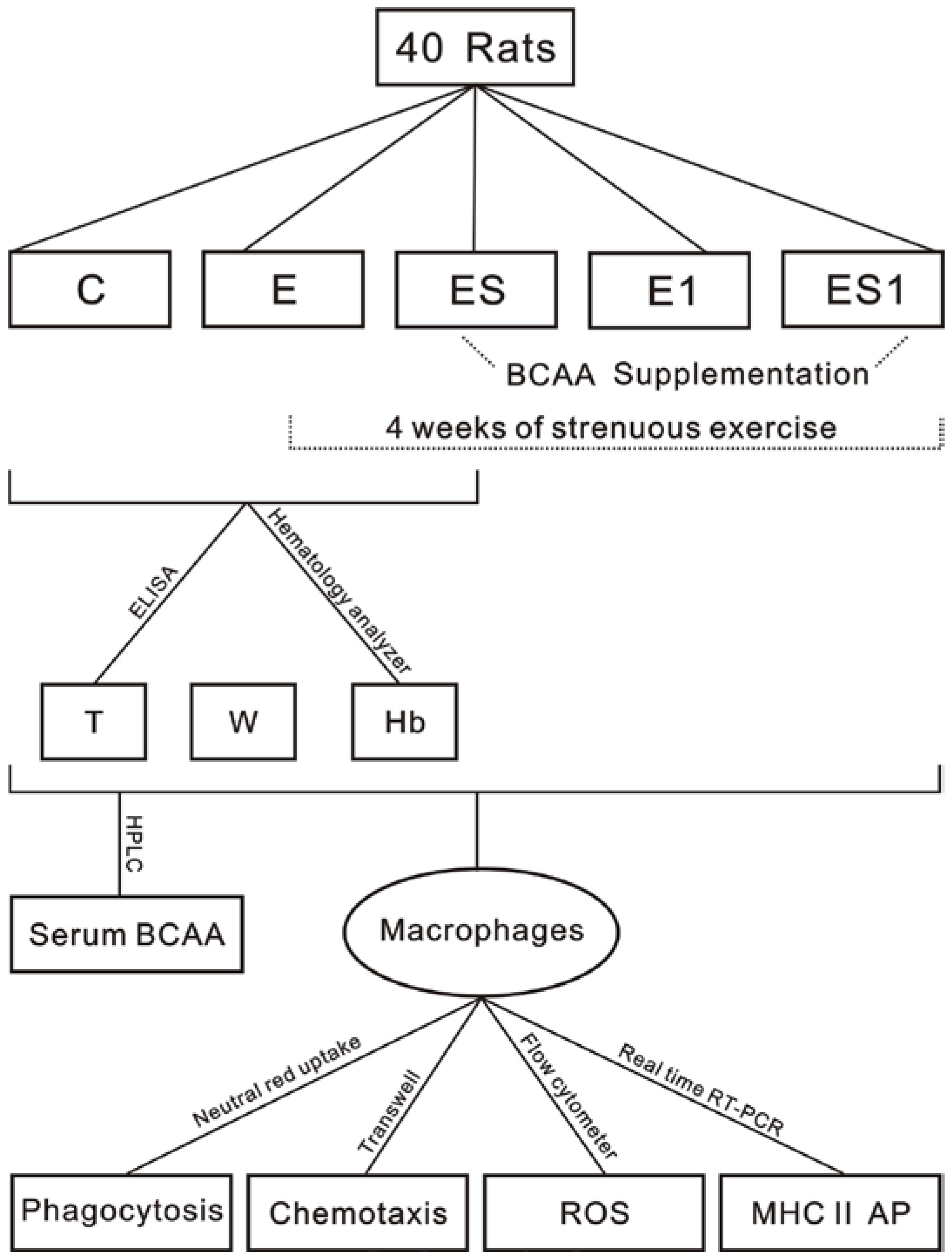
2. Materials and Methods
2.1. Animals
2.2. Exercise Program
2.3. BCAA Supplementation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | g | kcal |
|---|---|---|
| Casein | 140.000 | 560 |
| l-Cystine | 1.800 | 7.2 |
| Corn Starch | 495.692 | 1983 |
| Maltodextrin | 125 | 500 |
| Sucrose | 100.000 | 400 |
| Cellulose | 50.000 | 0 |
| Soybean Oil | 40.000 | 360 |
| t-Butylhydroquinone | 0.008 | 0 |
| Mineral mix | 35.000 | 0 |
| Vitamin mix | 10.000 | 40 |
| Choline bitartrate | 2.500 | 0 |
| Total | 1000 | 3850 |
2.4. Biochemical Analyses
2.5. Peritoneal Macrophages Preparation
2.6. Chemotaxis Assay
2.7. Phagocytosis Assay
2.8. Reactive Oxygen Species Determination
2.9. Real Time reverse-transcription polymerase chain reaction (Real Time RT-PCR)
2.10. Statistical Analysis
3. Results
3.1. Body Weight, Hemoglobin and Testosterone Levels
| Group | Body Weight (g) | Hemoglobin (g/L) | Testosterone (ng/mL) |
|---|---|---|---|
| C | 318.25 ± 11.57 | 142.88 ± 4.16 | 4.49 ± 0.55 |
| E | 274.13 ± 17.68 ** | 129.63 ± 10.56 ** | 3.08 ± 0.80 ** |
| ES | 277.75 ± 14.48 ** | 132.87 ± 9.50 * | 2.81 ± 0.73 ** |
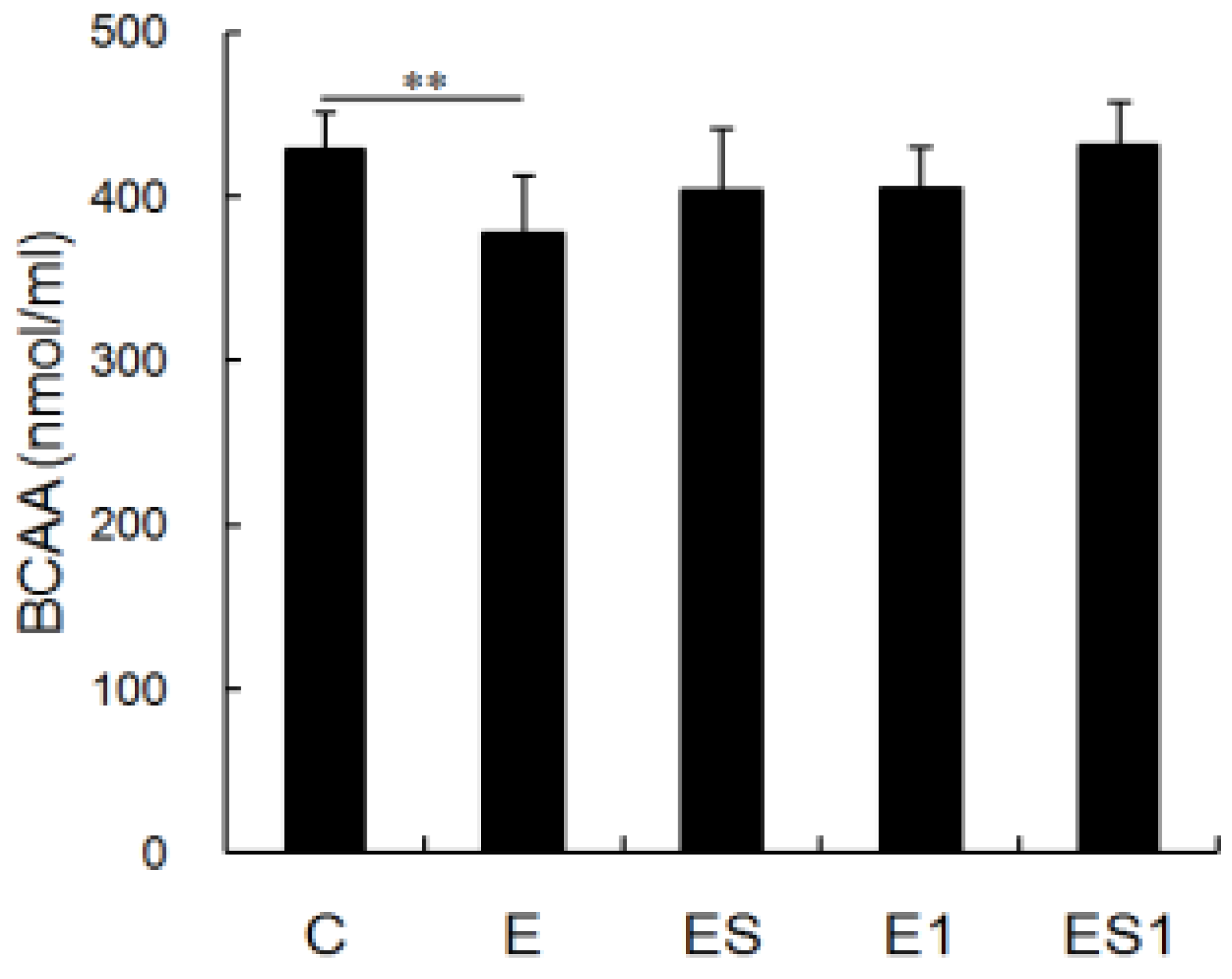
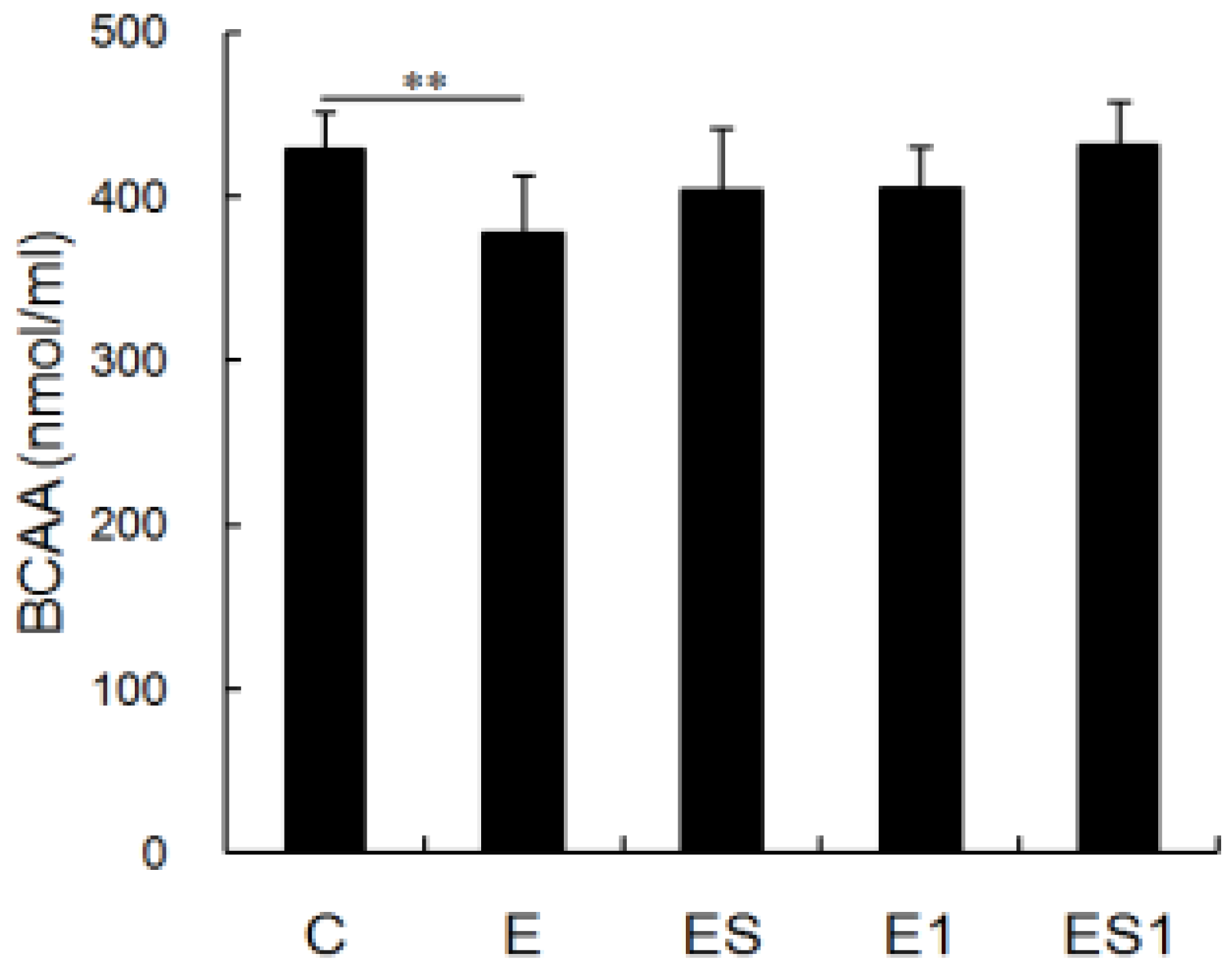
3.2. Serum BCAA Levels
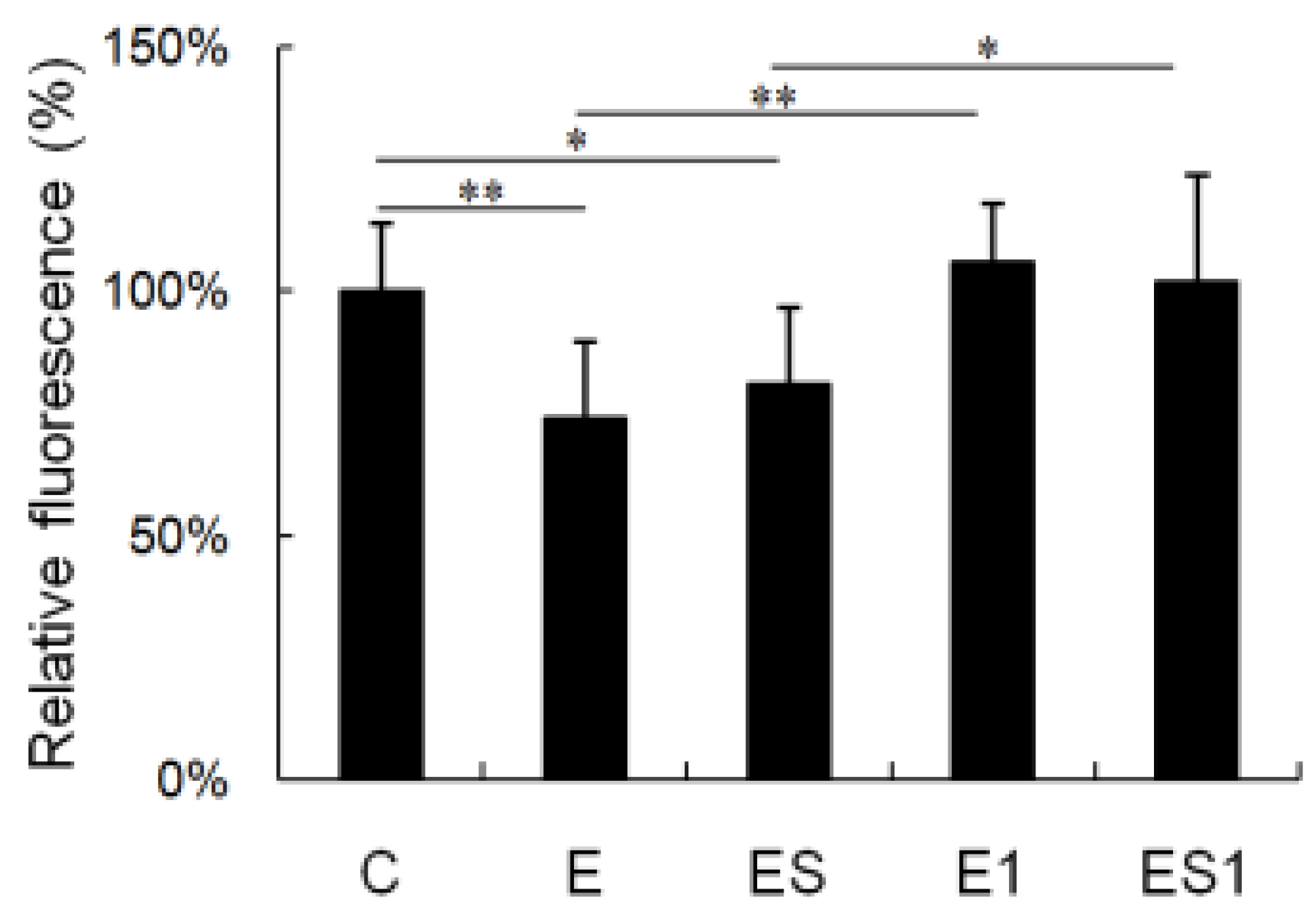
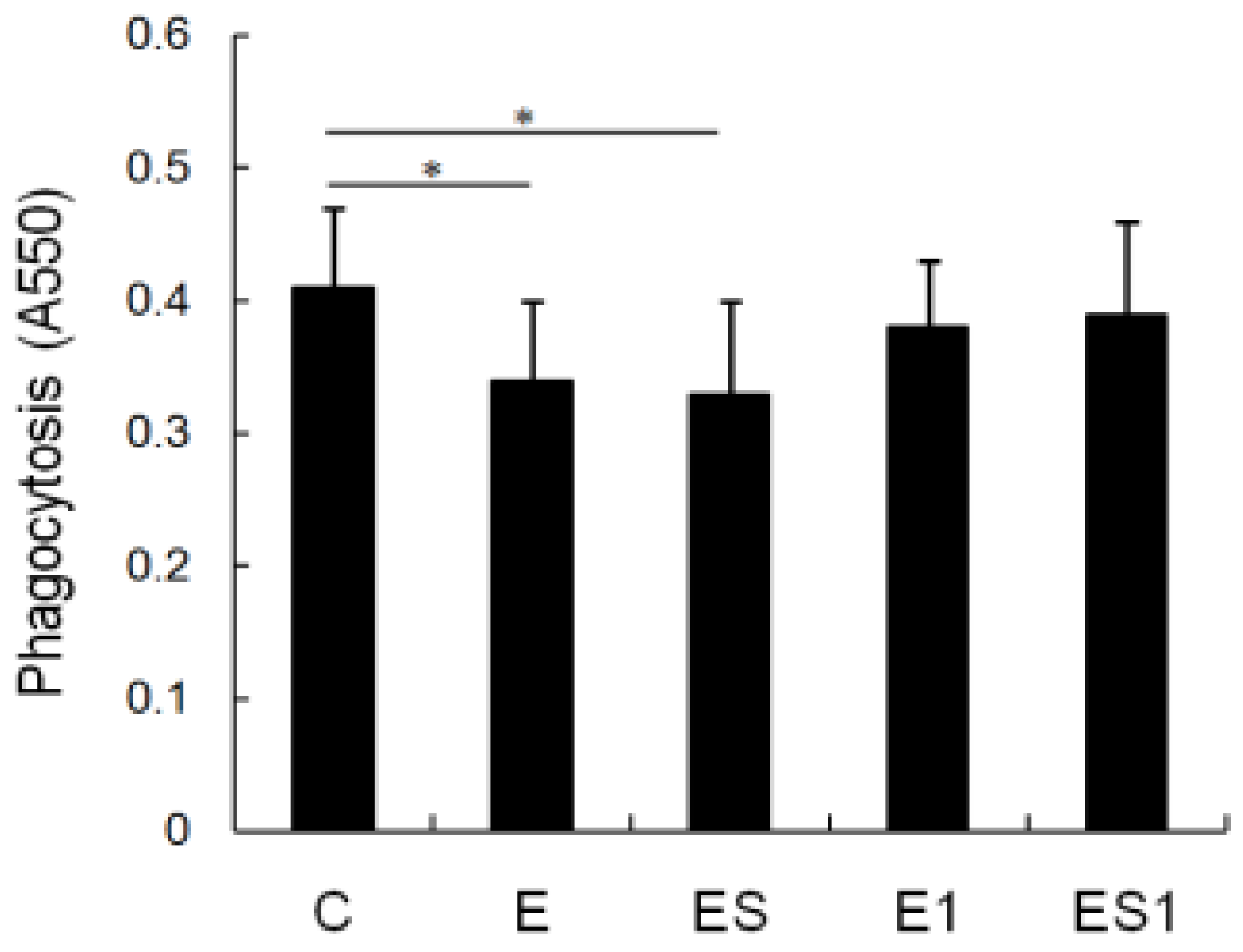
3.3. Phagocytosis
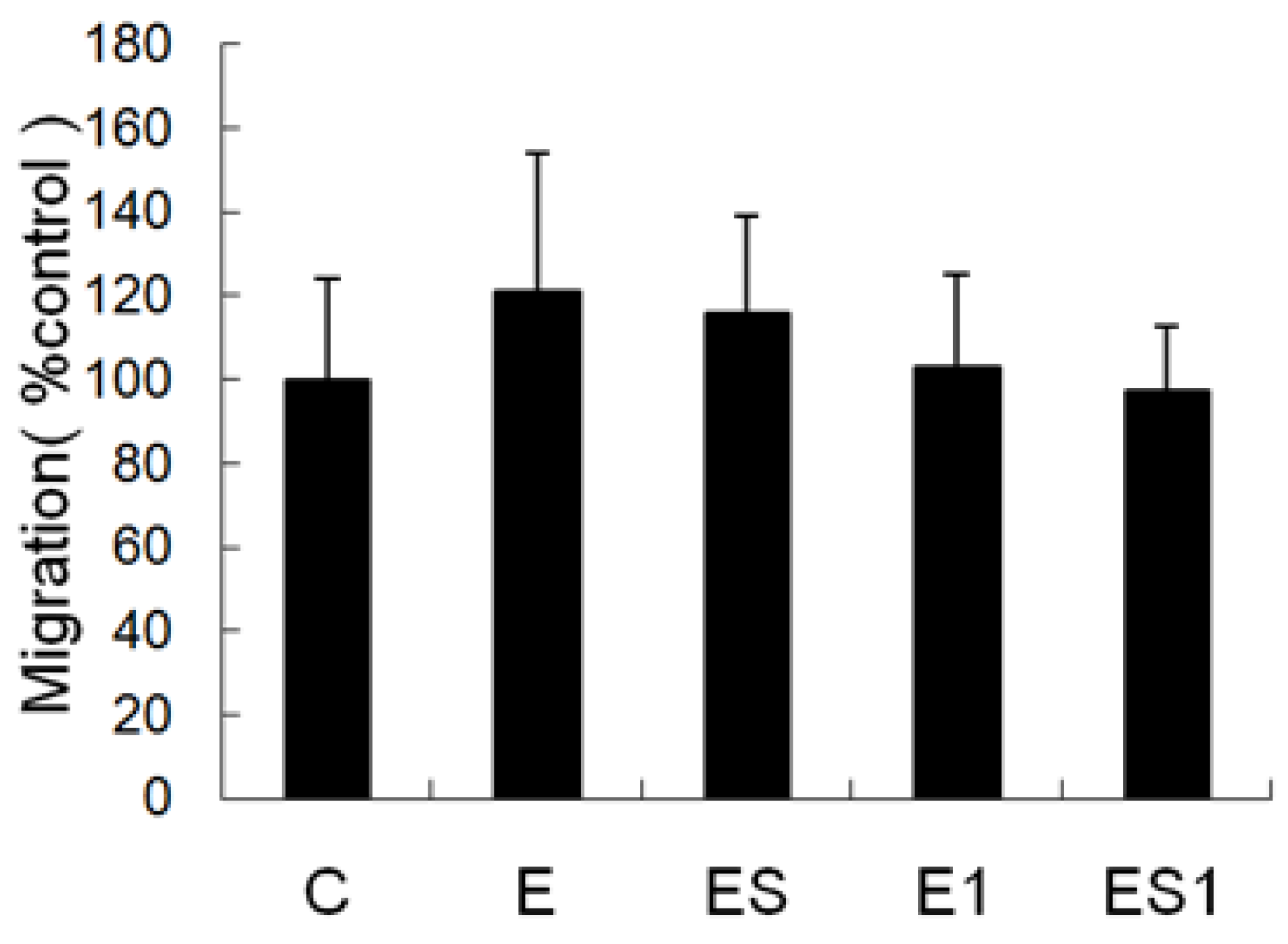
3.4. Chemotaxis
3.5. ROS Generation
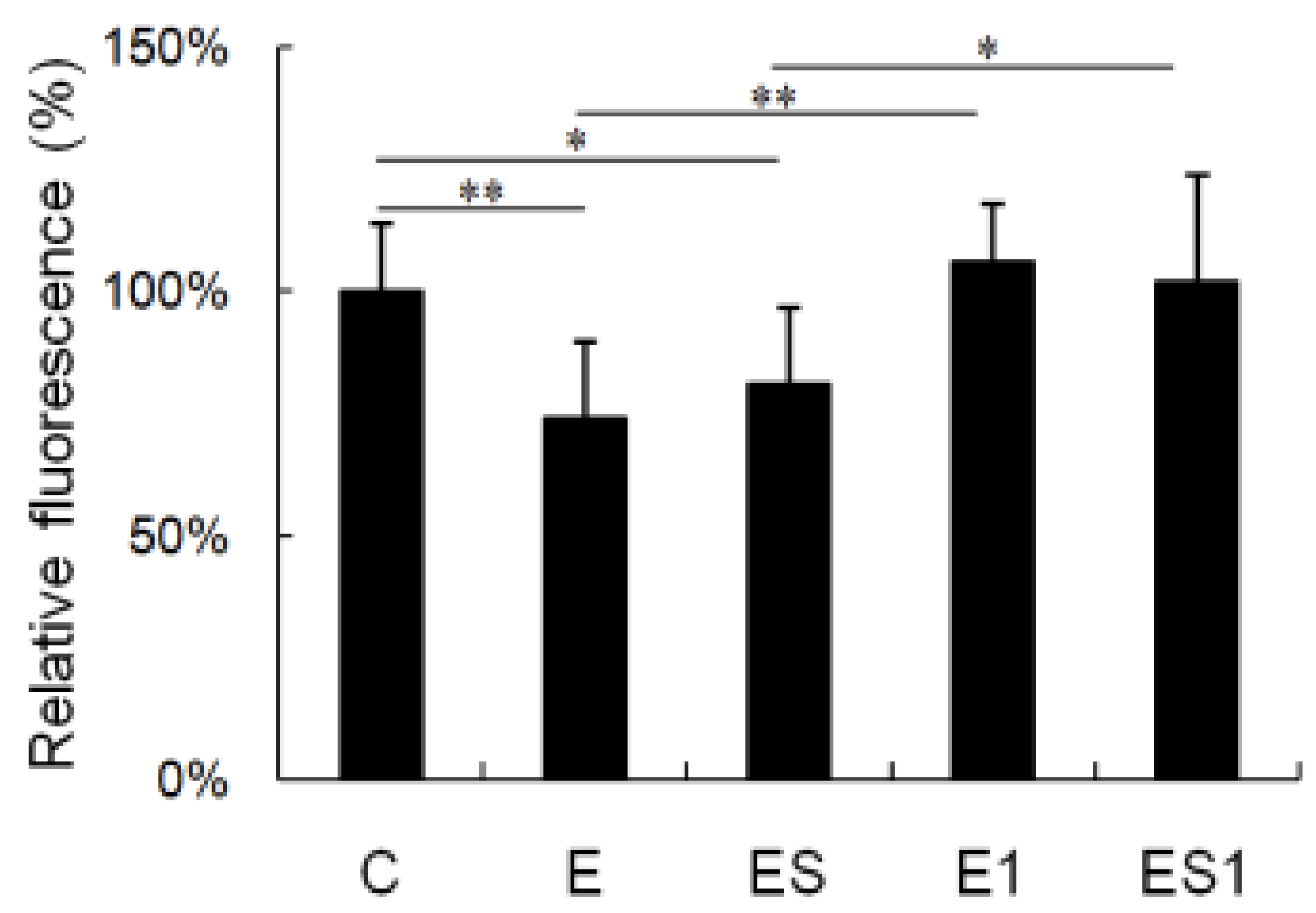
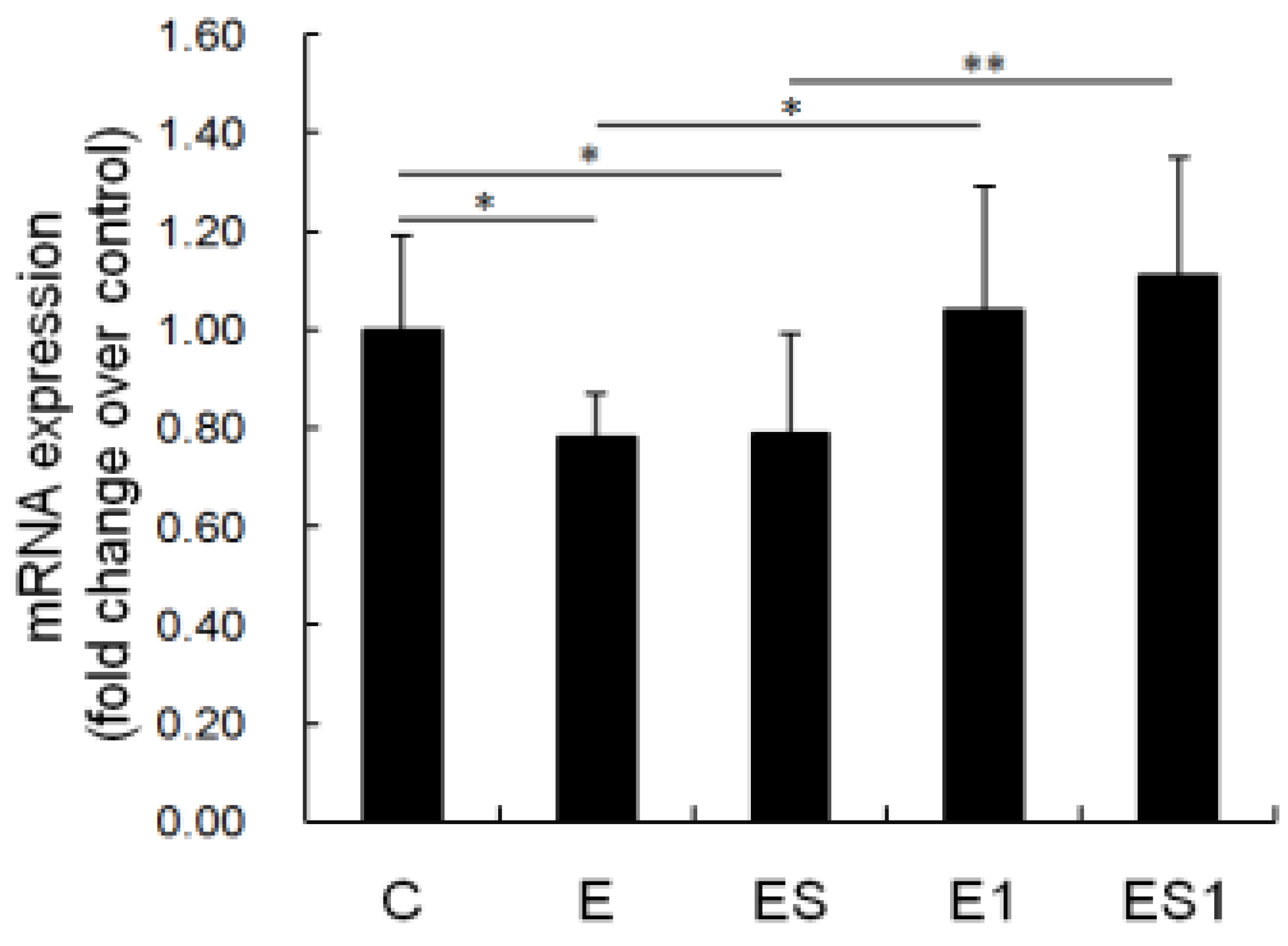
3.6. MHC II mRNA Level

4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Nieman, D.C. Clinical implications of exercise immunology. J. Sport Health Sci. 2012, 1, 12–17. [Google Scholar] [CrossRef]
- Peijie, C.; Hongwu, L.; Fengpeng, X.; Jie, R.; Jie, Z. Heavy load exercise induced dysfunction of immunity and neuroendocrine responses in rats. Life Sci. 2003, 72, 2255–2262. [Google Scholar] [CrossRef]
- Rama, L.; Teixeira, A.M.; Matos, A.; Borges, G.; Henriques, A.; Gleeson, M.; Pedreiro, S.; Filaire, E.; Alves, F.; Paiva, A. Changes in natural killer cell subpopulations over a winter training season in elite swimmers. Eur. J. Appl. Physiol. 2013, 113, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.H.; Chen, P.J.; Dong, J.M.; Wang, R.; Luo, B.B. Dietary glutamine supplementation partly reverses impaired macrophage function resulting from overload training in rats. Int. J. Sport Nutr. Exerc. Metab. 2015, 25, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.H.; Chen, P.J.; Wang, R.; Dong, J.M. Overload training inhibits phagocytosis and ROS generation of peritoneal macrophages: Role of IGF-1 and MGF. Eur. J. Appl. Physiol. 2013, 113, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, N.; Kobayashi, K. Macrophages in inflammation. Curr. Drug Targets Inflamm. Allergy 2005, 4, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Struder, H.K.; Hollmann, W.; Platen, P.; Wostmann, R.; Ferrauti, A.; Weber, K. Effect of exercise intensity on free tryptophan to branched-chain amino acids ratio and plasma prolactin during endurance exercise. Can. J. Appl. Physiol. Rev. Can. Physiol. Appl. 1997, 22, 280–291. [Google Scholar] [CrossRef]
- Holecek, M.; Kandar, R.; Sispera, L.; Kovarik, M. Acute hyperammonemia activates branched-chain amino acid catabolism and decreases their extracellular concentrations: Different sensitivity of red and white muscle. Amino Acids 2011, 40, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Coombes, J.S.; McNaughton, L.R. Effects of branched-chain amino acid supplementation on serum creatine kinase and lactate dehydrogenase after prolonged exercise. J. Sports Med. Phys. Fit. 2000, 40, 240–246. [Google Scholar]
- Shimomura, Y.; Yamamoto, Y.; Bajotto, G.; Sato, J.; Murakami, T.; Shimomura, N.; Kobayashi, H.; Mawatari, K. Nutraceutical effects of branched-chain amino acids on skeletal muscle. J. Nutr. 2006, 136, 529S–532S. [Google Scholar] [PubMed]
- Negro, M.; Giardina, S.; Marzani, B.; Marzatico, F. Branched-chain amino acid supplementation does not enhance athletic performance but affects muscle recovery and the immune system. J. Sports Med. Phys. Fit. 2008, 48, 347–351. [Google Scholar]
- Calder, P.C.; Yaqoob, P. Glutamine and the immune system. Amino Acids 1999, 17, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Newsholme, P. Why is l-glutamine metabolism important to cells of the immune system in health, postinjury, surgery or infection? J. Nutr. 2001, 131, 2515S–2522S. [Google Scholar] [PubMed]
- Calder, P.C. Branched-chain amino acids and immunity. J. Nutr. 2006, 136, 288S–293S. [Google Scholar] [PubMed]
- Li, P.; Yin, Y.L.; Li, D.; Kim, S.W.; Wu, G. Amino acids and immune function. Br. J. Nutr. 2007, 98, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Benito, B.; Gay-Jordi, G.; Serrano-Mollar, A.; Guasch, E.; Shi, Y.; Tardif, J.C.; Brugada, J.; Nattel, S.; Mont, L. Cardiac arrhythmogenic remodeling in a rat model of long-term intensive exercise training. Circulation 2011, 123, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Falavigna, G.; Alves de Araujo, J., Jr.; Rogero, M.M.; Pires, I.S.; Pedrosa, R.G.; Martins, E., Jr.; Alves de Castro, I.; Tirapegui, J. Effects of diets supplemented with branched-chain amino acids on the performance and fatigue mechanisms of rats submitted to prolonged physical exercise. Nutrients 2012, 4, 1767–1780. [Google Scholar] [CrossRef] [PubMed]
- Iwasa, M.; Kobayashi, Y.; Mifuji-Moroka, R.; Hara, N.; Miyachi, H.; Sugimoto, R.; Tanaka, H.; Fujita, N.; Gabazza, E.C.; Takei, Y. Branched-chain amino acid supplementation reduces oxidative stress and prolongs survival in rats with advanced liver cirrhosis. PLoS ONE 2013, 8, e70309. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, D.; Maki, T.; Herningtyas, E.H.; Ikeshita, N.; Shibahara, H.; Sugiyama, Y.; Nakanishi, S.; Iida, K.; Iguchi, G.; Takahashi, Y.; et al. Branched-chain amino acids protect against dexamethasone-induced soleus muscle atrophy in rats. Muscle Nerve 2010, 41, 819–827. [Google Scholar] [CrossRef] [PubMed]
- Deyl, Z.; Hyanek, J.; Horakova, M. Profiling of amino acids in body fluids and tissues by means of liquid chromatography. J. Chromatogr. 1986, 379, 177–250. [Google Scholar] [CrossRef]
- Yang, L.V.; Radu, C.G.; Wang, L.; Riedinger, M.; Witte, O.N. Gi-independent macrophage chemotaxis to lysophosphatidylcholine via the immunoregulatory GPCR G2A. Blood 2005, 105, 1127–1134. [Google Scholar] [CrossRef] [PubMed]
- Novak, M.L.; Bryer, S.C.; Cheng, M.; Nguyen, M.H.; Conley, K.L.; Cunningham, A.K.; Xue, B.; Sisson, T.H.; You, J.S.; Hornberger, T.A.; et al. Macrophage-specific expression of urokinase-type plasminogen activator promotes skeletal muscle regeneration. J. Immunol. 2011, 187, 1448–1457. [Google Scholar] [CrossRef] [PubMed]
- Long, F.; Wang, Y.X.; Liu, L.; Zhou, J.; Cui, R.Y.; Jiang, C.L. Rapid nongenomic inhibitory effects of glucocorticoids on phagocytosis and superoxide anion production by macrophages. Steroids 2005, 70, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Bae, Y.S.; Lee, J.H.; Choi, S.H.; Kim, S.; Almazan, F.; Witztum, J.L.; Miller, Y.I. Macrophages generate reactive oxygen species in response to minimally oxidized low-density lipoprotein: Toll-like receptor 4- and spleen tyrosine kinase-dependent activation of nadph oxidase 2. Circ. Res. 2009, 104, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Chirgwin, J.M.; Przybyla, A.E.; MacDonald, R.J.; Rutter, W.J. Isolation of biologically active ribonucleic acid from sources enriched in ribonuclease. Biochemistry 1979, 18, 5294–5299. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Chen, P.; Dong, J. Effects of overtraining on skeletal muscle growth and gene expression. Int. J. Sports Med. 2012, 33, 846–853. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.T.; Linderman, J.J.; Kirschner, D.E. Multiple mechanisms allow mycobacterium tuberculosis to continuously inhibit MHC class II-mediated antigen presentation by macrophages. Proc. Natl. Acad. Sci. USA 2005, 102, 4530–4535. [Google Scholar] [CrossRef] [PubMed]
- Noss, E.H.; Pai, R.K.; Sellati, T.J.; Radolf, J.D.; Belisle, J.; Golenbock, D.T.; Boom, W.H.; Harding, C.V. Toll-like receptor 2-dependent inhibition of macrophage class II MHC expression and antigen processing by 19-KDA lipoprotein of mycobacterium tuberculosis. J. Immunol. 2001, 167, 910–918. [Google Scholar] [CrossRef] [PubMed]
- Germain, R.N. MHC-dependent antigen processing and peptide presentation: Providing ligands for T lymphocyte activation. Cell 1994, 76, 287–299. [Google Scholar] [CrossRef]
- Chu, R.S.; Askew, D.; Noss, E.H.; Tobian, A.; Krieg, A.M.; Harding, C.V. CPG oligodeoxynucleotides down-regulate macrophage class II MHC antigen processing. J. Immunol. 1999, 163, 1188–1194. [Google Scholar] [PubMed]
- Taylor, A.E.; Finney-Hayward, T.K.; Quint, J.K.; Thomas, C.M.R.; Tudhope, S.J.; Wedzicha, J.A.; Barnes, P.J.; Donnelly, L.E. Defective macrophage phagocytosis of bacteria in copd. Eur. Respir. J. 2010, 35, 1039–1047. [Google Scholar] [CrossRef] [PubMed]
- Raidal, S.L.; Love, D.N.; Bailey, G.D.; Rose, R.J. The effect of high intensity exercise on the functional capacity of equine pulmonary alveolar macrophages and BAL-derived lymphocytes. Res. Vet. Sci. 2000, 68, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Woods, J.A.; Ceddia, M.A.; Kozak, C.; Wolters, B.W. Effects of exercise on the macrophage MHC II response to inflammation. Int. J. Sports Med. 1997, 18, 483–488. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.G. Cell signaling. H2O2, a necessary evil for cell signaling. Science 2006, 312, 1882–1883. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zeigler, M.M.; Lam, G.K.; Hunter, M.G.; Eubank, T.D.; Khramtsov, V.V.; Tridandapani, S.; Sen, C.K.; Marsh, C.B. The role of the NADPH oxidase complex, p38 MAPK, and AKT in regulating human monocyte/macrophage survival. Am. J. Respir. Cell Mol. Biol. 2007, 36, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.K.; Choi, Y.G.; Baik, J.Y.; Han, S.Y.; Jeong, D.W.; Bae, Y.S.; Kim, N.; Lee, S.Y. A crucial role for reactive oxygen species in RANKL-induced osteoclast differentiation. Blood 2005, 106, 852–859. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.M.; Donaldson, K.; Borm, P.J.; Schins, R.P.; Dehnhardt, M.; Gilmour, P.; Jimenez, L.A.; Stone, V. Calcium and ROS-mediated activation of transcription factors and TNF-α cytokine gene expression in macrophages exposed to ultrafine particles. Am. J. Physiol. Lung Cell. Mol. Physiol. 2004, 286, L344–L353. [Google Scholar] [CrossRef] [PubMed]
- Petro, T.M.; Bhattacharjee, J.K. Effect of dietary essential amino acid limitations upon the susceptibility to Salmonella typhimurium and the effect upon humoral and cellular immune responses in mice. Infect. Immun. 1981, 32, 251–259. [Google Scholar] [PubMed]
- Kitagawa, T.; Yokoyama, Y.; Kokuryo, T.; Nagino, M. Protective effects of branched-chain amino acids on hepatic ischemia-reperfusion-induced liver injury in rats: A direct attenuation of Kupffer cell activation. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 304, G346–G355. [Google Scholar] [CrossRef] [PubMed]
- De Simone, R.; Vissicchio, F.; Mingarelli, C.; De Nuccio, C.; Visentin, S.; Ajmone-Cat, M.A.; Minghetti, L. Branched-chain amino acids influence the immune properties of microglial cells and their responsiveness to pro-inflammatory signals. Biochim. Biophys. Acta 2013, 1832, 650–659. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, W.; Chen, P.; Liu, X.; Zhao, L. The Impaired Function of Macrophages Induced by Strenuous Exercise Could Not Be Ameliorated by BCAA Supplementation. Nutrients 2015, 7, 8645-8656. https://doi.org/10.3390/nu7105425
Xiao W, Chen P, Liu X, Zhao L. The Impaired Function of Macrophages Induced by Strenuous Exercise Could Not Be Ameliorated by BCAA Supplementation. Nutrients. 2015; 7(10):8645-8656. https://doi.org/10.3390/nu7105425
Chicago/Turabian StyleXiao, Weihua, Peijie Chen, Xiaoguang Liu, and Linlin Zhao. 2015. "The Impaired Function of Macrophages Induced by Strenuous Exercise Could Not Be Ameliorated by BCAA Supplementation" Nutrients 7, no. 10: 8645-8656. https://doi.org/10.3390/nu7105425
APA StyleXiao, W., Chen, P., Liu, X., & Zhao, L. (2015). The Impaired Function of Macrophages Induced by Strenuous Exercise Could Not Be Ameliorated by BCAA Supplementation. Nutrients, 7(10), 8645-8656. https://doi.org/10.3390/nu7105425