
Vaccinium oldhamii Fruit Inhibits Lipid Accumulation in 3T3-L1 Cells and Diet-Induced Obese Animals


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Extract and Fraction Preparation
2.2. Cell Culture
2.3. Cell Viability
2.4. Lipid Accumulation
2.5. Animals
2.6. Blood Biochemical Parameters
2.7. Western Blot
2.8. H&E Staining
2.9. HPLC Quantification of Malvidin-3-O-Galactoside and Chlorogenic Acid
2.10. Statistical Analysis
3. Results
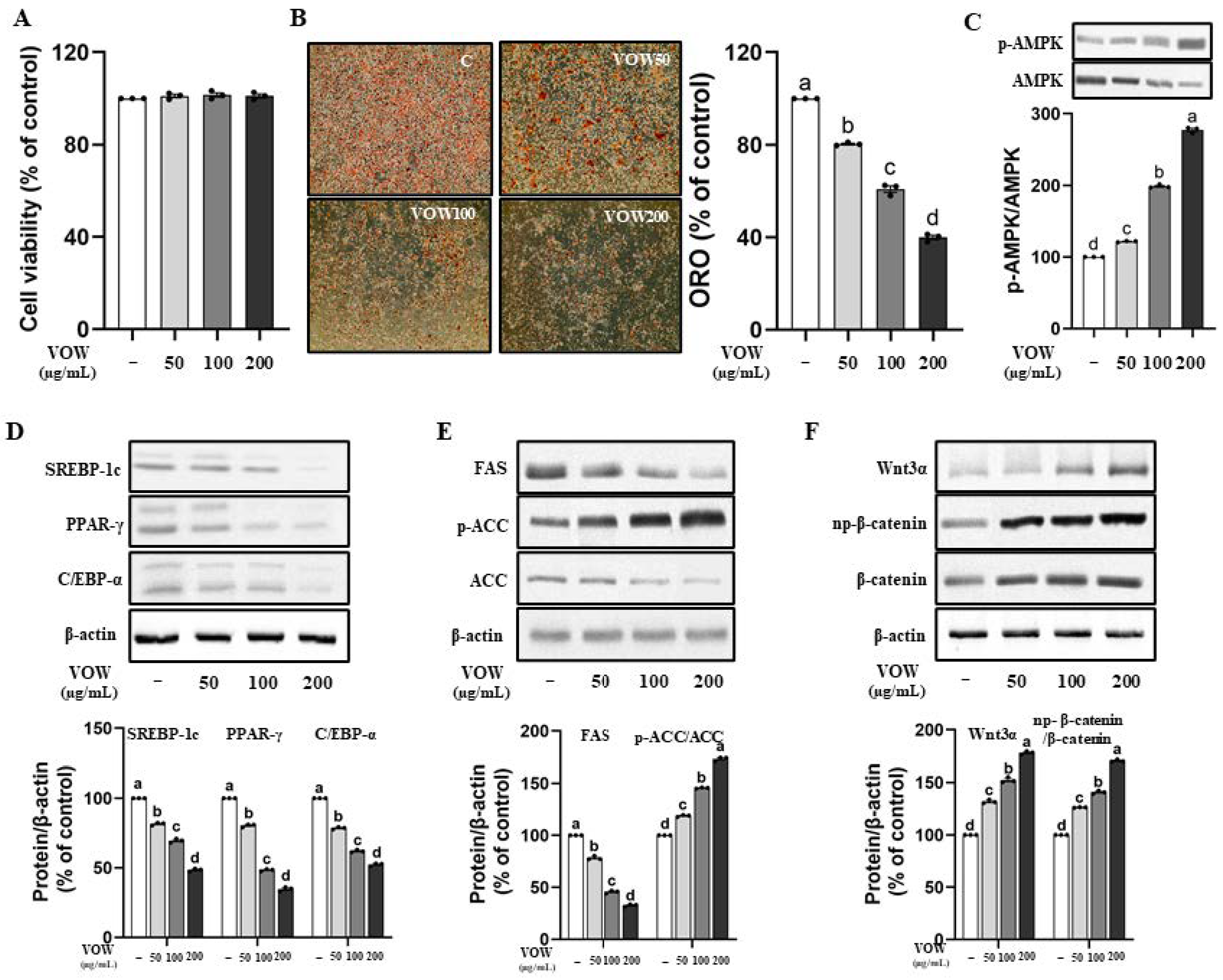
3.1. VOW Suppresses Lipid Accumulation and the Expression of Adipogenic Proteins in 3T3-L1 Cells
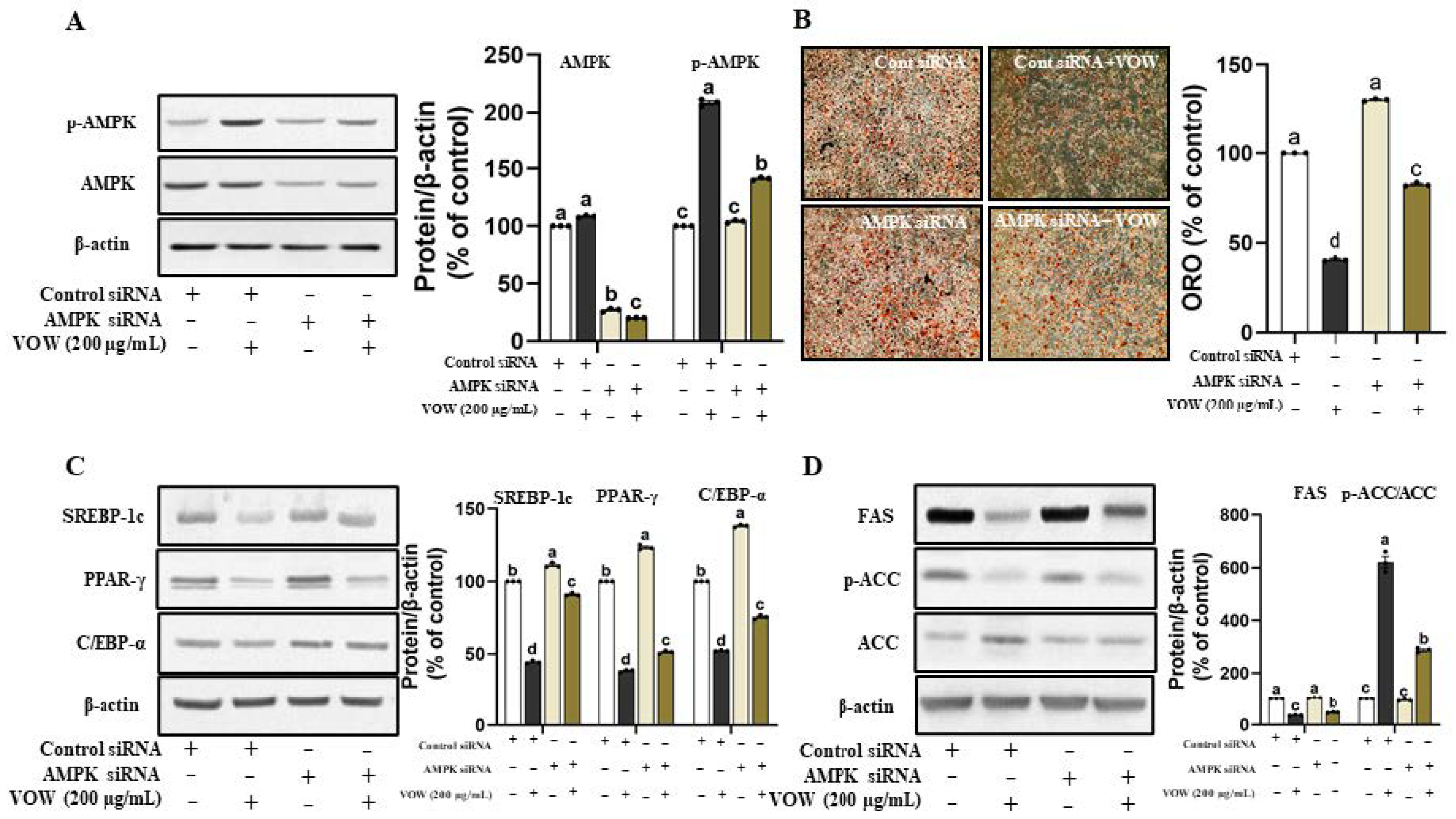
3.2. VOW-Induced Downregulation of Lipid Accumulation Is Associated with the AMPK Activation
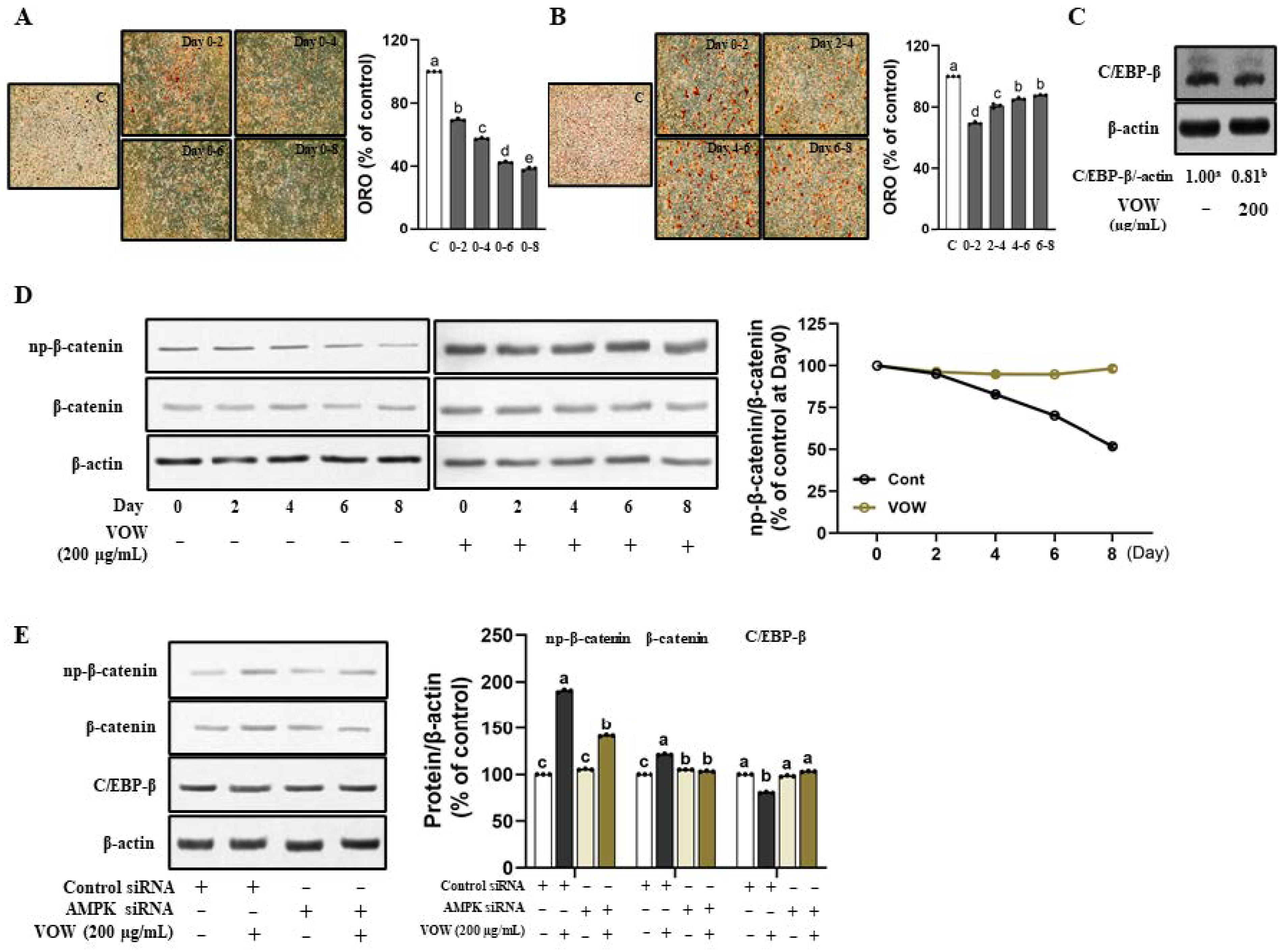
3.3. The Influence of VOW Depends on Timing and Period of Treatment
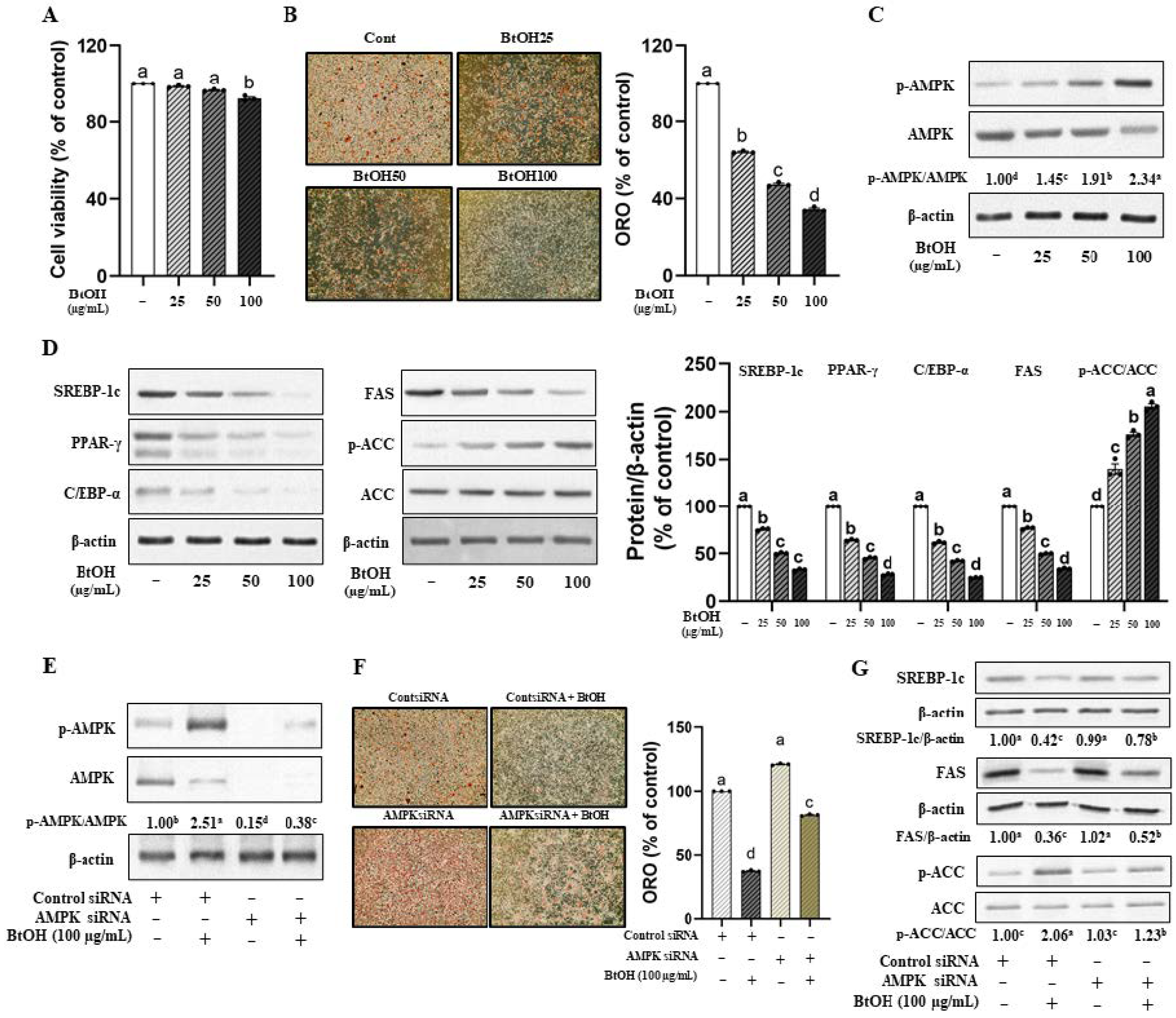
3.4. Butanol Fraction of VOW Is the Most Effective Among Various Solvent Fractions in Suppressing Lipid Accumulation in 3T3-L1 Cells
3.5. VOW Suppresses High-Fat and High-Sucrose Diet (HFHSD)-Induced Effects in Body Weight, Adipose Tissue Weight, and Serum Lipids
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Calle, E.E.; Kaaks, R. Overweight, obesity and cancer: Epidemiological evidence and proposed mechanisms. Nat. Rev. Cancer 2004, 4, 579–591. [Google Scholar] [CrossRef]
- Swinburn, B.A.; Sacks, G.; Hall, K.D.; McPherson, K.; Finegood, D.T.; Moodie, M.L.; Gortmaker, S.L. The global obesity pandemic: Shaped by global drivers and local environments. Lancet 2011, 378, 804–814. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.-H.; Lee, J.-H.; Kim, J.-W.; Cho, J.H.; Choi, Y.-H.; Ko, S.-H.; Zimmet, P.; Son, H.-Y. Epidemic obesity and type 2 diabetes in Asia. Lancet 2006, 368, 1681–1688. [Google Scholar] [CrossRef] [PubMed]
- Prins, J.B.; O’rahilly, S. Regulation of adipose cell number in man. Clin. Sci. 1997, 92, 3–11. [Google Scholar] [CrossRef]
- Warnke, I.; Goralczyk, R.; Fuhrer, E.; Schwager, J. Dietary constituents reduce lipid accumulation in murine C3H10 T1/2 adipocytes: A novel fluorescent method to quantify fat droplets. Nutr. Metab. 2011, 8, 30. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Y.; Della-Fera, M.A.; Hartzell, D.L.; Nelson-Dooley, C.; Hausman, D.B.; Baile, C.A. Esculetin Induces Apoptosis and inhibits adipogenesis in 3T3-L1 cells. Obesity 2006, 14, 1691–1699. [Google Scholar] [CrossRef]
- Hausman, D.; DiGirolamo, M.; Bartness, T.; Hausman, G.; Martin, R. The biology of white adipocyte proliferation. Obes. Rev. 2001, 2, 239–254. [Google Scholar] [CrossRef]
- Barzilai, N.; Gupta, G. Revisiting the role of fat mass in the life extension induced by caloric restriction. J. Gerontol. Ser. A Biomed. Sci. Med. Sci. 1999, 54, B89–B96. [Google Scholar] [CrossRef]
- Rohm, T.V.; Meier, D.T.; Olefsky, J.M.; Donath, M.Y. Inflammation in obesity, diabetes, and related disorders. Immunity 2022, 55, 31–55. [Google Scholar] [CrossRef]
- Wu, H.; Ballantyne, C.M. Metabolic inflammation and insulin resistance in obesity. Circ. Res. 2020, 126, 1549–1564. [Google Scholar] [CrossRef]
- Mohseni, R.; Teimouri, M.; Safaei, M.; Arab Sadeghabadi, Z. AMP-activated protein kinase is a key regulator of obesity-associated factors. Cell Biochem. Funct. 2023, 41, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Thrimawithana, T.; Shukla, R.; Adhikari, B. Managing obesity through natural polyphenols: A review. Future Foods 2020, 1, 100002. [Google Scholar] [CrossRef]
- Block, J.P.; Choudhry, N.K.; Carpenter, D.P.; Fischer, M.A.; Brennan, T.A.; Tong, A.Y.; Matlin, O.S.; Shrank, W.H. Time series analyses of the effect of FDA communications on use of prescription weight loss medications. Obesity 2014, 22, 943–949. [Google Scholar] [CrossRef]
- Filippatos, T.D.; Derdemezis, C.S.; Gazi, I.F.; Nakou, E.S.; Mikhailidis, D.P.; Elisaf, M.S. Orlistat-associated adverse effects and drug interactions: A critical review. Drug Saf. 2008, 31, 53–65. [Google Scholar] [CrossRef]
- González-Castejón, M.; Rodriguez-Casado, A. Dietary phytochemicals and their potential effects on obesity: A review. Pharmacol. Res. 2011, 64, 438–455. [Google Scholar] [CrossRef]
- Lee, J.H.; Lee, K.T.; Yang, J.H.; Baek, N.I.; Kim, D.K. Acetylcholinesterase inhibitors from the twigs of Vaccinium oldhami Miquel. Arch. Pharmacal Res. 2004, 27, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.-J.; Yim, E.-Y.; Song, G.; Kim, G.-O.; Hyun, C.-G. Inhibition of nitric oxide production in lipopolysaccharide-activated RAW 264.7 macrophages by Jeju plant extracts. Interdiscip. Toxicol. 2009, 2, 245–249. [Google Scholar] [CrossRef]
- Oh, S.-J.; Koh, S.-C. Screening of Antioxidative Activity and ${\alpha} $-Amylase Inhibitory Activity in Angiosperm Plants Native to Jeju Island. Korean J. Plant Resour. 2009, 22, 71–77. [Google Scholar]
- Tsuda, H.; Kunitake, H.; Kawasaki-Takaki, R.; Nishiyama, K.; Yamasaki, M.; Komatsu, H.; Yukizaki, C. Antioxidant activities and anti-cancer cell proliferation properties of Natsuhaze (Vaccinium oldhamii Miq.), Shashanbo (V. bracteatum Thunb.) and blueberry cultivars. Plants 2013, 2, 57–71. [Google Scholar] [CrossRef]
- Baba, T.; Hirose, D.; Sasaki, N.; Watanabe, N.; Kobayashi, N.; Kurashige, Y.; Karimi, F.; Ban, T. Mycorrhizal formation and diversity of endophytic fungi in hair roots of Vaccinium oldhamii Miq. in Japan. Microbes Environ. 2016, 31, 186–189. [Google Scholar] [CrossRef]
- Rhyu, J.; Kim, M.S.; You, M.-K.; Bang, M.-A.; Kim, H.-A. Pear pomace water extract inhibits adipogenesis and induces apoptosis in 3T3-L1 adipocytes. Nutr. Res. Pract. 2014, 8, 33–39. [Google Scholar] [CrossRef]
- Forbes-Hernandez, T.Y.; Afrin, S.; Cianciosi, D.; Manna, P.P.; Zhang, J.; Gasparrini, M.; Reboredo-Rodriguez, P. Strawberry extract attenuates oxidative stress in 3T3-L1 cells. J. Berry Res. 2018, 8, 193–203. [Google Scholar] [CrossRef]
- Dludla, P.V.; Jack, B.; Viraragavan, A.; Pheiffer, C.; Johnson, R.; Louw, J.; Muller, C.J. A dose-dependent effect of dimethyl sulfoxide on lipid content, cell viability and oxidative stress in 3T3-L1 adipocytes. Toxicol. Rep. 2018, 5, 1014–1020. [Google Scholar] [CrossRef]
- You, M.-K.; Go, G.-W.; Kim, H.-J.; Rhyu, J.; Kim, H.-A. Pear pomace water extract reduces adiposity in vivo and in vitro by activating the AMPK-dependent pathway. Asian Pac. J. Trop. Biomed. 2020, 10, 208–215. [Google Scholar]
- Guo, L.; Li, X.; Tang, Q.-Q. Transcriptional regulation of adipocyte differentiation: A central role for CCAAT/enhancer-binding protein (C/EBP) β. J. Biol. Chem. 2015, 290, 755–761. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-J.; You, M.-K.; Wang, Z.; Kim, H.-A. Red pepper seed inhibits differentiation of 3T3-L1 cells during the early phase of adipogenesis via the activation of AMPK. Am. J. Chin. Med. 2018, 46, 107–118. [Google Scholar] [CrossRef]
- Donati, G.; Proserpio, V.; Lichtenberger, B.M.; Natsuga, K.; Sinclair, R.; Fujiwara, H.; Watt, F.M. Epidermal Wnt/β-catenin signaling regulates adipocyte differentiation via secretion of adipogenic factors. Proc. Natl. Acad. Sci. USA 2014, 111, E1501–E1509. [Google Scholar] [CrossRef]
- Chen, T.-x.; Cheng, X.-y.; Wang, Y.; Yin, W. Toosendanin inhibits adipogenesis by activating Wnt/β-catenin signaling. Sci. Rep. 2018, 8, 4626. [Google Scholar] [CrossRef]
- Bagchi, D.P.; Nishii, A.; Li, Z.; DelProposto, J.B.; Corsa, C.A.; Mori, H.; Hardij, J.; Learman, B.S.; Lumeng, C.N.; MacDougald, O.A. Wnt/β-catenin signaling regulates adipose tissue lipogenesis and adipocyte-specific loss is rigorously defended by neighboring stromal-vascular cells. Mol. Metab. 2020, 42, 101078. [Google Scholar] [CrossRef]
- De Winter, T.J.; Nusse, R. Running against the Wnt: How Wnt/β-catenin suppresses adipogenesis. Front. Cell Dev. Biol. 2021, 9, 627429. [Google Scholar] [CrossRef]
- Moldes, M.; Zuo, Y.; Morrison, R.F.; Silva, D.; Park, B.-H.; Liu, J.; Farmer, S.R. Peroxisome-proliferator-activated receptor γ suppresses Wnt/β-catenin signalling during adipogenesis. Biochem. J. 2003, 376, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Bennett, C.N.; Ross, S.E.; Longo, K.A.; Bajnok, L.; Hemati, N.; Johnson, K.W.; Harrison, S.D.; MacDougald, O.A. Regulation of Wnt signaling during adipogenesis. J. Biol. Chem. 2002, 277, 30998–31004. [Google Scholar] [CrossRef]
- Li, H.-X.; Luo, X.; Liu, R.-X.; Yang, Y.-J.; Yang, G.-S. Roles of Wnt/β-catenin signaling in adipogenic differentiation potential of adipose-derived mesenchymal stem cells. Mol. Cell. Endocrinol. 2008, 291, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, H.; Zuo, Y.; Farmer, S.R. Functional interaction between peroxisome proliferator-activated receptor γ and β-catenin. Mol. Cell. Biol. 2006, 26, 5827–5837. [Google Scholar] [CrossRef]
- Zhao, J.; Yue, W.; Zhu, M.J.; Sreejayan, N.; Du, M. AMP-activated protein kinase (AMPK) cross-talks with canonical Wnt signaling via phosphorylation of β-catenin at Ser 552. Biochem. Biophys. Res. Commun. 2010, 395, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Kim, D.; Kee, S.H. Metformin-activated AMPK regulates β-catenin to reduce cell proliferation in colon carcinoma RKO cells. Oncol. Lett. 2019, 17, 2695–2702. [Google Scholar] [CrossRef]
- Xu, Y.; Ke, H.; Li, Y.; Xie, L.; Su, H.; Xie, J.; Mo, J.; Chen, W. Malvidin-3-O-glucoside from blueberry ameliorates nonalcoholic fatty liver disease by regulating transcription factor EB-mediated lysosomal function and activating the Nrf2/ARE signaling pathway. J. Agric. Food Chem. 2021, 69, 4663–4673. [Google Scholar] [CrossRef]
- Tie, F.; Ding, J.; Gao, Y.; Wang, H. Chlorogenic Acid and its Isomers Attenuate NAFLD by Mitigating Lipid Accumulation in Oleic Acid-Induced HepG2 Cells and High-Fat Diet-Fed Zebrafish. Chem. Biodivers. 2024, 21, e202400564. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, C.; Zhai, M.; Yu, T.; Pei, M.; Du, P.; Li, A.; Yan, J.; Li, C.; Zhang, G. Effect of chlorogenic acid on lipid metabolism in 3T3-L1 cells induced by oxidative stress. Food Biosci. 2023, 51, 102330. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.-H.; You, M.; Kim, H.-A. Vaccinium oldhamii Fruit Inhibits Lipid Accumulation in 3T3-L1 Cells and Diet-Induced Obese Animals. Nutrients 2025, 17, 1346. https://doi.org/10.3390/nu17081346
Lee Y-H, You M, Kim H-A. Vaccinium oldhamii Fruit Inhibits Lipid Accumulation in 3T3-L1 Cells and Diet-Induced Obese Animals. Nutrients. 2025; 17(8):1346. https://doi.org/10.3390/nu17081346
Chicago/Turabian StyleLee, Young-Hyeon, Mikyoung You, and Hyeon-A Kim. 2025. "Vaccinium oldhamii Fruit Inhibits Lipid Accumulation in 3T3-L1 Cells and Diet-Induced Obese Animals" Nutrients 17, no. 8: 1346. https://doi.org/10.3390/nu17081346
APA StyleLee, Y.-H., You, M., & Kim, H.-A. (2025). Vaccinium oldhamii Fruit Inhibits Lipid Accumulation in 3T3-L1 Cells and Diet-Induced Obese Animals. Nutrients, 17(8), 1346. https://doi.org/10.3390/nu17081346