
The Impact of Resveratrol and Melatonin on the Genome and Oxidative Status in Ageing Rats
 ,
,  , , , , , , , and
, , , , , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
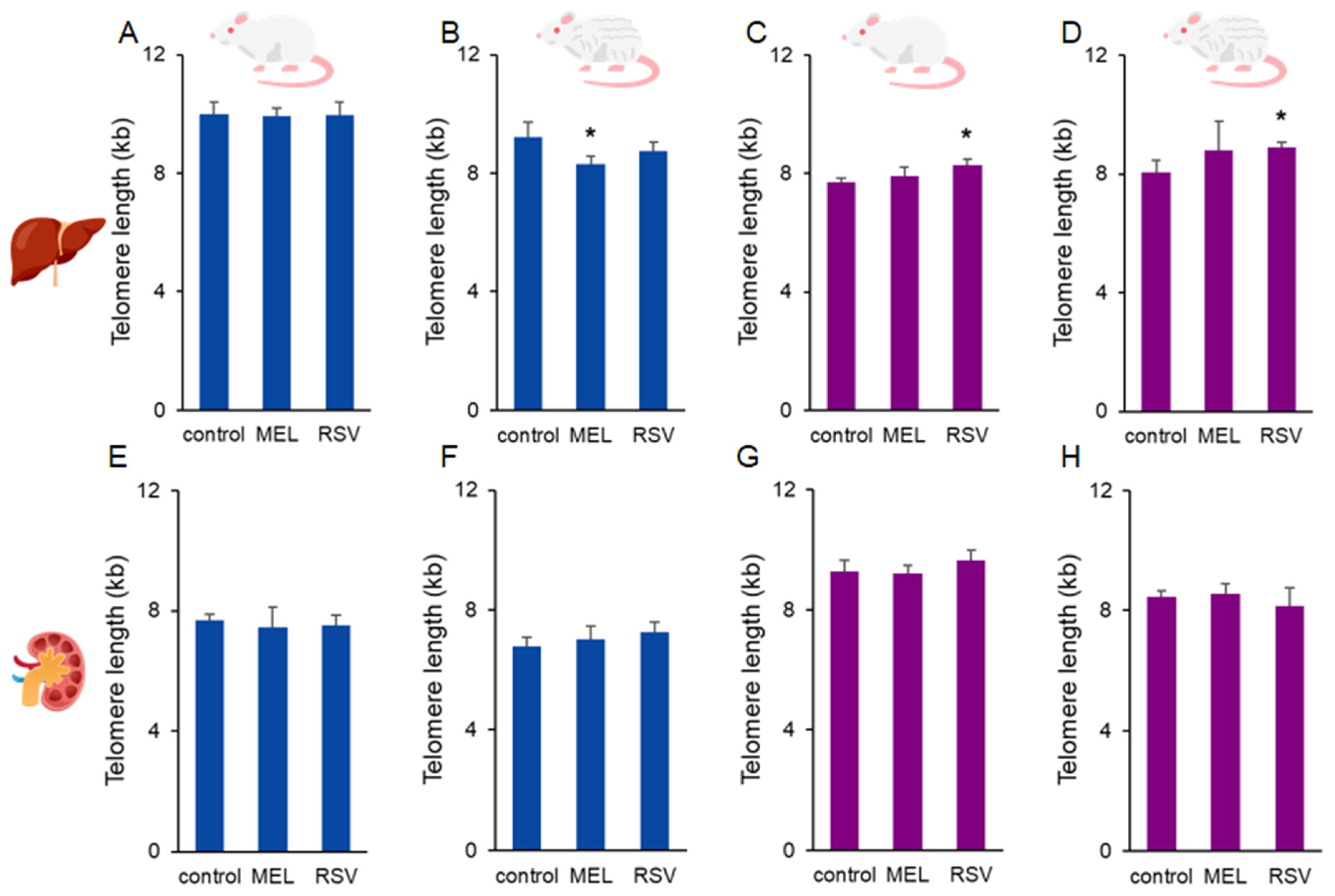
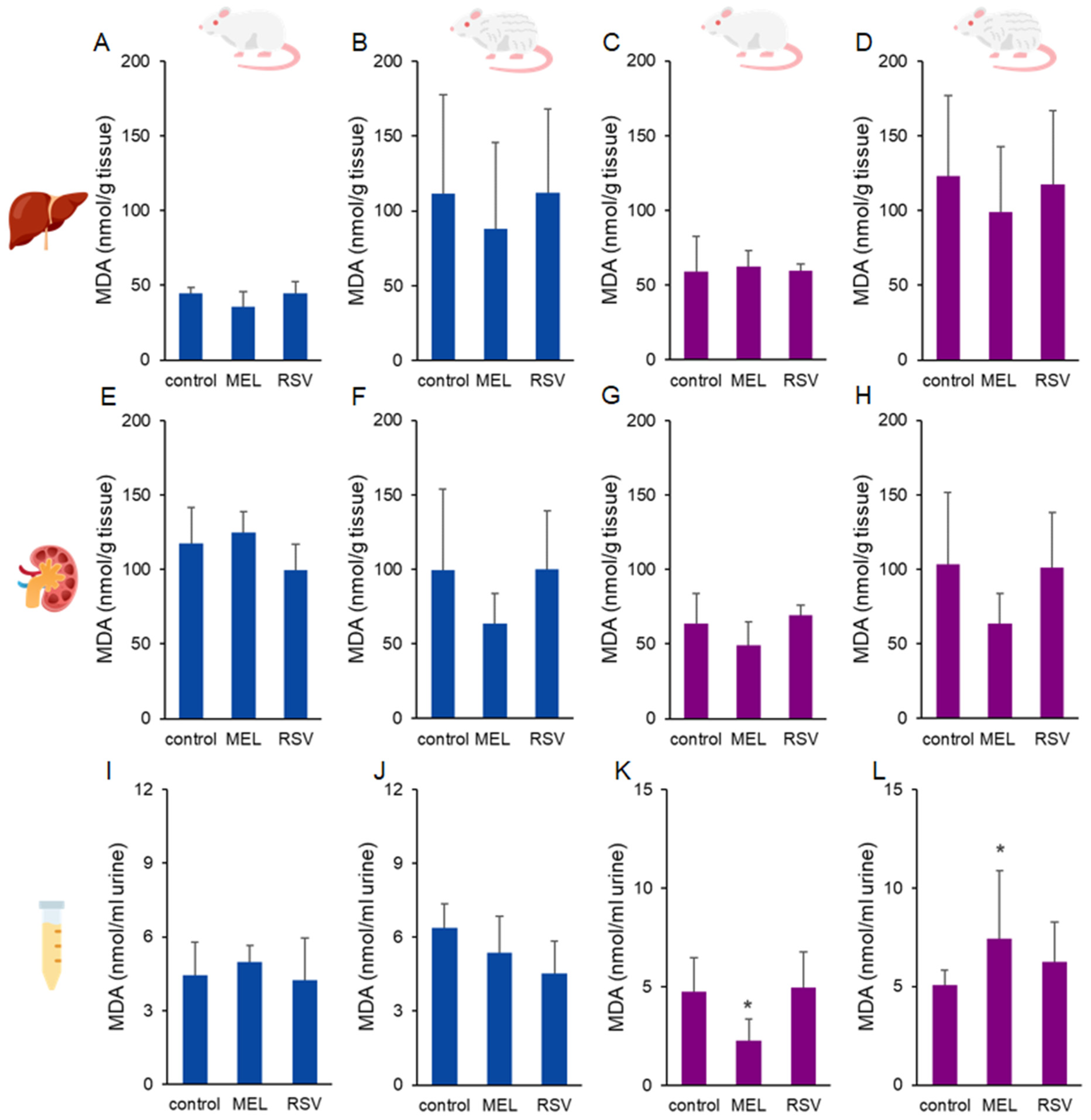
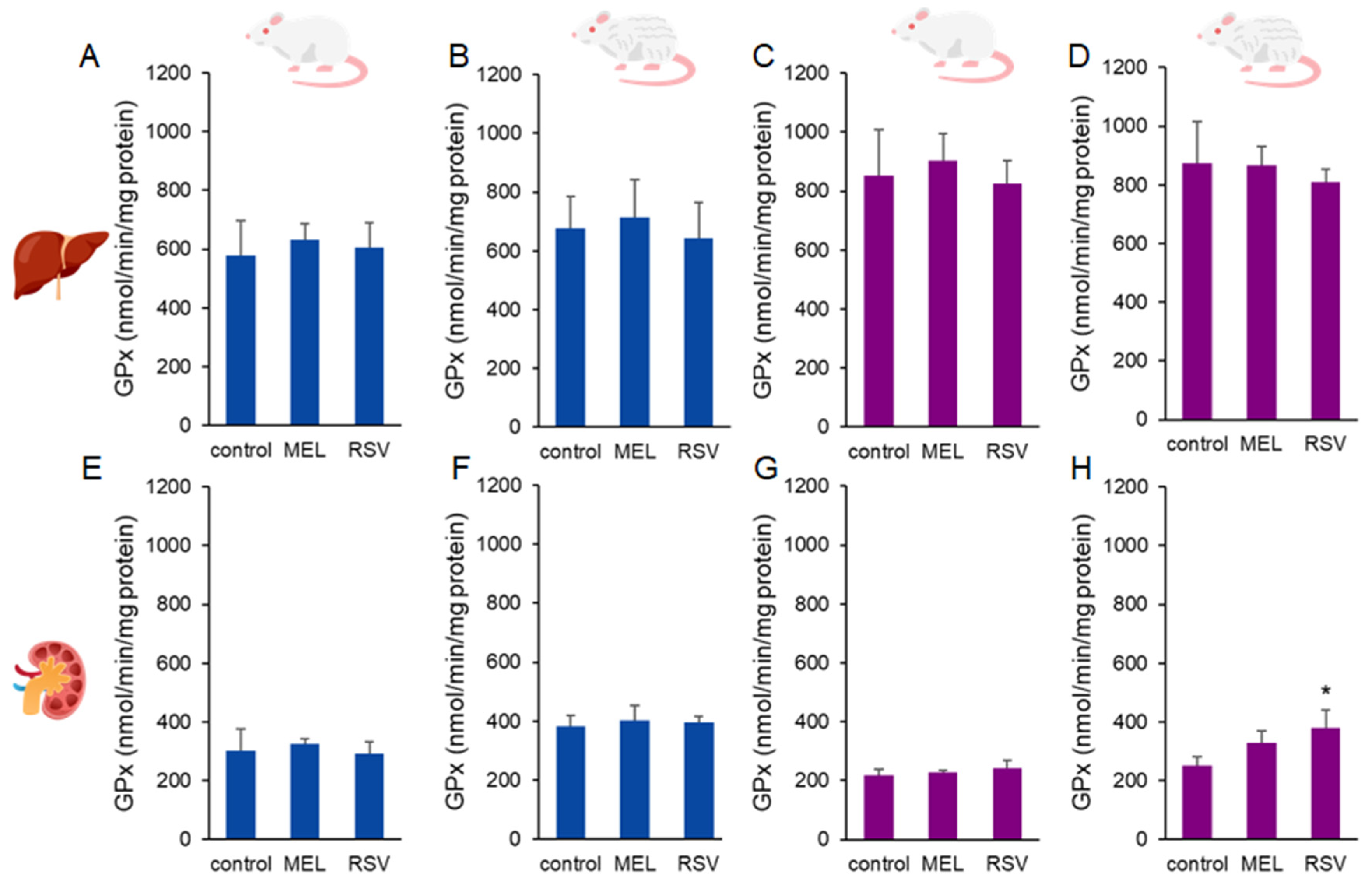
2. Results
3. Discussion
4. Conclusions
5. Materials and Methods
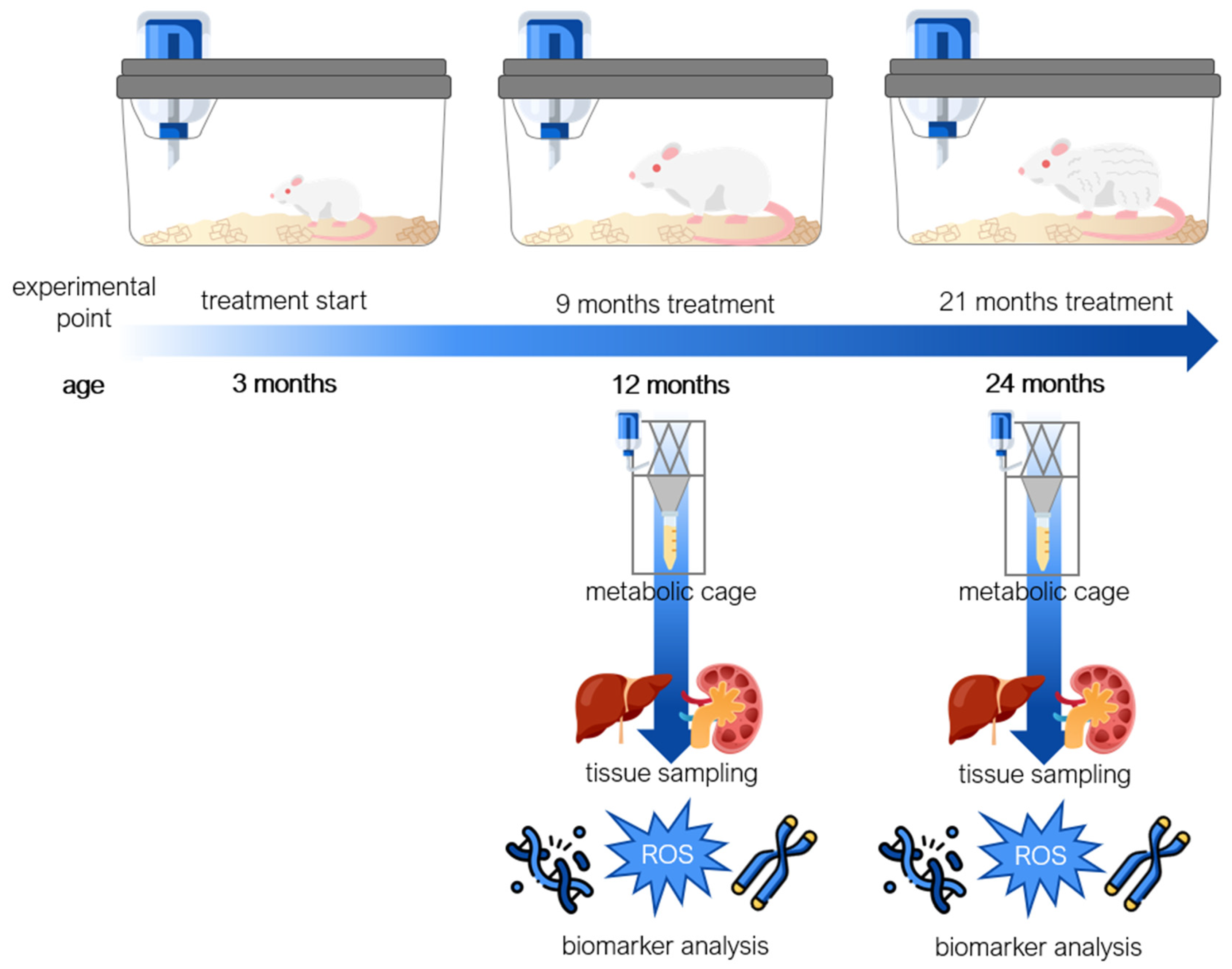
5.1. Animals
5.2. Animal Treatment and Sample Collection
5.3. Telomere Detection and Length Analysis
5.4. Alkaline Comet Assay Procedure
5.5. Urinary 8-OHdG
5.6. Glutathione Analysis
5.7. Malondialdehyde Analysis
5.8. Protein Carbonyls
5.9. SOD Activity Analysis
5.10. GPx Activity Analysis
5.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- UN—Depratment of Economic and Social Affairs. Leaving No One Behind in an Ageing World. In World Social Report 2023: Leaving No One Behind in an Ageing World; UN: New York, NY, USA, 2023; pp. 1–4. [Google Scholar]
- Michel, J.-P.; Leonardi, M.; Martinc, M.; Prina, M. WHO’s Report for the Decade of Healthy Ageing 2021–30 Sets the Stage for Globally Comparable Data on Healthy Ageing. Lancet 2021, 2, e121–e122. [Google Scholar] [CrossRef] [PubMed]
- Baker, D.J.; Wijshake, T.; Tchkonia, T.; LeBrasseur, N.K.; Childs, B.G.; van de Sluis, B.; Kirkland, J.L.; van Deursen, J.M. Clearance of P16Ink4a-Positive Senescent Cells Delays Ageing-Associated Disorders. Nature 2011, 479, 232–236. [Google Scholar] [CrossRef] [PubMed]
- von Zglinicki, T.; Saretzki, G.; Döcke, W.; Lotze, C. Mild Hyperoxia Shortens Telomeres and Inhibits Proliferation of Fibroblasts: A Model for Senescence? Exp. Cell Res. 1995, 220, 186–193. [Google Scholar] [CrossRef] [PubMed]
- von Zglinicki, T.; Saretzki, G.; Ladhoff, J.; d’Adda di Fagagna, F.; Jackson, S.P. Human Cell Senescence as a DNA Damage Response. Mech. Ageing Dev. 2005, 126, 111–117. [Google Scholar] [CrossRef]
- Serrano, M.; Lin, A.W.; McCurrach, M.E.; Beach, D.; Lowe, S.W. Oncogenic Ras Provokes Premature Cell Senescence Associated with Accumulation of P53 and P16INK4a. Cell 1997, 88, 593–602. [Google Scholar] [CrossRef]
- Vidacek, N.S.; Cukusić, A.; Ivanković, M.; Fulgosi, H.; Huzak, M.; Smith, J.R.; Rubelj, I. Abrupt Telomere Shortening in Normal Human Fibroblasts. Exp. Gerontol. 2010, 45, 235–242. [Google Scholar] [CrossRef]
- Rubelj, I.; Huzak, M.; Brdar, B. Sudden Senescence Syndrome Plays a Major Role in Cell Culture Proliferation. Mech. Ageing Dev. 2000, 112, 233–241. [Google Scholar] [CrossRef]
- Kim Sh, S.; Kaminker, P.; Campisi, J. Telomeres, Aging and Cancer: In Search of a Happy Ending. Oncogene 2002, 21, 503–511. [Google Scholar] [CrossRef]
- Cragg, G.M.; Pezzuto, J.M. Natural Products as a Vital Source for the Discovery of Cancer Chemotherapeutic and Chemopreventive Agents. Med. Princ. Pract. 2016, 25, 41–59. [Google Scholar] [CrossRef]
- Dave, A.; Parande, F.; Park, E.-J.; Pezzuto, J.M. Phytochemicals and Cancer Chemoprevention. J. Cancer Metastasis Treat. 2020, 6, 46. [Google Scholar] [CrossRef]
- Pezzuto, J.M. Resveratrol: Twenty Years of Growth, Development and Controversy. Biomol. Ther. 2019, 27, 1–14. [Google Scholar] [CrossRef]
- Howitz, K.T.; Bitterman, K.J.; Cohen, H.Y.; Lamming, D.W.; Lavu, S.; Wood, J.G.; Zipkin, R.E.; Chung, P.; Kisielewski, A.; Zhang, L.-L.; et al. Small Molecule Activators of Sirtuins Extend Saccharomyces Cerevisiae Lifespan. Nature 2003, 425, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, M.; Kim, S.K.; Berdichevsky, A.; Guarente, L. A Role for SIR-2.1 Regulation of ER Stress Response Genes in Determining C. Elegans Life Span. Dev. Cell 2005, 9, 605–615. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.G.; Rogina, B.; Lavu, S.; Howitz, K.; Helfand, S.L.; Tatar, M.; Sinclair, D. Sirtuin Activators Mimic Caloric Restriction and Delay Ageing in Metazoans. Nature 2004, 430, 686–689. [Google Scholar] [CrossRef]
- Labbé, A.; Garand, C.; Cogger, V.C.; Paquet, E.R.; Desbiens, M.; Le Couteur, D.G.; Lebel, M. Resveratrol Improves Insulin Resistance Hyperglycemia and Hepatosteatosis but Not Hypertriglyceridemia, Inflammation, and Life Span in a Mouse Model for Werner Syndrome. J. Gerontol. A Biol. Sci. Med. Sci. 2011, 66, 264–278. [Google Scholar] [CrossRef]
- da Luz, P.L.; Tanaka, L.; Brum, P.C.; Dourado, P.M.M.; Favarato, D.; Krieger, J.E.; Laurindo, F.R.M. Red Wine and Equivalent Oral Pharmacological Doses of Resveratrol Delay Vascular Aging but Do Not Extend Life Span in Rats. Atherosclerosis 2012, 224, 136–142. [Google Scholar] [CrossRef]
- Dubbels, R.; Reiter, R.J.; Klenke, E.; Goebel, A.; Schnakenberg, E.; Ehlers, C.; Schiwara, H.W.; Schloot, W. Melatonin in Edible Plants Identified by Radioimmunoassay and by High Performance Liquid Chromatography-Mass Spectrometry. J. Pineal Res. 1995, 18, 28–31. [Google Scholar] [CrossRef]
- Reiter, R.J.; Mayo, J.C.; Tan, D.-X.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an Antioxidant: Under Promises but over Delivers. J. Pineal Res. 2016, 61, 253–278. [Google Scholar] [CrossRef]
- Acuña-Castroviejo, D.; Escames, G.; Venegas, C.; Díaz-Casado, M.E.; Lima-Cabello, E.; López, L.C.; Rosales-Corral, S.; Tan, D.-X.; Reiter, R.J. Extrapineal Melatonin: Sources, Regulation, and Potential Functions. Cell. Mol. Life Sci. 2014, 71, 2997–3025. [Google Scholar] [CrossRef]
- Zhang, H.-M.; Zhang, Y. Melatonin: A Well-Documented Antioxidant with Conditional pro-Oxidant Actions. J. Pineal Res. 2014, 57, 131–146. [Google Scholar] [CrossRef]
- He, Y.; Li, Z.; Niu, Y.; Duan, Y.; Wang, Q.; Liu, X.; Dong, Z.; Zheng, Y.; Chen, Y.; Wang, Y.; et al. Progress in the Study of Aging Marker Criteria in Human Populations. Front. Public Health 2024, 12, 1305303. [Google Scholar] [CrossRef] [PubMed]
- Charles River Laboratories International, Inc. Wistar Rats NOMENCLATURE: Crl:WI; Strain Origin; Charles River: Boston, MA, USA, 2011. [Google Scholar]
- Breljak, D.; Micek, V.; Gerić, M.; Gajski, G.; Oguić, S.K.; Rašić, D.; Karaica, D.; Madunić, I.V.; Ljubojević, M.; Orct, T.; et al. Long-Term Effects of Melatonin and Resveratrol on Aging Rats: A Multi-Biomarker Approach. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2022, 876–877, 503443. [Google Scholar] [CrossRef]
- González-Navarro, H.; Dogan, S.; Tuna, B.G.; Potter, P.K.; Koks, G.; Koks, S. Aging in Rodents. In Encyclopedia of Biomedical Gerontology; Elsevier: Amsterdam, The Netherlands, 2020; pp. 73–87. [Google Scholar]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of Aging: An Expanding Universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef]
- Clayton, J.A. Studying Both Sexes: A Guiding Principle for Biomedicine. FASEB J. 2016, 30, 519–524. [Google Scholar] [CrossRef]
- Rowe, A. The Importance of Selection and Reporting of the Sex of Experimental Animals. Anim. Prod. Sci. 2019, 59, 401. [Google Scholar] [CrossRef]
- Ljubojević, M.; Orct, T.; Micek, V.; Karaica, D.; Jurasović, J.; Breljak, D.; Madunić, I.V.; Rašić, D.; Jovanović, I.N.; Peraica, M.; et al. Sex-Dependent Expression of Metallothioneins MT1 and MT2 and Concentrations of Trace Elements in Rat Liver and Kidney Tissues: Effect of Gonadectomy. J. Trace Elem. Med. Biol. 2019, 53, 98–108. [Google Scholar] [CrossRef]
- Sánchez-Hidalgo, M.; Guerrero Montávez, J.M.; Carrascosa-Salmoral, M.d.P.; Naranjo Gutierrez, M.d.C.; Lardone, P.J.; De La Lastra Romero, C.A. Decreased MT1 and MT2 Melatonin Receptor Expression in Extrapineal Tissues of the Rat during Physiological Aging. J. Pineal Res. 2009, 46, 29–35. [Google Scholar] [CrossRef]
- Sanchez-Hidalgo, M.; de la Lastra, C.A.; Carrascosa-Salmoral, M.P.; Naranjo, M.C.; Gomez-Corvera, A.; Caballero, B.; Guerrero, J.M. Age-Related Changes in Melatonin Synthesis in Rat Extrapineal Tissues. Exp. Gerontol. 2009, 44, 328–334. [Google Scholar] [CrossRef]
- Norwegian Scientific Committee for Food and Environment (VKM). Risk Assessment of Melatonin; Norwegian Scientific Committee for Food and Environment (VKM): Oslo, Norway, 2021. [Google Scholar]
- Martins Longaretti, L.; Luciano, J.A.; Strapazzon, G.; Pereira, M.; Damiani, A.P.; Rohr, P.; Rigo, F.K.; de Oliveira, C.A.; Steiner, B.T.; Vilela, T.C.; et al. Anti-Genotoxic and Anti-Mutagenic Effects of Melatonin Supplementation in a Mouse Model of Melanoma. Drug Chem. Toxicol. 2022, 45, 515–522. [Google Scholar] [CrossRef]
- Damiani, A.P.; Strapazzon, G.; De Oliveira Sardinha, T.T.; Rohr, P.; Gajski, G.; De Pinho, R.A.; De Andrade, V.M. Melatonin Supplementation over Different Time Periods until Ageing Modulates Genotoxic Parameters in Mice. Mutagenesis 2020, 35, 465–478. [Google Scholar] [CrossRef]
- Song, C.; Peng, W.; Yin, S.; Zhao, J.; Fu, B.; Zhang, J.; Mao, T.; Wu, H.; Zhang, Y. Melatonin Improves Age-Induced Fertility Decline and Attenuates Ovarian Mitochondrial Oxidative Stress in Mice. Sci. Rep. 2016, 6, 35165. [Google Scholar] [CrossRef]
- Akgun-Unal, N.; Ozyildirim, S.; Unal, O.; Gulbahce-Mutlu, E.; Mogulkoc, R.; Baltaci, A.K. The Effects of Resveratrol and Melatonin on Biochemical and Molecular Parameters in Diabetic Old Female Rat Hearts. Exp. Gerontol. 2023, 172, 112043. [Google Scholar] [CrossRef] [PubMed]
- Anisimov, V.N.; Zabezhinski, M.A.; Popovich, I.G.; Zaripova, E.A.; Musatov, S.A.; Andre, V.; Vigreux, C.; Godard, T.; Sichel, F. Inhibitory Effect of Melatonin on 7,12-Dimethylbenz[a]Anthracene-Induced Carcinogenesis of the Uterine Cervix and Vagina in Mice and Mutagenesis in Vitro. Cancer Lett. 2000, 156, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Baeeri, M.; Didari, T.; Khalid, M.; Mohammadi-Nejad, S.; Daghighi, S.M.; Farhadi, R.; Rahimifard, M.; Bayrami, Z.; Haghi-Aminjan, H.; Foroumadi, R.; et al. Molecular Evidence of the Inhibitory Potential of Melatonin against Naaso2-Induced Aging in Male Rats. Molecules 2021, 26, 6603. [Google Scholar] [CrossRef]
- Wessler, L.B.; Ise, K.; Lemos, I.C.; Rezende, V.L.; Duarte, M.B.; Damiani, A.P.; de Oliveira, J.; de Andrade, V.M.; Streck, E.L. Melatonin Ameliorates Oxidative Stress and DNA Damage of Rats Subjected to a Chemically Induced Chronic Model of Maple Syrup Urine Disease. Metab. Brain Dis. 2020, 35, 905–914. [Google Scholar] [CrossRef] [PubMed]
- Qasim, Z.; Ibrahim, Z.S.; Mahmoud, S.; Fahmy, M.; Shukry, M. Melatonin Mechanism to Mitigate Aging-Related Changes in the Liver, Kidney, and Brain in Rats. Egypt. J. Vet. Sci. 2025, 56, 993–1004. [Google Scholar] [CrossRef]
- Jiang, W.; Liu, J.; Zhao, X.; Yang, W. Melatonin-Induced Upregulation of Telomerase Activity Interferes with Macrophage Mitochondrial Metabolism and Suppresses NLRP3 Inflammasome Activation in the Treatment of Pneumonia. Heliyon 2024, 10, e29681. [Google Scholar] [CrossRef]
- Boutin, J.A.; Kennaway, D.J.; Jockers, R. Melatonin: Facts, Extrapolations and Clinical Trials. Biomolecules 2023, 13, 943. [Google Scholar] [CrossRef]
- Boutin, J.A.; Liberelle, M.; Yous, S.; Ferry, G.; Nepveu, F. Melatonin Facts: Lack of Evidence That Melatonin Is a Radical Scavenger in Living Systems. J. Pineal Res. 2024, 76, e12926. [Google Scholar] [CrossRef]
- Boutin, J.A.; Hamon de Almeida, V.; Coussay, N.; Legros, C.; Ferry, G.; Reybier, K. Melatonin Facts: Melatonin Lacks Immuno-Inflammation Boosting Capacities at the Molecular and Cellular Levels. Biochimie 2024, 222, 195–202. [Google Scholar] [CrossRef]
- Lieber, M.R.; Karanjawala, Z.E. Ageing, Repetitive Genomes and DNA Damage. Nat. Rev. Mol. Cell Biol. 2004, 5, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, E.; Paquet, C.; Beauchemin, M.; Bertrand, R. DNA-Damage Response Network at the Crossroads of Cell-Cycle Checkpoints, Cellular Senescence and Apoptosis. J. Zhejiang Univ. Sci. B 2007, 8, 377–397. [Google Scholar] [CrossRef] [PubMed]
- Baird, D.M.; Rowson, J.; Wynford-Thomas, D.; Kipling, D. Extensive Allelic Variation and Ultrashort Telomeres in Senescent Human Cells. Nat. Genet. 2003, 33, 203–207. [Google Scholar] [CrossRef]
- Londoño-Vallejo, J.A. Telomere Instability and Cancer. Biochimie 2008, 90, 73–82. [Google Scholar] [CrossRef]
- Salehi, B.; Mishra, A.P.; Nigam, M.; Sener, B.; Kilic, M.; Sharifi-Rad, M.; Fokou, P.V.T.; Martins, N.; Sharifi-Rad, J. Resveratrol: A Double-Edged Sword in Health Benefits. Biomedicines 2018, 6, 91. [Google Scholar] [CrossRef] [PubMed]
- Khafaga, A.F.; Noreldin, A.E.; Taha, A.E. The Adaptogenic Anti-Ageing Potential of Resveratrol against Heat Stress-Mediated Liver Injury in Aged Rats: Role of HSP70 and NF-KB Signalling. J. Therm. Biol. 2019, 83, 8–21. [Google Scholar] [CrossRef]
- Ran, Q.; Song, D.; Wang, Q.; Wang, D.; Chen, X.; Zhang, A.; Ma, L. Resveratrol Alleviates Arsenic Exposure-Induced Liver Fibrosis in Rats by Inhibiting Hepatocyte Senescence. Biol. Trace Elem. Res. 2024, 203, 1528–1538. [Google Scholar] [CrossRef]
- Majewski, M.; Ognik, K.; Thoene, M.; Rawicka, A.; Juśkiewicz, J. Resveratrol Modulates the Blood Plasma Levels of Cu and Zn, the Antioxidant Status and the Vascular Response of Thoracic Arteries in Copper Deficient Wistar Rats. Toxicol. Appl. Pharmacol. 2020, 390, 114877. [Google Scholar] [CrossRef]
- Hashemzaei, M.; Tabrizian, K.; Alizadeh, Z.; Pasandideh, S.; Rezaee, R.; Mamoulakis, C.; Tsatsakis, A.; Skaperda, Z.; Kouretas, D.; Shahraki, J. Resveratrol, Curcumin and Gallic Acid Attenuate Glyoxal-Induced Damage to Rat Renal Cells. Toxicol. Rep. 2020, 7, 1571–1577. [Google Scholar] [CrossRef]
- Rašić, D.; Želježić, D.; Kopjar, N.; Kifer, D.; Šegvić Klarić, M.; Peraica, M. DNA Damage in Rat Kidneys and Liver upon Subchronic Exposure to Single and Combined Ochratoxin A and Citrinin. World Mycotoxin J. 2019, 12, 163–172. [Google Scholar] [CrossRef]
- Virk, P.; Al-mukhaizeem, N.A.R.; Bin Morebah, S.H.; Fouad, D.; Elobeid, M. Protective Effect of Resveratrol against Toxicity Induced by the Mycotoxin, Zearalenone in a Rat Model. Food Chem. Toxicol. 2020, 146, 111840. [Google Scholar] [CrossRef] [PubMed]
- Boyaci, B.; Yalcin, R.; Cengel, A.; Erdem, O.; Dortlemez, O.; Dortlemez, H.; Sardas, S. Evaluation of DNA Damage in Lymphocytes of Cardiologists Exposed to Radiation during Cardiac Catheterization by the COMET ASSAY. Jpn. Heart J. 2004, 45, 845–853. [Google Scholar] [CrossRef]
- Brown, K.; Theofanous, D.; Britton, R.G.; Aburido, G.; Pepper, C.; Sri Undru, S.; Howells, L. Resveratrol for the Management of Human Health: How Far Have We Come? A Systematic Review of Resveratrol Clinical Trials to Highlight Gaps and Opportunities. Int. J. Mol. Sci. 2024, 25, 747. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Fang, G.; Song, W.; Cao, H.; Dong, R.; Huang, Y. Resveratrol’s Bibliometric and Visual Analysis from 2014 to 2023. Front. Plant Sci. 2024, 15, 1423323. [Google Scholar] [CrossRef]
- Rizvi, S.A.A.; Saleh, A.M. Applications of Nanoparticle Systems in Drug Delivery Technology. Saudi Pharm. J. 2018, 26, 64–70. [Google Scholar] [CrossRef]
- Cai, H.; Scott, E.; Kholghi, A.; Andreadi, C.; Rufini, A.; Karmokar, A.; Britton, R.G.; Horner-Glister, E.; Greaves, P.; Jawad, D.; et al. Cancer Chemoprevention: Evidence of a Nonlinear Dose Response for the Protective Effects of Resveratrol in Humans and Mice. Sci. Transl. Med. 2015, 7, 298ra117. [Google Scholar] [CrossRef]
- Vidaček, N.Š.; Nanić, L.; Ravlić, S.; Sopta, M.; Gerić, M.; Gajski, G.; Garaj-Vrhovac, V.; Rubelj, I. Telomeres, Nutrition, and Longevity: Can We Really Navigate Our Aging? J. Gerontol.: Ser. A 2018, 73, 39–47. [Google Scholar] [CrossRef]
- Breljak, D.; Brzica, H.; Sweet, D.H.; Anzai, N.; Sabolic, I. Sex-Dependent Expression of Oat3 (Slc22a8) and Oat1 (Slc22a6) Proteins in Murine Kidneys. Am. J. Physiol. Physiol. 2013, 304, F1114–F1126. [Google Scholar] [CrossRef]
- Brzica, H.; Breljak, D.; Krick, W.; Lovrić, M.; Burckhardt, G.; Burckhardt, B.C.; Sabolić, I. The Liver and Kidney Expression of Sulfate Anion Transporter Sat-1 in Rats Exhibits Male-Dominant Gender Differences. Pflügers Arch.—Eur. J. Physiol. 2009, 457, 1381–1392. [Google Scholar] [CrossRef]
- Sabolić, I.; Vrhovac, I.; Eror, D.B.; Gerasimova, M.; Rose, M.; Breljak, D.; Ljubojević, M.; Brzica, H.; Sebastiani, A.; Thal, S.C.; et al. Expression of Na+-ᴅ-Glucose Cotransporter SGLT2 in Rodents Is Kidney-Specific and Exhibits Sex and Species Differences. Am. J. Physiol. Physiol. 2012, 302, C1174–C1188. [Google Scholar] [CrossRef]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem 2023 Update. Nucleic Acids Res. 2023, 51, D1373–D1380. [Google Scholar] [CrossRef] [PubMed]
- Harley, C.B.; Futcher, A.B.; Greider, C.W. Telomeres Shorten during Ageing of Human Fibroblasts. Nature 1990, 345, 458–460. [Google Scholar] [CrossRef]
- Gajski, G.; Žegura, B.; Ladeira, C.; Pourrut, B.; Del Bo’, C.; Novak, M.; Sramkova, M.; Milić, M.; Gutzkow, K.B.; Costa, S.; et al. The Comet Assay in Animal Models: From Bugs to Whales—(Part 1 Invertebrates). Mutat. Res. Rev. Mutat. Res. 2019, 779, 82–113. [Google Scholar] [CrossRef] [PubMed]
- Gajski, G.; Žegura, B.; Ladeira, C.; Novak, M.; Sramkova, M.; Pourrut, B.; Del Bo’, C.; Milić, M.; Gutzkow, K.B.; Costa, S.; et al. The Comet Assay in Animal Models: From Bugs to Whales—(Part 2 Vertebrates). Mutat. Res. Mutat. Res. 2019, 781, 130–164. [Google Scholar] [CrossRef] [PubMed]
- Ladeira, C.; Møller, P.; Giovannelli, L.; Gajski, G.; Haveric, A.; Bankoglu, E.E.; Azqueta, A.; Gerić, M.; Stopper, H.; Cabêda, J.; et al. The Comet Assay as a Tool in Human Biomonitoring Studies of Environmental and Occupational Exposure to Chemicals-A Systematic Scoping Review. Toxics 2024, 12, 270. [Google Scholar] [CrossRef]
- Collins, A.; Møller, P.; Gajski, G.; Vodenková, S.; Abdulwahed, A.; Anderson, D.; Bankoglu, E.E.; Bonassi, S.; Boutet-Robinet, E.; Brunborg, G.; et al. Measuring DNA Modifications with the Comet Assay: A Compendium of Protocols. Nat. Protoc. 2023, 18, 929–989. [Google Scholar] [CrossRef]
- Møller, P.; Azqueta, A.; Boutet-Robinet, E.; Koppen, G.; Bonassi, S.; Milić, M.; Gajski, G.; Costa, S.; Teixeira, J.P.; Costa Pereira, C.; et al. Minimum Information for Reporting on the Comet Assay (MIRCA): Recommendations for Describing Comet Assay Procedures and Results. Nat. Protoc. 2020, 15, 3817–3826. [Google Scholar] [CrossRef]
- Toora, B.D.; Rajagopal, G. Measurement of Creatinine by Jaffe’s Reaction--Determination of Concentration of Sodium Hydroxide Required for Maximum Color Development in Standard, Urine and Protein Free Filtrate of Serum. Indian J. Exp. Biol. 2002, 40, 352–354. [Google Scholar]
- Ellman, G.L. A Colorimetric Method for Determining Low Concentrations of Mercaptans. Arch. Biochem. Biophys. 1958, 74, 443–450. [Google Scholar] [CrossRef]
- Drury, J.A.; Nycyk, J.A.; Cooke, R.W. Comparison of Urinary and Plasma Malondialdehyde in Preterm Infants. Clin. Chim. Acta. 1997, 263, 177–185. [Google Scholar] [CrossRef]
- Mercier, Y.; Gatellier, P.; Renerre, M. Lipid and Protein Oxidation in Vitro, and Antioxidant Potential in Meat from Charolais Cows Finished on Pasture or Mixed Diet. Meat Sci. 2004, 66, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gerić, M.; Nanić, L.; Micek, V.; Novak Jovanović, I.; Gajski, G.; Rašić, D.; Orct, T.; Ljubojević, M.; Karaica, D.; Jurasović, J.; et al. The Impact of Resveratrol and Melatonin on the Genome and Oxidative Status in Ageing Rats. Nutrients 2025, 17, 1187. https://doi.org/10.3390/nu17071187
Gerić M, Nanić L, Micek V, Novak Jovanović I, Gajski G, Rašić D, Orct T, Ljubojević M, Karaica D, Jurasović J, et al. The Impact of Resveratrol and Melatonin on the Genome and Oxidative Status in Ageing Rats. Nutrients. 2025; 17(7):1187. https://doi.org/10.3390/nu17071187
Chicago/Turabian StyleGerić, Marko, Lucia Nanić, Vedran Micek, Ivana Novak Jovanović, Goran Gajski, Dubravka Rašić, Tatjana Orct, Marija Ljubojević, Dean Karaica, Jasna Jurasović, and et al. 2025. "The Impact of Resveratrol and Melatonin on the Genome and Oxidative Status in Ageing Rats" Nutrients 17, no. 7: 1187. https://doi.org/10.3390/nu17071187
APA StyleGerić, M., Nanić, L., Micek, V., Novak Jovanović, I., Gajski, G., Rašić, D., Orct, T., Ljubojević, M., Karaica, D., Jurasović, J., Vrhovac Madunić, I., Peraica, M., Sabolić, I., de Andrade, V. M., Breljak, D., & Rubelj, I. (2025). The Impact of Resveratrol and Melatonin on the Genome and Oxidative Status in Ageing Rats. Nutrients, 17(7), 1187. https://doi.org/10.3390/nu17071187