Potential of Phytase-Mediated Iron Release from Cereal-Based Foods: A Quantitative View


Abstract
:1. Introduction
2. Dietary Iron Bioavailability
3. Function, Structure and Properties of Phytic Acid
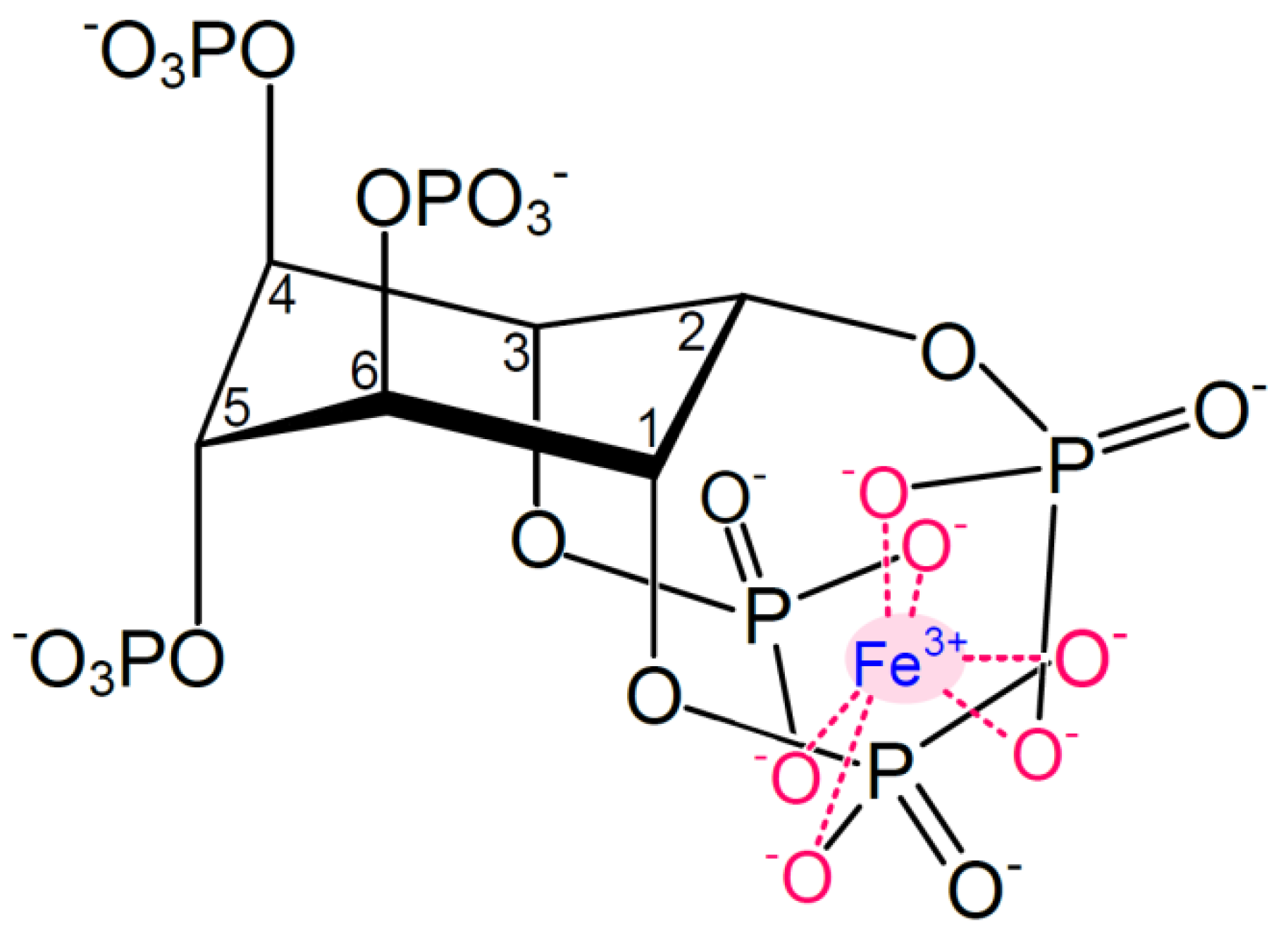

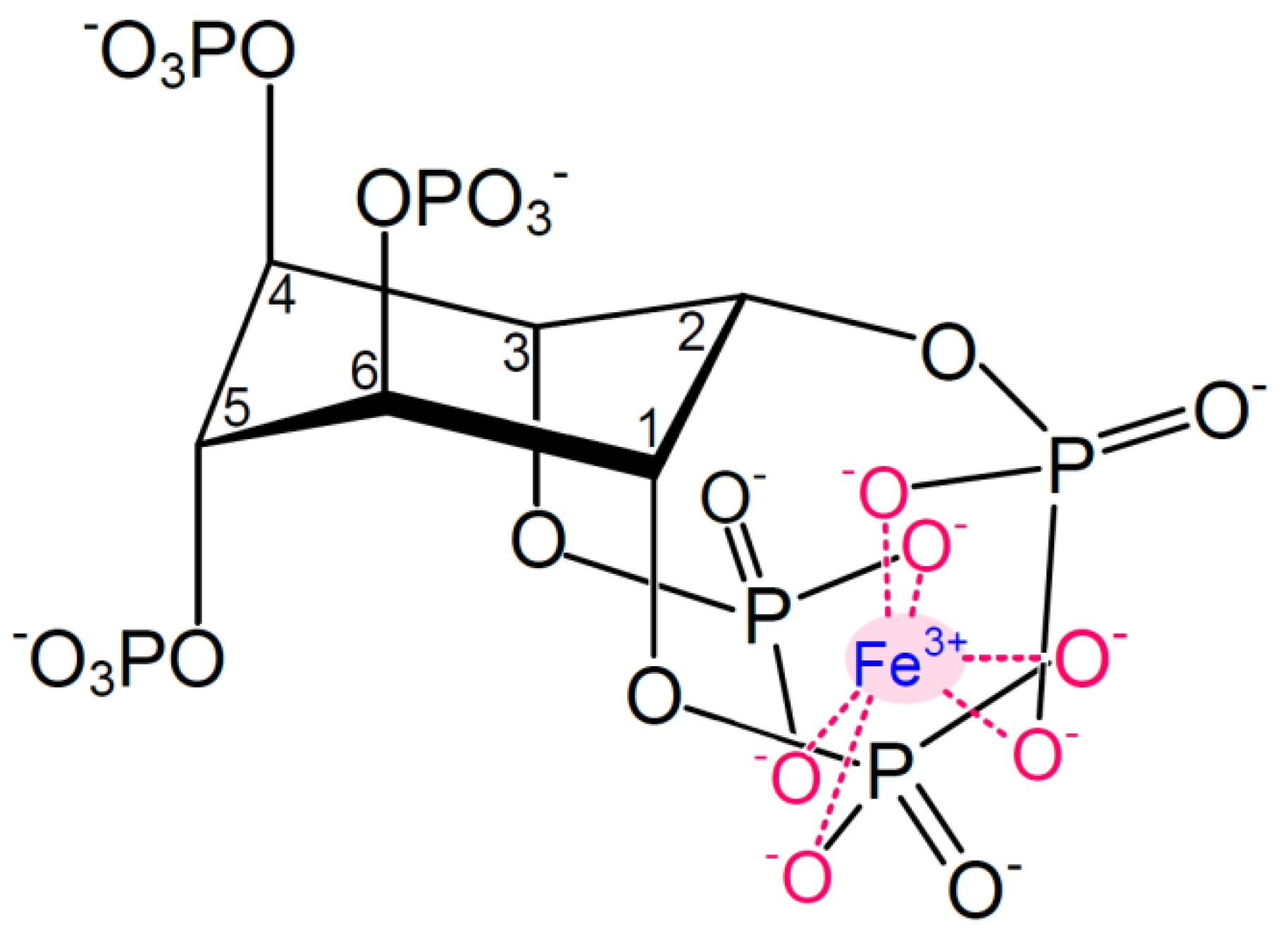{kind=link}
| Food | Phytic acid (g/100 g) | Iron content (mg/100 g) 1 | Estimated 2 molar ratio PA:Fe |
|---|---|---|---|
| Oatmeal | 0.80–1.03 | 3.9 | 17–22 |
| White bread | 0.28–1.00 | 1.0 | 24–85 |
| Whole grain bread | 0.43–1.05 | 1.4 | 26–64 |
| Rye bread (whole meal) | 0.03–0.41 | 1.5 | 2–23 |
| Rice | 0.06–2.20 | 1.2 | 4–155 |
4. Iron Intake and Requirements
5. Non-Haem Iron Absorption in Humans
6. Chemical Form of Iron Phytate Complexes in the Gastrointestinal System
7. Degradation of Phytate in the Human Gastrointestinal System
8. Phytases
9. Degradation of Iron Phytate Complexes in the Gastrointestinal System
9.1. Activity of Phytases in the Gastrointestinal System
9.2. Role of Exogenous Phytases in Gastrointestinal Phytate Degradation
9.3. Extent of Phytate Degradation and Iron Absorption
10. Potential of Phytase-Mediated Iron Release from Plant Foods
10.1. Effect of Evaluation Parameters in in Vitro Studies
- Extent of phytate degradation and content of lower inositol phosphates;
- Dialysability of iron;
- Cellular availability and/or uptake of iron.
10.1.1. Phytate Degradation
10.1.2. Dialysability of Iron
10.1.3. Caco-2 Cellular Iron Uptake
10.2. Effect Evaluation Parameters in in Vivo Studies
10.3. Previous Studies on Phytases for Food Processing in Vitro and in Vivo
10.3.1. Studies Evaluating Efficacy of in Vivo Phytase Catalysis
| Treatment | Study design and duration | Iron dosage | Test group | Evaluation parameter | Result | Reference |
|---|---|---|---|---|---|---|
| Active or inactivated endogenous wheat bran phytase | Meal study 2 × 4 day periods | n/a | 8 M, 1 F ileostomy patients (no information on iron status) | Phytate degradation (Fe absorption not evaluated) | ↑ from ~5% with deactivated wheat bran phytase to ~60% with active wheat bran phytase | [64] |
| Active or inactivated endogenous wheat phytase or AN phytase | 2 separate meal studies with wheat rolls: (1) Active or inactivated wheat phytase; (2) Phytase-inactivated wheat bran w/wo AN phytase | 3.7 mg labelled Fe + 0.4 mg intrinsic Fe/meal | 9 M, 11 F split into 10 in each substudy (no information on iron status) | Fe absorption (measurement of radiolabelled 55Fe/59Fe in blood samples) | (1) No difference; (2) ↑ 83% from 14.3% ± 2.6% to 26.1% ± 3.8% with AN phytase | [75] |
| Maize porridge taken with different micronutrient powders containing Fe (as NaFeEDTA or FeSO4) with AN phytase, AA, l-α-glycerophospho-choline and/or Ca | Meal study Crossover design (2 days) | 3 mg Fe/meal | 101 F (21 ID, 1 ID anaemic) allocated to 6 separate iron absorption studies (n = 16–18 in each substudy) | Fe absorption (stable isotope labelled 57Fe/58Fe measurement in erythrocytes 14 days later) | Phytase: ↑ 23%–75% from 2.4%–5.0% to 4.1%–7.4%. Highest effect of phytase was observed in combination with ascorbic acid, where total increase as result of phytase, ascorbic acid, and NaFeEDTA resulted in 400% absorption increase compared to iron absorption from FeSO4 (1.5% to 7.4%). No significant effect of other single factors | [74] |
| High-phytate porridge taken with micronutrient powder containing Zn, Fe as NaFeEDTA, AA and AN phytase | Diet study (5 days/week for 23 weeks) Double-blind controlled study Control: No Fe supplement | 2.5 mg Fe/meal | 200 C (low iron status) | Fe status (SF, Hb, CRP, TfR content in blood samples) | Occurrence of Fe deficiency ↓ 75% (↓ 35% in control group) Body Fe ↑ 100% (↑ 40% in control) Fe absorption ~7%–8% | [98] |
| Fe-rich bread w/wo phytase supplement compared with FeSO4 supplement | Meal study | 5–8 mg Fe/meal | 24 F (borderline anaemic) allocated to 5 test meals | Fe status (serum iron measured 180 and 210 min after ingestion) | Serum Fe ↓ in all groups at 180 and 210 min after intake except for the positive control group given FeSO4 supplement. Decrease in serum Fe was largest in the order of iron-rich bread > control bread > iron-rich bread with 0.010% (w/w) phytase > iron-rich bread with 0.015% (w/w) phytase | [99] |
10.3.2. Studies Evaluating Efficacy of ex Vivo Phytase Catalysis on in Vivo Parameters
10.3.3. Studies Evaluating Efficacy of ex Vivo Phytase Catalysis on in Vitro Parameters
| Treatment | Study design and duration | Iron dosage | Test group | Evaluation parameter | Result | Reference |
|---|---|---|---|---|---|---|
| AN phytase addition to cereal porridges during manufacturing | Meal study | 2.5 mg Fe/meal | 34 M (1 ID), 44 F (13 ID), all NA | Fe absorption (measurement of radiolabel-led 55Fe/59Fe in blood samples) | ↑ 209%–1066% from 0.3% to 2.4%→2.8%–11.5% | [105] |
| AN phytase added during making of fibre-rich wheat bread | Intervention study; 4 months | 6 mg Fe/meal, 14 mg/day | 41 iron-sufficient F | Fe absorption in vivo (SF) InsP3-6 content in bread | No effect on SF by phytase treatment; InsP5+6 contents ↓ 17%; InsP3-6 content ↓ 22% | [5] |
| CA and AN phytase addition to oat beverage as pretreatment | Meal study, 4 days | 1.3 mg Fe/meal | 23 M, 22 F (non-ID) | Fe absorption in vivo (radio-labelled 55Fe/59Fe count in whole body and eryth-rocytes); InsP3-6 content in oat beverage | CA: ↑54% (from 3.9% to 6.0%) Phytase added with CA further increased iron absorption by 78% (6.0%→10.7%); Phytase reduced InsP3-6 to undetectable amounts (<1 mg/portion), phytate-P ↓ 83% | [101] |
| Treatment | Evaluation parameter | Result | Reference |
|---|---|---|---|
| Addition of AN, A. fumigatus or E. coli phytase w/wo CA and AA to whole-wheat bread dough | Phytate 1 degradation
Fe dialysability from bread | CA: Phytate degradation ↑ from 42% in control to 69%; Fe dialysability ↑ 12-fold AN phytase alone: Phytate degradation 57%, no significant effect on dialysability Microbial phytases + CA: AN: Phytate degradation 74%–85% (dose-dependent); Fe dialysability ↑ 15-fold. E. coli: Phytate degradation 63%–76% (dose-dependent); Fe dialysability not assessed A. fumigatus: Phytate degradation 83%–85% (dose-dependent); Fe dialysability not assessed AN phytase + CA + AA: Fe dialysability ↑ 24-fold compared to control | [21] |
| AN phytase addition during making of different bread | Phytate 1 degradation | Phytate degradation ↑ 12%–57% compared to control depending on bread type, total phytate degradation with phytase: 49%–90% | [106] |
| High-phytase producing yeast strains addition during in vitro digestion of wheat gruel | Phytate degradation | ≤59% | [91] |
| Bifidobacteria cell suspensions or AN phytase addition during wheat bread making | InsP3-6 content in bread | Phytase: each InsP ↓ 67%–100% (mean ± standard deviation 91% ± 10%) compared to control; Bifidobacteria: each ↓ −8%–67% (19% ± 27%) compared to control | [107] |
| Highly phytase-producing yeasts (experimental strains), S. cerevisiae or A. ficuum phytase addition to togwa prior to ingestion | Phytate degradation after 48 h fermentation | S. cerevisiae: 85% A. ficuum phytase: 89% Experimental strains: 95% Control: 51% | [108] |
| Activating endogenous phytase in fortified and non-fortified flour/oilseed mixtures | Fe dialysability | Wheat/soy: ↑ 43%–162% (non-fortified); ↑ 40%–168% (fortified); Wheat/groundnut: ↑ 83%–192% (non-fortified); 97%–240% (fortified) Dialysable Fe in controls: 1.3%–9.7% | [109] |
| AN phytase addition during in vitro digestion of bread | Phytate degradation
Fe bioavailability in Caco-2 cell model | Phytate degradation 89%–97%
Cellular uptake of Fe ↑ 152%–156% | [102] |
| Fungal phytase or phytase-producing bifidobacteria addition during whole wheat bread- or whole wheat sourbread-making | InsP5 + InsP6 degradation
Fe dialysability Ferritin formation in Caco-2 cells | Whole wheat bread: Fungal phytase: InsP5+6 degradation ~97%; dialysable Fe ↑ 17%, dialysability ↓ 9%; Caco-2 cell uptake ↑ 92% Bifidobacteria: InsP5+6 degradation ~76%; dialysable Fe ↑ 127%, dialysability ↑ 118%; Caco-2 cell uptake ↑ 33% (not significant) Whole wheat sourbread: Fungal phytase: InsP5+6 degradation ~68%; dialysable Fe ↑ 45%, dialysability ↑ 82%; Caco-2 cell uptake ↑ 67% (not significant) Bifidobacteria: InsP5+6 degradation of ~58%–70%; dialysable Fe ↑ 283%–456%; dialysability ↑ 236%–391%; Caco-2 cell uptake ↑ 0%–25% (not significant) | [93] |
| A. oryzae phytase or phytase-producing bifidobacteria addition during pretreatment of infant cereals | Phytate degradation
Fe dialysability | A. oryzae phytase: InsP6 ↓ 45%–67% compared to control; InsP3-6 (total) ↓ 25%–28%; no change in Fe dialysability Bifidobacteria: InsP6 ↓ 75%–87% compared to control; InsP3-6 (total) ↓ 23%–63%; no change in Fe dialysability | [103] |
| Exogenous wheat phytase addition to wheat flour | Phytate 1 degradation
Fe dialysability | Phytate ↓ 35%–69%
Fe dialysability ↑ 11%–52% (from 12% to 16% to 15%–25%) | [104] |
11. Other Options for Enhancing Iron Absorption
12. Conclusions
Conflict of Interest
References
- World Health Organization, Worldwide Prevalence of Anaemia 1993–2005; WHO Press: Geneva, Switzerland, 2008.
- Bohn, L.; Meyer, A.S.; Rasmussen, S.K. Phytate: Impact on environment and human nutrition. A challenge for molecular breeding. J. Zhejiang Univ. Sci. B 2008, 9, 165–191. [Google Scholar]
- Schlemmer, U.; Frølich, W.; Prieto, R.M.; Grases, F. Phytate in foods and significance for humans: Food sources, intake, processing, bioavailability, protective role and analysis. Mol. Nutr. Food Res. 2009, 53, S330–S375. [Google Scholar] [CrossRef]
- Hunt, J.R. Bioavailability of iron, zinc, and other trace minerals from vegetarian diets. Am. J. Clin. Nutr. 2003, 78, 633S–639S. [Google Scholar]
- Kristensen, M.B.; Tetens, I.; Jørgensen, A.B.; Thomsen, A.D.; Milman, N.; Hels, O.; Sandström, B.; Hansen, M. A decrease in iron status in young healthy women after long-term daily consumption of the recommended intake of fibre-rich wheat bread. Eur. J. Nutr. 2005, 44, 334–340. [Google Scholar] [CrossRef]
- Minihane, A.M.; Rimbach, G. Iron absorption and the iron binding and anti-oxidant properties of phytic acid. Int. J. Food Sci. Technol. 2002, 37, 741–748. [Google Scholar] [CrossRef]
- Hurrell, R.; Egli, I. Iron bioavailability and dietary reference values. Am. J. Clin. Nutr. 2010, 91, 1461S–1467S. [Google Scholar] [CrossRef]
- Hunt, J.R.; Roughead, Z.K. Nonheme-iron absorption, fecal ferritin excretion, and blood indexes of iron status in women consuming controlled lactoovovegetarian diets for 8 wk. Am. J. Clin. Nutr. 1999, 69, 944–952. [Google Scholar]
- Monsen, E.R. Ascorbic Acid: An Enhancing Factor in Iron Absorption. In Nutritional Bioavailability of Iron; American Chemical Society: Washington, DC, USA, 1982; pp. 85–95. [Google Scholar]
- Cook, J.D. Adaptation in iron metabolism. Am. J. Clin. Nutr. 1990, 51, 301–308. [Google Scholar]
- Monsen, E.R. Iron nutrition and absorption: Dietary factors which impact iron bioavailability. J. Am. Diet. Assoc. 1988, 88, 786–790. [Google Scholar]
- Hurrell, R.F. Influence of vegetable protein sources on trace element and mineral bioavailability. J. Nutr. 2003, 133, 2973S–2977S. [Google Scholar]
- Theil, E.C.; Briat, J.-F. Plant Ferritin and Non-Heme Iron Nutrition in Humans; International Food Policy Research Institute and International Center for Tropical Agriculture: Washington, DC, USA, 2004. [Google Scholar]
- May, L.; Morris, E.R.; Ellis, R. Chemical identity of iron in wheat by Mössbauer spectroscopy. J. Agric. Food Chem. 1980, 28, 1004–1006. [Google Scholar] [CrossRef]
- Morris, E.R.; Ellis, R. Isolation of monoferric phytate from wheat bran and its biological value as an iron source to the rat. J. Nutr. 1976, 106, 753–760. [Google Scholar]
- Grusak, M.A.; Pearson, J.N.; Marentes, E. The physiology of micronutrient homeostasis in field crops. Field Crop. Res. 1999, 60, 41–56. [Google Scholar]
- Harrison, P.M.; Arosio, P. The ferritins: Molecular properties, iron storage function and cellular regulation. Biochim. Biophys. Acta Bioenerg. 1996, 1275, 161–203. [Google Scholar] [CrossRef]
- Theil, E.C.; Chen, H.; Miranda, C.; Janser, H.; Elsenhans, B.; Núñes, M.T.; Pizarro, F.; Schümann, K. Absorption of iron from ferritin is independent of heme iron and ferrous salts in women and rat intestinal segments. J. Nutr. 2012, 142, 478–483. [Google Scholar] [CrossRef]
- Kalgaonkar, S.; Lönnerdal, B. Receptor-mediated uptake of ferritin-bound iron by human intestinal Caco-2 cells. J. Nutr. Biochem. 2009, 20, 304–311. [Google Scholar] [CrossRef]
- Reddy, M.B.; Hurrell, R.F.; Cook, J.D. Estimation of nonheme-iron bioavailability from meal composition. Am. J. Clin. Nutr. 2000, 71, 937–943. [Google Scholar]
- Porres, J.M.; Etcheverry, P.; Miller, D.D.; Lei, X.G. Phytase and citric acid supplementation in whole-wheat bread improves phytate-phosphorus release and iron dialyzability. J. Food Sci. 2001, 66, 614–619. [Google Scholar] [CrossRef]
- Tetens, I.; Bendtsen, K.M.; Henriksen, M.; Ersbøll, A.K.; Milman, N. The impact of a meat- versus a vegetable-based diet on iron status in women of childbearing age with small iron stores. Eur. J. Nutr. 2007, 46, 439–445. [Google Scholar] [CrossRef]
- García-Casal, M.N.; Layrisse, M.; Solano, L.; Barón, M.A.; Arguello, F.; Llovera, D.; Ramírez, J.; Leets, I.; Tropper, E. Vitamin A and beta-carotene can improve nonheme iron absorption from rice, wheat and corn by humans. J. Nutr. 1998, 128, 646–650. [Google Scholar]
- Layrisse, M.; García-Casal, M.N.; Solano, L.; Barón, M.A.; Arguello, F.; Llovera, D.; Ramírez, J.; Leets, I.; Tropper, E. New property of vitamin A and B-carotene on human iron absorption: Effect on phytate and polyphenols as inhibitors of iron absorption. Arch. Latinoam. Nutr. 2000, 50, 243–248. [Google Scholar]
- Andrews, N.C. Iron Metabolism. In Principles of Molecular Medicine; Runge, M.S., Patterson, C., Eds.; Humana Press: New York, NY, USA, 2006; pp. 848–853. [Google Scholar]
- Petry, N.; Egli, I.; Zeder, C.; Walczyk, T.; Hurrell, R. Polyphenols and phytic acid contribute to the low iron bioavailability from common beans in young women. J. Nutr. 2010, 140, 1977–1982. [Google Scholar] [CrossRef]
- Janet, R. Hunt algorithms for iron and zinc bioavailability: Are they accurate? Int. J. Vitam. Nutr. Res. 2010, 80, 257–262. [Google Scholar] [CrossRef]
- Fairweather-Tait, S.J.; Collings, R. Estimating the bioavailability factors needed for setting dietary reference values. Int. J. Vitam. Nutr. Res. 2010, 80, 249–256. [Google Scholar] [CrossRef]
- Hambidge, K.M. Micronutrient bioavailability: Dietary reference intakes and a future perspective. Am. J. Clin. Nutr. 2010, 91, 1430S–1432S. [Google Scholar] [CrossRef]
- McKie, A.T. The role of Dcytb in iron metabolism: an update. Biochem. Soc. Trans. 2008, 36, 1239–1241. [Google Scholar] [CrossRef]
- Hurrell, R.F. Fortification: Overcoming technical and practical barriers. J. Nutr. 2002, 132, 806S–812S. [Google Scholar]
- Hallberg, L. Wheat fiber, phytates and iron absorption. Scand. J. Gastroenterol. Suppl. 1987, 22, 73–79. [Google Scholar] [CrossRef]
- Sandberg, A.-S. Bioavailability of minerals in legumes. Br. J. Nutr. 2002, 88, S281–S285. [Google Scholar] [CrossRef]
- Hurrell, R.F. Phytic acid degradation as a means of improving iron absorption. Int. J. Vitam. Nutr. Res. 2004, 74, 445–452. [Google Scholar] [CrossRef]
- Jin, F.; Frohman, C.; Thannhauser, T.W.; Welch, R.M.; Glahn, R.P. Effects of ascorbic acid, phytic acid and tannic acid on iron bioavailability from reconstituted ferritin measured by an in vitro digestion-Caco-2 cell model. Br. J. Nutr. 2009, 101, 972–981. [Google Scholar] [CrossRef]
- Lönnerdal, B. The importance and bioavailability of phytoferritin-bound iron in cereals and legume foods. Int. J. Vitam. Nutr. Res. 2007, 77, 152–157. [Google Scholar] [CrossRef]
- Loewus, F.A. Biosynthesis of Phytate in Food Grains and Seeds. In Food Phytates; Reddy, N.R., Sathe, S.K., Eds.; CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- Raboy, V. Progress in breeding low phytate crops. J. Nutr. 2002, 132, 503S–505S. [Google Scholar]
- Lott, J.N.A. Accumulation of Seed Reserves of Phosphorous and Other Minerals. In Seed Physiology; Academic Press Australia: Toowong, Australia, 1984; pp. 139–166. [Google Scholar]
- Bohn, L.; Josefsen, L.; Meyer, A.S.; Rasmussen, S.K. Quantitative analysis of phytate globoids isolated from wheat bran and characterization of their sequential dephosphorylation by wheat phytase. J. Agric. Food Chem. 2007, 55, 7547–7552. [Google Scholar] [CrossRef]
- Borg, S.; Brinch-Pedersen, H.; Tauris, B.; Holm, P.B. Iron transport, deposition and bioavailability in the wheat and barley grain. Plant Soil 2009, 325, 15–24. [Google Scholar] [CrossRef]
- Costello, A.J.R.; Glonek, T.; Myers, T.C. 31P nuclear magnetic resonance-pH titrations of myo-inositol hexaphosphate. Carbohydr. Res. 1976, 46, 159–171. [Google Scholar] [CrossRef]
- Reddy, N.R. Occurrence, Distribution, Content, and Dietary Intake of Phytate. In Food Phytates; Reddy, N.R., Sathe, S.K., Eds.; CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar]
- Lott, J.N.; Ockenden, I.; Raboy, V.; Batten, G.D. Phytic acid and phosphorus in crop seeds and fruits: A global estimate. Seed Sci. Res. 2000, 10, 11–33. [Google Scholar]
- Saxholt, E.; Christensen, A.T.; Møller, A.; Hartkopp, H.B.; Hess Ygil, K.; Hels, O.H. Danish Food Composition Databank, Revision 7; Department of Nutrition, National Food Institute, Technical University of Denmark: Lyngby, Denmark, 2008; Available online: http://www.foodcomp.dk/ (accessed on 28 March 2013).
- Pantopoulos, K.; Porwal, S.K.; Tartakoff, A.; Devireddy, L. Mechanisms of mammalian iron homeostasis. Biochemistry 2012, 51, 5705–5724. [Google Scholar] [CrossRef]
- Council, N. Nordic Nutrition Recommendations 2004: Integrating Nutrition and Physical Activity; Nordic Council of Ministers: Copenhagen, Denmark, 2005. [Google Scholar]
- Lynch, S. The precision of in vitro methods and algorithms for predicting the bioavailability of dietary iron. Int. J. Vitam. Nutr. Res. 2005, 75, 436–445. [Google Scholar] [CrossRef]
- West, A.R.; Oates, P.S. Mechanisms of heme iron absorption: Current questions and controversies. World J. Gastroenterol. 2008, 14, 4101–4110. [Google Scholar] [CrossRef]
- Alagozlu, H.; Karakan, T. Probiotics are sensitive microorganisms: Lactobacillus can be affected by stomach acid, antibiotic, and bowel cleansing. Dig. Dis. Sci. 2012, 57, 2477–2478. [Google Scholar] [CrossRef]
- Dressman, J.B.; Berardi, R.R.; Dermentzoglou, L.C.; Russell, T.L.; Schmaltz, S.P.; Barnett, J.L.; Jarvenpaa, K.M. Upper gastrointestinal (GI) pH in young, healthy men and women. Pharm. Res. 1990, 7, 756–761. [Google Scholar] [CrossRef]
- Malagelada, J.-R.; Go, V.L.W.; Summerskill, W.H.J. Different gastric, pancreatic, and biliary responses to solid-liquid or homogenized meals. Dig. Dis. Sci. 1979, 24, 101–110. [Google Scholar] [CrossRef]
- Bretti, C.; Cigala, R.M.; Lando, G.; Milea, D.; Sammartano, S. Sequestering ability of phytate toward biologically and environmentally relevant trivalent metal cations. J. Agric. Food Chem. 2012, 60, 8075–8082. [Google Scholar] [CrossRef]
- Ismail-Beigi, F.; Faraji, B.; Reinhold, J.G. Binding of zinc and iron to wheat bread, wheat bran, and their components. Am. J. Clin. Nutr. 1977, 30, 1721–1725. [Google Scholar]
- Veiga, N.; Torres, J.; Mansell, D.; Freeman, S.; Domínguez, S.; Barker, C.J.; Díaz, A.; Kremer, C. “Chelatable iron pool”: Inositol 1,2,3-trisphosphate fulfils the conditions required to be a safe cellular iron ligand. J. Biol. Inorg. Chem. 2009, 14, 51–59. [Google Scholar] [CrossRef]
- Šala, M.; Makuc, D.; Kolar, J.; Plavec, J.; Pihlar, B. Potentiometric and 31P NMR studies on inositol phosphates and their interaction with iron(III) ions. Carbohydr. Res. 2011, 346, 488–494. [Google Scholar] [CrossRef]
- Sandberg, A.-S.; Andlid, T. Phytogenic and microbial phytases in human nutrition. Int. J. Food Sci. Technol. 2002, 37, 823–833. [Google Scholar] [CrossRef]
- McCance, R.A.; Widdowson, E.M. Phytin in human nutrition. Biochem. J. 1935, 29, 2694–2699. [Google Scholar]
- Hoff-Jørgensen, E.; Andersen, O.; Nielsen, G. The effect of phytic acid on the absorption of calcium and phosphorus. 3. In Children. Biochem. J. 1946, 40, 555–557. [Google Scholar]
- Sandberg, A.-S.; Andersson, H.; Kivistö, B.; Sandström, B. Extrusion cooking of a high-fibre cereal product. 1. Effects on digestibility and absorption of protein, fat, starch, dietary fibre and phytate in the small intestine. Br. J. Nutr. 1986, 55, 245–254. [Google Scholar] [CrossRef]
- Kivistö, B.; Andersson, H.; Cederblad, G.; Sandberg, A.-S.; Sandström, B. Extrusion cooking of a high-fibre cereal product. 2. Effects on apparent absorption of zinc, iron, calcium, magnesium and phosphorus in humans. Br. J. Nutr. 1986, 55, 255–260. [Google Scholar] [CrossRef]
- Sandberg, A.-S.; Andersson, H.; Carlsson, N.-G.; Sandström, B. Degradation products of bran phytate formed during digestion in the human small intestine: Effect of extrusion cooking on digestibility. J. Nutr. 1987, 117, 2061–2065. [Google Scholar]
- Iqbal, T.H.; Lewis, K.O.; Cooper, B.T. Phytase activity in the human and rat small intestine. Gut 1994, 35, 1233–1236. [Google Scholar] [CrossRef]
- Sandberg, A.-S.; Andersson, H. Effect of dietary phytase on the digestion of phytate in the stomach and small intestine of humans. J. Nutr. 1988, 118, 469–473. [Google Scholar]
- Lei, X.G.; Weaver, J.D.; Mullaney, E.; Ullah, A.H.; Azain, M.J. Phytase, a new life for an “old” enzyme. Annu. Rev. Anim. Biosci. 2013, 1, 283–309. [Google Scholar] [CrossRef]
- Greiner, R.; Konietzny, U. Phytase for food application. Food Technol. Biotechnol. 2006, 44, 125–140. [Google Scholar]
- Lei, X.G.; Porres, J.M. Phytase enzymology, applications, and biotechnology. Biotechnol. Lett. 2003, 25, 1787–1794. [Google Scholar] [CrossRef]
- Phillippy, B. Stability of Plant and Microbial Phytases. In Food Phytates; Reddy, N.R., Sathe, S.K., Eds.; CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- Pandey, A.; Szakacs, G.; Soccol, C.R.; Rodriguez-Leon, J.A.; Soccol, V.T. Production, purification and properties of microbial phytases. Bioresour. Technol. 2001, 77, 203–214. [Google Scholar] [CrossRef]
- Lei, X.G.; Porres, J.M.; Mullaney, E.J.; Brinch-Pedersen, H. Phytase: Source, Structure and Application. In Industrial Enzymes; Polaina, J., MacCabe, A.P., Eds.; Springer: Heidelberg, The Netherlands, 2007; pp. 505–529. [Google Scholar]
- World Health Organization, Safety Evaluation of Certain Food Additives. In WHO Food Additives Series: 67; WHO Press: Geneva, Switzerland, 2012.
- Kraemer, K.; Steiger, G.; Wieprecht, T. Iron Fortification Nutritional Blend. EP 2166876 B1. 10 April 2013. [Google Scholar]
- Schlemmer, U.; Jany, K.D.; Berk, A.; Schulz, E.; Rechkemmer, G. Degradation of phytate in the gut of pigs—Pathway of gastrointestinal inositol phosphate hydrolysis and enzymes involved. Arch. Anim. Nutr. 2001, 55, 255–280. [Google Scholar]
- Troesch, B.; Egli, I.; Zeder, C.; Hurrell, R.F.; de Pee, S.; Zimmermann, M.B. Optimization of a phytase-containing micronutrient powder with low amounts of highly bioavailable iron for in-home fortification of complementary foods. Am. J. Clin. Nutr. 2009, 89, 539–544. [Google Scholar] [CrossRef]
- Sandberg, A.-S.; Hulthén, L.R.; Türk, M. Dietary Aspergillus niger phytase increases iron absorption in humans. J. Nutr. 1996, 126, 476–480. [Google Scholar]
- Kemme, P.A.; Schlemmer, U.; Mroz, Z.; Jongbloed, A.W. Monitoring the stepwise phytate degradation in the upper gastrointestinal tract of pigs. J. Sci. Food Agric. 2006, 86, 612–622. [Google Scholar] [CrossRef]
- Phillippy, B.Q. Susceptibility of wheat and Aspergillus niger phytases to inactivation by gastrointestinal enzymes. J. Agric. Food Chem. 1999, 47, 1385–1388. [Google Scholar] [CrossRef]
- Yu, S.; Cowieson, A.; Gilbert, C.; Plumstead, P.; Dalsgaard, S. Interactions of phytate and myo-inositol phosphate esters (IP 1–5) including IP5 isomers with dietary protein and iron and inhibition of pepsin. J. Anim. Sci. 2012, 90, 1824–1832. [Google Scholar] [CrossRef]
- Brune, M.; Rossander-Hultén, L.; Hallberg, L.; Sandberg, A.-S. Iron absorption from bread in humans: Inhibiting effects of cereal fiber, phytate and inositol phosphates with different numbers of phosphate groups. J. Nutr. 1992, 122, 442–449. [Google Scholar]
- Sandberg, A.-S.; Brune, M.; Carlsson, N.-G.; Hallberg, L.; Skoglund, E.; Rossander-Hulthén, L. Inositol phosphates with different numbers of phosphate groups influence iron absorption in humans. Am. J. Clin. Nutr. 1999, 70, 240–246. [Google Scholar]
- Tang, J.; Leung, A.; Leung, C.; Lim, B.L. Hydrolysis of precipitated phytate by three distinct families of phytases. Soil Biol. Biochem. 2006, 38, 1316–1324. [Google Scholar] [CrossRef]
- Heubner, W.; Stadler, H. Über eine titration-methode zur bestimmung des phytins. Biochem. Z. 1914, 64, 432–437. [Google Scholar]
- Latta, M.; Eskin, M. A simple and rapid colorimetric method for phytate determination. J. Agric. Food Chem. 1980, 28, 1313–1315. [Google Scholar] [CrossRef]
- Carlsson, N.-G.; Bergman, E.-L.; Skoglund, E.; Hasselblad, K.; Sandberg, A.-S. Rapid analysis of inositol phosphates. J. Agric. Food Chem. 2001, 49, 1695–1701. [Google Scholar] [CrossRef]
- Haraldsson, A.-K.; Rimsten, L.; Alminger, M.; Andersson, R.; Åman, P.; Sandberg, A.-S. Digestion of barley malt porridges in a gastrointestinal model: Iron dialysability, iron uptake by Caco-2 cells and degradation of β-glucan. J. Cereal Sci. 2005, 42, 243–254. [Google Scholar] [CrossRef]
- Sandberg, A.-S. Methods and options in vitro dialyzability; benefits and limitations. Int. J. Vitam. Nutr. Res. 2005, 75, 395–404. [Google Scholar] [CrossRef]
- Argyri, K.; Theophanidi, E.; Kapna, A.; Staikidou, C.; Pounis, G.; Komaitis, M.; Georgiou, C.; Kapsokefalou, M. Iron or zinc dialyzability obtained from a modified in vitro digestion procedure compare well with iron or zinc absorption from meals. Food Chem. 2011, 127, 716–721. [Google Scholar] [CrossRef]
- Fairweather-Tait, S.; Lynch, S.; Hotz, C.; Hurrell, R.; Beebe, S.; Bering, S.; Bukhave, K.; Glahn, R.; Hambidge, M.; Hunt, J.; et al. The usefulness of in vitro models to predict the bioavailability of iron and zinc: A consensus statement from the HarvestPlus expert consultation. Int. J. Vitam. Nutr. Res. 2005, 75, 371–374. [Google Scholar] [CrossRef]
- Laparra, J.M.; Glahn, R.P.; Miller, D.D. Different responses of Fe transporters in Caco-2/HT29-MTX cocultures than in independent Caco-2 cell cultures. Cell Biol. Int. 2009, 33, 971–977. [Google Scholar] [CrossRef]
- Sandberg, A.-S. The use of Caco-2 cells to estimate Fe absorption in humans—A critical appraisal. Int. J. Vitam. Nutr. Res. 2010, 80, 307–313. [Google Scholar] [CrossRef]
- Haraldsson, A.-K.; Veide, J.; Andlid, T.; Alminger, M.L.; Sandberg, A.-S. Degradation of phytate by high-phytase Saccharomyces cerevisiae strains during simulated gastrointestinal digestion. J. Agric. Food Chem. 2005, 53, 5438–5444. [Google Scholar] [CrossRef]
- Yun, S.; Habicht, J.-P.; Miller, D.D.; Glahn, R.P. An in vitro digestion/Caco-2 cell culture system accurately predicts the effects of ascorbic acid and polyphenolic compounds on iron bioavailability in humans. J. Nutr. 2004, 134, 2717–2721. [Google Scholar]
- Sanz-Penella, J.M.; Laparra, J.M.; Sanz, Y.; Haros, M. Assessment of iron bioavailability in whole wheat bread by addition of phytase-producing bifidobacteria. J. Agric. Food Chem. 2012, 60, 3190–3195. [Google Scholar] [CrossRef]
- World Health Organization, Assessing the Iron Status of Populations; WHO Press: Geneva, Switzerland, 2007.
- Lynch, S.R. Why nutritional iron deficiency persists as a worldwide problem. J. Nutr. 2011, 141, 763S–768S. [Google Scholar] [CrossRef]
- Casgrain, A.; Collings, R.; Harvey, L.J.; Hooper, L.; Fairweather-Tait, S.J. Effect of iron intake on iron status: A systematic review and meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2012, 96, 768–780. [Google Scholar] [CrossRef]
- Zimmermann, M.B.; Molinari, L.; Staubli-Asobayire, F.; Hess, S.Y.; Chaouki, N.; Adou, P.; Hurrell, R.F. Serum transferrin receptor and zinc protoporphyrin as indicators of iron status in African children. Am. J. Clin. Nutr. 2005, 81, 615–623. [Google Scholar]
- Troesch, B.; van Stujivenberg, M.E.; Smuts, C.M.; Kruger, H.S.; Biebinger, R.; Hurrell, R.F.; Baumgartner, J.; Zimmermann, M.B. A Micronutrient powder with low doses of highly absorbable iron and zinc reduces iron and zinc deficiency and improves weight-for-age Z-scores in South African children. J. Nutr. 2011, 141, 237–242. [Google Scholar] [CrossRef]
- Bokhari, F.; Derbyshire, E.; Li, W.; Brennan, C.S.; Stojceska, V. A study to establish whether food-based approaches can improve serum iron levels in child-bearing aged women. J. Hum. Nutr. Diet. 2012, 25, 95–100. [Google Scholar] [CrossRef]
- Conway, R.E.; Geissler, C.A.; Hider, R.C.; Thompson, R.P.H.; Powell, J.J. Serum iron curves can be used to estimate dietary iron bioavailability in humans. J. Nutr. 2006, 136, 1910–1914. [Google Scholar]
- Zhang, H.; Onning, G.; Öste, R.; Gramatkovski, E.; Hulthén, L. Improved iron bioavailability in an oat-based beverage: The combined effect of citric acid addition, dephytinization and iron supplementation. Eur. J. Nutr. 2007, 46, 95–102. [Google Scholar] [CrossRef]
- Shockravi, S.; Almgren, A.; Carlsson, N.-G.; Sandberg, A.-S. Dephytinisation of Sangak and Barbari bread made from different extraction rate flours increases iron and zinc bioaccessibility in Caco-2 cells. Int. J. Food Sci. Technol. 2012, 47, 2252–2258. [Google Scholar] [CrossRef]
- Sanz-Penella, J.M.; Frontela, C.; Ros, G.; Martinez, C.; Monedero, V.; Haros, M. Application of bifidobacterial phytases in infant cereals: Effect on phytate contents and mineral dialyzability. J. Agric. Food Chem. 2012, 60, 11787–11792. [Google Scholar] [CrossRef]
- Akhter, S.; Saeed, A.; Irfan, M.; Malik, K.A. In vitro dephytinization and bioavailability of essential minerals in several wheat varieties. J. Cereal Sci. 2012, 56, 741–746. [Google Scholar] [CrossRef]
- Hurrell, R.F.; Reddy, M.B.; Juillerat, M.-A.; Cook, J.D. Degradation of phytic acid in cereal porridges improves iron absorption by human subjects. Am. J. Clin. Nutr. 2003, 77, 1213–1219. [Google Scholar]
- Haros, M.; Rosell, C.M.; Benedito, C. Fungal phytase as a potential breadmaking additive. Eur. Food Res. Technol. 2001, 213, 317–322. [Google Scholar] [CrossRef]
- Sanz-Penella, J.M.; Tamayo-Ramos, J.A.; Sanz, Y.; Haros, M. Phytate reduction in bran-enriched bread by phytase-producing bifidobacteria. J. Agric. Food Chem. 2009, 57, 10239–10244. [Google Scholar] [CrossRef]
- Hellström, A.M.; Almgren, A.; Carlsson, N.-G.; Svanberg, U.; Andlid, T.A. Degradation of phytate by Pichia kudriavzevii TY13 and Hanseniaspora guilliermondii TY14 in Tanzanian togwa. Int J. Food Microbiol. 2012, 153, 73–77. [Google Scholar] [CrossRef]
- Shilpa, K.S.; Lakshmi, A.J. Comparison of enhancement in bioaccessible iron and zinc in native and fortified high-phytate oilseed and cereal composites by activating endogenous phytase. Int. J. Food Sci. Technol. 2012, 47, 1613–1619. [Google Scholar] [CrossRef]
- Zhao, F.-J.; Shewry, P.R. Recent developments in modifying crops and agronomic practice to improve human health. Food Policy 2011, 36, S94–S101. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Nielsen, A.V.F.; Tetens, I.; Meyer, A.S. Potential of Phytase-Mediated Iron Release from Cereal-Based Foods: A Quantitative View. Nutrients 2013, 5, 3074-3098. https://doi.org/10.3390/nu5083074
Nielsen AVF, Tetens I, Meyer AS. Potential of Phytase-Mediated Iron Release from Cereal-Based Foods: A Quantitative View. Nutrients. 2013; 5(8):3074-3098. https://doi.org/10.3390/nu5083074
Chicago/Turabian StyleNielsen, Anne V. F., Inge Tetens, and Anne S. Meyer. 2013. "Potential of Phytase-Mediated Iron Release from Cereal-Based Foods: A Quantitative View" Nutrients 5, no. 8: 3074-3098. https://doi.org/10.3390/nu5083074
APA StyleNielsen, A. V. F., Tetens, I., & Meyer, A. S. (2013). Potential of Phytase-Mediated Iron Release from Cereal-Based Foods: A Quantitative View. Nutrients, 5(8), 3074-3098. https://doi.org/10.3390/nu5083074