
Nitrate as Warden of Nitric Oxide Homeostasis in Mammals

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Nitric Oxide Cycle
2.1. Short Story of NO Discovery
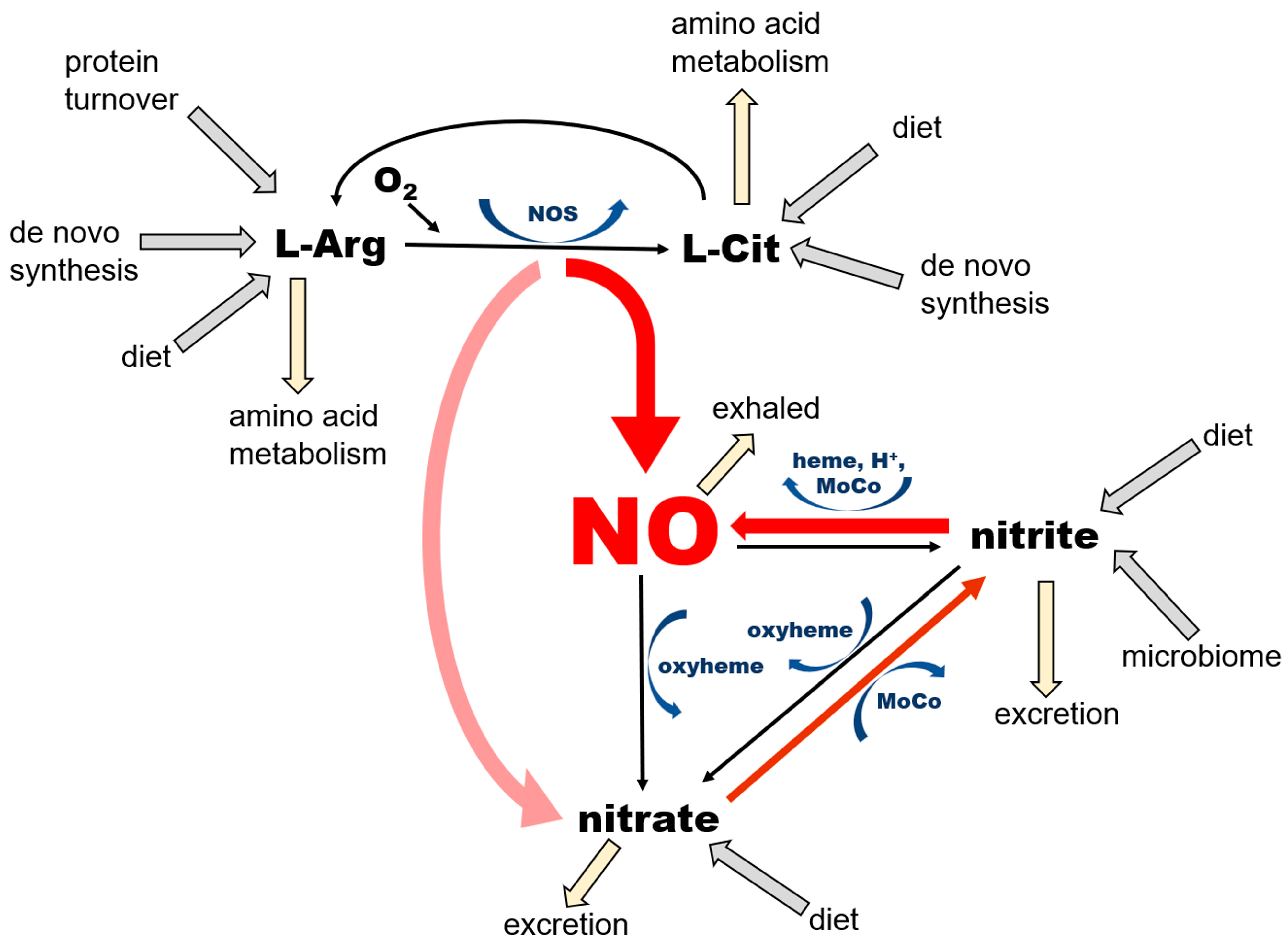
2.2. Organization of the NO Pathway with Emphasis on Its Cyclicity and Self-Sustaining Nature
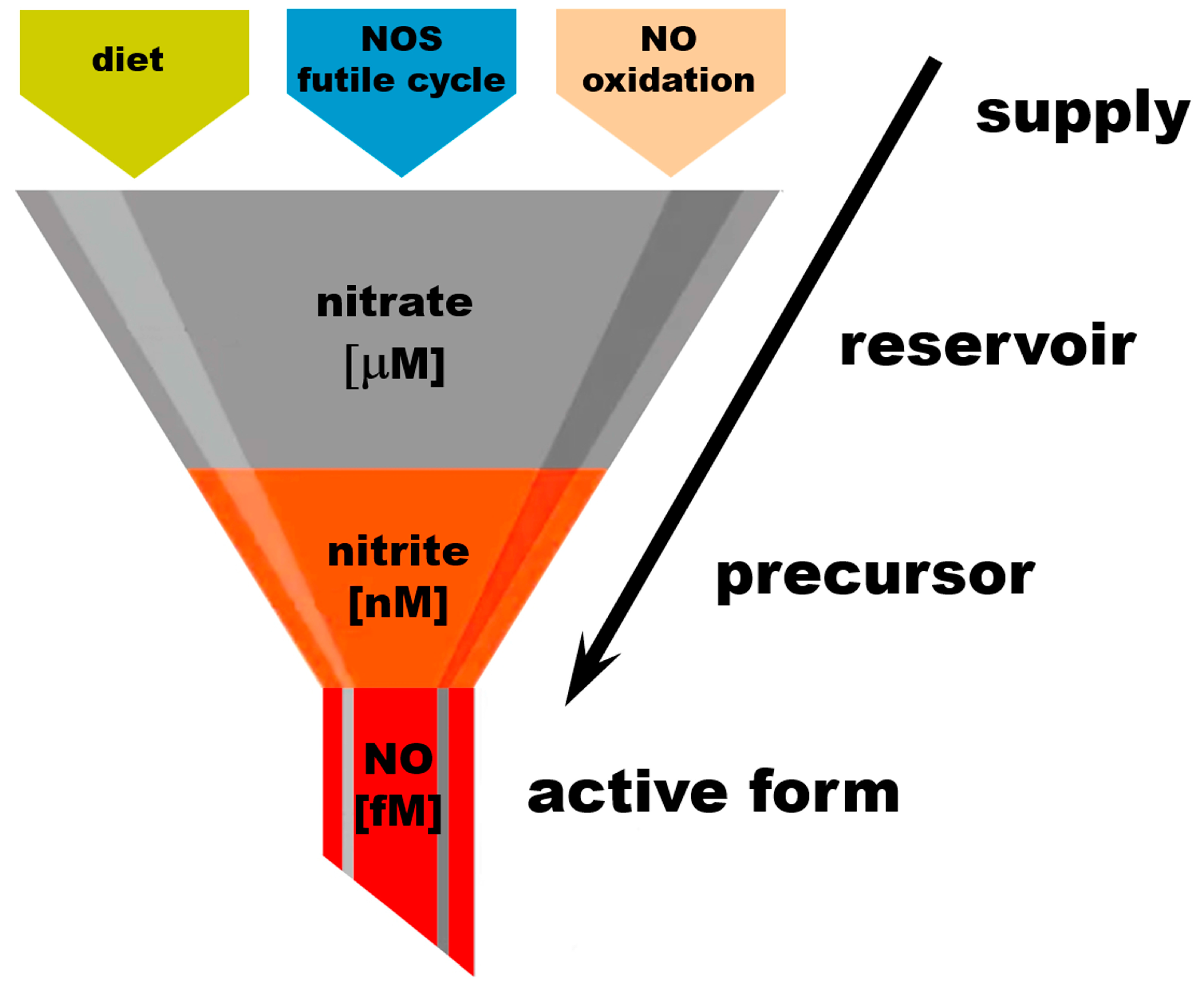
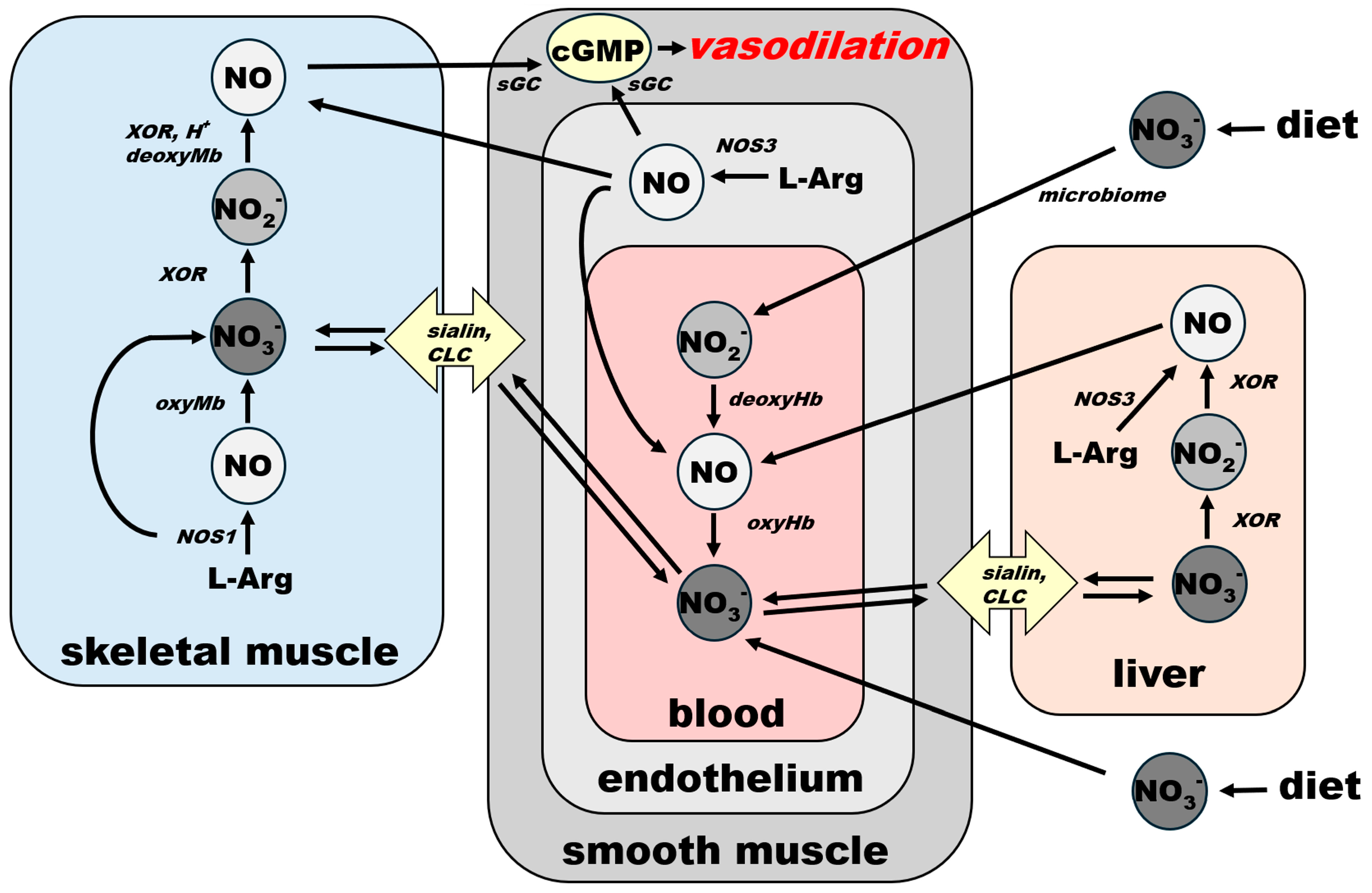
2.3. Nitrate as an Exclusive Storage Molecule of NO Cycle
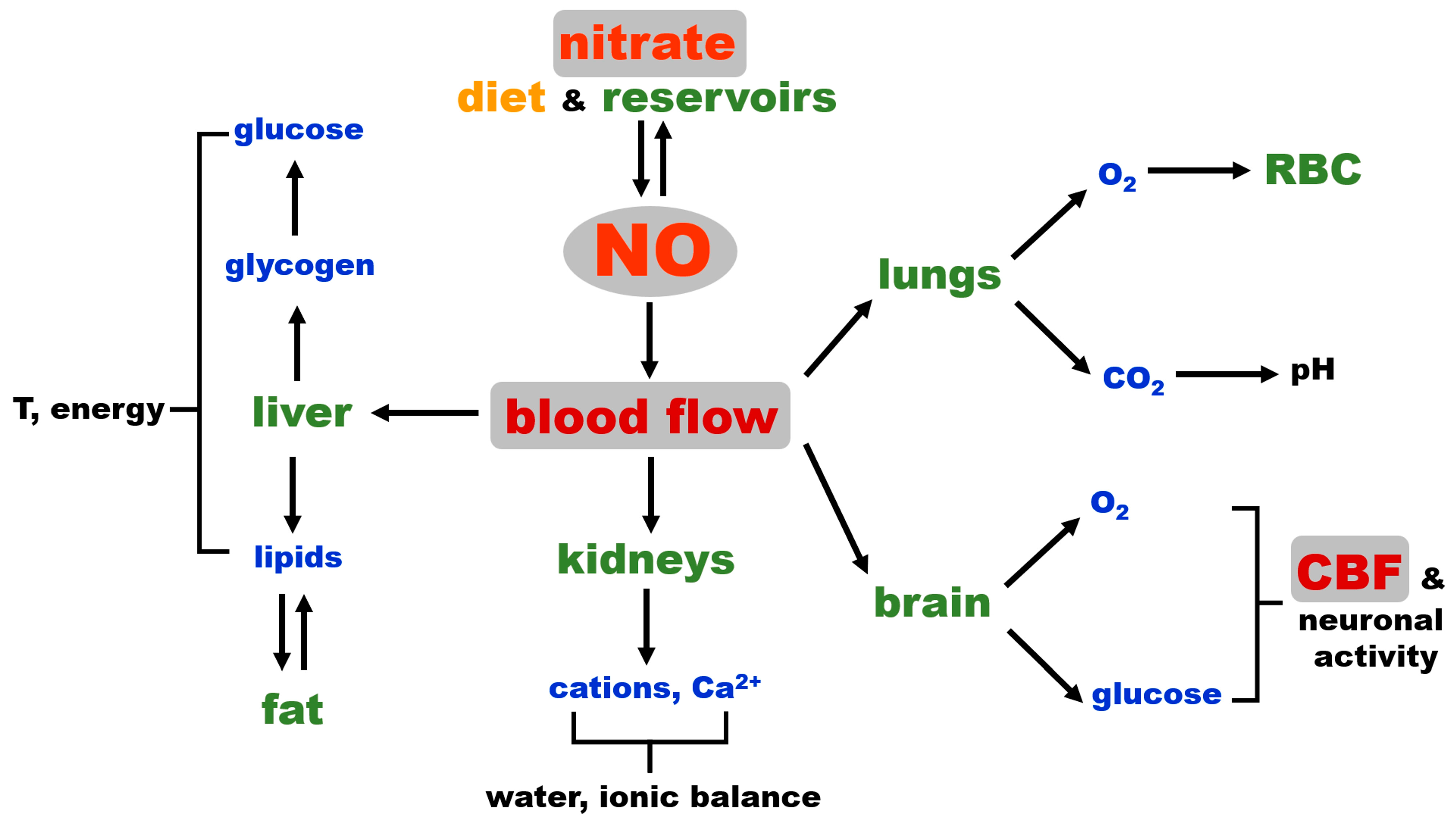
2.4. NO and Blood Flow Control as a Critical Factor for Maintaining General Homeostasis in Mammals
3. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AO | aldehyde oxidase |
| BH4 | tetrahydrobiopterin |
| BAEC | bovine aortic endothelial cells |
| CBF | cerebral blood flow |
| cGMP | guanosine 3′,5′-cyclic monophosphate |
| CLC | chloride channels and transporters |
| COPD | chronic pulmonary obstructive disease |
| eNOS/NOS3 | endothelial nitric oxide synthase |
| nNOS/NOS1 | neuronal nitric oxide synthase |
| FAD | flavin adenine dinucleotide |
| FMN | flavin mononucleotide |
| IPF | idiopathic pulmonary fibrosis |
| mARC | mitochondrial amidoxime reducing component |
| MoCo proteins | molybdopterin motive containing proteins |
| NADPH | nicotinamide adenine dinucleotide phosphate |
| NO | nitric oxide |
| NO2− | nitrite |
| NO3− | nitrate |
| NOS | nitric oxide synthase |
| ONOO | peroxinitrite |
| RBC | red blood cell |
| sGC | soluble guanylate cyclase |
| SO | sulfite oxidase |
| XOR | xanthine oxidoreductase |
References
- Moroz, L.L.; Mukherjee, K.; Romanova, D.Y. Nitric oxide signaling in ctenophores. Front. Neurosci. 2023, 17, 1125433. [Google Scholar] [CrossRef]
- Cristino, L.; Guglielmotti, V.; Cotugno, A.; Musio, C.; Santillo, S. Nitric oxide signaling pathways at neural level in invertebrates: Functional implications in cnidarians. Brain Res. 2008, 1225, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, K.; Almeida-Souza, T.H.; Silva, R.S.; Santos, H.F.; Santos, E.V.; Gois, A.M.; Leal, P.C.; Santos, J.R. Involvement of nitric oxide in the neurobiology of fear-like behavior. Nitric Oxide 2022, 124, 24–31. [Google Scholar] [CrossRef]
- Donald, J.A.; Forgan, L.G.; Cameron, M.S. The evolution of nitric oxide signalling in vertebrate blood vessels. J. Comp. Physiol. B 2015, 185, 153–171. [Google Scholar] [CrossRef] [PubMed]
- Andrabi, S.M.; Sharma, N.S.; Karan, A.; Shahriar, S.M.S.; Cordon, B.; Ma, B.; Xie, J. Nitric Oxide: Physiological Functions, Delivery, and Biomedical Applications. Adv. Sci. 2023, 10, e2303259. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, J.R., Jr. Historical origins of the discovery of mammalian nitric oxide (nitrogen monoxide) production/physiology/pathophysiology. Biochem. Pharmacol. 2020, 176, 113793. [Google Scholar] [CrossRef]
- Lundberg, J.O.; Weitzberg, E.; Lundberg, J.M.; Alving, K. Intragastric nitric oxide production in humans: Measurements in expelled air. Gut 1994, 35, 1543–1546. [Google Scholar] [CrossRef]
- Zweier, J.L.; Wang, P.; Samouilov, A.; Kuppusamy, P. Enzyme-independent formation of nitric oxide in biological tissues. Nat. Med. 1995, 1, 804–809. [Google Scholar] [CrossRef]
- Zweier, J.L.; Samouilov, A.; Kuppusamy, P. Non-enzymatic nitric oxide synthesis in biological systems. Biochim. Biophys. Acta 1999, 1411, 250–262. [Google Scholar] [CrossRef]
- Cosby, K.; Partovi, K.S.; Crawford, J.H.; Patel, R.P.; Reiter, C.D.; Martyr, S.; Yang, B.K.; Waclawiw, M.A.; Zalos, G.; Xu, X.; et al. Nitrite reduction to nitric oxide by deoxyhemoglobin vasodilates the human circulation. Nat. Med. 2003, 9, 1498–1505. [Google Scholar] [CrossRef]
- Gladwin, M.T.; Schechter, A.N.; Kim-Shapiro, D.B.; Patel, R.P.; Hogg, N.; Shiva, S.; Cannon, R.O., 3rd; Kelm, M.; Wink, D.A.; Espey, M.G.; et al. The emerging biology of the nitrite anion. Nat. Chem. Biol. 2005, 1, 308–314. [Google Scholar] [CrossRef]
- Dejam, A.; Hunter, C.J.; Tremonti, C.; Pluta, R.M.; Hon, Y.Y.; Grimes, G.; Partovi, K.; Pelletier, M.M.; Oldfield, E.H.; Cannon, R.O., 3rd; et al. Nitrite infusion in humans and nonhuman primates: Endocrine effects, pharmacokinetics, and tolerance formation. Circulation 2007, 116, 1821–1831. [Google Scholar] [CrossRef] [PubMed]
- Bryan, N.S. Nitrite in nitric oxide biology: Cause or consequence? A systems-based review. Free Radic. Biol. Med. 2006, 41, 691–701. [Google Scholar] [CrossRef] [PubMed]
- Totzeck, M.; Hendgen-Cotta, U.B.; Luedike, P.; Berenbrink, M.; Klare, J.P.; Steinhoff, H.J.; Semmler, D.; Shiva, S.; Williams, D.; Kipar, A.; et al. Nitrite regulates hypoxic vasodilation via myoglobin-dependent nitric oxide generation. Circulation 2012, 126, 325–334. [Google Scholar] [CrossRef] [PubMed]
- van Faassen, E.E.; Babrami, S.; Feelisch, M.; Hogg, N.; Kelm, M.; Kim-Shapiro, D.B.; Kozlov, A.V.; Li, H.T.; Lundberg, J.O.; Mason, R.; et al. Nitrite as Regulator of Hypoxic Signaling in Mammalian Physiology. Med. Res. Rev. 2009, 29, 683–741. [Google Scholar] [CrossRef]
- Pinder, A.G.; Pittaway, E.; Morris, K.; James, P.E. Nitrite directly vasodilates hypoxic vasculature via nitric oxide-dependent and -independent pathways. Br. J. Pharmacol. 2009, 157, 1523–1530. [Google Scholar] [CrossRef]
- Duncan, C.; Li, H.; Dykhuizen, R.; Frazer, R.; Johnston, P.; MacKnight, G.; Smith, L.; Lamza, K.; McKenzie, H.; Batt, L.; et al. Protection against oral and gastrointestinal diseases: Importance of dietary nitrate intake, oral nitrate reduction and enterosalivary nitrate circulation. Comp. Biochem. Physiol. A Physiol. 1997, 118, 939–948. [Google Scholar] [CrossRef]
- Lundberg, J.O.; Weitzberg, E.; Cole, J.A.; Benjamin, N. Nitrate, bacteria and human health. Nat. Rev. Microbiol. 2004, 2, 593–602. [Google Scholar] [CrossRef]
- Bryan, N.S.; Ahmed, S.; Lefer, D.J.; Hord, N.; von Schwarz, E.R. Dietary nitrate biochemistry and physiology. An update on clinical benefits and mechanisms of action. Nitric Oxide 2023, 132, 1–7. [Google Scholar] [CrossRef]
- Liu, H.; Huang, Y.; Huang, M.; Wang, M.; Ming, Y.; Chen, W.; Chen, Y.; Tang, Z.; Jia, B. From nitrate to NO: Potential effects of nitrate-reducing bacteria on systemic health and disease. Eur. J. Med. Res. 2023, 28, 425. [Google Scholar] [CrossRef]
- Volino-Souza, M.; Oliveira, G.V.; Pinheiro, V.D.S.; Conte-Junior, C.A.; Alvares, T.D.S. The effect of dietary nitrate on macro- and microvascular function: A systematic review. Crit. Rev. Food Sci. Nutr. 2024, 64, 1225–1236. [Google Scholar] [CrossRef] [PubMed]
- Derella, C.C.; Anderson, K.C.; Woessner, M.N.; Paterson, C.; Allen, J.D. Ergogenic Effect of Nitrate Supplementation in Clinical Populations: A Systematic Review and Meta-Analysis. Nutrients 2024, 16, 3832. [Google Scholar] [CrossRef]
- Tripodi, G.; Lombardo, M.; Kerav, S.; Aiello, G.; Baldelli, S. Nitric Oxide in Parkinson’s Disease: The Potential Role of Dietary Nitrate in Enhancing Cognitive and Motor Health via the Nitrate-Nitrite-Nitric Oxide Pathway. Nutrients 2025, 17, 393. [Google Scholar] [CrossRef] [PubMed]
- Jansson, E.A.; Huang, L.; Malkey, R.; Govoni, M.; Nihlen, C.; Olsson, A.; Stensdotter, M.; Petersson, J.; Holm, L.; Weitzberg, E.; et al. A mammalian functional nitrate reductase that regulates nitrite and nitric oxide homeostasis. Nat. Chem. Biol. 2008, 4, 411–417. [Google Scholar] [CrossRef]
- Huang, L.; Borniquel, S.; Lundberg, J.O. Enhanced xanthine oxidoreductase expression and tissue nitrate reduction in germ free mice. Nitric Oxide 2010, 22, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Koch, C.D.; Gladwin, M.T.; Freeman, B.A.; Lundberg, J.O.; Weitzberg, E.; Morris, A. Enterosalivary nitrate metabolism and the microbiome: Intersection of microbial metabolism, nitric oxide and diet in cardiac and pulmonary vascular health. Free Radic. Biol. Med. 2017, 105, 48–67. [Google Scholar] [CrossRef]
- Archer, D.L. Evidence that ingested nitrate and nitrite are beneficial to health. J. Food Prot. 2002, 65, 872–875. [Google Scholar] [CrossRef]
- Piknova, B.; Park, J.W.; Swanson, K.M.; Dey, S.; Noguchi, C.T.; Schechter, A.N. Skeletal muscle as an endogenous nitrate reservoir. Nitric Oxide 2015, 47, 10–16. [Google Scholar] [CrossRef]
- Piknova, B.; Park, J.W.; Tunau-Spencer, K.J.; Jenkins, A.; Hellinga, D.G.; Walter, P.J.; Cai, H.; Schechter, A.N. Skeletal Muscle, Skin, and Bone as Three Major Nitrate Reservoirs in Mammals: Chemiluminescence and (15)N-Tracer Studies in Yorkshire Pigs. Nutrients 2024, 16, 2674. [Google Scholar] [CrossRef]
- Stuehr, D.J.; Santolini, J.; Wang, Z.Q.; Wei, C.C.; Adak, S. Update on mechanism and catalytic regulation in the NO synthases. J. Biol. Chem. 2004, 279, 36167–36170. [Google Scholar] [CrossRef]
- Shiva, S.; Wang, X.; Ringwood, L.A.; Xu, X.; Yuditskaya, S.; Annavajjhala, V.; Miyajima, H.; Hogg, N.; Harris, Z.L.; Gladwin, M.T. Ceruloplasmin is a NO oxidase and nitrite synthase that determines endocrine NO homeostasis. Nat. Chem. Biol. 2006, 2, 486–493. [Google Scholar] [CrossRef]
- Stuehr, D.J.; Haque, M.M. Nitric oxide synthase enzymology in the 20 years after the Nobel Prize. Br. J. Pharmacol. 2019, 176, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Blekkenhorst, L.C.; Bondonno, N.P.; Sim, M.; Woodman, R.J.; Croft, K.D.; Lewis, J.R.; Hodgson, J.M.; Bondonno, C.P. A food composition database for assessing nitrate intake from plant-based foods. Food Chem. 2022, 394, 133411. [Google Scholar] [CrossRef]
- Hosseini, M.J.; Dezhangah, S.; Esmi, F.; Gharavi-nakhjavani, M.S.; Hashempour-Baltork, F.; Mirza Alizadeh, A. A worldwide systematic review, meta-analysis and meta-regression of nitrate and nitrite in vegetables and fruits. Ecotoxicol. Environ. Saf. 2023, 257, 114934. [Google Scholar] [CrossRef] [PubMed]
- Lidder, S.; Webb, A.J. Vascular effects of dietary nitrate (as found in green leafy vegetables and beetroot) via the nitrate-nitrite-nitric oxide pathway. Br. J. Clin. Pharmacol. 2013, 75, 677–696. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Liu, X.; Sun, Q.; Fan, Z.; Xia, D.; Ding, G.; Ong, H.L.; Adams, D.; Gahl, W.A.; Zheng, C.; et al. Sialin (SLC17A5) functions as a nitrate transporter in the plasma membrane. Proc. Natl. Acad. Sci. USA 2012, 109, 13434–13439. [Google Scholar] [CrossRef]
- Qu, X.M.; Wu, Z.F.; Pang, B.X.; Jin, L.Y.; Qin, L.Z.; Wang, S.L. From Nitrate to Nitric Oxide: The Role of Salivary Glands and Oral Bacteria. J. Dent. Res. 2016, 95, 1452–1456. [Google Scholar] [CrossRef]
- Srihirun, S.; Park, J.W.; Teng, R.; Sawaengdee, W.; Piknova, B.; Schechter, A.N. Nitrate uptake and metabolism in human skeletal muscle cell cultures. Nitric Oxide 2020, 94, 1–8. [Google Scholar] [CrossRef]
- Akhtar, S.; Sagar, K.; Singh, A.; Hote, M.P.; Roy, A.; Sharma, A. Inflammation-induced sialin mediates nitrate efflux in dysfunctional endothelium affecting NO bioavailability. Nitric Oxide 2024, 146, 37–47. [Google Scholar] [CrossRef]
- Jentsch, T.J.; Pusch, M. CLC Chloride Channels and Transporters: Structure, Function, Physiology, and Disease. Physiol. Rev. 2018, 98, 1493–1590. [Google Scholar] [CrossRef]
- Piknova, B.; Park, J.W.; Lam, K.K.; Schechter, A.N. Nitrate as a source of nitrite and nitric oxide during exercise hyperemia in rat skeletal muscle. Nitric Oxide—Biol. Chem. 2016, 55–56, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Suschek, C.V.; Feibel, D.; von Kohout, M.; Oplander, C. Enhancement of Nitric Oxide Bioavailability by Modulation of Cutaneous Nitric Oxide Stores. Biomedicines 2022, 10, 2124. [Google Scholar] [CrossRef]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef] [PubMed]
- Lüneburg, N.; Xanthakis, V.; Schwedhelm, E.; Sullivan, L.M.; Maas, R.; Anderssohn, M.; Riederer, U.; Glazer, N.L.; Vasan, R.S.; Böger, R.H. Reference Intervals for Plasma L-Arginine and the L-Arginine:Asymmetric Dimethylarginine Ratio in the Framingham Offspring Cohort. J. Nutr. 2011, 141, 2186–2190. [Google Scholar] [CrossRef]
- Maas, R.; Xanthakis, V.; Goen, T.; Muller, J.; Schwedhelm, E.; Boger, R.H.; Vasan, R.S. Plasma Nitrate and Incidence of Cardiovascular Disease and All-Cause Mortality in the Community: The Framingham Offspring Study. J. Am. Heart Assoc. 2017, 6, e006224. [Google Scholar] [CrossRef]
- Boger, R.H. The pharmacodynamics of L-arginine. J. Nutr. 2007, 137, 1650S–1655S. [Google Scholar] [CrossRef] [PubMed]
- Baydoun, A.R.; Emery, P.W.; Pearson, J.D.; Mann, G.E. Substrate-dependent regulation of intracellular amino acid concentrations in cultured bovine aortic endothelial cells. Biochem. Biophys. Res. Commun. 1990, 173, 940–948. [Google Scholar] [CrossRef]
- Wyatt, A.W.; Steinert, J.R.; Mann, G.E. Modulation of the L-arginine/nitric oxide signalling pathway in vascular endothelial cells. Biochem. Soc. Symp. 2004, 71, 143–156. [Google Scholar] [CrossRef]
- Gornik, H.L.; Creager, M.A. Arginine and endothelial and vascular health. J. Nutr. 2004, 134, 2880S–2887S; discussion 2895S. [Google Scholar] [CrossRef]
- Morris, S.M., Jr. Arginine Metabolism Revisited. J. Nutr. 2016, 146, 2579S–2586S. [Google Scholar] [CrossRef]
- Jonvik, K.L.; Nyakayiru, J.; Pinckaers, P.J.; Senden, J.M.; van Loon, L.J.; Verdijk, L.B. Nitrate-Rich Vegetables Increase Plasma Nitrate and Nitrite Concentrations and Lower Blood Pressure in Healthy Adults. J. Nutr. 2016, 146, 986–993. [Google Scholar] [CrossRef]
- Gilliard, C.N.; Lam, J.K.; Cassel, K.S.; Park, J.W.; Schechter, A.N.; Piknova, B. Effect of dietary nitrate levels on nitrate fluxes in rat skeletal muscle and liver. Nitric Oxide 2018, 75, 1–7. [Google Scholar] [CrossRef]
- Piknova, B.; Schechter, A.N.; Park, J.W.; Vanhatalo, A.; Jones, A.M. Skeletal Muscle Nitrate as a Regulator of Systemic Nitric Oxide Homeostasis. Exerc. Sport Sci. Rev. 2022, 50, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Ehmsen, J.; Poon, E.; Davies, K. The dystrophin-associated protein complex. J. Cell Sci. 2002, 115, 2801–2803. [Google Scholar] [CrossRef]
- Gao, Q.Q.; McNally, E.M. The Dystrophin Complex: Structure, Function, and Implications for Therapy. Compr. Physiol. 2015, 5, 1223–1239. [Google Scholar] [CrossRef] [PubMed]
- Tejero, J.; Hunt, A.P.; Santolini, J.; Lehnert, N.; Stuehr, D.J. Mechanism and regulation of ferrous heme-nitric oxide (NO) oxidation in NO synthases. J. Biol. Chem. 2019, 294, 7904–7916. [Google Scholar] [CrossRef] [PubMed]
- Garbincius, J.F.; Michele, D.E. Dystrophin-glycoprotein complex regulates muscle nitric oxide production through mechanoregulation of AMPK signaling. Proc. Natl. Acad. Sci. USA 2015, 112, 13663–13668. [Google Scholar] [CrossRef]
- Dombernowsky, N.W.; Olmestig, J.N.E.; Witting, N.; Kruuse, C. Role of neuronal nitric oxide synthase (nNOS) in Duchenne and Becker muscular dystrophies—Still a possible treatment modality? Neuromuscul. Disord. 2018, 28, 914–926. [Google Scholar] [CrossRef]
- Sweeney, H.L.; Barton, E.R. The dystrophin-associated glycoprotein complex: What parts can you do without? Proc. Natl. Acad. Sci. USA 2000, 97, 13464–13466. [Google Scholar] [CrossRef]
- Sander, M.; Chavoshan, B.; Harris, S.A.; Iannaccone, S.T.; Stull, J.T.; Thomas, G.D.; Victor, R.G. Functional muscle ischemia in neuronal nitric oxide synthase-deficient skeletal muscle of children with Duchenne muscular dystrophy. Proc. Natl. Acad. Sci. USA 2000, 97, 13818–13823. [Google Scholar] [CrossRef]
- Nelson, M.D.; Rosenberry, R.; Barresi, R.; Tsimerinov, E.I.; Rader, F.; Tang, X.; Mason, O.; Schwartz, A.; Stabler, T.; Shidban, S.; et al. Sodium nitrate alleviates functional muscle ischaemia in patients with Becker muscular dystrophy. J. Physiol. 2015, 593, 5183–5200. [Google Scholar] [CrossRef]
- Wylie, L.J.; Park, J.W.; Vanhatalo, A.; Kadach, S.; Black, M.I.; Stoyanov, Z.; Schechter, A.N.; Jones, A.M.; Piknova, B. Human skeletal muscle nitrate store: Influence of dietary nitrate supplementation and exercise. J. Physiol. 2019, 597, 5565–5576. [Google Scholar] [CrossRef] [PubMed]
- Black, M.I.; Wylie, L.J.; Kadach, S.; Piknova, B.; Park, J.W.; Stoyanov, Z.; L’Heureux, J.E.; Schechter, A.N.; Vanhatalo, A.; Jones, A.M. Effects of low and high dietary nitrate intake on human saliva, plasma and skeletal muscle nitrate and nitrite concentrations and their functional consequences. Free Radic. Biol. Med. 2024, 225, 881–893. [Google Scholar] [CrossRef]
- Park, J.W.; Piknova, B.; Walter, P.J.; Cai, H.; Upanan, S.; Thomas, S.M.; Tunau-Spencer, K.J.; Schechter, A.N. Distribution of dietary nitrate and its metabolites in rat tissues after (15)N-labeled nitrate administration. Sci. Rep. 2023, 13, 3499. [Google Scholar] [CrossRef]
- Park, J.W.; Piknova, B.; Dey, S.; Noguchi, C.T.; Schechter, A.N. Compensatory mechanisms in myoglobin deficient mice preserve NO homeostasis. Nitric Oxide 2019, 90, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Kadach, S.; Piknova, B.; Black, M.I.; Park, J.W.; Wylie, L.J.; Stoyanov, Z.; Thomas, S.M.; McMahon, N.F.; Vanhatalo, A.; Schechter, A.N.; et al. Time course of human skeletal muscle nitrate and nitrite concentration changes following dietary nitrate ingestion. Nitric Oxide 2022, 121, 1–10. [Google Scholar] [CrossRef]
- Frisbee, J.C. Reduced nitric oxide bioavailability contributes to skeletal muscle microvessel rarefaction in the metabolic syndrome. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 289, R307–R316. [Google Scholar] [CrossRef]
- Ghimire, K.; Altmann, H.M.; Straub, A.C.; Isenberg, J.S. Nitric oxide: What’s new to NO? Am. J. Physiol. Cell Physiol. 2017, 312, C254–C262. [Google Scholar] [CrossRef]
- Wang, Y.; Simons, M. Flow-regulated lymphatic vasculature development and signaling. Vasc. Cell 2014, 6, 14. [Google Scholar] [CrossRef]
- Ohhashi, T.; Kawai, Y.; Maejima, D.; Hayashi, M.; Watanabe-Asaka, T. Physiological Roles of Lymph Flow-Mediated Nitric Oxide in Lymphatic System. Lymphat. Res. Biol. 2023, 21, 253–261. [Google Scholar]
- Esplugues, J.V. NO as a signalling molecule in the nervous system. Br. J. Pharmacol. 2002, 135, 1079–1095. [Google Scholar] [CrossRef] [PubMed]
- Picon-Pages, P.; Garcia-Buendia, J.; Munoz, F.J. Functions and dysfunctions of nitric oxide in brain. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 1949–1967. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, A.; Jeddi, S.; Kashfi, K. Brain glucose metabolism: Role of nitric oxide. Biochem. Pharmacol. 2025, 232, 116728. [Google Scholar] [CrossRef] [PubMed]
- Roger, N.; Barbera, J.A.; Roca, J.; Rovira, I.; Gomez, F.P.; Rodriguez-Roisin, R. Nitric oxide inhalation during exercise in chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 1997, 156, 800–806. [Google Scholar] [CrossRef]
- Hasuda, T.; Satoh, T.; Shimouchi, A.; Sakamaki, F.; Kyotani, S.; Matsumoto, T.; Goto, Y.; Nakanishi, N. Improvement in exercise capacity with nitric oxide inhalation in patients with precapillary pulmonary hypertension. Circulation 2000, 101, 2066–2070. [Google Scholar] [CrossRef]
- Blanco, I.; Ribas, J.; Xaubet, A.; Gómez, F.P.; Roca, J.; Rodriguez-Roisin, R.; Barberà, J.A. Effects of inhaled nitric oxide at rest and during exercise in idiopathic pulmonary fibrosis. J. Appl. Physiol. 2011, 110, 638–645. [Google Scholar] [CrossRef]
- Gwyn, D.R.; Lindeman, K.S.; Hirshman, C.A. Inhaled nitric oxide attenuates bronchoconstriction in canine peripheral airways. Am. J. Respir. Crit. Care Med. 1996, 153, 604–609. [Google Scholar] [CrossRef]
- Hunter, C.J.; Dejam, A.; Blood, A.B.; Shields, H.; Kim-Shapiro, D.B.; Machado, R.F.; Tarekegn, S.; Mulla, N.; Hopper, A.O.; Schechter, A.N.; et al. Inhaled nebulized nitrite is a hypoxia-sensitive NO-dependent selective pulmonary vasodilator. Nat. Med. 2004, 10, 1122–1127. [Google Scholar] [CrossRef]
- Casey, D.B.; Badejo, A.M., Jr.; Dhaliwal, J.S.; Murthy, S.N.; Hyman, A.L.; Nossaman, B.D.; Kadowitz, P.J. Pulmonary vasodilator responses to sodium nitrite are mediated by an allopurinol-sensitive mechanism in the rat. Am. J. Physiol. Heart Circ. Physiol. 2009, 296, H524–H533. [Google Scholar] [CrossRef]
- Reddy, Y.N.V.; Stewart, G.M.; Obokata, M.; Koepp, K.E.; Borlaug, B.A. Peripheral and pulmonary effects of inorganic nitrite during exercise in heart failure with preserved ejection fraction. Eur. J. Heart Fail. 2021, 23, 814–823. [Google Scholar] [CrossRef]
- Pana, R.; Hornby, L.; Shemie, S.D.; Dhanani, S.; Teitelbaum, J. Time to loss of brain function and activity during circulatory arrest. J. Crit. Care 2016, 34, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Dienel, G.A. Brain Glucose Metabolism: Integration of Energetics with Function. Physiol. Rev. 2019, 99, 949–1045. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piknova, B.; Park, J.W.; Schechter, A.N. Nitrate as Warden of Nitric Oxide Homeostasis in Mammals. Nutrients 2025, 17, 1544. https://doi.org/10.3390/nu17091544
Piknova B, Park JW, Schechter AN. Nitrate as Warden of Nitric Oxide Homeostasis in Mammals. Nutrients. 2025; 17(9):1544. https://doi.org/10.3390/nu17091544
Chicago/Turabian StylePiknova, Barbora, Ji Won Park, and Alan N. Schechter. 2025. "Nitrate as Warden of Nitric Oxide Homeostasis in Mammals" Nutrients 17, no. 9: 1544. https://doi.org/10.3390/nu17091544
APA StylePiknova, B., Park, J. W., & Schechter, A. N. (2025). Nitrate as Warden of Nitric Oxide Homeostasis in Mammals. Nutrients, 17(9), 1544. https://doi.org/10.3390/nu17091544