Perspectives on Immunoglobulins in Colostrum and Milk

Abstract
:
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Immune Milk
2.1. Overview
2.2. Homologous Transfer of Passive Immunity
2.3. Heterologous Transfer of Passive Immunity
2.4. Immune Milk and Diseases Causing Diarrhea
2.5. Immune Milk and Dental Caries
2.6. Immune Milk and Intestinal Parasites
2.7. Immunization to Boost Colostrum and Milk Antibodies
3. Immunoglobulin Structure and Function
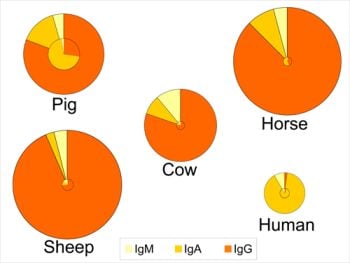
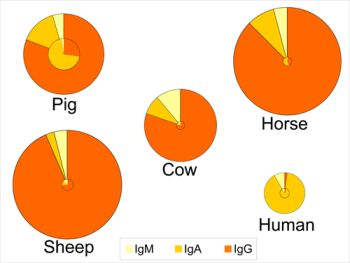
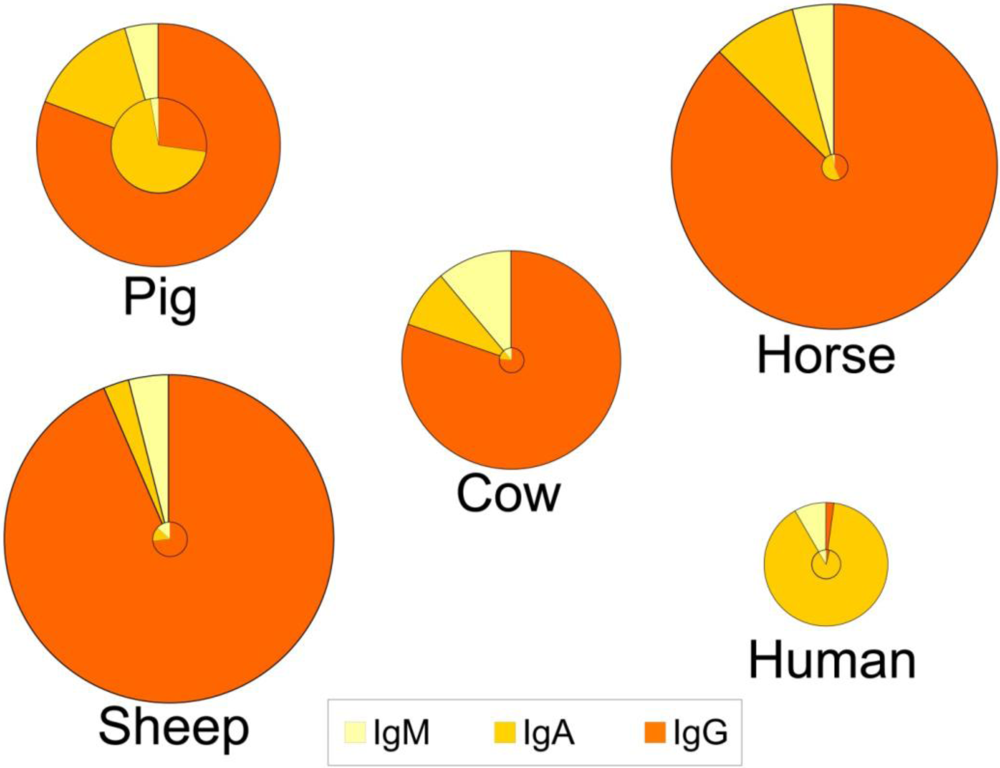
4. Concentrations in Colostrum and Milk-Physiological Conditions
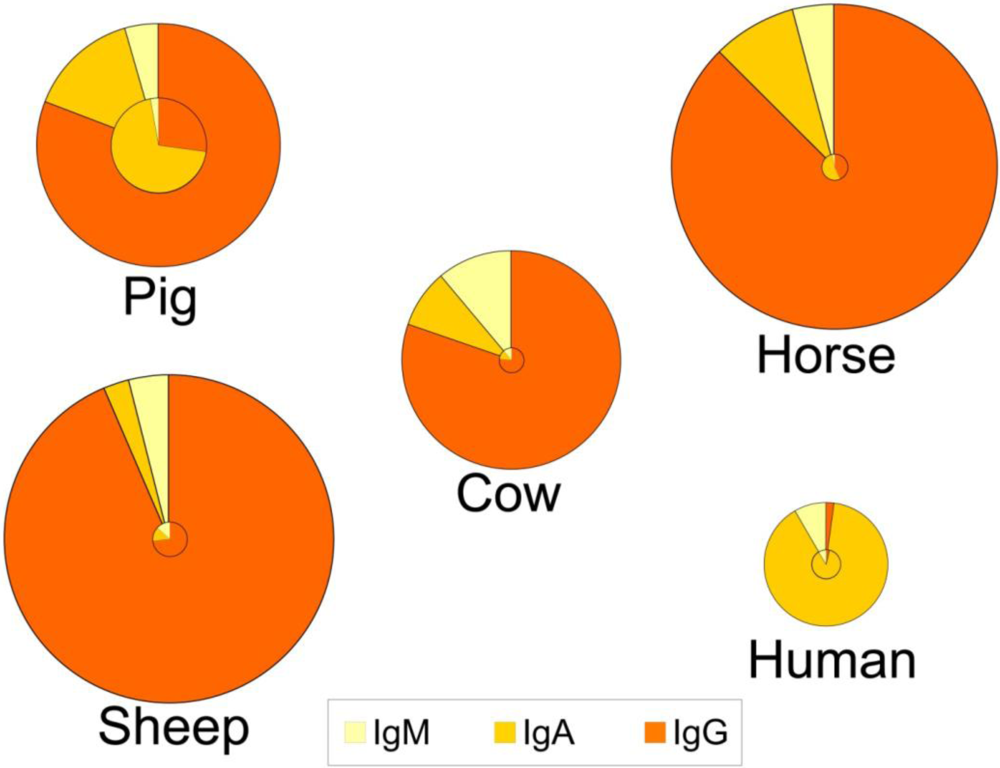
5. Sources and Transport of Immunoglobulins
5.1. IgG
5.2. Secretory IgA and IgM
6. Role of Immunoglobulins in the Intestine
6.1. Uptake of Immunoglobulins
6.2. Immunoglobulin-Intestinal Interactions
6.3. Intestinal Actions of Colostrum and Milk Immunoglobulins
7. Immunoglobulin Isolation and Stability
7.1. Overview
7.2. Isolation of Immunoglobulins from Mammary Secretions
7.3. Effects of Digestive Enzymes
7.4. Effects of pH
7.5. Effects of Heat Treatment
7.6. Heating Colostrum
8. Conclusions
References
- Butler, J.E.; Kehrli, M.E., Jr. Immunoglobulins and immunocytes in the mammary gland and its secretions. In Mucosal Immunology, 3rd; Mestecky , J., Lamm, M., Strober, W., Bienenstock, J., McGhee , J.R., Mayer, L., Eds.; Elsevier Academic Press: Burlington, MA, USA, 2005. [Google Scholar]
- Wheeler, T.T.; Hodgkinson, A.J.; Prosser, C.G.; Davis, S.R. Immune components of colostrum and milk-A historical perspective. J. Mammary Gland Biol. Neoplasia 2007, 12, 237–247. [Google Scholar]
- Butler, J.E. Immunoglobulins of the mammary secretions. In Lactation: A Comprehensive Treatise; Larson, B.L., Smith, V.R., Eds.; Academic Press: New York, NY, USA, 1974. [Google Scholar]
- McClelland, D.B.L. Antibodies in milk. J. Reprod. Fertil. 1982, 65, 537–543. [Google Scholar]
- Butler, J.E. Bovine immunoglobulins: An augmented review. Vet. Immunol. Immunopathol. 1983, 4, 43–152. [Google Scholar]
- Chernishov, V.P.; Slukvin, I.I. Mucosal immunity of the mammary gland and immunology of mother/newborn interrelation. Arch. Immunol. Ther. Exp. (Warsz.) 1990, 38, 145–164. [Google Scholar] [PubMed]
- Larson, B.L. Immunoglobulins of the mammary secretions. In Advanced Dairy Chemistry: Proteins, 2nd; Fox, P.F., Ed.; Elsevier Applied Science: London, UK, 1992. [Google Scholar]
- Telemo, E.; Hanson, L.A. Antibodies in milk. J. Mammary Gland Biol. Neoplasia 1996, 1, 243–249. [Google Scholar]
- Korhonen, H.; Marnila, P.; Gill, H.S. Milk immunoglobulins and complement factors. Br. J. Nutr. 2000, 84 (Suppl. 1), S75–S80. [Google Scholar] [PubMed]
- Hanson, L.Å.; Silfverdal, S.-A.; Stromback, L.; Erling, V.; Zaman, S.; Olcen, P.; Telemo, E. The immunological role of breast feeding. Pediatr. Allergy Immunol. 2001, 12 (Suppl. 14), S15–S19. [Google Scholar]
- Lilius, E.-M.; Marnila, P. The role of colostral antibodies in prevention of microbial infections. Curr. Opin. Infect. Dis. 2001, 14, 295–300. [Google Scholar]
- Tizard, I. The protective properties of milk and colostrum in non-human species. In Advances in Nutritional Research: Immunological Properties of Milk; Woodward, B., Draper, H.H., Eds.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2001. [Google Scholar]
- Uruakpa, F.O.; Ismond, M.A.H.; Akobundu, E.N.T. Colostrum and its benefits: A review. Nutr. Rev. 2002, 22, 755–767. [Google Scholar]
- Hurley, W.L. Immunoglobulins of the mammary secretions. In Advanced Dairy Chemistry: Proteins, 3rd; Fox, P.F., McSweeney, P.L.H., Eds.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2003. [Google Scholar]
- Van de Perre, P. Transfer of antibody via mother’s milk. Vaccine 2003, 21, 3374–3376. [Google Scholar]
- Gapper, L.W.; Copstake, D.E.J.; Otter, D.E.; Indyk, H.E. Analysis of bovine immunoglobulin G in milk, colostrum and dietary supplements: A review. Anal. Bioanal. Chem. 2007, 389, 93–109. [Google Scholar]
- Stelwagen, K.; Carpenter, E.; Haugh, B.; Hodgkinson, A.; Wheeler, T.T. Immune components of bovine colostrum and milk. J. Anim. Sci. 2009, 87, 3–9. [Google Scholar]
- Brandtzaeg, P. The mucosal immune system and its integration with the mammary glands. J. Pediatr. 2010, 156, S8–S15. [Google Scholar]
- Campbell, B.; Petersen, W.E. Immune milk-A historical survey. Dairy Sci. Abstr. 1963, 25, 345–358. [Google Scholar]
- Lascelles, A.K. A review of the literature on some aspects of immune milk. Dairy Sci. Abstr. 1963, 25, 359–364. [Google Scholar]
- Korhonen, H.; Marnila, P.; Gill, H.S. Bovine milk antibodies for health. Br. J. Nutr. 2000, 84 (Suppl. 1), S135–S146. [Google Scholar] [PubMed]
- Struff, W.G.; Sprotte, G. ovine colostrum as a biologic in clinical medicine; a review-Part I: Biotechnological standards, pharmacodynamic and pharmacokinetic characteristics and principles of treatment. Int. J. Clin. Pharmacol. Ther. 2997, 45, 193–202. [Google Scholar]
- Struff, W.G.; Sprotte, G. Bovine colostrum as a biologic in clinical medicine; a review-Part II: Clinical studies. Int. J. Clin. Pharmacol. Ther. 2008, 46, 211–225. [Google Scholar] [PubMed]
- Alisky, J. Bovine and human-derived passive immunization could help slow a future avian influenza pandemic. Med. Hypotheses 2009, 72, 74–75. [Google Scholar]
- Levine, M.M. Vaccines and milk immunoglobulin concentrates for prevention of infectious diarrhea. J. Pediatr. 1991, 118, S129–S136. [Google Scholar]
- Facon, M.; Skura, B.J.; Nakai, S. Potential for immunological supplementation of foods. Food Agric. Immunol. 1993, 5, 85–91. [Google Scholar]
- Davidson, G.P. Passive protection against diarrhea disease. J. Pediatr. Gastroenterol. Nutr. 1996, 23, 207–212. [Google Scholar]
- Mestecky, J.; Russell, M.W. Passive and active protection against disorders of the gut. Vet. Q. 1998, 20, S83–S87. [Google Scholar]
- Weiner, C.; Pan, Q.; Hurtig, M.; Borén, T.; Bostwick, E.; Hammarstom, L. Passive immunity against human pathogens using bovine antibodies. Clin. Exp. Immunol. 1999, 116, 193–205. [Google Scholar]
- Zeitlin, L.; Cone, R.A.; Moench, T.R.; Whaley, K.J. Preventing infectious disease with passive immunization. Microbes Infect. 2000, 1, 701–708. [Google Scholar]
- Zinkernagel, R. Maternal antibodies, childhood infections, and autoimmune disease. N. Engl. J. Med. 2001, 345, 1331–1335. [Google Scholar]
- McFadden, T.B.; Besser, T.E.; Barrington, G.M. Regulation of immunoglobulin transfer into mammary secretions of ruminants. In Milk Composition, Production and Biotechnology; Welch, R.A.S., Burns, D.J.W., Davis, S.R., Popay, A.I., Prosser, C.G., Eds.; CAB International: New York, NY, USA, 1997; pp. 133–152. [Google Scholar]
- Bernhisel-Broadbent, J.; Yolken, R.H.; Sampson, H.A. Allergenicity of orally administered preparations in food-allergic children. Pediatrics 1991, 87, 208–214. [Google Scholar]
- Colker, C.M.; Swain, M.; Lynch, L.; Gingerich, D.A. Effects of a milk-based bioactive micronutrient beverage on pain symptoms and activity of adults with osteoarthritis: A double-blind, placebo-controlled clinical evaluation. Nutrition 2002, 18, 388–392. [Google Scholar]
- Gingerich, D.A.; McPhillips, C.A. Analytical approach to determination of safety of milk ingredients from hyperimmunized cows. Regul. Toxicol. Pharmacol. 2005, 41, 102–112. [Google Scholar]
- Glass, R.I.; Svennerholm, A.-M.; Stoll, B.J.; Khan, M.R.; Hossain, K.M.B.; Huq, M.I.; Holmgren, J. Protection against cholera in breast-fed children by antibodies in breast milk. N. Engl. J. Med. 1983, 398, 1389–1392. [Google Scholar]
- Ruiz-Palacios, G.M.; Calva, J.J.; Pickering, L.K.; Lopez-Vidal, Y.; Volkow, P.; Pezzarossi, H.; West, M.S. Protection of breast-fed infants against Campylobacter diarrhea by antibodies in human milk. J. Pediatr. 1990, 116, 707–713. [Google Scholar]
- Moon, H.W.; Bunn, T.O. Vaccines for preventing enterotoxigenic Escherichia coli infections in farm animals. Vaccine 1993, 11, 213–220. [Google Scholar]
- Wilson, M.R.; Brown, P.; Svendsen, J. Immunity to Escherichia coli in pigs: Antibody secretion by the mammary gland after intramammary or intramuscular vaccination with E. coli vaccine. Can. J. Comp. Med. 1972, 36, 44–48. [Google Scholar]
- Kortbeek-Jacobs, J.M.C.; van Kooten, P.J.S.; van der Donk, J.A.; van Dijk, J.E.; Rutten, V.P. The effect of oral immunization on the population of lymphocytes migrating to the mammary gland of the sow. Vet. Microbiol. 1984, 9, 287–299. [Google Scholar]
- Saif, L.J.; Smith, K.L.; Landmeier, B.J.; Bohl, E.H.; Theil, K.W.; Todhunter, D.A. Immune response of pregnant cows to bovine rotavirus immunization. Am. J. Vet. Res. 1984, 45, 49–58. [Google Scholar]
- Lanza, I.; Shoup, D.I.; Saif, L.J. Lactogenic immunity and milk antibody isotypes to transmissible gastroenteritis virus in sows exposed to porcine respiratory coronavirus during pregnancy. Am. J. Vet. Res. 1995, 56, 739–748. [Google Scholar]
- Parreño, V.; Béjar, C.; Vagnozzi, A.; Barrandeguy, M.; Costantini, V.; Craig, M.I.; Yuan, L.; Hodgins, D.; Saif, L.; Ferñandez, F. Modulation by colostrum-acquired maternal antibodies of systemic and mucosal antibody responses to rotavirus in calves experimentally challenged with bovine rotavirus. Vet. Immunol. Immunopathol. 2004, 100, 7–24. [Google Scholar]
- Parreño, V.; Marcoppido, G.; Vega, C.; Garaicoechea, L.; Rodriguez, D.; Saif, L.; Ferñandez, F. Milk supplemented with immune colostrum: Protection against rotavirus diarrhea and modulatory effect on the systemic and mucosal antibody responses in calves experimentally challenged with bovine rotavirus. Vet. Immunol. Immunopathol. 2010, 136, 12–27. [Google Scholar]
- Snodgrass, D.R.; Campbell, I.; Mwenda, J.M.; Chege, G.; Suleman, M.A.; Morein, B.; Hart, C.A. Stimulation of rotavirus IgA, IgG and neutralizing antibodies in baboon milk by parenteral vaccination. Vaccine 1995, 13, 408–413. [Google Scholar]
- Shahid, N.S.; Steinhoff, M.C.; Roy, E.; Begum, T.; Thompson, C.M.; Siber, G.R. Placental and breast transfer of antibodies after maternal immunization with polysaccharide meningococcal vaccine: A randomized, controlled evaluation. Vaccine 2002, 20, 2404–2409. [Google Scholar]
- Schaller, J.P.; Saif, L.J.; Cordle, C.T.; Candler, E., Jr.; Winship, T.R.; Smith, K.L. Prevention of human rotavirus-induced diarrhea in gnotobiotic piglets using bovine antibody. J. Infect. Dis. 1992, 165, 623–630. [Google Scholar]
- Jenkins, M.C.; O’Brien, C.; Trout, J.; Guidry, A.; Fayer, R. Hyperimmune bovine colostrum specific for recombinant Cryptosporidium parvum antigen confers partial protection against cryptosporidiosis in immunosuppressed adult mice. Vaccine 1999, 17, 2453–2460. [Google Scholar]
- Huang, X.H.; Chen, L.; Gao, W.; Zhang, W.; Chen, S.J.; Xu, L.B.; Zhang, S.Q. Specific IgG activity of bovine immune milk against diarrhea bacteria and its protective effects on pathogen-infected intestinal damages. Vaccine 2008, 26, 5973–5980. [Google Scholar]
- Stolle, R.J.; Beck, L.R. Prevention and treatment of rheumatoid arthritis. U.S. Patent 4,732,757,22, March 1988. [Google Scholar]
- Golay, A.; Ferrara, J.-M.; Felber, J.-P.; Schneider, H. Cholesterol-lowering effect of skim milk from immunized cows in hypercholesterolemic patients. Am. J. Clin. Nutr. 1990, 52, 1014–1019. [Google Scholar]
- Sharpe, S.J.; Gamble, G.D.; Sharpe, D.N. Cholesterol-lowering and blood pressure effects of immune milk. Am. J. Clin. Nutr. 1994, 59, 929–934. [Google Scholar]
- Tai, Y.S.; Liu, B.Y.; Wang, J.T.; Sun, A.; Kwan, H.W.; Chiang, C.P. Oral administration of milk from cows immunized with human intestinal bacteria leads to significant improvements of symptoms and signs in patients with oral submucous fibrosis. J. Oral Pathol. Med. 2001, 30, 618–625. [Google Scholar]
- Yolken, R.H.; Losonsky, G.A.; Vonderfecht, S.; Leister, F.; Wee, S.-B. Antibody to human rotavirus in cow’s milk. N. Engl. J. Med. 1985, 312, 605–610. [Google Scholar]
- Li-Chan, E.; Kummer, A.; Losso, J.N.; Nakai, S. Survey of immunoglobulin G content and antibody specificity in cow’s milk from British Columbia. Food Agric. Immunol. 1994, 6, 443–451. [Google Scholar]
- Ebina, T.; Sato, A.; Umezu, K.; Ishida, N.; Ohyama, S.; Oizumi, A.; Aikawa, K.; Katagiri, S.; Katsushima, N.; Imai, A.; Kitaoka, S.; Suzuki, H.; Konno, T. Prevention of rotavirus infection by cow colostrum containing antibody against human rotavirus. Lancet 1983, 322, 1029–1030. [Google Scholar]
- Ebina, T.; Sato, A.; Umezu, K.; Ishida, N.; Ohyama, S.; Oizumi, A.; Aikawa, K.; Katagiri, S.; Katsushima, N.; Imai, A.; Kitaoka, S.; Suzuki, H.; Konno, T. Prevention of rotavirus infection by oral administration of cow colostrum containing antihumanrotavirus antibody. Med. Microbiol. Immunol. 1985, 174, 177–185. [Google Scholar]
- Hilpert, H.; Brussow, H.; Mietens, C.; Sidoti, J.; Lerner, L.; Werchau, H. Use of bovine milk concentrate containing antibody to rotavirus to treat rotavirus gastroenteritis in infants. J. Infect. Dis. 1987, 156, 158–166. [Google Scholar]
- Mitra, A.K.; Mahalanabis, D.; Ashraf, H.; Unicomb, L.; Eeckels, R.; Tzipori, S. Hyperimmune cow colostrum reduces diarrhoea due to rotavirus: A double-blind, controlled clinical trial. Acta Paediatr. 1995, 84, 996–1001. [Google Scholar]
- Davidson, G.P.; Whyte, P.B.D.; Daniels, E.; Franklin, K.; Nunan, H.; McCloud, P.I.; Moore, A.G.; Moore, D.J. Passive immunization of children with bovine colostrum containing antibodies to human rotavirus. Lancet 1989, 334, 709–712. [Google Scholar]
- Mietens, C.; Keinhorst, H.; Hilpert, H.; Gerber, H.; Amster, H.; Pahud, J.J. Treatment of infantile E. coli gastroenteritis with specific bovine anti-E. coli milk immunoglobulins. Eur. J. Pediatr. 1979, 132, 239–252. [Google Scholar] [PubMed]
- Tacket, C.O.; Binion, S.B.; Bostwick, E.; Losonsky, G.; Roy, M.J.; Edelman, R. Efficacy of bovine milk immunoglobulin concentrate in preventing illness after Shigella flexneri challenge. Am. J. Trop. Med. Hyg. 1992, 47, 276–283. [Google Scholar]
- Ashraf, H.; Mahalanabis, D.; Mitra, A.K.; Tzipori, S.; Fuchs, G.J. Hyperimmune bovine colostrum in the treatment of shigellosis in children: A double-blind, randomized, controlled tria. Acta Paediatr. 2001, 90, 1373–1378. [Google Scholar]
- Tacket, C.O.; Losonsky, G.; Link, H.; Hoang, Y.; Guesry, P.; Hilpert, H.; Levine, M.M. Protection by milk immunoglobulin concentrate against oral challenge with enterotoxigenic Escherichia coli. N. Engl. J. Med. 1988, 318, 1240–1243. [Google Scholar]
- Freedman, D.J.; Tacket, C.O.; Delehanty, A.; Maneval, D.R.; Nataro, J.; Crabb, J.H. Milk immunoglobulin with specific activity against purified colonization factor antigens can protect against oral challenge with enterotoxigenic Escherichia coli. J. Infect. Dis. 1998, 177, 662–667. [Google Scholar]
- Tacket, C.O.; Losonsky, G.; Livio, S.; Edelman, R.; Crabb, J.; Freedman, D. Lack of prophylactic efficacy of an enteric-coated bovine hyperimmune milk product against enterotoxigenic Escherichia coli challenge administered during a standard meal. J. Infect. Dis. 1999, 180, 2056–2059. [Google Scholar]
- Huppertz, H.-I.; Rutkowski, S.; Busch, D.; Eisebit, R.; Lissner, R.; Karch, H. Bovine colostrum ameliorates diarrhea in infection with diarrheagenic Escherichia coli, Shiga toxin-producing E.coli, and E. coli expressing intimin and hemolysin. J. Pediatr. Gastroenterol. Nutr. 1999, 29, 452–456. [Google Scholar] [CrossRef] [PubMed]
- Stephan, W.; Dichtelmuller, H.; Lissner, R. Antibodies from colostrum in oral immunotherapy. J. Clin. Chem. Clin. Biochem. 1990, 28, 19–23. [Google Scholar]
- Koga, T.; Oho, T.; Shimazaki, Y.; Nakano, Y. Immunization against dental caries. Vaccine 2002, 20, 2027–2044. [Google Scholar]
- Michalek, S.M.; McGhee, J.R. Effective immunity to dental caries: Passive transfer to rats of antibodies to Streptococcus mutans elicits protection. Infect. Immun. 1977, 17, 644–650. [Google Scholar]
- Loimaranta, V.; Carlén, A.; Olsson, J.; Tenovuo, J.; Syvaoja, E.-L.; Korhonen, H. Concentrated bovine colostral whey proteins from Streptococcus mutans/Strep. sorbinus immunized cows inhibit the adherence of Strep. mutans and promote the aggregation of mutans streptococci. J. Dairy Res. 1998, 65, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Loimaranta, V.; Tenovuo, J.; Virtanen, S.; Marnila, P.; Syvaoja, E.-L.; Tupasela, T.; Korhonen, H. Generation of bovine immune colostrum against Streptococcus mutans and Streptococcus sobrinus and its effect on glucose uptake and extracellular polysaccharide formation by mutans streptococci. Vaccine 1997, 15, 1261–1268. [Google Scholar] [CrossRef] [PubMed]
- Loimaranta, V.; Tenovuo, J.; Korhonen, H. Combined inhibitory effect of bovine immune whey and peroxidase-generated hypothiocyanite against glucose uptake by Streptococcus mutans. Oral Microbiol. Immunol. 1998, 13, 378–381. [Google Scholar]
- Loimaranta, V.; Laine, M.; Soderling, E.; Vasara, E.; Rokka, S.; Marnila, P.; Tossavainen, O.; Tenovuo, J. Effects of bovine immune and non-immune whey preparations on the composition and pH response of human dental plaque. Eur. J. Oral Sci. 1999, 107, 244–250. [Google Scholar]
- Wei, H.; Loimaranta, V.; Tenovuo, J.; Rokka, S.; Syvaoja, E.-L.; Korhonen, H.; Joutsjoki, V.; Marnila, P. Stability and activity of specific antibodies against Streptococcus mutans and Streptococcus sobrinus in bovine milk fermented with Lactobacillus rhamnosus strain GG or treated at ultra-high temperature. Oral Microbiol. Immunol. 2002, 17, 9–15. [Google Scholar]
- Filler, S.J.; Gregory, R.L.; Michalek, S.M.; Katz, J.; McGhee, J.R. Effect of immune bovine milk on Streptococcus mutans in human dental plaque. Archs. Oral Biol. 1991, 36, 41–47. [Google Scholar]
- Oho, T.; Shimazaki, Y.; Mitoma, M.; Yoshimura, M.; Yamashita, Y.; Okano, K.; Nakano, Y.; Kawagoe, H.; Fukuyama, M.; Fujihara, N.; Koga, T. Bovine milk antibodies against cell surface protein antigen PAc-glucosyltransferase fusion protein suppress cell adhesion and alter glucan synthesis of Streptococcus mutans. J. Nutr. 1999, 129, 1836–1841. [Google Scholar]
- Shimazaki, Y.; Mitoma, M.; Oho, T.; Nakano, Y.; Yamashita, Y.; Okano, K.; Nakano, Y.; Fukuyama, M.; Fujihara, N.; Nada, Y.; Koga, T. Passive immunization with milk produced from an immunized cow prevents oral recolonization by Streptococcus mutans. Clin. Diagn. Lab. Immunol. 2001, 8, 1136–1139. [Google Scholar]
- Tzipori, S.; Roberton, D.; Chapman, C. Remission of diarrhoea due to cryptosporidiosis in an immunodeficient child treated with hyperimmune bovine colostrum. Br. Med. J. 1986, 293, 1276–1277. [Google Scholar]
- Shield, J.; Melville, C.; Novelli, V.; Anderson, G.; Scheimberg, I.; Gibb, D.; Milla, P. Bovine colostrum immunoglobulin concentrate for cryptosporidiosis in AIDS. Arch. Dis. Child. 1993, 69, 451–453. [Google Scholar]
- Hurley, W.L. Mammary gland function during involution. J. Dairy Sci. 1989, 72, 1637–1646. [Google Scholar]
- Guidry, A.J.; Butler, J.E.; Pearson, R.E.; Weiland, B. IgA, IgG1, IgG2, IgM and BSA secretion by the bovine mammary gland throughout lactati. Vet. Immunol. Immunopathol. 1980, 1, 329–341. [Google Scholar]
- Barkema, H.W.; Green, M.J.; Bradley, A.J.; Zadoks, R.N. Invited review: The role of contagious disease in udder health. J. Dairy Sci. 2009, 92, 4717–4729. [Google Scholar]
- Rinaldi, M.; Li, R.W.; Capuco, A.V. Mastitis associate transcriptomic disruptions in cattle. Vet. Immunol. Immunopathol. 2010, 138, 267–279. [Google Scholar]
- Colditz, I.G.; Watson, D.L. The immunophysiological basis for vaccinating ruminants against mastitis. Aust. Vet. J. 1985, 62, 145–153. [Google Scholar]
- Denis, M.; Wedlock, D.N.; Lacy-Hulbert, S.J.; Hillerton, J.E.; Buddle, B.M. Vaccines against bovine mastitis in the New Zealand context: What is the best way forward? N. Z. Vet. J. 2009, 57, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Erskine, R.J.; Brockett, A.R.; Beeching, N.D.; Hull, R.W.; Bartlett, P.C. Effect of changes in number of doses and anatomic location for administration of an Escherichia coli bacterin on serum IgG1 and IgG2 concentrations in dairy cows. Am. J. Vet. Res. 2010, 71, 120–124. [Google Scholar]
- Dosogne, H.; Vangroenweghe, F.; Burvenich, C. Potential mechanism of action of J5 vaccine in protection against bovine coliform mastitis. Vet. Res. 2002, 33, 1–12. [Google Scholar]
- Mallard, B.A.; Wagter, L.C.; Ireland, M.J.; Dekkers, J.C.M. Effects of growth hormone, insulin-like growth factor-I, and cortisol on periparturient antibody response profiles of dairy cattl. Vet. Immunol. Immunopathol. 1997, 60, 61–76. [Google Scholar]
- Hogan, J.; Smith, K.L. Coliform mastitis. Vet. Res. 2003, 34, 507–519. [Google Scholar]
- Hogan, J.S.; Weiss, W.P.; Smith, K.L.; Todhunter, D.A.; Schoenberger, P.S.; Sordillo, L.M. Effects of an Escherichia coli J5 vaccine on mild clinical coliform mastitis. J. Dairy Sci. 1995, 78, 285–290. [Google Scholar]
- Hogan, J.S.; Bogacz, V.L.; Aslam, M.; Smith, K.L. Efficacy of an Escherichia coli J5 bacterin administered to primigravid heifers. J. Dairy Sci. 1999, 82, 939–943. [Google Scholar]
- Hogan, J.S.; Cannon, V.B.; Smith, K.L.; Rinehart, C.; Miller, S. Effects of adjuvants on safety and efficacy of an Escherichia coli J5 bacterin. J. Dairy Sci. 2005, 88, 534–542. [Google Scholar]
- Erskine, R.J.; vanDyk, E.J.; Bartlett, P.C.; Burton, J.L.; Boyle, M.C. Effect of hyperimmunization with an Escherichia coli J5 bacterin in adult lactating dairy cows. J. Am. Vet. Med. Assoc. 2007, 231, 1092–1097. [Google Scholar]
- Wilson, D.J.; Mallard, B.A.; Burton, J.L.; Schukken, Y.H.; Grohn, Y.T. Milk and serum J5-specific antibody responses, milk production change, and clinical effects following intramammary Escherichia coli challenge for J5 vaccinate and control cow. Clin. Vaccine Immunol. 2007, 14, 693–699. [Google Scholar]
- Wilson, D.J.; Mallard, B.A.; Burton, J.L.; Schukken, Y.H.; Grohn, Y.T. Association of Escherichia coli J5-specific serum antibody responses with clinical mastitis outcome for J5 vaccinate and control dairy cattle. Clin. Vaccine Immunol. 2009, 16, 209–217. [Google Scholar]
- Prenafeta, A.; March, R.; Foix, A.; Casals, I.; Costa, L. Study of the humoral immunological response after vaccination with a Staphylococcus aureus biofilm-embedded bacterin in dairy cows: Possible role of the exopolysaccharide specific antibody production in the protection from Staphylococcus aureus induced mastitis. Vet. Immunol. Immunopathol. 2010, 134, 208–217. [Google Scholar]
- Middleton, J.R.; Luby, C.D.; Adams, D.S. Efficacy of vaccination against staphylococcal mastitis: A review and new data. Vet. Microbiol. 2009, 134, 192–198. [Google Scholar]
- Boothby, J.T.; Schore, C.E.; Jasper, D.E.; Osburn, B.I.; Thomas, C.B. Immune responses to Mycoplasma bovis vaccination and experimental infection in the bovine mammary gland. Can. J. Vet. Res. 1988, 52, 355–359. [Google Scholar]
- Chang, C.C.; Winter, A.J.; Norcross, N.L. Immune response in the bovine mammary gland after intestinal, local, and systemic immunizatio. Infect. Immun. 1981, 31, 650–659. [Google Scholar]
- Sheldrake, R.F.; Husband, A.J.; Watson, D.L.; Cripps, A.W. The effect of intraperitoneal and intramammary immunization of sheep on the numbers of antibody-containing cells in the mammary gland, and antibody titres in blood serum and mammary secretions. Immunology 1985, 56, 605–614. [Google Scholar]
- Finch, J.M.; Hill, A.W.; Field, T.R.; Leigh, J.A. Local vaccination with killed Streptococcus uberis protects bovine mammary gland against experimental intramammary challenge with the homologous strain. Infect. Immun. 1994, 62, 3599–3603. [Google Scholar]
- Takemura, K.; Hogan, J.S.; Smith, K.L. Efficacy of immunization with ferric citrate receptor FecA from Escherichia coli on induced coliform mastitis. J. Dairy Sci. 2002, 85, 774–781. [Google Scholar]
- Kelly, C.P.; Pothoulakis, C.; Vavva, F.; Castagliuolo, I.; Bostwick, E.F.; O’Keane, J.C.; Keates, S.; LaMont, J.T. Anti-Clostridium difficile bovine immunoglobulin concentrate inhibits cytotoxicity and enterotoxicity of C. difficile toxins. Antimicrob. Agents Chemother. 1996, 40, 373–379. [Google Scholar] [PubMed]
- Brüssow, H.; Hilpert, H.; Walther, I.; Sidoti, J.; Mietens, C.; Bachman, P. Bovine milk immunoglobulins for passive immunity to infantile rotavirus gastroenteritis. J. Clin. Microbiol. 1987, 25, 982–986. [Google Scholar]
- Hodgkinson, A.J.; Cannon, R.D.; Holmes, A.R.; Fischer, F.J.; Willix-Payne, D.J. Production from dairy cows of semi-industrial quantities of milk-protein concentrate (MPC) containing efficacious anti-Candida albicans IgA antibodies. J. Dairy Res. 2007, 74, 269–275. [Google Scholar]
- Beck, L.R. Method for treating inflammation using bovine milk. U.S. Patent 4,284,623,18, August 1981. [Google Scholar]
- Dorosko, S.M.; Ayres, S.L.; Connor, R.L. Induction of HIV-1 MPR649-684-specific IgA and IgG antibodies in caprine colostrum using a peptide-based vaccine. Vaccine 2008, 26, 5416–5422. [Google Scholar]
- Shkreta, L.; Talbot, B.G.; Diarra, M.S.; Lacasse, P. Immune responses to a DNA/protein vaccination strategy against Staphylococcus aureus induced mastitis in dairy cows. Vaccine 2004, 23, 114–126. [Google Scholar]
- Castagliuolo, I.; Piccinini, R.; Beggiao, E.; Palu, G.; Mengoli, C.; Ditadi, F.; Vicenzoni, G.; Zecconi, A. Mucosal genetic immunization against four adhesions protects against Staphylococcus aureus-induced mastitis in mice. Vaccine 2006, 24, 4393–4402. [Google Scholar]
- O’Brien, C.N.; Guidry, A.J.; Douglass, L.W.; Westhoff, D.C. Immunization with Staphylococcus aureus lysate incorporated into microspheres. J. Dairy Sci. 2001, 84, 1791–1799. [Google Scholar]
- Liu, G.L.; Wang, J.Q.; Bu, D.P.; Cheng, J.B.; Zhang, C.G.; Wei, H.Y.; Zhou, L.Y.; Liu, K.L.; Dong, X.L. Specific immune milk production of cows implanted with antigen-release devices. J. Dairy Sci. 2009, 92, 100–108. [Google Scholar]
- Toledo, J.R.; Sanchez, O.; Montesino, R.; Farnos, O.; Rodriguez, M.P.; Alfonso, P.; Oramas, N.; Rodriguez, E.; Santana, E.; Vega, E.; Ganges, L.; Frias, M.T.; Cremata, J.; Barrera, M. Highly protective E2-CSFV vaccine candidate produced in the mammary gland of adenoviral transduced goats. J. Biotechnol. 2008, 133, 370–376. [Google Scholar]
- Mix, E.; Goertsches, R.; Zettl, U.K. Immunoglobulins-basic considerations. J. Neurol. 2006, 253, 9–17. [Google Scholar]
- Woof, J.M. The structure of IgA. In Mucosal Immune Defense: Immunoglobulin A; Kaetzel, C.S., Ed.; Springer: New York, NY, USA, 2007. [Google Scholar]
- Johansen, F.E.; Braathen, R.; Brandtzaeg, P. Role of J chain in secretory immunoglobulin formation. Scand. J. Immunol. 2000, 52, 240–248. [Google Scholar]
- Rouse, B.T.; Ingram, D.G. The total protein and immunoglobulin profile of equine colostrum and milk. Immunology 1970, 19, 901–907. [Google Scholar]
- Lascelles, A.K. The immune system of the ruminant mammary gland and its role in the control of mastitis. J. Dairy Sci. 1979, 62, 154–160. [Google Scholar]
- Hunziker, W.; Kraehenbuhl, J.-P. Epithelial transcytosis of immunoglobulins. J. Mammary Gland Biol. Neoplasia 1998, 3, 287–302. [Google Scholar]
- Cianga, P.; Medesan, C.; Richardson, J.A.; Ghetie, V.; Ward, E.S. Identification and function of neonatal Fc receptor in mammary gland of lactating mice. Eur. J. Immunol. 1999, 29, 2515–2523. [Google Scholar]
- He, W.; Ladinsky, M.S.; Huey-Tubman, K.E.; Jensen, G.J.; McIntosh, R.; Bjorkman, P.J. FcRn-mediated antibody transport across epithelial cells revealed by electron tomography. Nature 2008, 455, 542–546. [Google Scholar]
- Cervenak, J.; Kacskovics, I. The neonatal Fc receptor plays a crucial role in the metabolism of IgG in livestock animals. Vet. Immunol. Immunopathol. 2009, 128, 171–177. [Google Scholar]
- Rodewald, R.; Kraehenbuhl, J.P. Receptor-mediated transport of IgG. J. Cell Biol. 1984, 99, 159s–164s. [Google Scholar]
- Simister, N.E.; Rees, A.R. Isolation and characterization of an Fc receptor from neonatal rat small intestine. Eur. J. Immunol. 1985, 15, 733–738. [Google Scholar]
- Simister, N.E.; Story, C.M. Human placental Fc receptors and the transmission of antibodies from mother to fetus. J. Reprod. Immunol. 1997, 37, 1–23. [Google Scholar]
- Simister, N.E. Placental transport of immunoglobulin G. Vaccine 2003, 21, 3365–3369. [Google Scholar]
- Pentsuk, N.; van der Laan, J.W. An interspecies comparison of placental antibody transfer: New insights into developmental toxicity testing of monoclonal antibodies. Birth Defects Res. B Dev. Reprod. Toxicol. 2009, 86, 328–344. [Google Scholar]
- Fuchs, R.; Ellinger, I. Endocytic and transcytotic processes in villus syncytiotrophoblast: Role in nutrient transport to the human fetus. Traffic 2004, 5, 725–738. [Google Scholar]
- Simister, N.E.; Mostov, K.E. An Fc receptor structurally related to MHC class I antigens. Nature (Lond.) 1989, 337, 184–187. [Google Scholar] [CrossRef]
- Junghans, R.P.; Anderson, C.L. The protection receptor for IgG catabolism is the β2-microglobulin-containing neonatal intestinal transport receptor. Proc. Natl. Acad. Sci. USA 1996, 93, 5512–5516. [Google Scholar]
- Junghans, R.P. Finally! The Brambell receptor (FcRB). Immunol. Res. 1997, 16, 29–57. [Google Scholar]
- Telleman, P.; Junghans, R.P. The role of the Brambell receptor (FcRB) in liver: Protection of endocytosed immunoglobulin G (IgG) from catabolism in hepatocytes rather than transport of IgG to bile. Immunology 2000, 100, 245–251. [Google Scholar]
- Bender, B.; Bodrogi, L.; Mayer, B.; Schneider, Z.; Zhao, Y.; Hammarstrom, L.; Eggen, I.; Bosze, Z. Position independent and copy-number-related expression of the neonatal Fc receptor α-chain in transgenic mice carrying a 102 kb genomic fragment. Transgenic Res. 2007, 16, 613–627. [Google Scholar]
- Lu, W.; Zhao, Z.; Zhao, Y.; Yu, S.; Zhao, Y.; Fan, B.; Kasckovics, I.; Hammarstrom, L.; Li, N. Over-expression of the bovine FcRn in the mammary gland results in increased IgG levels in both milk and serum of transgenic mice. Immunology 2007, 122, 401–408. [Google Scholar]
- Cervenak, J.; Bender, B.; Schneider, Z.; Magna, M.; Carstea, B.V.; Liliom, K.; Erdei, A.; Bosze, Z.; Kacskovics, I. Neonatal FcR overexpression boosts humoral immune responses in transgenic mice. J. Immunol. 2010, 186, 959–968. [Google Scholar]
- Mayer, B.; Zolnai, A.; Frenyo, L.V.; Jancsik, V.; Szentirmay, Z.; Hammarstrom, L.; Kasckovics, I. Redistribution of the sheep neonatal Fc receptor in the mammary gland around the time of parturition in ewes and its localization in the small intestine of neonatal lambs. Immunology 2002, 107, 288–296. [Google Scholar]
- Mayer, B.; Zolnai, A.; Frenyo, L.V.; Jancsik, V.; Szentirmay, Z.; Hammarstrom, L.; Kasckovics, I. Localization of the sheep FcRn in the mammary gland. Vet. Immunol. Immunopathol. 2002, 87, 327–330. [Google Scholar]
- Mayer, B.; Doleschall, M.; Bender, B.; Bartyik, J.; Bosze, Z.; Frenyo, L.V.; Kacskovics, I. Expression of the neonatal Fc receptor (FcRn) in the bovine mammary gland. J. Dairy Sci. 2005, 72, 107–112. [Google Scholar]
- Sayed-Ahmed, A.; Kassab, M.; Abd-Elmaksoud, A.; Elnasharty, M.; El-Kirdasy, A. Expression and immunohistochemical localization of the neonatal Fc receptor (FcRn) in the mammary gland of the Egyptian water buffalo. Acta Histochem. 2010, 112, 383–391. [Google Scholar]
- Zou, S.; Hurley, W.L.; Hegarty, H.M.; Larson, B.L.; Nelson, D.R. Immunohistological localization of IgG1, IgA and secretory component in the bovine mammary gland during involution. Cell Tissue Res. 1988, 251, 81–86. [Google Scholar]
- Laegreid, W.W.; Heaton, M.P.; Keen, J.E.; Grosse, W.M.; Chitko-McKnown, C.G.; Smith, T.P.L.; Keele, J.W.; Bennett, G.J.; Besser, T.E. Association of bovine neonatal Fc receptor α-chain gene (FCGRT) haplotypes with serum IgG concentration in newborn calves. Mamm. Genome 2002, 13, 704–710. [Google Scholar]
- Zhang, R.; Zhao, Z.; Zhao, Y.; Kacskovics, I.; van der Eijk, M.; de Groot, N. Association of FcRn heavy chain encoding gene (FCGRT) polymorphisms with IgG content in bovine colostrum. Anim. Biotechnol. 2009, 20, 242–246. [Google Scholar]
- Clawson, M.L.; Heaton, M.P.; Chitko-McKnown, C.G.; Fox, J.M.; Smith, T.P.L.; Snelling, W.M.; Keele, J.W.; Laegreid, W.W. Beta-2-microglobulin haplotypes in U.S. beef cattle and association with failure of passive transfer in newborn calves. Mamm. Genome 2004, 15, 228–236. [Google Scholar]
- Baumrucker, C.R.; Burkett, A.M.; Magliaro-Macrina, A.L.; Dechow, C.D. Colostrogenesis: Mass transfer of immunoglobulin G1 into colostrum. J. Dairy Sci. 2010, 93, 3031–3038. [Google Scholar]
- Sasaki, M.; Davis, C.L.; Larson, B.L. Production and turnover of IgG1 and IgG2 immunoglobulins in the bovine around parturition. J. Dairy Sci. 1976, 59, 2046–2055. [Google Scholar]
- Ishikawa, H.; Kanamori, Y.; Hamada, H.; Kiyono, H. Development and function of organized gut-associated lymphoid tissues. In Mucosal Immunology,3rd ed.; Mestecky, J., Lamm, M., Strober, W., Bienenstock, J., McGhee, J.R., Mayer, L., Eds.; Elsevier Academic Press: Burlington, MA, USA, 2005. [Google Scholar]
- Spenser, J.; Boursier, L.; Edgeworth, J.D. IgA plasma cell developmen. In Mucosal Immune Defense: Immunoglobulin A; Kaetzel, C.S., Ed.; Springer: New York, NY, USA, 2007; pp. 25–42. [Google Scholar]
- Brandtzaeg, P. Mucosal immunity: Integration between mother and the breast-fed infant. Vaccine 2003, 21, 3382–3388. [Google Scholar]
- Mostov, K.; Kaetzel, C.S. Immunoglobulin transport and the polymeric immunoglobulin receptor. In Mucosal Immunology, 2nd; Ogra, P.L., Mestecky, J., Lamm, M.E., Strober, W., Bienenstock, J., McGhee, J.R., Eds.; Academic Press: New York, NY, USA, 1999; pp. 181–211. [Google Scholar]
- Kaetzel, C.S.; Bruno, M.E.C. Epithelial transport of IgA by the polymeric immunoglobulin receptor. In Mucosal Immune Defense: Immunoglobulin A; Kaetzel, C.S., Ed.; Springer: New York, NY, USA, 2007; pp. 43–89. [Google Scholar]
- Johansen, F.E.; Braathen, R.; Brandtzaeg, P. The J chain is essential for polymeric Ig receptor-mediated epithelial transport of IgA. J. Immunol. 2001, 167, 5185–5192. [Google Scholar]
- Braathen, R.; Hohman, V.S.; Brandtzaeg, P.; Johansen, F.E. Secretory antibody formation: Conserved binding interactions between J chain and polymeric Ig receptor from humans and amphibians. J. Immunol. 2007, 178, 1589–1597. [Google Scholar]
- Rincheval-Arnold, A.; Belair, J.; Djiane, J. Developmental expression of pIgR gene in sheep mammary gland and hormonal regulation. J. Dairy Res. 2002, 69, 13–26. [Google Scholar]
- Blum, J.W. Nutritional physiology of neonatal calves. J. Anim. Physiol. Anim. Nutr. 2006, 90, 1–11. [Google Scholar]
- Staley, T.E.; Bush, L.J. Receptor mechanisms of the neonatal intestine and their relationship to immunoglobulin absorption and disease. J. Dairy Sci. 1985, 68, 184–205. [Google Scholar]
- Godden, S. Colostrum management for dairy calves. Vet. Clin. Food Anim. 2008, 24, 19–39. [Google Scholar]
- Husband, A.J.; Brandon, M.R.; Lascelles, A.K. Absorption and endogenous production of immunoglobulins in calves. Aust. J. Exp. Biol. Med. Sci. 1972, 50, 491–498. [Google Scholar]
- Brandtzaeg, P.; Johansen, F.E. IgA and intestinal homeostasis. In Mucosal Immune Defense: Immunoglobulin A; Kaetzel, C.S., Ed.; Springer: New York, NY, USA, 2007; pp. 221–268. [Google Scholar]
- Russell, M.W. Biological functions of IgA. In Mucosal Immune Defense: Immunoglobulin A; Kaetzel, C.S., Ed.; Springer: New York, NY, USA, 2007; pp. 144–172. [Google Scholar]
- Hanson, L.A.; Korotkova, M.; Telemo, E. Human milk: Its components and their immunobiologic functions. In Mucosal Immunology, 3rd; Mestecky, J., Bienenstock, J., Lamm, M.E., Mayer, L., McGhee, J.R., Strober, W., Eds.; Elsevier Academic Press: Amsterdam, The Netherlands, 2005; pp. 1795–1827. [Google Scholar]
- Sait, L.C.; Galic, M.; Price, J.D.; Simpfendorfer, K.R.; Diavatopoulos, D.A.; Uren, T.K.; Wijburg, O.L.C.; Strugnell, R.A. Secretory antibodies reduce systemic antibody responses against the gastrointestinal commensal flora. Int. Immunol. 2007, 19, 257–265. [Google Scholar]
- Barrington, G.M.; Parish, S.M. Bovine neonatal immunology. Vet. Clin. North Am. Food Anim. Pract. 2001, 17, 463–475. [Google Scholar]
- Gill, H. Dairy products and the immune function in the elderly. In Functional Dairy Products; Mattila-Sandholm, T., Saarela, M., Eds.; CRC Press: New York, NY, USA, 2003; pp. 133–168. [Google Scholar]
- Lonnerdal, B. Nutritional and physiological significance of human milk proteins. Am. J. Clin. Nutr. 2003, 77, 1537S–1543S. [Google Scholar]
- Pakkanen, R.; Aalto, J. Growth factors and antimicrobial factors of bovine colostrum. Int. Dairy J. 1997, 7, 285–297. [Google Scholar]
- Playford, R.J.; Macdonald, C.E.; Johnson, W.S. Colostrum and milk-derived peptide growth factors for the treatment of gastrointestinal disorders. Am. J. Clin. Nutr. 2000, 72, 5–14. [Google Scholar]
- Newburg, D.S.; Walker, W.A. Protection of the neonate by the innate immune system of developing gut and of human milk. Pediatr. Res. 2007, 61, 2–8. [Google Scholar]
- Mehta, R.; Petrova, A. Biologically active breast milk proteins in association with very preterm delivery and stage of lactation. J. Perinatol. 2010, 31, 58–62. [Google Scholar]
- Siccardi, D.; Turner, J.R.; Mrsny, R.J. Regulation of intestinal epithelial function: A link between opportunities for macromolecular drug delivery and inflammatory bowel disease. Adv. Drug Deliv. Rev. 2005, 57, 219–235. [Google Scholar]
- Bikker, P.; Kranendonk, G.; Gerritsen, R.; Russell, L.; Campbell, J.; Crenshaw, J.; Rodriguez, C.; Rodenas, J.; Polo, J. Absorption of orally supplied immunoglobulins in neonatal piglets. Livest. Sci. 2010, 134, 139–142. [Google Scholar]
- Weiner, M.L. Intestinal transport of some macromolecules in food. Food Chem. Toxicol. 1988, 26, 867–880. [Google Scholar]
- Hardin, J.A.; Kimm, M.H.; Wirasinghe, M.; Gall, D.G. Macromolecular transport across the rabbit proximal and distal colon. Gut 1999, 44, 218–225. [Google Scholar]
- Davids, B.J.; Palm, J.E.D.; Housley, M.P.; Smith, J.R.; Anderson, Y.S.; Martin, M.G.; Hendrickson, B.A.; Johansen, F.-E.; Svard, S.G.; Gillin, F.D.; Eckmann, L. Polymeric immunoglobulin receptor in intestinal immune defense against the lumen-dwelling protozoan parasite Giardia. J. Immunol. 2006, 177, 6281–6290. [Google Scholar]
- Fernandez, M.I.; Pedron, T.; Tournebize, R.; Olivo-Marin, J.-C.; Sansonetti, P.J.; Phalipon, A. Anti-inflammatory role for intracellular dimeric immunoglobulin A by neutralization of lipopolysaccharide in epithelial cells. Immunity 2003, 18, 739–749. [Google Scholar]
- Israel, E.J.; Taylor, S.; Wu, Z.; Mizoguchi, E.; Blumberg, R.S.; Bhan, A.; Simister, E. Expression of the neonatal Fc receptor, FcRn, on human intestinal epithelial cell. Immunology 1997, 92, 69–74. [Google Scholar]
- Dickinson, B.L.; Badizadegan, K.; Wu, Z.; Ahouse, J.C.; Zhu, X.; Simister, N.E.; Blumberg, R.S.; Lencer, W.I. Bidirectional FcRn-dependent IgG transport in a polarized human intestinal epithelial cell line. J. Clin. Invest. 1999, 104, 903–911. [Google Scholar]
- Rojas, R.; Apodaca, G. Immunoglobulin transport across polarized epithelia cells. Nat. Rev. Mol. Cell Biol. 2002, 3, 1–12. [Google Scholar]
- Yoshida, M.; Claypool, S.M.; Wagner, J.S.; Mizoguchi, E.; Mizoguchi, A.; Roopenian, D.C.; Lencer, W.I.; Blumberg, R.S. Human neonatal Fc receptor mediates transport of IgG into luminal secretions for delivery of antigens to mucosal dendritic cells. Immunity 2004, 20, 769–783. [Google Scholar]
- Guarner, F.; Malagelada, J.-R. Gut flora in health and disease. Lancet 2003, 361, 512–519. [Google Scholar]
- Kobayashi, K.; Blaser, M.J.; Brown, W.R. Identification of a unique IgG Fc binding site in human intestinal epithelium. J. Immunol. 1989, 143, 2567–2574. [Google Scholar]
- Harada, N.; Iijima, S.; Kobayashi, K.; Yoshida, T.; Brown, W.R.; Hibi, T.; Oshima, A.; Morikawa, M. Human IgGFc binding protein (FcγBP) in colonic epithelial cells exhibits mucin-like structure. J. Biol. Chem. 1997, 272, 15232–15241. [Google Scholar]
- Kobayashi, K.; Ogata, H.; Morikawa, M.; Iijima, S.; Harada, N.; Yoshida, T.; Brown, W.R.; Inoue, N.; Hamada, Y.; Ishii, H.; Watanabe, M.; Hibi, T. Distribution and partial characterization of IgG Fc binding protein in various mucin producing cells and body fluids. Gut 2002, 51, 169–176. [Google Scholar]
- Siegrist, C.-A. Mechanisms by which maternal antibodies influence infant vaccine responses: Review of hypotheses and definition of main determinants. Vaccine 2003, 21, 3406–3412. [Google Scholar]
- Milstein, C.P.; Feinstein, A. Comparative studies of two types of bovine immunoglobulin G heavy chains. Biochem. J. 1968, 107, 559–564. [Google Scholar]
- Porto, A.C.R.C.; Oliveira, L.L.; Ferraz, L.C.; Ferraz, L.E.S.; Thomaz, S.M.O.; Rosa, J.C.; Roque-Barreira, M.C. Isolation of bovine immunoglobulins resistant to peptic digestion: New perspectives in the prevention of failure in passive immunization of neonatal calves. J. Dairy Sci. 2007, 90, 955–962. [Google Scholar]
- Zettlitz, K.A. Protein A/G chromatography. In Antibody Engineering; Kontermann, R., Dübel, S., Eds.; Springer-Verlag: Berlin, Germany, 2010. [Google Scholar]
- Darcy, E.; Leonard, P.; Fitzgerald, J.; Danaher, M.; O’Kennedy, R. Purification of antibodies using affinity chromatography. Methods Mol. Biol. 2011, 681, 369–382. [Google Scholar]
- Ma, Z.; Lan, Z.; Matsuura, T.; Ramakrishna, S. Electrospun polyethersulfone affinity membrane: Membrane preparation and performance evaluation. J. Chromatogr. B 2009, 877, 3686–3694. [Google Scholar]
- Kaneko, T.; Wu, B.T.; Nakai, S. Selective concentration of bovine immunoglobulins and α-lactalbumin from acid whey using FeCl3. J. Food Sci. 1985, 50, 1531–1536. [Google Scholar]
- Al-Mashikhi, S.A.; Nakai, S. Separation of immunoglobulin and transferring from blood serum and plasma by metal chelate interaction chromatography. J. Dairy Sci. 1988, 71, 1756–1763. [Google Scholar]
- Carrillo-Conde, B.; Garza, A.; Anderegg, J.; Narasimhan, B. Protein adsorption on biodegradable polyanhydride microparticles. J. Biomed. Mater. Res. 2010, 95A, 40–48. [Google Scholar]
- Mancini, G.; Carbonara, A.O.; Heremans, J.F. Immunochemical quantification of antigens by single radial immunodiffusion. Immunochemistry 1965, 2, 235–254. [Google Scholar]
- Kummer, A.; Kitts, D.D.; Li-Chan, E.; Losso, J.N.; Skura, B.J.; Nakai, S. Quantification of bovine IgG in milk using enzyme-linked immunosorbant assay. Food Agric. Immunol. 1992, 4, 93–102. [Google Scholar]
- Ma, L.; Wang, C.; Hong, Y.; Zhang, M.; Su, M. Thermally addressed immunosorbent assay for multiplexed protein detections using phase change nanoparticles. Anal. Chem. 2010, 82, 1186–1190. [Google Scholar]
- Crosson, C.; Thomas, D.; Rossi, C. Quantification of immunoglobulin G in bovine and caprine milk using a surface plasmon resonance-based immunosensor. J. Agric. Food Chem. 2010, 58, 3259–3264. [Google Scholar]
- Nisonoff, A.; Wissler, F.C.; Lipman, L.N.; Woernley, D.L. Separation of univalent fragments from the bivalent rabbit antibody molecule by reduction of disulfide bonds. Arch. Biochem. Biophys. 1960, 89, 230–244. [Google Scholar]
- Fang, W.D.; Mukkur, T.K.S. Physicochemical characteristics of proteolytic cleavage fragments of bovine colostral immunoglobulin G1 (IgG1). Biochem. J. 1976, 155, 25–30. [Google Scholar]
- Carter, P.J. Potent antibody therapeutics by design. Nat. Rev. Immunol. 2006, 6, 343–357. [Google Scholar]
- Brock, J.H.; Arzabe, F.A.; Pineiro, A.; Olivito, A.-M. The effect of trypsin and chymotrypsin on the bactericidal antibody activity of bovine colostrum. Immunology 1977, 32, 207–213. [Google Scholar]
- De Rham, O.; Isliker, H. Proteolysis of bovine immunoglobulins. Int. Arch. Allergy Appl. Immunol. 1977, 55, 61–69. [Google Scholar]
- Yvon, M.; Levieux, D.; Valluv, M.-C.; Pelissier, J.-P.; Mirand, P.P. Colostrum protein digestion in newborn lambs. J. Nutr. 1993, 123, 586–596. [Google Scholar]
- Roos, N.; Mahe, S.; Benamouzig, R.; Sick, H.; Rautureau, J.; Tome, D. 15N-labeled immunoglobulins from bovine colostrum are partially resistant to digestion in human intestine. J. Nutr. 1995, 125, 1238–1244. [Google Scholar] [PubMed]
- Mahe, S.; Huneau, J.-F.; Marteau, P.; Thuille, F.; Tome, D. Gastroileal nitrogen and electrolyte movements after bovine milk ingestion in humans. Am. J. Clin. Nutr. 1992, 56, 410–416. [Google Scholar]
- Kelly, C.P.; Chetham, S.; Keates, S.; Bostwick, E.F.; Roush, A.M.; Castagliuolo, I.; LaMont, J.T.; Pothoulakis, C. Survival of anti-Colostridium difficile bovine immunoglobulin concentrate in the human gastrointestinal tract. Antimicrob. Agents Chemother. 1997, 41, 236–241. [Google Scholar]
- Warny, M.; Fatimi, A.; Bostwick, E.F.; Laine, D.C.; Lebei, F.; LaMont, J.T.; Pothoulakis, C.; Kelly, C.P. Bovine immunoglobulin concentrate-Colostridium difficile retains C. difficile toxin neutralizing activity after passage through the human stomach and small intestine. Gut 1999, 44, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Hurley, W.L. Mammary function during the nonlactating period: Enzyme, lactose, protein concentrations, and pH of mammary secretions. J. Dairy Sci. 1987, 70, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M.; Nagashima, H.; Hasimoto, K. Comparative studies in molecular stability of immunoglobulin G from different species. Comp. Biochem. Physiol. B 1993, 106, 255–261. [Google Scholar]
- Chen, C.-C.; Chang, H.-M. Effect of thermal protectants on the stability of bovine milk immunoglobulin G. J. Agric. Food Chem. 1998, 46, 3570–3576. [Google Scholar]
- Dominguez, E.; Perez, M.D.; Puyol, P.; Sanchez, L.; Calvo, M. Effect of pH on antigen-binding activity of IgG from bovine colostrum upon heating. J. Dairy Res. 2001, 68, 511–518. [Google Scholar]
- Gao, W.; Chen, L.; Xu, L.B.; Huang, X.H. Specific activity against diarrheagenic bacteria in bovine immune milk and effect of pH on its antigen-binding activity upon heating. J. Dairy Res. 2010, 77, 220–224. [Google Scholar]
- Chen, C.-C.; Tu, Y.-Y.; Chang, H.-M. Efficiency and protective effect of encapsulation of milk immunoglobulin G in multiple emulsion. J. Agric. Food Chem. 1999, 47, 407–410. [Google Scholar]
- Calmettes, P.; Cser, L.; Rajnavolgy, E. Temperature and pH dependence of immunoglobulin G conformation. Arch. Biochem. Biophys. 1991, 291, 277–283. [Google Scholar]
- Dominguez, E.; Perez, M.D.; Calvo, M. Effect of heat treatment on the antigen-binding activity of anti-peroxidase immunoglobulins in bovine colostrum. J. Dairy Sci. 1997, 80, 3182–3187. [Google Scholar]
- Mainer, G.; Sanchez, L.; Ena, J.M.; Calvo, M. Kinetic and thermodynamic parameters for heat denaturation of bovine milk IgG, IgA and IgM. J. Food Sci. 1997, 62, 1034–1038. [Google Scholar]
- Chen, C.-C.; Tu, Y.-Y.; Chang, H.-M. Thermal stability of bovine milk immunoglobulin G (IgG) and the effect of added thermal protectants on the stability. J. Food Sci. 2000, 65, 188–193. [Google Scholar]
- Li-Chan, E.; Kummer, A.; Loso, J.N.; Kitts, D.D.; Nakai, S. Stability of bovine immunoglobulins to thermal treatment and processing. Food Res. Int. 1995, 28, 9–16. [Google Scholar]
- Mainer, G.; Dominguez, E.; Randrup, M.; Sanchez, L.; Calvo, M. Effect of heat treatment on anti-rotavirus activity of bovine colostrum. J. Dairy Res. 1999, 66, 131–137. [Google Scholar]
- Chantry, C.J.; Israel-Ballard, K.; Moldoveanu, Z.; Peerson, J.M.; Coutsoudis, A.; Sibeko, L.; Abrams, B. Effect of flash-heat treatment on immunoglobulins in breastmilk. J. Acquir. Immune Defic. Syndr. 2009, 51, 264–267. [Google Scholar]
- Chantry, C.J.; Wiedeman, J.; Buehring, G.; Peerson, J.M.; Hayton, K.; K’Aluoch, O.; Lonnerdal, B.; Israel-Ballard, K.; Coutsoudis, A.; Abrams, B. Effect of flash-heat treatment on antimicrobial activity of breastmilk. Breastfeed. Med. 2010. [Google Scholar]
- Evrendilek, G.A.; Li, S.; Dantzer, W.R.; Zhang, Q.-H. Pulsed electric field processing of beer: Microbial, sensory, and quality analyse. J. Food Sci. 2004, 69, M228–M232. [Google Scholar]
- Li, S.-Q.; Zhang, Q.-H. Inactivation of E. coli 8739 in enriched soymilk using pulsed electric fields. J. Food Sci. 2004, 69, M169–M174. [Google Scholar]
- Yang, R.-J.; Li, S.-Q.; Zhang, Q.-H. Effects of pulsed electric fields on the activity of enzymes in aqueous solution. J. Food Sci. 2004, 69, FCT241–FCT248. [Google Scholar]
- Li, S.-Q.; Zhang, Q.-H.; Lee, Y.-Z.; Pham, T.-V. Effects of pulsed electric fields and thermal processing on the stability of bovine immunoglobulin G (IgG) in enriched soymilk. J. Food Sci. 2003, 68, 1201–1207. [Google Scholar]
- Li, S.-Q.; Bomser, J.A.; Zhang, Q.-H. Effects of pulsed electric fields and heat treatment on stability and secondary structure of bovine immunoglobulin G. J. Agric. Food Chem. 2005, 53, 663–670. [Google Scholar]
- Krishnamurthy, K.; Demirci, A.; Irudayaraj, J.M. Inactivation of Staphylococcus aureus in milk using flow-through pulsed UV-light treatment system. J. Food Sci. 2007, 72, M233–M239. [Google Scholar]
- Balasubramaniam, V.M.; Ting, E.Y.; Stewart, C.M.; Robbins, J.A. Recommended laboratory practices for conducting high-pressure microbial inactivation experiments. Innov. Food Sci. Emerg. Technol. 2004, 5, 299–306. [Google Scholar]
- Li, S.-Q.; Zhang, Q.-H.; Balasubramaniam, V.M.; Lee, Y.-Z.; Bomser, J.A.; Schwartz, S.J.; Dunne, C.P. Comparison of effects of high-pressure processing and heat treatment on immunoactivity of bovine milk immunoglobulin G in enriched soymilk under equivalent microbial inactivation levels. J. Agric. Food Chem. 2006, 54, 739–746. [Google Scholar]
- Balasubramaniam, S.; Balasubramaniam, V.M. Compression heating influence of pressure transmitting fluids on bacteria inactivation during high pressure processing. Food Res. Int. 2003, 36, 661–668. [Google Scholar]
- Trujillo, A.J.; Castro, N.; Quevedo, J.M.; Arguello, A.; Capote, J.; Guamis, B. Effect of heat and high-pressure treatments on microbiological quality and immunoglobulin G stability of caprine colostrum. J. Dairy Sci. 2007, 90, 833–839. [Google Scholar]
- Permanyer, M.; Castellote, C.; Ramirez-Santana, C.; Audi, C.; Pérez-Cano, F.J.; Castell, M.; Lopez-Sabater, M.C.; Franch, A. Maintenance of breast milk immunoglobulin A after high-pressure processing. J. Dairy Sci. 2009, 93, 877–883. [Google Scholar]
- McMartin, S.; Godden, S.; Metzger, L.; Feirtag, J.; Bey, R.; Stabel, J.; Goyal, S.; Fetrow, J.; Wells, S.; Chester-Jones, H. Heat treatment of bovine colostrum. I: Effects of temperature on viscosity and immunoglobulin G level. J. Dairy Sci. 2006, 89, 2110–2118. [Google Scholar] [CrossRef] [PubMed]
- Godden, S.M.; Smith, S.; Feirtag, J.M.; Green, L.R.; Wells, S.J.; Fetrow, J.P. Effect of on-farm commercial batch pasteurization of colostrum on colostrum and serum immunoglobulin concentrations in dairy calves. J. Dairy Sci. 2003, 86, 1503–1512. [Google Scholar]
- Elizondo-Salazar, J.A.; Jayarao, B.M.; Heinrichs, A.J. Effect of heat treatment of bovine colostrum on bacterial counts, viscosity, and immunoglobulin G concentratio. J. Dairy Sci. 2010, 93, 961–967. [Google Scholar]
- Godden, S.; McMartin, S.; Feirtag, J.; Stabel, J.; Bey, R.; Goyal, S.; Metzger, L.; Fetrow, J.; Wells, S.; Chester-Jones, H. Heat-treatment of bovine colostrum. II: Effects of heating duration on pathogen viability and immunoglobulin G. J. Dairy Sci. 2006, 89, 3476–3483. [Google Scholar] [PubMed]
- Johnson, J.L.; Godden, S.M.; Molitor, T.; Ames, T.; Hagman, D. Effects of feeding heat-treated colostrum on passive transfer of immune and nutritional parameters in neonatal dairy calves. J. Dairy Sci. 2007, 90, 5189–5198. [Google Scholar]
- Elizondo-Salazar, J.A.; Heinrichs, A.J. Feeding heat-treated colostrum or unheated colostrum with two different bacterial concentrations to neonatal dairy calves. J. Dairy Sci. 2009, 92, 4565–4571. [Google Scholar]
- Elizondo-Salazar, J.A.; Heinrichs, A.J. Feeding heat-treated colostrum to neonatal dairy heifers: Effects on growth characteristics and blood parameters. J. Dairy Sci. 2009, 92, 3265–3273. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hurley, W.L.; Theil, P.K. Perspectives on Immunoglobulins in Colostrum and Milk. Nutrients 2011, 3, 442-474. https://doi.org/10.3390/nu3040442
Hurley WL, Theil PK. Perspectives on Immunoglobulins in Colostrum and Milk. Nutrients. 2011; 3(4):442-474. https://doi.org/10.3390/nu3040442
Chicago/Turabian StyleHurley, Walter L., and Peter K. Theil. 2011. "Perspectives on Immunoglobulins in Colostrum and Milk" Nutrients 3, no. 4: 442-474. https://doi.org/10.3390/nu3040442
APA StyleHurley, W. L., & Theil, P. K. (2011). Perspectives on Immunoglobulins in Colostrum and Milk. Nutrients, 3(4), 442-474. https://doi.org/10.3390/nu3040442