Red Ginseng Treatment for Two Weeks Promotes Fat Metabolism during Exercise in Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Treatment

{kind=link}
{kind=link}
{kind=link}
| Ingredient | Content |
|---|---|
| Rg1 | 0.71 |
| Re | 0.93 |
| Rf | 1.21 |
| Rf1 | 0.78 |
| Rg2(s) | 1.92 |
| Rg2® | 1.29 |
| Rb1 | 4.62 |
| Rc | 2.41 |
| Rb2 | 1.83 |
| Rd | 0.89 |
| Rg3(s) | 2.14 |
| Rg3® | 0.91 |
| Total | 19.64 |
2.2. Red Ginseng Extract
2.3. Training Methods
 ).
). 2.4. Measurement of Metabolic Rates
2.5. Gas Analysis
2.6. Blood Analysis
2.7. Glycogen Analysis
2.8. Statistical Analysis
3. Results
3.1. Body Weight and Food Intake
| CON | RG | |
|---|---|---|
| Initial body weight (g) | 35.1 ± 1.6 | 34.7 ± 1.4 |
| Final body weight (g) | 36.9 ± 2.0 | 36.1 ± 1.2 |
| Body weight gain (g) | 1.6 ± 1.2 | 1.4 ± 1.0 |
| Total amount of food intake (g/2weeks) | 73.0 ± 6.8 | 77.6 ± 5.9 |
| Daily food intake (g/day) | 4.5 ± 0.4 | 4.8 ± 0.3 |
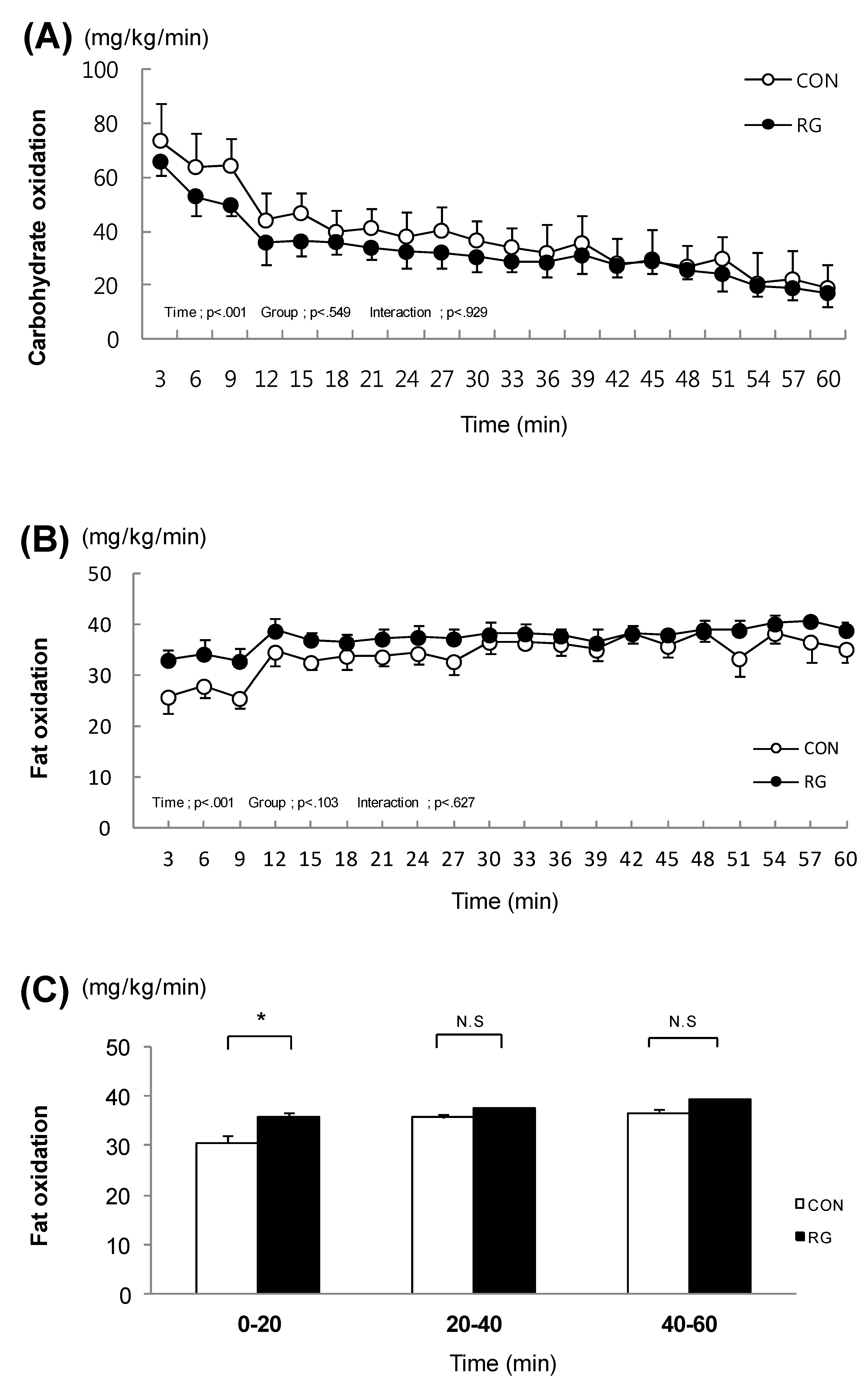
3.2. Carbohydrate Oxidation and Fat Oxidation during Exercise
. After the two-week experimental period, carbohydrate oxidation during exercise was not different between the two groups (Figure 1A). In addition, after the experimental period, fat oxidation during exercise was not different between the two groups (Figure 1B). In contrast, when examining each session of exercise separately, fat oxidation during the initial 20 min of the one-hour exercise was significantly higher in the RG group (35.5 ± 0.8 mg/kg/min) than in the CON group (30.5 ± 1.5 mg/kg/min) (Figure 1C).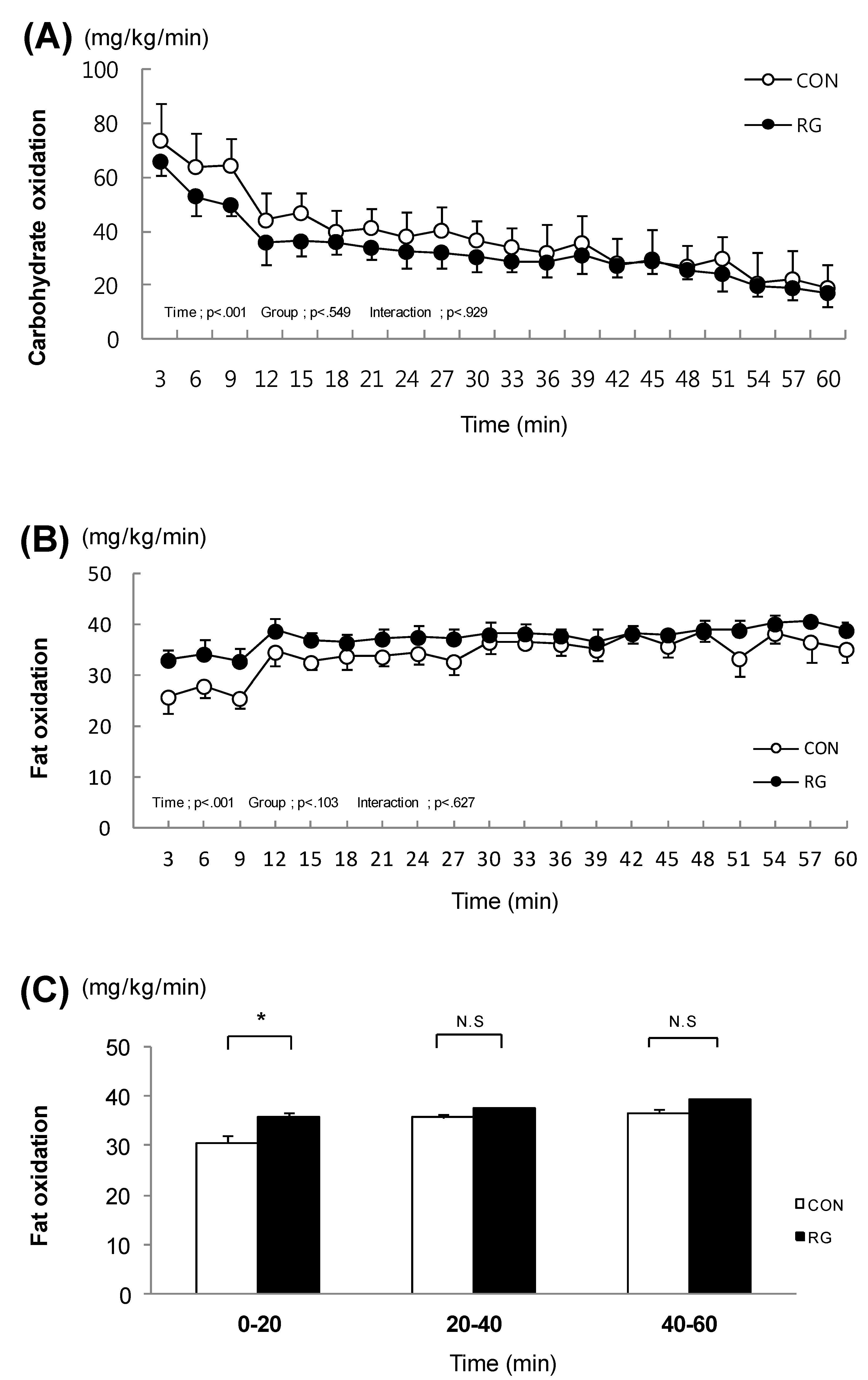
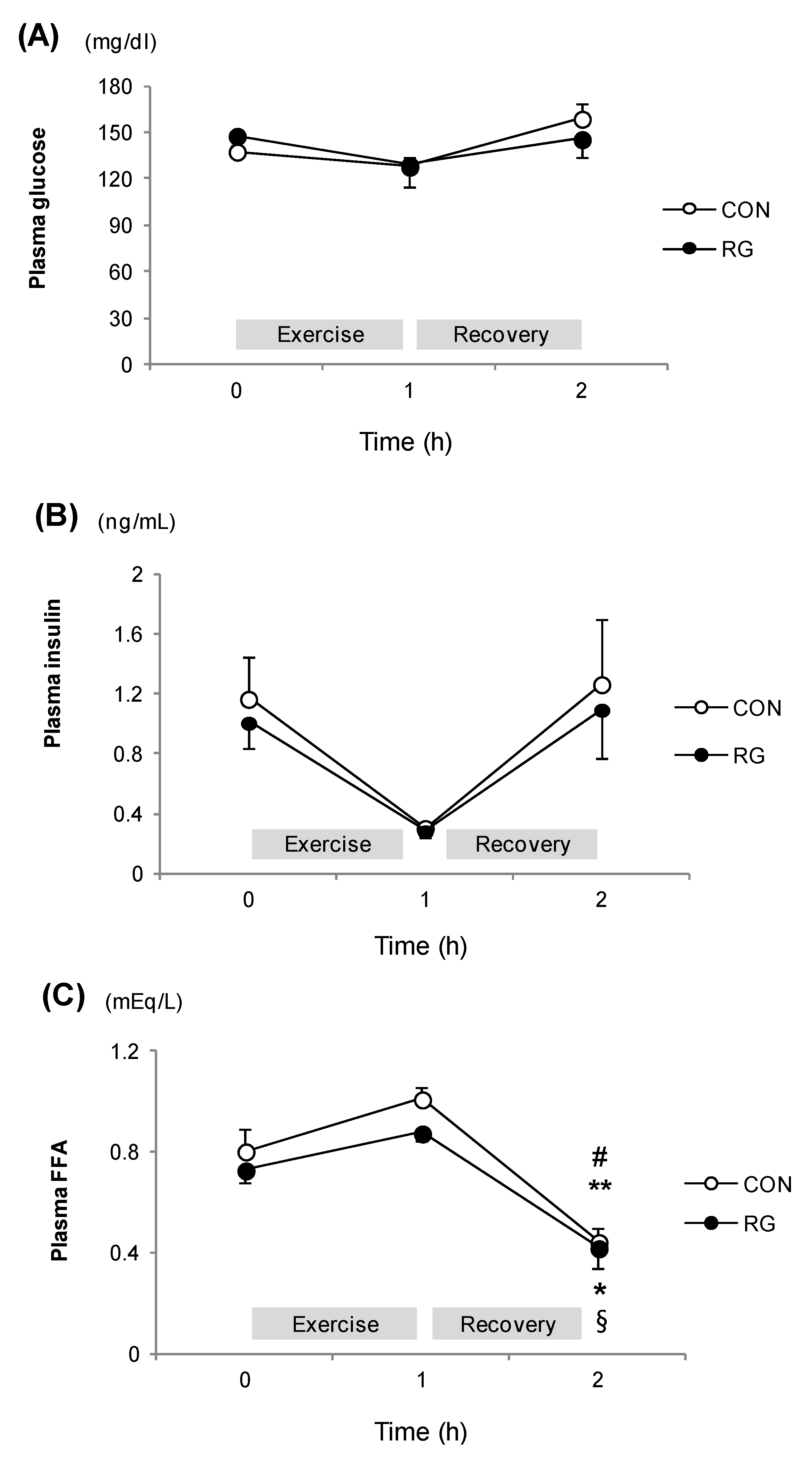
3.3. Plasma Parameters

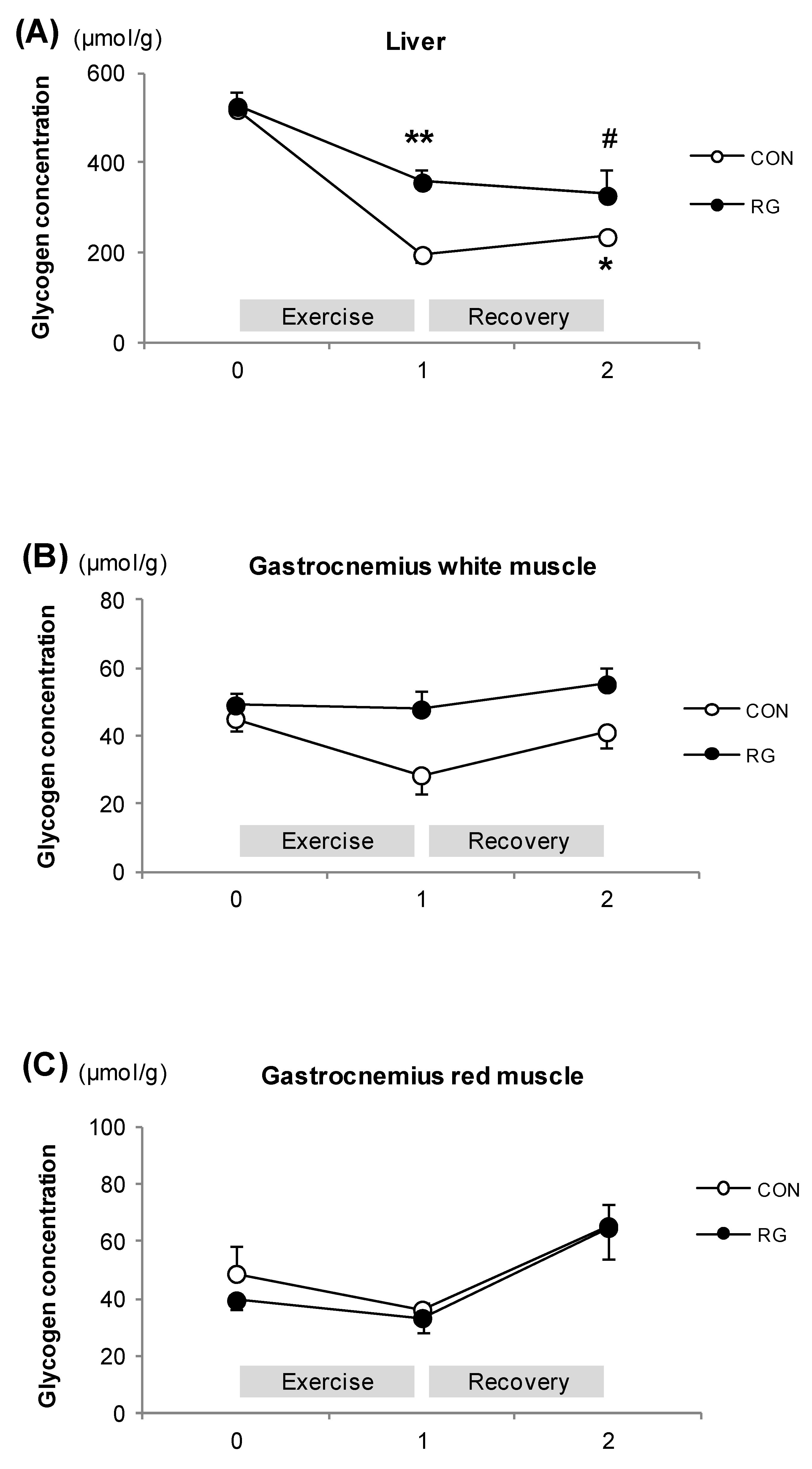
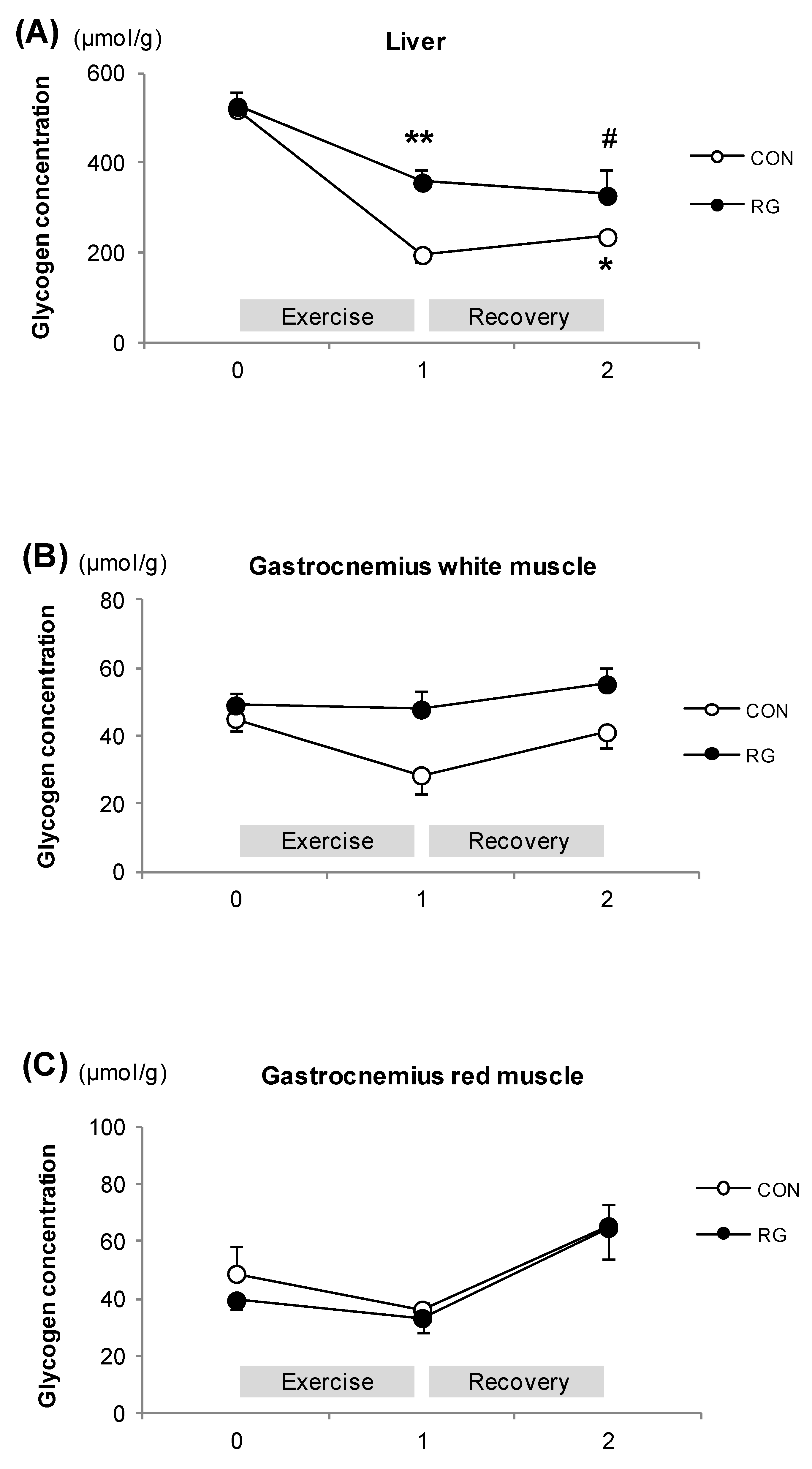
3.4. Glycogen Concentrations
4. Discussion
5. Conclusions
Acknowledgment
Author Contributions
Conflicts of Interest
References
- Lee, H.; Hong, M.K.; Shin, S.A.; Son, T.Y.; Ann, E.S.; Kim, W.S.; Hong, Y. The effect of BCAA administration on the central fatigue and endurance exercise capacity. Exerc. Sci. 2002, 11, 25–34. [Google Scholar]
- Inoue, K.; Fushiki, T. Exercise and central fatigue. Korean J. Exerc. Nutr. 2003, 7, 227–233. [Google Scholar]
- Lim, K.W. Effects of branched chain amino acid ingestion on m RNA expression of lactate transpoters and lactate metabolism during endurance exercise in rats and athletes. Korean J. Exerc. Nutr. 2006, 10, 9–16. [Google Scholar]
- Ikeuchi, M.; Koyama, T.; Takahashi, J.; Yazawa, K. Effects of astaxanthin supplementation on exercise-induced fatigue in mice. Biol. Pharm. Bull. 2006, 29, 2106–2110. [Google Scholar] [CrossRef]
- Kiefer, D.; Pantuso, T. Panax ginseng. Am Fam Physician. 2003, 15, 1539–1542. [Google Scholar]
- Shin, Y.W.; Choi, H.J.; Park, J.E.; Kim, D.H.; Park, J.H.; Kim, N.J. Effect of Heat Processed Ginseng on Anti-Fatigue. Korean J. Pharmacogn. 2006, 37, 246–252. [Google Scholar]
- Chen, C.F.; Chiou, W.F.; Zhang, J.T. Comparison of the pharmacological effects of Panax ginseng and Panax quinquefolium. Acta Pharmacol. Sin. 2008, 29, 1103–1108. [Google Scholar] [CrossRef]
- Lee, H.J.; Lee, Y.H.; Park, S.K.; Kang, E.S.; Kim, H.J.; Lee, Y.C.; Choi, C.S.; Park, S.E.; Ahn, C.W.; Cha, B.S.; et al. Korean red ginseng (Panax ginseng) improves insulin sensitivity and attenuates the development of diabetes in Otsuka Long-Evans Tokushima fatty rats. Metabolism 2009, 58, 1170–1177. [Google Scholar] [CrossRef]
- Yoon, M.; Lee, H.; Jeong, S.; Kim, J.J.; Nicol, C.J.; Nam, K.W.; Kim, M.; Cho, B.G.; Oh, G.T. Peroxisome proliferator-activated receptor a is involved in the regulation of lipid metabolism by ginseng. Br. J. Pharmacol. 2003, 138, 1295–1302. [Google Scholar] [CrossRef]
- Song, Y.B.; An, Y.R.; Kim, S.J.; Park, H.W.; Jung, J.W.; Kyung, J.S.; Hwang, S.Y.; Kim, Y.S. Lipid metabolic effect of Korean red ginseng extract in mice fed on a high-fat diet. J. Sci. Food Agric. 2012, 30, 388–396. [Google Scholar]
- Avakian, E.V.; Sugimoto, R.B.; Taguchi, S.; Horvath, S.M. Effect of Panax ginseng Extract on Energy Metabolism during Exercise in Rats. Planta Med. 1984, 50, 151–154. [Google Scholar] [CrossRef]
- Avakian, E.V.; Evonuk, E. Effect of Panax ginseng extract on tissue glycogen and adrenal cholesterol depletion during prolonged exercise. Planta Med. 1979, 36, 43–48. [Google Scholar] [CrossRef]
- Hwang, H.J.; Kim, J.S.; Kim, B.K.; Lee, C.H.; Suh, H.J.; Yun, H.Y.; Park, J.H.; Lim, K.W. Effects of Korean red ginseng intake during endurance training on resting energy metabolism in male mice. J. Exerc. Nutr. Biochem. 2013, 17, 1–5. [Google Scholar]
- Jeon, Y.R.; Kim, J.S.; Hwang, H.J.; Lim, K.W. Effects of endurance training for 4 weeks on resting metabolic rate and excess post-exercise oxygen consumption in mouse. J. Exerc. Nutr. Biochem. 2012, 16, 113–122. [Google Scholar]
- Kim, J.S.; Hwang, H.J.; Yun, H.Y.; Kim, B.K.; Lee, C.H.; Suh, H.J.; Lim, K.W. Silk peptide intake increases fat oxidation at rest in exercised mice. J. Nutr. Sci. Vitaminol. 2013, 59, 250–255. [Google Scholar] [CrossRef]
- Lim, K.W.; Kim, J.S.; Jeon, Y.R.; Hwang, H.J.; Suh, H.J. Measurement of resting metabolic rate using metabolic chamber in resting rats. J. Exerc. Nutr. Biochem. 2011, 15, 35–40. [Google Scholar]
- Passonneau, J.V.; Lauderdale, V.R. A comparison of three methods of glycogen measurement in tissues. Anal. Biochem. 1974, 60, 405–412. [Google Scholar] [CrossRef]
- Karu, N.; Reifen, R.; Kerem, Z. Weight gain reduction in mice fed Panax ginseng saponin, a pancreatic lipase inhibitor. J. Agric. Food Chem. 2007, 18, 2824–2828. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, S.A.; Han, S.M.; Shim, I. Comparison of the antiobesity effects of the protopanaxadiol- and protopanaxatriol-type saponins of red ginseng. Phytother. Res. 2009, 23, 78–85. [Google Scholar] [CrossRef]
- Lee, S.H.; Lee, H.J.; Lee, Y.H.; Lee, B.W.; Cha, B.S.; Kang, E.S.; Ahn, C.W.; Park, J.S.; Kim, H.J.; Lee, E.Y.; et al. Korean red ginseng (Panax ginseng) improves insulin sensitivity in high fat fed Sprague-Dawley rats. Phytother. Res. 2012, 26, 142–147. [Google Scholar] [CrossRef]
- Jeukendrup, A.E.; Saris, W.H.; Wagenmakers, A.J. Fat Metabolism During Exercise: A Review. Part I: Fatty Acid Mobilization and Muscle Metabolism. Int. J. Sports Med. 1998, 19, 231–244. [Google Scholar] [CrossRef]
- Tang, W.; Zhang, Y.; Gao, J.; Ding, X.; Gao, S. The Anti-fatigue Effect of 20(R)-Ginsenoside Rg3 in Mice by Intranasally Administration. Biol. Pharm. Bull. 2008, 31, 2024–2027. [Google Scholar] [CrossRef]
- Xiong, Y.; Shen, L.; Liu, K.J.; Tso, P.; Xiong, Y.; Wang, G.; Woods, S.C.; Liu, M. Antiobesity and antihyperglycemic effects of ginsenoside Rb1 in rats. Diabetes 2010, 59, 2505–2512. [Google Scholar] [CrossRef]
- Richter, E.A.; Hargreaves, M. Exercise, GLUT4, and Skeletal Muscle Glucose Uptake. Physiol. Rev. 2013, 93, 993–1017. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hwang, H.; Kim, J.; Park, J.; Yun, H.; Cheon, W.-K.; Kim, B.; Lee, C.-H.; Suh, H.; Lim, K. Red Ginseng Treatment for Two Weeks Promotes Fat Metabolism during Exercise in Mice. Nutrients 2014, 6, 1874-1885. https://doi.org/10.3390/nu6051874
Hwang H, Kim J, Park J, Yun H, Cheon W-K, Kim B, Lee C-H, Suh H, Lim K. Red Ginseng Treatment for Two Weeks Promotes Fat Metabolism during Exercise in Mice. Nutrients. 2014; 6(5):1874-1885. https://doi.org/10.3390/nu6051874
Chicago/Turabian StyleHwang, Hyejung, Jisu Kim, Jonghoon Park, Heayeon Yun, Woo-Kwang Cheon, Bokyung Kim, Chi-Ho Lee, Heajung Suh, and Kiwon Lim. 2014. "Red Ginseng Treatment for Two Weeks Promotes Fat Metabolism during Exercise in Mice" Nutrients 6, no. 5: 1874-1885. https://doi.org/10.3390/nu6051874
APA StyleHwang, H., Kim, J., Park, J., Yun, H., Cheon, W.-K., Kim, B., Lee, C.-H., Suh, H., & Lim, K. (2014). Red Ginseng Treatment for Two Weeks Promotes Fat Metabolism during Exercise in Mice. Nutrients, 6(5), 1874-1885. https://doi.org/10.3390/nu6051874