
Mechanism of Green Tea Peptides in Lowering Blood Pressure and Alleviating Renal Injury Induced by Hypertension Through the Ang II/TGF-β1/SMAD Signaling Pathway

 ,
, Highlights
- Green tea peptides exert protective effects through hypotensive, antifibrotic, and anti-inflammatory effects.
- Green tea peptides exert antihypertensive effects by inhibiting the activity of angiotensin-converting enzyme (ACE) and thereby reducing the production of angiotensinⅡ(Ang Ⅱ).
- Green tea peptides attenuate renal fibrosis by modulating TGF-β1/SMAD signaling and blocking EMT activation.
- Green tea peptides alleviate renal inflammation by modulating MyD88/NF-κB p65/iNOS signaling.
Abstract

1. Introduction
2. Materials and Methods
2.1. Green Tea Peptide Preparation
2.2. Animals and Treatment
2.3. Blood Pressure Measurements
2.4. Renal Function and Histopathology
2.5. Real-Time Quantitative Polymerase Chain (RT-qPCR) Analysis
2.6. Western Blotting
2.7. Statistical Analysis
3. Results
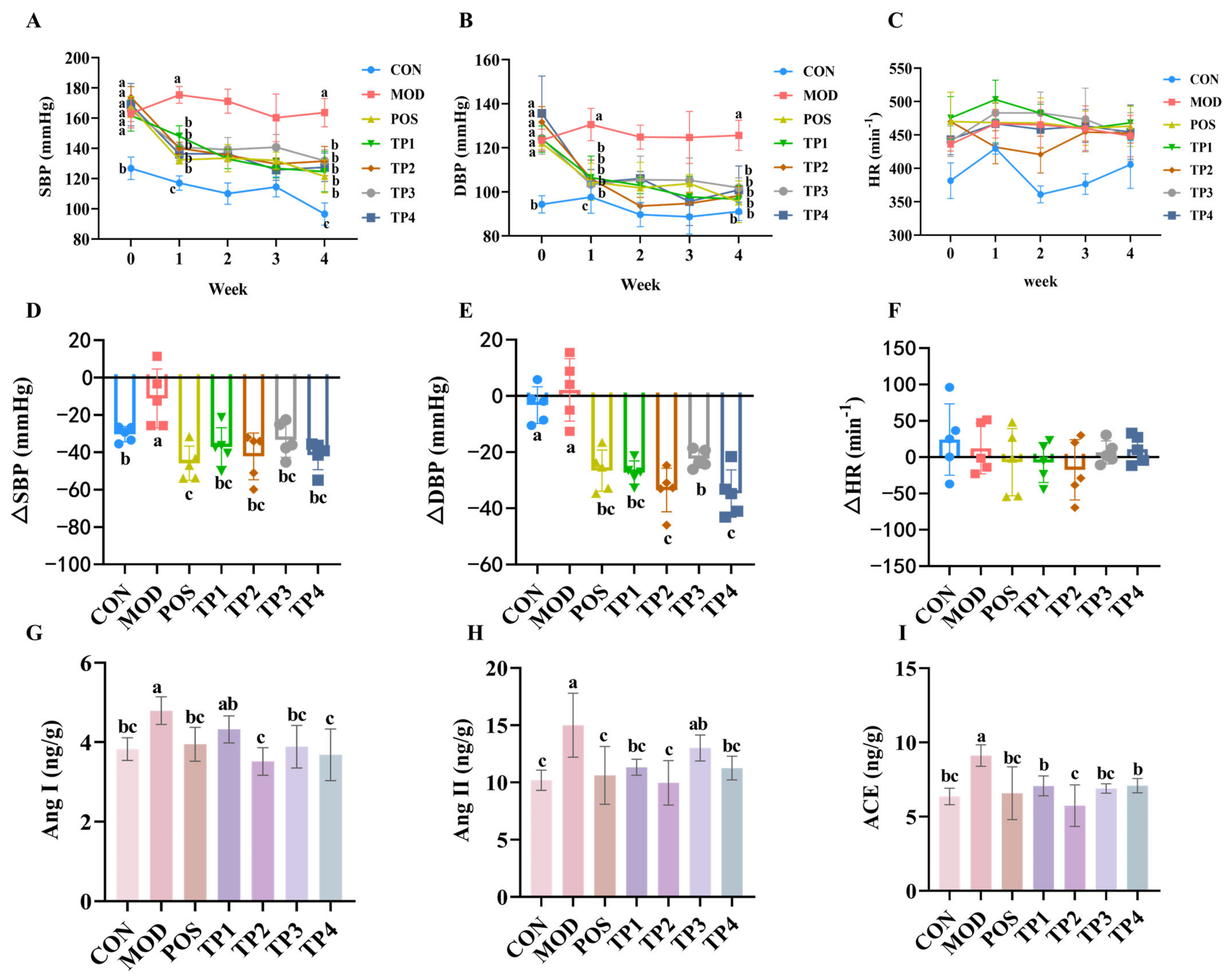
3.1. Green Tea Peptide Attenuated Hypertension in SHRs
3.2. Green Tea Peptide Ameliorated Renal Injury in SHRs
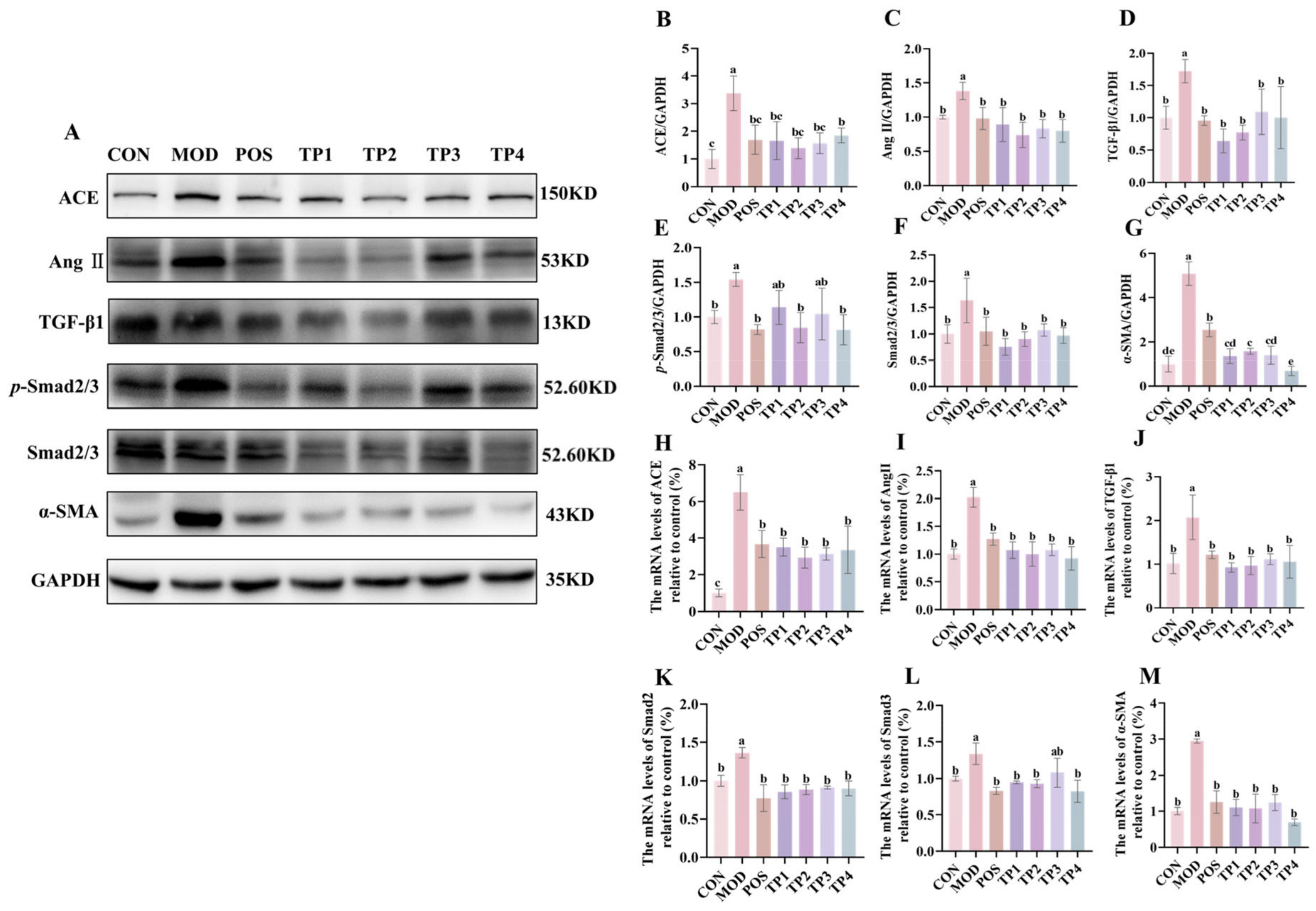
3.3. Green Tea Peptides Through Ang II/TGF-β1/SMAD Pathway to Lower Blood Pressure and Attenuated Renal Fibrosis
3.4. Green Tea Peptide Relieves Kidney Inflammation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| TP | Tea peptides |
| SHR | Spontaneously hypertensive rat |
| WKY | Wistar-Kyoto rat |
| SBP | Systolic blood pressure |
| DBP | Diastolic blood pressure |
| HR | Heart rate |
| ACE | Angiotensin-converting enzyme |
| Ang I | Angiotensin I |
| Ang II | Angiotensin II |
| CRE | Creatinine |
| BUN | Blood urea nitrogen |
| p-Smad2/3 | Phosphorylated-Smad2/3 |
| Smad2/3 | Sma- and Mad-related proteins 2/3 |
| TGF-β1 | Transforming growth factor-β1 |
| α-SMA | α-smooth muscle actin |
| MyD88 | Medullary differentiation factor 88 |
| NF-κB p65 | Nuclear factor-κB p65 |
| iNOS | inducible nitric oxide synthase |
| EMT | Epithelial-to-mesenchymal transition |
| ESRD | End-stage renal disease |
| RAAS | Renin–angiotensin–aldosterone system |
References
- Zhang, C.; Yan, W.; Sun, X.; Lin, F. Association between dietary fiber intake and chronic kidney disease in adults with and without hypertension in the United States: A cross-sectional study of NHANES 2009–2020. Ren. Fail. 2024, 46, 2415514. [Google Scholar] [PubMed]
- Varleta, P.; Tagle, R. A feared combination: Hypertension and chronic kidney disease. J. Clin. Hypertens. 2019, 21, 102–104. [Google Scholar]
- Costantino, V.V.; Gil, L.A.; Bocanegra, V.; Valles, P.G. Molecular Mechanisms of Hypertensive Nephropathy: Renoprotective Effect of Losartan Through Hsp70. Cells 2021, 10, 3146. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Wang, X.; Zhu, J.; Huang, X.; Liu, M.; Qiao, Z.; Zhang, Y.; Sun, Y.; Wang, Z.; Zhan, P.; et al. GPR97 deficiency ameliorates renal interstitial fibrosis in mouse hypertensive nephropathy. Acta Pharmacol. Sin. 2023, 44, 1206–1216. [Google Scholar]
- Kao, T.W.; Huang, C.C. Blood pressure management and renal protection: Revisiting hypertensive nephropathy. J. Chin. Med. Assoc. 2021, 84, 911–916. [Google Scholar]
- Carriazo, S.; Vanessa Perez-Gomez, M.; Ortiz, A. Hypertensive nephropathy: A major roadblock hindering the advance of precision nephrology. Clin. Kidney J. 2020, 13, 504–509. [Google Scholar] [CrossRef]
- Bae, E.; Yu, M.Y.; Moon, J.J.; Kim, J.E.; Lee, S.; Han, S.W.; Park, D.J.; Kim, Y.S.; Yang, S.H. Renoprotective Effect of KLF2 on Glomerular Endothelial Dysfunction in Hypertensive Nephropathy. Cells 2022, 11, 762. [Google Scholar] [CrossRef]
- Nwia, S.M.; Leite, A.P.O.; Li, X.C.; Zhuo, J.L. Sex differences in the renin-angiotensin-aldosterone system and its roles in hypertension, cardiovascular, and kidney diseases. Front. Cardiovasc. Med. 2023, 10, 1198090. [Google Scholar]
- Jingjing Dong, S.W.X.Y. Angiotensin I converting enzyme (ACE) inhibitory activity and antihypertensive effects of rice peptides. Food Sci. Hum. Wellness 2022, 6, 1539–1543. [Google Scholar]
- Long, L.; Tan, L.; Xu, F.; Yang, W.; Li, H.; Liu, J.; Wang, K.; Zhao, Z.; Wang, Y.; Wang, C.; et al. Qingda Granule Attenuates Hypertension-Induced Cardiac Damage via Regulating Renin-Angiotensin System Pathway. Chin. J. Integr. Med. 2024. [Google Scholar] [CrossRef]
- Cai, W.; Zhang, Z.; Huang, Y.; Sun, H.; Qiu, L. Vaccarin alleviates hypertension and nephropathy in renovascular hypertensive rats. Exp. Ther. Med. 2018, 15, 924–932. [Google Scholar] [CrossRef] [PubMed]
- Duan, T.; Li, M.; Lin, Z.; Meng, L.; Li, M.; Xia, T.; Zhang, X.; Lin, G.; Yan, L.; Liang, M.; et al. The Protective Effect of Vitexin on Hypertensive Nephropathy Rats. Kidney Blood Press. Res. 2024, 49, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Son, M.; Oh, S.; Choi, J.; Jang, J.T.; Son, K.H.; Byun, K. Attenuating Effects of Dieckol on Hypertensive Nephropathy in Spontaneously Hypertensive Rats. Int. J. Mol. Sci. 2021, 22, 4230. [Google Scholar] [CrossRef]
- Lee, J.H.; Massagué, J. TGF-β in developmental and fibrogenic EMTs. Semin. Cancer Biol. 2022, 86, 136–145. [Google Scholar] [CrossRef]
- Shin, S.; Park, J.; Choi, H.; Lee, K. Hypotensive and Endothelium-Dependent Vasorelaxant Effects of Grayblue Spicebush Ethanol Extract in Rats. Foods 2023, 12, 4282. [Google Scholar] [CrossRef]
- Kretchy, I.A.; Owusu-Daaku, F.T.; Danquah, S.A.; Asampong, E. A psychosocial perspective of medication side effects, experiences, coping approaches and implications for adherence in hypertension management. Clin. Hypertens. 2015, 21, 19. [Google Scholar] [CrossRef]
- Jeong, E.; Park, S.; Yang, Y.; Baek, Y.; Yun, D.; Kim, H.; Go, G.; Lee, H. Black Soybean and Adzuki Bean Extracts Lower Blood Pressure by Modulating the Renin-Angiotensin System in Spontaneously Hypertensive Rats. Foods 2021, 10, 1571. [Google Scholar] [CrossRef]
- Chupeerach, C.; Aursalung, A.; Watcharachaisoponsiri, T.; Whanmek, K.; Thiyajai, P.; Yosphan, K.; Sritalahareuthai, V.; Sahasakul, Y.; Santivarangkna, C.; Suttisansanee, U. The Effect of Steaming and Fermentation on Nutritive Values, Antioxidant Activities, and Inhibitory Properties of Tea Leaves. Foods 2021, 10, 117. [Google Scholar] [CrossRef]
- Xue, W.; Zhao, W.; Wu, S.; Yu, Z. Underlying anti-hypertensive mechanism of the Mizuhopecten yessoensis derived peptide NCW in spontaneously hypertensive rats via widely targeted kidney metabolomics. Food Sci. Hum. Wellness 2024, 13, 472–481. [Google Scholar] [CrossRef]
- Tang, Z.; Zhan, L.; He, R.; Zhou, Y.; Tang, Q.; Liu, Z.; Zhang, S.; Liu, A. Hepatoprotective Effect of Tea Composite Solid Beverage on Alcohol-Caused Rat Liver Injury. Foods 2023, 12, 4126. [Google Scholar] [CrossRef]
- Xu, Y.; Liu, S.; Zhu, L.; Dai, L.; Qian, W.; Zhang, J.; Li, X.; Pan, W. Green tea protects against hippocampal neuronal apoptosis in diabetic encephalopathy by inhibiting JNK/MLCK signaling. Mol. Med. Rep. 2021, 24, 575. [Google Scholar] [PubMed]
- Cao, S.; Zhao, C.; Gan, R.; Xu, X.; Wei, X.; Corke, H.; Atanasov, A.G.; Li, H. Effects and Mechanisms of Tea and Its Bioactive Compounds for the Prevention and Treatment of Cardiovascular Diseases: An Updated Review. Antioxidants 2019, 8, 166. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, C.; Fang, W.; Tang, Q.; Zhan, L.; Shi, Y.; Tang, M.; Liu, Z.; Zhang, S.; Liu, A. Research progress on the lipid-lowering and weight loss effects of tea and the mechanism of its functional components. J. Nutr. Biochem. 2023, 112, 109210. [Google Scholar]
- An, R.; Sun, L.; Xiang, L.; Zhang, W.; Li, Q.; Lai, X.; Wen, S.; Huo, M.; Li, D.; Sun, S. Effect of yellowing time on bioactive compounds in yellow tea and their antiproliferative capacity in HepG2 cells. Food Sci. Nutr. 2019, 7, 1838–1847. [Google Scholar]
- Wu, M.; Wu, X.; Zhu, J.; Li, F.; Wei, X.; Wang, Y. Selenium-enriched and ordinary green tea extracts prevent high blood pressure and alter gut microbiota composition of hypertensive rats caused by high-salt diet. Food Sci. Hum. Wellness 2022, 11, 738–751. [Google Scholar] [CrossRef]
- Xingfei, L.; Shunshun, P.; Wenji, Z.; Lingli, S.; Qiuhua, L.; Ruohong, C.; Shili, S. Properties of ACE inhibitory peptide prepared from protein in green tea residue and evaluation of its anti-hypertensive activity. Process Biochem. 2020, 92, 277–287. [Google Scholar] [CrossRef]
- Chen, X.; Sun, L.; Li, D.; Lai, X.; Wen, S.; Chen, R.; Zhang, Z.; Li, Q.; Sun, S. Green tea peptides ameliorate diabetic nephropathy by inhibiting the TGF-β/Smad signaling pathway in mice. Food Funct. 2022, 13, 3258–3270. [Google Scholar] [CrossRef]
- Yao, M.; Lian, D.; Wu, M.; Zhou, Y.; Fang, Y.; Zhang, S.; Zhang, W.; Yang, Y.; Li, R.; Chen, H.; et al. Isoliensinine Attenuates Renal Fibrosis and Inhibits TGF-β1/Smad2/3 Signaling Pathway in Spontaneously Hypertensive Rats. Drug Des. Dev. Ther. 2023, 17, 2749–2762. [Google Scholar] [CrossRef]
- Isaka, Y. Targeting TGF-β Signaling in Kidney Fibrosis. Int. J. Mol. Sci. 2018, 19, 2532. [Google Scholar] [CrossRef]
- Hernandez-Ledesma, B.; Del Mar Contreras, M.; Recio, I. Antihypertensive peptides: Production, bioavailability and incorporation into foods. Adv. Colloid Interface Sci. 2011, 165, 23–35. [Google Scholar]
- Ngamsuk, S.; Huang, T.; Hsu, J. ACE Inhibitory Activity and Molecular Docking of Gac Seed Protein Hydrolysate Purified by HILIC and RP-HPLC. Molecules 2020, 25, 4635. [Google Scholar] [CrossRef] [PubMed]
- World Medical Association. World Medical Association Declaration of Helsinki Ethical Principles for Medical Research Involving Human Subjects. JAMA 2013, 310, 2191–2194. [Google Scholar]
- Huang, M.; Zhu, S.; Huang, H.; He, J.; Tsuji, K.; Jin, W.W.; Xie, D.; Ham, O.; Capen, D.E.; Lu, W.; et al. Integrin-Linked Kinase Deficiency in Collecting Duct Principal Cell Promotes Necroptosis of Principal Cell and Contributes to Kidney Inflammation and Fibrosis. J. Am. Soc. Nephrol. 2019, 30, 2073–2090. [Google Scholar] [PubMed]
- Zheng, W.; Huang, T.; Tang, Q.Z.; Li, S.; Qin, J.; Chen, F. Astragalus Polysaccharide Reduces Blood Pressure, Renal Damage, and Dysfunction Through the TGF-beta1-ILK Pathway. Front. Pharmacol. 2021, 12, 706617. [Google Scholar]
- Shi, H.H.; Zhang, L.Y.; Chen, L.P.; Yang, J.Y.; Wang, C.C.; Xue, C.H.; Wang, Y.M.; Zhang, T.T. EPA-Enriched Phospholipids Alleviate Renal Interstitial Fibrosis in Spontaneously Hypertensive Rats by Regulating TGF-beta Signaling Pathways. Mar. Drugs 2022, 20, 152. [Google Scholar]
- Leader, C.J.; Kelly, D.J.; Sammut, I.A.; Wilkins, G.T.; Walker, R.J. Spironolactone mitigates, but does not reverse, the progression of renal fibrosis in a transgenic hypertensive rat. Physiol. Rep. 2020, 8, e14448. [Google Scholar]
- Zhao, L.; Zhang, Z.; Li, P.; Gao, Y.; Shi, Y. Bakuchiol regulates TLR4/MyD88/NF-kappaB and Keap1/Nrf2/HO-1 pathways to protect against LPS-induced acute lung injury in vitro and in vivo. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2024, 397, 3301–3312. [Google Scholar]
- Lu, Y.; Xie, X.N.; Xin, Q.Q.; Yuan, R.; Miao, Y.; Cong, W.H.; Chen, K.J. Advance on Chinese Medicine for Hypertensive Renal Damage: Focus on the Complex Molecular Mechanisms. Chin. J. Integr. Med. 2024, 30, 938–948. [Google Scholar]
- Shen, Y.; Miao, N.; Xu, J.; Gan, X.; Xu, D.; Zhou, L.; Xue, H.; Zhang, W.; Lu, L. Metformin Prevents Renal Fibrosis in Mice with Unilateral Ureteral Obstruction and Inhibits Ang II-Induced ECM Production in Renal Fibroblasts. Int. J. Mol. Sci. 2016, 17, 146. [Google Scholar] [CrossRef]
- Siltari, A.; Vapaatalo, H.; Korpela, R. Milk and milk-derived peptides combat against hypertension and vascular dysfunction: A review. Int. J. Food Sci. Technol. 2019, 54, 1920–1929. [Google Scholar]
- Zambrowicz, A.; Pokora, M.; Setner, B.; Dąbrowska, A.; Szołtysik, M.; Babij, K.; Szewczuk, Z.; Trziszka, T.; Lubec, G.; Chrzanowska, J. Multifunctional peptides derived from an egg yolk protein hydrolysate: Isolation and characterization. Amino Acids 2015, 47, 369–380. [Google Scholar] [PubMed]
- Geng, X.; Tian, G.; Zhang, W.; Zhao, Y.; Zhao, L.; Wang, H.; Ng, T.B. A Tricholoma matsutake Peptide with Angiotensin Converting Enzyme Inhibitory and Antioxidative Activities and Antihypertensive Effects in Spontaneously Hypertensive Rats. Sci. Rep. 2016, 6, 24130. [Google Scholar]
- Zhang, Y.; Dai, Z.; Zhao, X.; Chen, C.; Li, S.; Meng, Y.; Suonan, Z.; Sun, Y.; Shen, Q.; Wang, L.; et al. Deep learning drives efficient discovery of novel antihypertensive peptides from soybean protein isolate. Food Chem. 2023, 404, 134690. [Google Scholar]
- Owoicho Orgah, J.; Wang, M.; Yang, X.; Wang, Z.; Wang, D.; Zhang, Q.; Fan, G.; Han, J.; Qin, G.; Gao, X.; et al. Danhong Injection Protects Against Hypertension-Induced Renal Injury Via Down-Regulation of Myoglobin Expression in Spontaneously Hypertensive Rats. Kidney Blood Press. Res. 2018, 43, 12–24. [Google Scholar]
- Safari, S.; Yousefifard, M.; Hashemi, B.; Baratloo, A.; Forouzanfar, M.M.; Rahmati, F.; Motamedi, M.; Najafi, I. The value of serum creatine kinase in predicting the risk of rhabdomyolysis-induced acute kidney injury: A systematic review and meta-analysis. Clin. Exp. Nephrol. 2016, 20, 153–161. [Google Scholar]
- Wong, C.K.S.; Falkenham, A.; Myers, T.; Légaré, J. Connective tissue growth factor expression after angiotensin II exposure is dependent on transforming growth factor-β signaling via the canonical Smad-dependent pathway in hypertensive induced myocardial fibrosis. J. Renin-Angiotensin-Aldosterone Syst. 2018, 19, 1605484098. [Google Scholar]
- Kovarik, J.J.; Kaltenecker, C.C.; Domenig, O.; Antlanger, M.; Poglitsch, M.; Kopecky, C.; Säemann, M.D. Effect of Mineralocorticoid Receptor Antagonism and ACE Inhibition on Angiotensin Profiles in Diabetic Kidney Disease: An Exploratory Study. Diabetes Ther. 2021, 12, 2485–2498. [Google Scholar]
- Biernacka, A.; Dobaczewski, M.; Frangogiannis, N.G. The Association Between Salt Taste Threshold and Hypertensive Nephropathy in Patients with Essential Hypertension. Am. J. Hypertens. 2011, 35, 965. [Google Scholar]
- Wang, H.; Jiang, Q.; Zhang, L. Baicalin protects against renal interstitial fibrosis in mice by inhibiting the TGF-beta/Smad signalling pathway. Pharm. Biol. 2022, 60, 1407–1416. [Google Scholar]
- Juan, Y.; Yu, Y.; Tsai, Y.; Lee, C.; Chen, Y.; Ting, Y.; Tsai, J.; Hsieh, Y. Alpha-mangostin alleviate renal interstitial fibrosis via suppression of TGF-beta1/Smad/ERK signaling axis in vitro and in vivo. Biochem. Pharmacol. 2023, 218, 115935. [Google Scholar]
- Inazaki, K.; Kanamaru, Y.; Kojima, Y.; Sueyoshi, N.; Okumura, K.; Kaneko, K.; Yamashiro, Y.; Ogawa, H.; Nakao, A. Smad3 deficiency attenuates renal fibrosis, inflammation, and apoptosis after unilateral ureteral obstruction. Kidney Int. 2004, 66, 597–604. [Google Scholar] [PubMed]
- Yang, F.; Zhang, K.; Dai, X.; Jiang, W.; Eulogio, J.L.M.N.; Llorent Mart Nez, E.J. Preliminary Exploration of Potential Active Ingredients and Molecular Mechanisms of Yanggan Yishui Granules for Treating Hypertensive Nephropathy Using UPLC-Q-TOF/MS Coupled with Network Pharmacology and Molecular Docking Strategy. J. Anal. Methods Chem. 2024, 2024, 7967918–7967999. [Google Scholar]
- Chen, Y.; Wang, K.; Yang, J.; Zhang, A.; Dong, X.; Zhou, Z.; Li, T.; Fan, R. Mechanism of ferroptosis in hypertensive nephropathy. Transl. Androl. Urol. 2022, 11, 617–626. [Google Scholar]
- Huang, Y.; Peng, K.; Yan, J.; Chen, H.; Jiang, P.; Du, Y.; Ling, X.; Zhang, S.; Wu, J. The Participation of Ferroptosis in Fibrosis of the Heart and Kidney Tissues in Dahl Salt-Sensitive Hypertensive Rats. Am. J. Hypertens. 2024, 37, 784–791. [Google Scholar]
- Xiong, D.; Hu, W.; Ye, S.; Yuan-Sheng, T. Isoliquiritigenin alleviated the Ang II-induced hypertensive renal injury through suppressing inflammation cytokines and oxidative stress-induced apoptosis via Nrf2 and NF-kB pathways. Biochem. Biophys. Res. Commun. 2018, 506, 161–168. [Google Scholar]
- Hashmi, S.F.; Rathore, H.A.; Sattar, M.A.; Johns, E.J.; Gan, C.-Y.; Chia, T.Y.; Ahmad, A. Hydrogen Sulphide Treatment Prevents Renal Ischemia-Reperfusion Injury by Inhibiting the Expression of ICAM-1 and NF-kB Concentration in Normotensive and Hypertensive Rats. Biomolecules 2021, 11, 1549. [Google Scholar] [CrossRef]
- Hu, N.; Wang, C.; Dai, X.; Zhou, M.; Gong, L.; Yu, L.; Peng, C.; Li, Y. Phillygenin inhibits LPS-induced activation and inflammation of LX2 cells by TLR4/MyD88/NF-κB signaling pathway. J. Ethnopharmacol. 2020, 248, 112361. [Google Scholar]
- Li, H.B.; Li, X.; Huo, C.J.; Su, Q.; Guo, J.; Yuan, Z.Y.; Zhu, G.Q.; Shi, X.L.; Liu, J.J.; Kang, Y.M. TLR4/MyD88/NF-κB signaling and PPAR-γ within the paraventricular nucleus are involved in the effects of telmisartan in hypertension. Toxicol. Appl. Pharmacol. 2016, 305, 93–102. [Google Scholar]
- Kang, Y.; Ma, Y.; Zheng, J.; Elks, C.; Sriramula, S.; Yang, Z.; Francis, J. Brain nuclear factor-kappa B activation contributes to neurohumoral excitation in angiotensin II-induced hypertension. Cardiovasc. Res. 2009, 82, 503–512. [Google Scholar]
- Shi, M.; Guo, F.; Liao, D.; Huang, R.; Feng, Y.; Zeng, X.; Ma, L.; Fu, P. Pharmacological inhibition of fatty acid-binding protein 4 alleviated kidney inflammation and fibrosis in hyperuricemic nephropathy. Eur. J. Pharmacol. 2020, 887, 173570. [Google Scholar]
- Bhandari, D.; Rafiq, S.; Gat, Y.; Gat, P.; Waghmare, R.; Kumar, V. A Review on Bioactive Peptides: Physiological Functions, Bioavailability and Safety. Int. J. Pept. Res. Ther. 2020, 26, 139–150. [Google Scholar] [CrossRef]
- Liu, P.; Lan, X.; Yaseen, M.; Wu, S.; Feng, X.; Zhou, L.; Sun, J.; Liao, A.; Liao, D.; Sun, L. Purification, Characterization and Evaluation of Inhibitory Mechanism of ACE Inhibitory Peptides from Pearl Oyster (Pinctada fucata martensii) Meat Protein Hydrolysate. Mar. Drugs 2019, 17, 463. [Google Scholar] [CrossRef] [PubMed]
- Daskaya-Dikmen, C.; Yucetepe, A.; Karbancioglu-Guler, F.; Daskaya, H.; Ozcelik, B. Angiotensin-I-Converting Enzyme (ACE)-Inhibitory Peptides from Plants. Nutrients 2017, 9, 316. [Google Scholar] [CrossRef] [PubMed]
- Lammi, C.; Aiello, G.; Boschin, G.; Arnoldi, A. Multifunctional peptides for the prevention of cardiovascular disease: A new concept in the area of bioactive food-derived peptides. J. Funct. Foods 2019, 55, 135–145. [Google Scholar] [CrossRef]
- Aiello, G.; Cropotova, J.; Kvangarsnes, K.; d’Adduzio, L.; Fanzaga, M.; Bollati, C.; Boschin, G.; Roda, G.; Lammi, C. Ultrasonicated Atlantic herring side streams as source of multifunctional bioactive and bioavailable peptides. npj Sci. Food. 2025, 9, 25. [Google Scholar] [CrossRef]
- Xiang, L.; Qiu, Z.; Zhao, R.; Zheng, Z.; Qiao, X. Advancement and prospects of production, transport, functional activity and structure-activity relationship of food-derived angiotensin converting enzyme (ACE) inhibitory peptides. Crit. Rev. Food Sci. Nutr. 2023, 63, 1437–1463. [Google Scholar] [CrossRef]
- Li, G.; Le, G.; Shi, Y.; Shrestha, S. Angiotensin I–converting enzyme inhibitory peptides derived from food proteins and their physiological and pharmacological effects. Nutr. Res. 2004, 24, 469–486. [Google Scholar] [CrossRef]
- Cao, X.; Zhou, H.; Xie, J.; Huang, L.; Guo, S.; Zhang, Z.; Chen, Q.; Meng, C.; Zhang, F.; Hong, J. Preparation and stability of angiotensin-I-converting enzyme (ACE) inhibitory peptides from protein hydrolysate of Wuyi rock tea residues. CYTA J. Food 2024, 22, 2401032. [Google Scholar] [CrossRef]
- Sullivan, J.C.; Bhatia, K.; Yamamoto, T.; Elmarakby, A.A. Angiotensin (1–7) Receptor Antagonism Equalizes Angiotensin II–Induced Hypertension in Male and Female Spontaneously Hypertensive Rats. Hypertension 2010, 56, 658–666. [Google Scholar] [CrossRef]
- Shiraly, R.; Khani Jeihooni, A.; Bakhshizadeh Shirazi, R. Perception of risk of hypertension related complications and adherence to antihypertensive drugs: A primary healthcare based cross-sectional study. BMC Prim. Care 2022, 23, 303. [Google Scholar] [CrossRef]
- Unger, T.; Borghi, C.; Charchar, F.; Khan, N.A.; Poulter, N.R.; Prabhakaran, D.; Ramirez, A.; Schlaich, M.; Stergiou, G.S.; Tomaszewski, M.; et al. 2020 International Society of Hypertension Global Hypertension Practice Guidelines. Hypertension 2020, 75, 1334–1357. [Google Scholar] [PubMed]
- Lee, D.; Kim, M.; Lee, H.; Han, J.; Myung, C. The combined effects of telmisartan and ramipril on hypertension and cardiovascular injury. J. Pharm. Investig. 2022, 52, 443–451. [Google Scholar]
- Borghi, C.; Soldati, M.; Bragagni, A.; Cicero, A.F.G. Safety implications of combining ACE inhibitors with thiazides for the treatment of hypertensive patients. Expert Opin. Drug Saf. 2020, 19, 1577–1583. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Names | Sequences |
|---|---|
| GAPDH | Forward: 5′-AGACAGCCGCATCTTCTTGT-3′ |
| Reverse: 5′-CTTGCCGTGGGTAGAGTCAT-3′ | |
| TGF-β1 | Forward: 5′-ACCTGCAAGACCATCGACATG-3′ |
| Reverse: 5′-CGAGCCTTAGTTTGGACAGGAT-3′ | |
| Smad2 | Forward: 5′-TTTGCCGAGTGCCTAAGTGATA-3′ |
| Reverse: 5′-TTCTTATGGTGCACATTCGAGTC-3′ | |
| Smad3 | Forward: 5′-GCTGTCTACCAGTTGACTCGCAT-3′ |
| Reverse: 5′-GGGTGCTGGTCACTGTCTGTCT-3′ | |
| ACE | Forward: 5′-TCCTATTCCCGCTCATCT-3′ |
| Reverse: 5′-CCAGCCCTTCTGTACCATT-3′ | |
| Ang II | Forward: 5′-CACCCCTTTCATCTCCTCTACTA-3′ |
| Reverse: 5′-TCTTGCCTCACTCAGCATCTT-3′ | |
| α-SMA | Forward: 5′-AACACGGCATCATCACCAAC-3′ |
| Reverse: 5′-CACAGCCTGAATAGCCACATAC-3′ | |
| NF-κB p65 | Forward: 5′-AAGATCAATGGCTACACAGG-3′ |
| Reverse: 5′-CCTCAATGTCTTCTTTCTGC-3′ | |
| iNOS | Forward: 5′-GACCAAACTGTGTGCCTGGA-3′ |
| Reverse: 5′-TACTCTGAGGGCTGACACAAGG-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Sun, S.; Lai, X.; Li, Q.; Chen, R.; Zhang, Z.; Hao, M.; Zhang, S.; Sun, L.; Li, D. Mechanism of Green Tea Peptides in Lowering Blood Pressure and Alleviating Renal Injury Induced by Hypertension Through the Ang II/TGF-β1/SMAD Signaling Pathway. Nutrients 2025, 17, 1300. https://doi.org/10.3390/nu17081300
Li L, Sun S, Lai X, Li Q, Chen R, Zhang Z, Hao M, Zhang S, Sun L, Li D. Mechanism of Green Tea Peptides in Lowering Blood Pressure and Alleviating Renal Injury Induced by Hypertension Through the Ang II/TGF-β1/SMAD Signaling Pathway. Nutrients. 2025; 17(8):1300. https://doi.org/10.3390/nu17081300
Chicago/Turabian StyleLi, Lulu, Shili Sun, Xingfei Lai, Qiuhua Li, Ruohong Chen, Zhenbiao Zhang, Mengjiao Hao, Suwan Zhang, Lingli Sun, and Dongli Li. 2025. "Mechanism of Green Tea Peptides in Lowering Blood Pressure and Alleviating Renal Injury Induced by Hypertension Through the Ang II/TGF-β1/SMAD Signaling Pathway" Nutrients 17, no. 8: 1300. https://doi.org/10.3390/nu17081300
APA StyleLi, L., Sun, S., Lai, X., Li, Q., Chen, R., Zhang, Z., Hao, M., Zhang, S., Sun, L., & Li, D. (2025). Mechanism of Green Tea Peptides in Lowering Blood Pressure and Alleviating Renal Injury Induced by Hypertension Through the Ang II/TGF-β1/SMAD Signaling Pathway. Nutrients, 17(8), 1300. https://doi.org/10.3390/nu17081300