
The Allergy Crossroads of Subtropical Regions: Mites, Crustaceans, and the Rise of Edible Insects

 , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
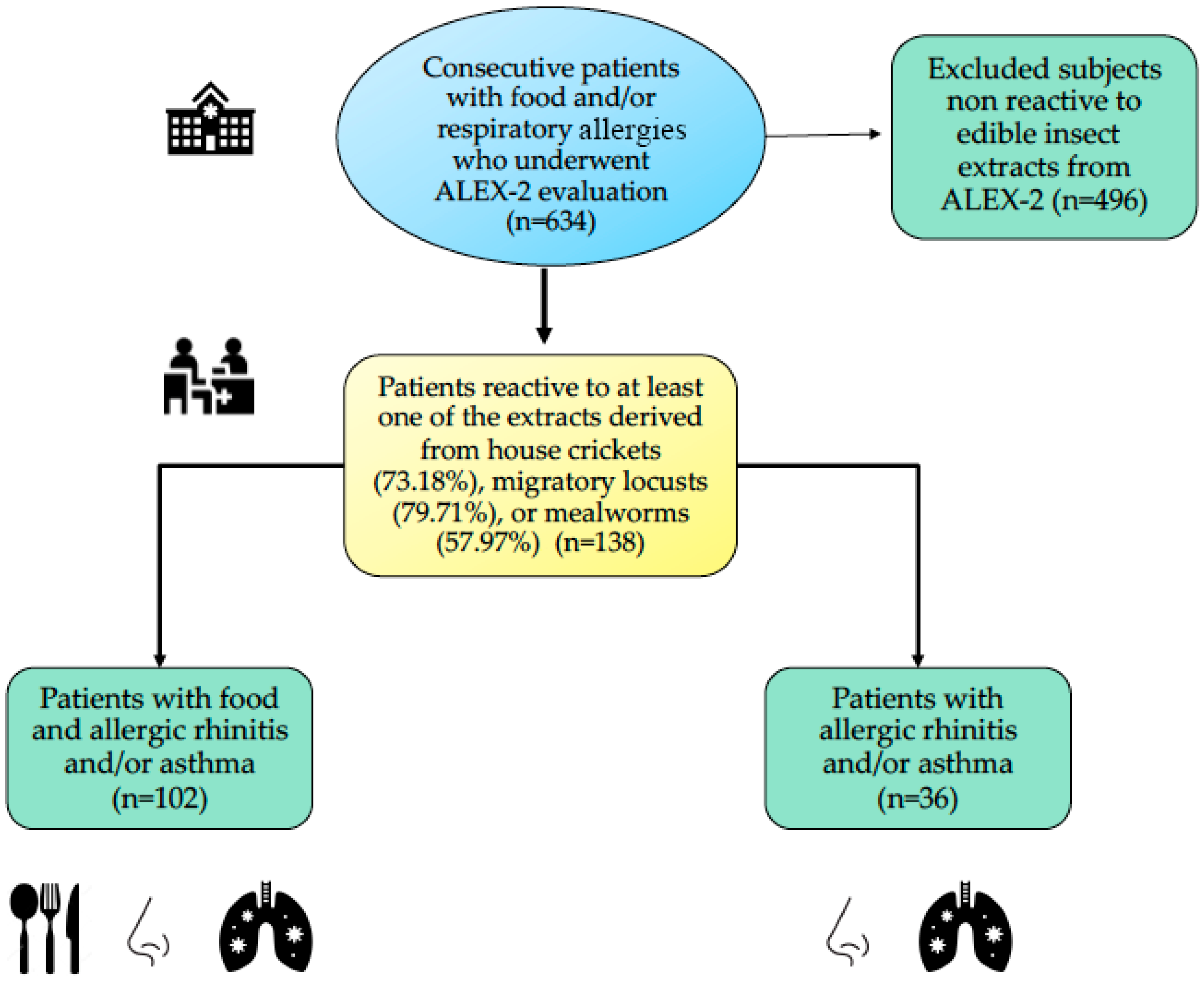
2.1. Subjects
2.2. Serological Analysis
2.3. Statistical Analysis
3. Results
3.1. Study Population
3.2. Specific IgE Profile in Patients with a Sensitization to EIs
3.3. Multiplex IgE Reactivity Profiles in Patients with Sensitization to EIs and Exclusively Affected by Respiratory Allergies
3.4. Allergen-Specific IgE Levels to TMs, AKs, and Different EI Extracts Were Significantly Correlated
4. Discussion
4.1. Molecular Sensitization Patterns in the Investigated Cohort
4.2. Cross-Reactivity Among EIs and Other Allergens
4.3. Insect-Specific Proteins and Sensitization Mechanisms
4.4. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Tm | Tenebrio molitor |
| EIs | Edible Insects |
| EFSA | European Food Safety Authority |
| HDMs | House Dust Mites |
| TM | Tropomyosin |
| Lm | Locusta migratoria |
| Ad | Acheta domesticus |
| AK | Arginine Kinase |
Appendix A

{kind=link}
| Mite Allergen | Acheta domesticus r (p Value) | Locusta migratoria r (p Value) | Tenebrio mollitor r (p Value) |
|---|---|---|---|
| Der f 2 | 0.13 (0.1) | 0.1 (0.21) | 0.02 (0.79) |
| Der p 2 | −0.28 (0.06) | −0.05 (0.34) | −0.15 (0.3) |
| Der p 1 | 0.32 (0.05) | 0.29 (0.08) | 0.14 (0.09) |
| Der p 23 | 0.28 (0.09) | 0.25 (0.13) | 0.24 (0.1) |
| Der f 1 | 0.18 (0.27) | 0.13 (0.43) | 0.03 (0.8) |
| Der p 5 | 0.25 (0.12) | 0.29 (0.08) | 0.17 (0.31) |
| Der p 7 | 0.24 (0.1) | 0.21 (0.12) | 0.22 (0.2) |
| Blo t 21 | 0.22 (0.18) | 0.14 (0.39) | 0.06 (0.69) |
| Gly d 2 | 0.06 (0.68) | −0.05 (0.72) | 0.003 (0.98) |
| Tyr p 2 | 0.12 (0.09) | 0.15 (0.1) | 0.23 (0.16) |
| Der p 21 | −0.07 (0.66) | −0.11 (0.51) | −0.15 (0.37) |
| Blo t 5 | 0.16 (0.33) | 0.09 (0.58) | 0.02 (0.88) |
| Lep d 2 | 0.23 (0.15) | 0.09 (0.56) | 0.18 (0.28) |
| Der p 20 | 0.29 (0.001) | 0.31 (0.0019) | 0.26 (0.0017) |
| Der p 10 | 0.71 (<0.0001) | 0.77 (<0.0001) | 0.82 (<0.0001) |
| Blo t 10 | 0.7 (<0.0001) | 0.71 (<0.0001) | 0.78 (<0.0001) |
| Der p 11 | 0.0 (0.0) | 0.0 (0.0) | 0.0 (0.0) |
References
- Commission Implementing Regulation (Eu) 2025/89 of 20 January 2025. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=OJ:L_202500089 (accessed on 3 February 2025).
- Edible Insects: The Science of Novel Food Evaluations. Available online: https://www.efsa.europa.eu/en/news/edible-insects-science-novel-food-evaluations (accessed on 7 February 2025).
- Francis, F.; Doyen, V.; Debaugnies, F.; Mazzucchelli, G.; Caparros, R.; Alabi, T.; Blecker, C.; Haubruge, E.; Corazza, F. Limited cross reactivity among arginine kinase allergens from mealworm and cricket edible insects. Food Chem. 2019, 276, 714–718. [Google Scholar] [CrossRef] [PubMed]
- Barre, A.; Pichereaux, C.; Simplicien, M.; Burlet-Schiltz, O.; Benoist, H.; Rougé, P. A Proteomic- and Bioinformatic-Based Identification of Specific Allergens from Edible Insects: Probes for Future Detection as Food Ingredients. Foods 2021, 10, 280. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- de Gier, S.; Verhoeckx, K. Insect (food) allergy and allergens. Mol. Immunol. 2018, 100, 82–106. [Google Scholar] [CrossRef] [PubMed]
- Lamberti, C.; Nebbia, S.; Cirrincione, S.; Brussino, L.; Giorgis, V.; Romito, A.; Marchese, C.; Manfredi, M.; Marengo, E.; Giuffrida, M.G.; et al. Thermal processing of insect allergens and IgE cross-recognition in Italian patients allergic to shrimp, house dust mite and mealworm. Food Res. Int. 2021, 148, 110567. [Google Scholar] [CrossRef] [PubMed]
- Jeong, K.Y.; Park, J.W. Insect Allergens on the Dining Table. Curr. Protein Pept. Sci. 2020, 21, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Skotnicka, M.; Karwowska, K.; Kłobukowski, F.; Borkowska, A.; Pieszko, M. Possibilities of the Development of Edible Insect-Based Foods in Europe. Foods 2021, 10, 766. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Barre, A.; Pichereaux, C.; Velazquez, E.; Maudouit, A.; Simplicien, M.; Garnier, L.; Bienvenu, F.; Bienvenu, J.; Burlet-Schiltz, O.; Auriol, C.; et al. Insights into the Allergenic Potential of the Edible Yellow Mealworm (Tenebrio molitor). Foods 2019, 8, 515. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- van Broekhoven, S.; Bastiaan-Net, S.; de Jong, N.W.; Wichers, H.J. Influence of processing and in vitro digestion on the allergic cross-reactivity of three mealworm species. Food Chem. 2016, 196, 1075–1083. [Google Scholar] [CrossRef] [PubMed]
- Sokol, W.N. Grasshopper sensitization in patients allergic to crustaceans, mites, and cockroaches: Should grasshopper-containing products carry a warning? Ann. Allergy Asthma Immunol. 2020, 124, 518–520. [Google Scholar] [CrossRef] [PubMed]
- Purohit, A.; Shao, J.; Degreef, J.M.; van Leeuwen, A.; van Ree, R.; Pauli, G.; de Blay, F. Role of tropomyosin as a cross-reacting allergen in sensitization to cockroach in patients from Martinique (French Caribbean island) with a respiratory allergy to mite and a food allergy to crab and shrimp. Eur. Ann. Allergy Clin. Immunol. 2007, 39, 85–88. [Google Scholar] [PubMed]
- Ribeiro, J.C.; Cunha, L.M.; Sousa-Pinto, B.; Fonseca, J. Allergic risks of consuming edible insects: A systematic review. Mol. Nutr. Food Res. 2017, 62, 1700030. [Google Scholar] [CrossRef] [PubMed]
- Sozener, Z.C.; Ozturk, B.O.; Cerci, P.; Turk, M.; Akin, B.G.; Akdis, M.; Altiner, S.; Ozbey, U.; Ogulur, I.; Mitamura, Y.; et al. Epithelial barrier hypothesis: Effect of the external exposome on the microbiome and epithelial barriers in allergic disease. Allergy 2022, 77, 1418–1449. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Caraballo, L.; Zakzuk, J.; Lee, B.W.; Acevedo, N.; Soh, J.Y.; Sánchez-Borges, M.; Hossny, E.; García, E.; Rosario, N.; Ansotegui, I.; et al. Particularities of allergy in the Tropics. World Allergy Organ. J. 2016, 9, 20. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Muddaluru, V.; Valenta, R.; Vrtala, S.; Schlederer, T.; Hindley, J.; Hickey, P.; Larché, M.; Tonti, E. Comparison of house dust mite sensitization profiles in allergic adults from Canada, Europe, South Africa and USA. Allergy 2021, 76, 2177–2188. [Google Scholar] [CrossRef] [PubMed]
- González-Pérez, R.; Galván-Calle, C.A.; Galán, T.; Poza-Guedes, P.; Sánchez-Machín, I.; Enrique-Calderón, O.M.; Pineda, F. Molecular Signatures of Aeroallergen Sensitization in Respiratory Allergy: A Comparative Study Across Climate-Matched Populations. Int. J. Mol. Sci. 2024, 26, 284. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Goodess, C.M.; Giorgi, F.; Hamaoui-Laguel, L.; Semenov, M.A.; Solmon, F.; Storkey, J.; Vautard, R.; Epstein, M.M. Climate Change and Future Pollen Allergy in Europe. Environ. Heal. Perspect. 2017, 125, 385–391. [Google Scholar]
- Beggs, P.J.; Clot, B.; Sofiev, M.; Johnston, F.H. Climate change, airborne allergens, and three translational mitigation approaches. eBioMedicine 2023, 93, 104478. [Google Scholar] [CrossRef] [PubMed]
- Plume Labs. Available online: https://plumelabs.com/en/air/ (accessed on 16 April 2025).
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef]
- Ansotegui, I.J.; Melioli, G.; Canonica, G.W.; Caraballo, L.; Villa, E.; Ebisawa, M.; Passalacqua, G.; Savi, E.; Ebo, D.; Gómez, R.M.; et al. IgE allergy diagnostics and other relevant tests in allergy, a World Allergy Organization position paper. World Allergy Organ. J. 2020, 13, 100080. [Google Scholar] [CrossRef]
- Santos, A.F.; Riggioni, C.; Agache, I.; Akdis, C.A.; Akdis, M.; Alvarez-Perea, A.; Alvaro-Lozano, M.; Ballmer-Weber, B.; Barni, S.; Beyer, K.; et al. EAACI guidelines on the diagnosis of IgE-mediated food allergy. Allergy 2023, 78, 3057–3076. [Google Scholar] [CrossRef] [PubMed]
- Kleine-Tebbe, J.; Jakob, T. Molecular allergy diagnostics using IgE singleplex determinations: Methodological and practical considerations for use in clinical routine: Part 18 of the Series Molecular Allergology. Allergo J. Int. 2015, 24, 185–197. [Google Scholar] [CrossRef]
- Kleine-Tebbe, J.; Jappe, U. Molecular allergy diagnostic tests: Development and relevance in clinical practice. Allergologie 2017, 1, 169–189. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Platteel, A.C.; van der Pol, P.; Murk, J.-L.; Verbrugge-Bakker, I.; Hack-Steemers, M.; Roovers, T.H.; Heron, M. A comprehensive comparison between ISAC and ALEX2 multiplex test systems. Clin. Chem. Lab. Med. 2022, 60, 1046–1052. [Google Scholar] [CrossRef] [PubMed]
- González-Pérez, R.; Poza-Guedes, P.; Pineda, F.; Galán, T.; Mederos-Luis, E.; Abel-Fernández, E.; Martínez, M.J.; Sánchez-Machín, I. Molecular Mapping of Allergen Exposome among Different Atopic Phenotypes. Int. J. Mol. Sci. 2023, 24, 10467. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Soil-Transmitted Helminthiases: Eliminating Soil-Transmitted Helminthiases as a Public Health Problem in Children: Progress Report 2001–2010 and Strategic Plan 2011–2020. Available online: https://apps.who.int/iris/bitstream/handle/10665/44804/9789241503129_eng.pdf (accessed on 13 February 2025).
- Bousquet, J.; Schünemann, H.J.; Togias, A.; Bachert, C.; Erhola, M.; Hellings, P.W.; Klimek, L.; Pfaar, O.; Wallace, D.; Ansotegui, I.; et al. Allergic Rhinitis and Its Impact on Asthma Working Group. Next-generation Allergic Rhinitis and Its Impact on Asthma (ARIA) guidelines for allergic rhinitis based on Grading of Recommendations Assessment, Development and Evaluation (GRADE) and real-world evidence. J. Allergy Clin. Immunol. 2020, 145, 70–80.e3. [Google Scholar]
- 2022 GINA Main Report. Available online: https://ginasthma.org/gina-reports/ (accessed on 15 March 2025).
- Bojcukova, J.; Vlas, T.; Forstenlechner, P.; Panzner, P. Comparison of two multiplex arrays in the diagnostics of allergy. Clin. Transl. Allergy 2019, 9, 31. [Google Scholar] [CrossRef]
- Lis, K.; Bartuzi, Z. Selected Technical Aspects of Molecular Allergy Diagnostics. Curr. Issues Mol. Biol. 2023, 45, 5481–5493. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Nösslinger, H.; Mair, E.; Oostingh, G.J.; Ahlgrimm-Siess, V.; Ringauf, A.; Lang, R. Multiplex Assays in Allergy Diagnosis: Allergy Explorer 2 versus ImmunoCAP ISAC E112i. Diagnostics 2024, 14, 976. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Altmann, F. Coping with cross-reactive carbohydrate determinants in allergy diagnosis. Allergo J. Int. 2016, 25, 98–105. [Google Scholar] [CrossRef]
- Chen, H.; Jiang, Q.; Yang, Y.; Zhang, W.; Yang, L.; Zhu, R. Cross-Reacting Carbohydrate Determinants Inhibitor Can Improve the Diagnostic Accuracy in Pollen and Food Allergy. J. Asthma Allergy 2022, 15, 713–725. [Google Scholar] [CrossRef]
- Omuse, E.R.; Tonnang, H.E.Z.; Yusuf, A.A.; Machekano, H.; Egonyu, J.P.; Kimathi, E.; Mohamed, S.F.; Kassie, M.; Subramanian, S.; Onditi, J.; et al. The global atlas of edible insects: Analysis of diversity and commonality contributing to food systems and sustainability. Sci. Rep. 2024, 14, 5045. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Siddiqui, S.A.; Tettey, E.; Yunusa, B.M.; Ngah, N.; Debrah, S.K.; Yang, X.; Fernando, I.; Povetkin, S.N.; Shah, M.A. Legal situation and consumer acceptance of insects being eaten as human food in different nations across the world-A comprehensive review. Compr. Rev. Food Sci. Food Saf. 2023, 22, 4786–4830. [Google Scholar] [CrossRef] [PubMed]
- Sokol, W.N.; Wünschmann, S.; Agah, S. Grasshopper anaphylaxis in patients allergic to dust mite, cockroach, and crustaceans: Is tropomyosin the cause? Ann. Allergy Asthma Immunol. 2017, 119, 91–92. [Google Scholar] [CrossRef] [PubMed]
- Ayuso, R.; Reese, G.; Leong-Kee, S.; Plante, M.; Lehrer, S.B. Molecular basis of arthropod cross-reactivity: IgE-binding cross-reactive epitopes of shrimp, house dust mite and cockroach tropomyosins. Int. Arch. Allergy Immunol. 2002, 129, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Olivieri, B.; Stoenchev, K.V.; Skypala, I.J. Anaphylaxis across Europe: Are pollen food syndrome and lipid transfer protein allergy so far apart? Curr. Opin. Allergy Clin. Immunol. 2022, 22, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Betancor, D.; Gomez-Lopez, A.; Villalobos-Vilda, C.; Nuñez-Borque, E.; Fernández-Bravo, S.; De Las Heras Gozalo, M.; Pastor-Vargas, C.; Esteban, V.; Cuesta-Herranz, J. LTP Allergy Follow-Up Study: Development of Allergy to New Plant Foods 10 Years Later. Nutrients 2021, 13, 2165. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Mahammed, L.L.; Belaid, B.; Berkani, L.M.; Merah, F.; Rahali, S.Y.; Kaci, A.A.; Berkane, I.; Sayah, W.; Allam, I.; Djidjik, R. Shrimp sensitization in house dust mite algerian allergic patients: A single center experience. World Allergy Organ. J. 2022, 15, 100642. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Farioli, L.; Losappio, L.M.; Giuffrida, M.G.; Pravettoni, V.; Micarelli, G.; Nichelatti, M.; Scibilia, J.; Mirone, C.; Cavallarin, L.; Lamberti, C.; et al. Mite-Induced Asthma and IgE Levels to Shrimp, Mite, Tropomyosin, Arginine Kinase, and Der p 10 Are the Most Relevant Risk Factors for Challenge-Proven Shrimp Allergy. Int. Arch. Allergy Immunol. 2017, 174, 133–143. [Google Scholar] [CrossRef] [PubMed]
- del Giudice, M.M.; Dinardo, G.; Klain, A.; D’addio, E.; Bencivenga, C.L.; Decimo, F.; Indolfi, C. Anaphylaxis after Shrimp Intake in a European Pediatric Population: Role of Molecular Diagnostics and Implications for Novel Foods. Children 2023, 10, 1583. [Google Scholar] [CrossRef]
- Giusti, D.; Guemari, A.; Perotin, J.-M.; Fontaine, J.-F.; Libyh, M.T.; Gatouillat, G.; Tabary, T.; Pham, B.-N.; Vitte, J. Molecular allergology: A clinical laboratory tool for precision diagnosis, stratification and follow-up of allergic patients. Clin. Chem. Lab. Med. 2024, 62, 2339–2355. [Google Scholar] [CrossRef] [PubMed]
- Riggioni, C.; Leung, A.S.; Wai, C.Y.; Davies, J.M.; Sompornrattanaphan, M.; Pacharn, P.; Chamani, S.; Brettig, T.; Peters, R.L. Exploring geographical variances in component-resolved diagnosis within the Asia-Pacific region. Pediatr. Allergy Immunol. 2025, 36, e70054. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Mittermann, I.; Lupinek, C.; Wieser, S.; Aumayr, M.; Kuchler, W.W.; Chan, A.W.; Lee, T.H.; Zieglmayer, P. IgE reactivity patterns in Asian and central European cockroach-sensitized patients reveal differences in primary sensitizing allergen sources. J. Allergy Clin. Immunol. Glob. 2022, 1, 145–153. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wangorsch, A.; Jamin, A.; Spiric, J.; Vieths, S.; Scheurer, S.; Mahler, V.; Hofmann, S.C. Allergic Reaction to a Commercially Available Insect Snack Caused by House Cricket (Acheta domesticus) Tropomyosin. Mol. Nutr. Food Res. 2024, 68, e2300420. [Google Scholar] [CrossRef] [PubMed]
- Li, J.C.; Rotter, N.S.; Stieb, E.S.; Stockbridge, J.L.; Theodorakakis, M.D.; Shreffler, W.G. Utility of food allergy thresholds. Ann. Allergy Asthma Immunol. 2023, 132, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Valenta, R.; Karaulov, A.; Niederberger, V.; Gattinger, P.; van Hage, M.; Flicker, S.; Linhart, B.; Campana, R.; Focke-Tejkl, M.; Curin, M.; et al. Molecular Aspects of Allergens and Allergy. Adv. Immunol. 2018, 138, 195–256. [Google Scholar] [CrossRef] [PubMed]
- Sharma, E.; Vitte, J. A systematic review of allergen cross-reactivity: Translating basic concepts into clinical relevance. J. Allergy Clin. Immunol. Glob. 2024, 3, 100230. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Dramburg, S.; Hilger, C.; Santos, A.F.; Vecillas, L.d.L.; Aalberse, R.C.; Acevedo, N.; Aglas, L.; Altmann, F.; Arruda, K.L.; Asero, R.; et al. EAACI Molecular Allergology User’s Guide 2.0. Pediatr. Allergy Immunol. 2023, 34, e13854. [Google Scholar] [CrossRef] [PubMed]
- De Marchi, L.; Wangorsch, A.; Zoccatelli, G. Allergens from Edible Insects: Cross-reactivity and Effects of Processing. Curr. Allergy Asthma Rep. 2021, 21, 35. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Scala, E.; Abeni, D.; Villella, V.; ViIlalta, D.; Cecchi, L.; Caprini, E.; Asero, R. Investigating Novel Food Sensitization: A Real-Life Prevalence Study of Cricket, Locust, and Mealworm IgE-Reactivity in Naïve allergic Individuals. J. Investig. Allergol. Clin. Immunol. 2024, 35, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Wong, L.; Huang, C.H.; Lee, B.W. Shellfish and House Dust Mite Allergies: Is the Link Tropomyosin? Allergy Asthma Immunol. Res. 2016, 8, 101–106. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Emilia, M.; Magdalena, C.; Weronika, G.; Julia, W.; Danuta, K.; Jakub, S.; Bożena, C.; Krzysztof, K. IgE-based analysis of sensitization and cross-reactivity to yellow mealworm and edible insect allergens before their widespread dietary introduction. Sci. Rep. 2025, 15, 1466. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Linacero, R.; Cuadrado, C. New Research in Food Allergen Detection. Foods 2022, 11, 1520. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Retzinger, A.C.; Retzinger, G.S. The Acari Hypothesis, II: Interspecies Operability of Pattern Recognition Receptors. Pathogens 2021, 10, 1220. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Trompette, A.; Divanovic, S.; Visintin, A.; Blanchard, C.; Hegde, R.S.; Madan, R.; Thorne, P.S.; Wills-Karp, M.; Gioannini, T.L.; Weiss, J.P.; et al. Allergenicity resulting from functional mimicry of a Toll-like receptor complex protein. Nature 2009, 457, 585–588. [Google Scholar] [CrossRef] [PubMed]
- Mueller, G.A.; Edwards, L.L.; Aloor, J.J.; Fessler, M.B.; Glesner, J.; Pomés, A.; Chapman, M.D.; London, R.E.; Pedersen, L.C. The structure of the dust mite allergen Der p 7 reveals similarities to innate immune proteins. J. Allergy Clin. Immunol. 2010, 125, 909–917.e4. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Pali-Schöll, I.; Meinlschmidt, P.; Larenas-Linnemann, D.; Purschke, B.; Hofstetter, G.; Rodríguez-Monroy, F.A.; Einhorn, L.; Mothes-Luksch, N.; Jensen-Jarolim, E.; Jäger, H. Edible insects: Cross-recognition of IgE from crustacean- and house dust mite allergic patients, and reduction of allergenicity by food processing. World Allergy Organ. J. 2019, 12, 100006. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Popescu, F.D. Cross-reactivity between aeroallergens and food allergens. World J. Methodol. 2015, 5, 31–50. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Shroba, J.; Rath, N.; Barnes, C. Possible Role of Environmental Factors in the Development of Food Allergies. Clin. Rev. Allergy Immunol. 2018, 57, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Belluco, S.; Losasso, C.; Maggioletti, M.; Alonzi, C.C.; Paoletti, M.G.; Ricci, A. Edible Insects in a Food Safety and Nutritional Perspective: A Critical Review. Compr. Rev. Food Sci. Food Saf. 2013, 12, 296–313. [Google Scholar] [CrossRef]
- Conway, A.; Jaiswal, S.; Jaiswal, A.K. The Potential of Edible Insects as a Safe, Palatable, and Sustainable Food Source in the European Union. Foods 2024, 13, 387. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhao, L.; Zhang, Y.; Zhang, S.; Zhang, L.; Lan, F. The effect of immunotherapy on cross-reactivity between house dust mite and other allergens in house dust mite -sensitized patients with allergic rhinitis. Expert Rev. Clin. Immunol. 2021, 17, 969–975. [Google Scholar] [CrossRef] [PubMed]
- Verhoeckx, K.C.; van Broekhoven, S.; den Hartog-Jager, C.F.; Gaspari, M.; de Jong, G.A.; Wichers, H.J.; van Hoffen, E.; Houben, G.F.; Knulst, A.C. House dust mite (Der p 10) and crustacean allergic patients may react to food containing Yellow mealworm proteins. Food Chem. Toxicol. 2014, 65, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Broekman, H.C.H.P.; Knulst, A.C.; de Jong, G.; Gaspari, M.; Jager, C.F.D.H.; Houben, G.F.; Verhoeckx, K.C.M. Is mealworm or shrimp allergy indicative for food allergy to insects? Mol. Nutr. Food Res. 2017, 61, 1601061. [Google Scholar] [CrossRef] [PubMed]
- Gałęcki, R.; Bakuła, T.; Gołaszewski, J. Foodborne Diseases in the Edible Insect Industry in Europe-New Challenges and Old Problems. Foods 2023, 12, 770. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Abro, Z.; Sibhatu, K.T.; Fetene, G.M.; Alemu, M.H.; Tanga, C.M.; Sevgan, S.; Kassie, M. Global review of consumer preferences and willingness to pay for edible insects and derived products. Glob. Food Secur. 2025, 44, 100834. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Quintieri, L.; Nitride, C.; De Angelis, E.; Lamonaca, A.; Pilolli, R.; Russo, F.; Monaci, L. Alternative Protein Sources and Novel Foods: Benefits, Food Applications and Safety Issues. Nutrients 2023, 15, 1509. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Sicherer, S.H.; Sampson, H.A. Food allergy. J. Allergy Clin. Immunol. 2010, 125, S116–S125. [Google Scholar] [CrossRef] [PubMed]
- Schussler, E.; Kattan, J. Allergen Component Testing in the Diagnosis of Food Allergy. Curr. Allergy Asthma Rep. 2015, 15, 55. [Google Scholar] [CrossRef] [PubMed]
| Characteristics | (n = 138) |
|---|---|
| Age (y.o.) median (range) | 17 (3–75) |
| Sex (F/M) | 43/95 |
| Food and respiratory allergy | 80 (58%) |
| Food allergy | 22 (15.9%) |
| Seafood allergy | 40 (50%) |
| Respiratory allergy | 36 (26%) |
| Allergic rhinitis | 21 (58.3%) |
| Allergic rhinitis and asthma | 15 (41.6% |
| Total IgE (IU/mL) median (range) | 532 (54–2500) |
| Family history of atopy (%) | 105 (77.08) |
| Pan-Allergens in 138 Subjects Sensitized to at Least One Edible Insect | Acheta domesticus (n = 101) | Locusta migratoria (n = 110) | Tenebrio molitor (n = 80) |
|---|---|---|---|
| Tropomyosin (any) molecules in 88/138 subjects (63.76%) | |||
| Ani s 3 (n = 76) | 69 (68.31) | 67 (60.9) | 58 (72.5) |
| Blo t 10 (n = 79) | 71 (70.29) | 68 (61.81) | 62 (77.5) |
| Der p 10 (n = 64) | 62 (61.38) | 58 (52.72) | 57 (71.25) |
| Per a 7 (n = 68) | 64 (63.36) | 60 (54.54) | 58 (72.5) |
| Pen m 1 (n = 63) | 62 (61.38) | 58 (52.72) | 54 (67.5) |
| Arginine kinase (any) molecules in 46/138 subjects (33.33%) | |||
| Bla g 9 (n = 36) | 27 (26.73) | 33 (30) | 22 (27.5) |
| Der p 20 (n = 39) | 32 (31.68) | 35 (31.81) | 27 (33.75) |
| Pen m 2 (n = 26) | 23 (22.77) | 25 (22.72) | 23 (28.75) |
| Paramyosin Der p 11 in 1/138 subjects (0.72%) | 1 (0.99) | 1 (0.9) | 1 (1.25) |
| Troponin-C Cra c 6 in 40/138 subjects (28.98%) | 34 (33.66) | 37 (33.63) | 31 (76.25) |
| Myosin light chain Pen m 3 in 9/138 subjects (6.52%) | 8 (7.92) | 8 (7.27) | 8 (10) |
| Sarcoplasmic calcium-binding protein Pen m 4 in 12/138 subjects (8.69%) | 8 (7.92) | 10 (9.09) | 8 (10) |
| None (32/138 (23.18%) subjects) | 24 (23.76) | 22 (20) | 10 (10) |
| Mite Allergen | Median sIgE M (IQR) | No. of Sensitized Patients (%) |
|---|---|---|
| Der f 2 | 17.41 (37.21) | 32 (88.88) |
| Der p 2 | 24.19 (41.42) | 31 (86.11) |
| Der p 1 | 9.46 (27.31) | 30 (83.33) |
| Der p 23 | 7.44 (24.89) | 28 (77.77) |
| Der f 1 | 2.47 (9.95) | 25 (69.44) |
| Der p 5 | 3.45 (27.02) | 22 (61.11) |
| Der p 7 | 2.53 (25.03) | 21 (58.33) |
| Blo t 21 | 0.2 (10.64) | 19 (52.77) |
| Gly d 2 | 0.11 (3.71) | 17 (47.22) |
| Tyr p 2 | 0.1 (1.75) | 15 (41.66) |
| Der p 21 | 0.1 (9.83) | 14 (38.88) |
| Blo t 5 | 0.1 (6.84) | 14 (38.88) |
| Lep d 2 | 0.1 (2.19) | 13 (36.11) |
| Der p 20 | 0.1 (0.0) | 3 (8.33) |
| Der p 10 | 0.1 (0.0) | 1 (2.77) |
| Blo t 10 | 0.1 (0.0) | 2 (5.55) |
| Der p 11 | 0.1 (0.0) | 0 (0) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Pérez, R.; Poza-Guedes, P.; Figueiras-Rincón, M.A.; Colque-Bayona, M.; Sánchez-Machín, I. The Allergy Crossroads of Subtropical Regions: Mites, Crustaceans, and the Rise of Edible Insects. Nutrients 2025, 17, 1405. https://doi.org/10.3390/nu17091405
González-Pérez R, Poza-Guedes P, Figueiras-Rincón MA, Colque-Bayona M, Sánchez-Machín I. The Allergy Crossroads of Subtropical Regions: Mites, Crustaceans, and the Rise of Edible Insects. Nutrients. 2025; 17(9):1405. https://doi.org/10.3390/nu17091405
Chicago/Turabian StyleGonzález-Pérez, Ruperto, Paloma Poza-Guedes, Manuel Alberto Figueiras-Rincón, Mónica Colque-Bayona, and Inmaculada Sánchez-Machín. 2025. "The Allergy Crossroads of Subtropical Regions: Mites, Crustaceans, and the Rise of Edible Insects" Nutrients 17, no. 9: 1405. https://doi.org/10.3390/nu17091405
APA StyleGonzález-Pérez, R., Poza-Guedes, P., Figueiras-Rincón, M. A., Colque-Bayona, M., & Sánchez-Machín, I. (2025). The Allergy Crossroads of Subtropical Regions: Mites, Crustaceans, and the Rise of Edible Insects. Nutrients, 17(9), 1405. https://doi.org/10.3390/nu17091405