
Organ-Specific Responses to Chronic High-Fat Diets in Mice: Insights into Phospholipid Fatty Acid Distribution

 ,
,  ,
,  , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Chronic High-Fat Diets
2.3. Organ Sample Collection and Preparation
2.4. Analysis of Phospholipid Fatty Acids in Brain, Kidney, and White Adipose Tissue
2.5. Estimation of Enzyme System Activities
- 1.
- Elongase Activity (EA):
- 2.
- Δ9 Desaturase Activity (D9D):
- 3.
- Δ6 Desaturase Activity (D6D):
- 4.
- Δ5 Desaturase Activity (D5D):
2.6. Analysis of Serum Lipid Profile, Cardiovascular Indices, and C-Reactive Protein
- 1.
- Atherogenic Index (AI): ,
- 2.
- Atherogenic Coefficient (AC): ,
- 3.
- Cardiac Risk Ratio (CRR): ,
- 4.
- Cardioprotective Index (CPI): .
2.7. Data and Statistical Evaluation
3. Results
3.1. Brain Phospholipid Fatty Acid Distributions
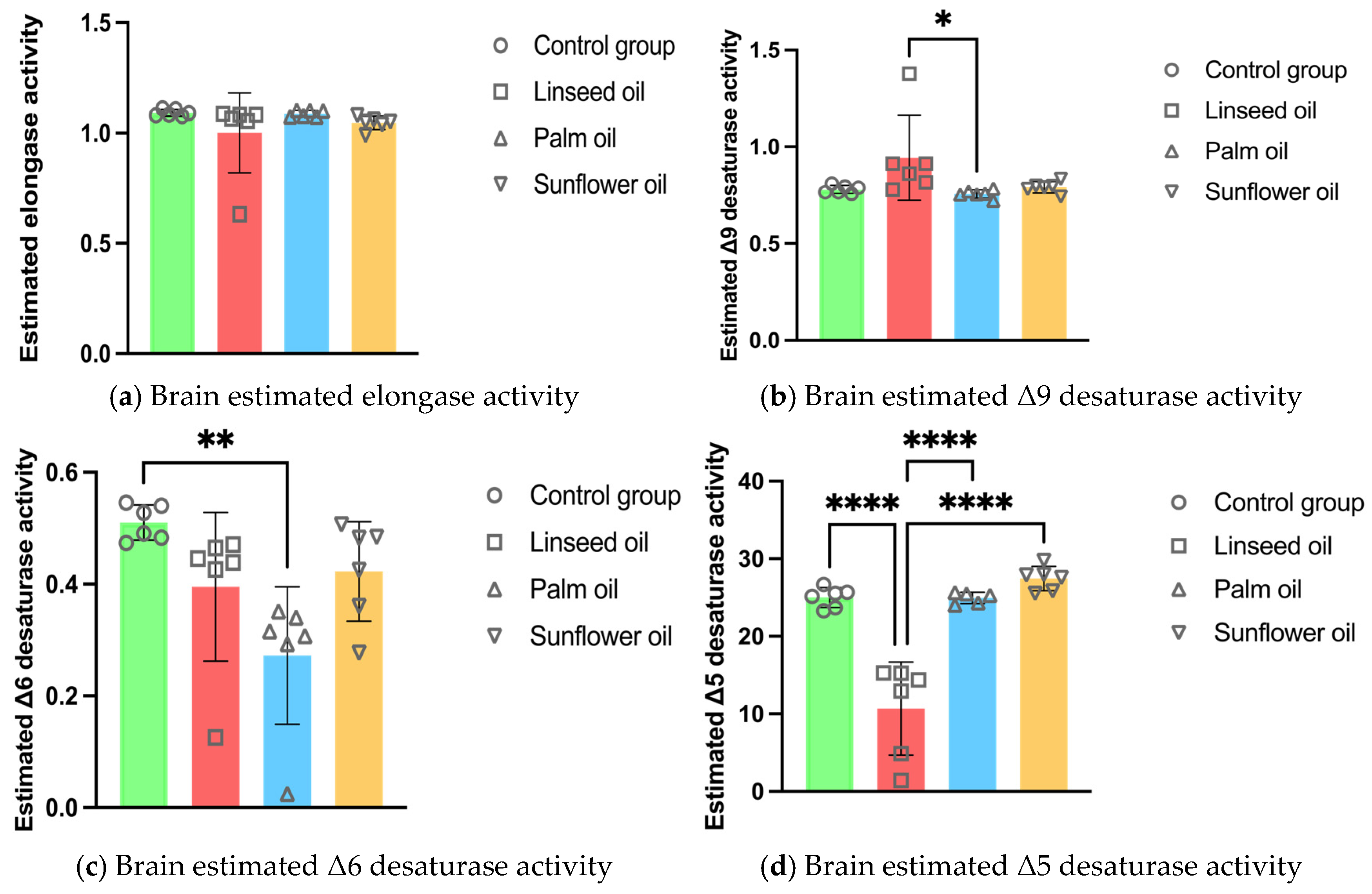
3.2. Brain Enzyme Activity Estimations
3.3. Kidney Phospholipid Fatty Acid Distributions
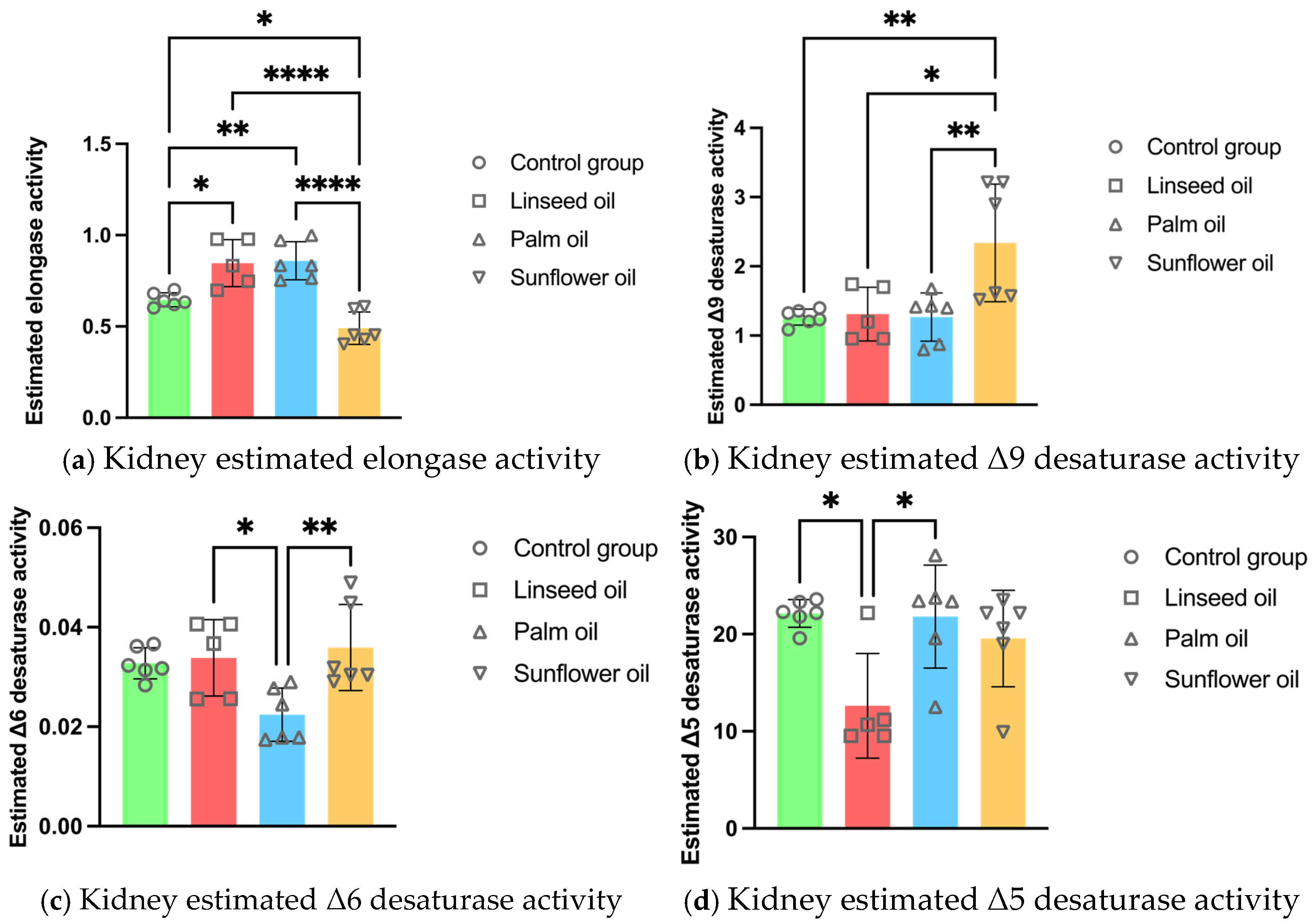
3.4. Kidney Enzyme Activity Estimations
3.5. White Adipose Tissue Phospholipid Fatty Acid Distributions
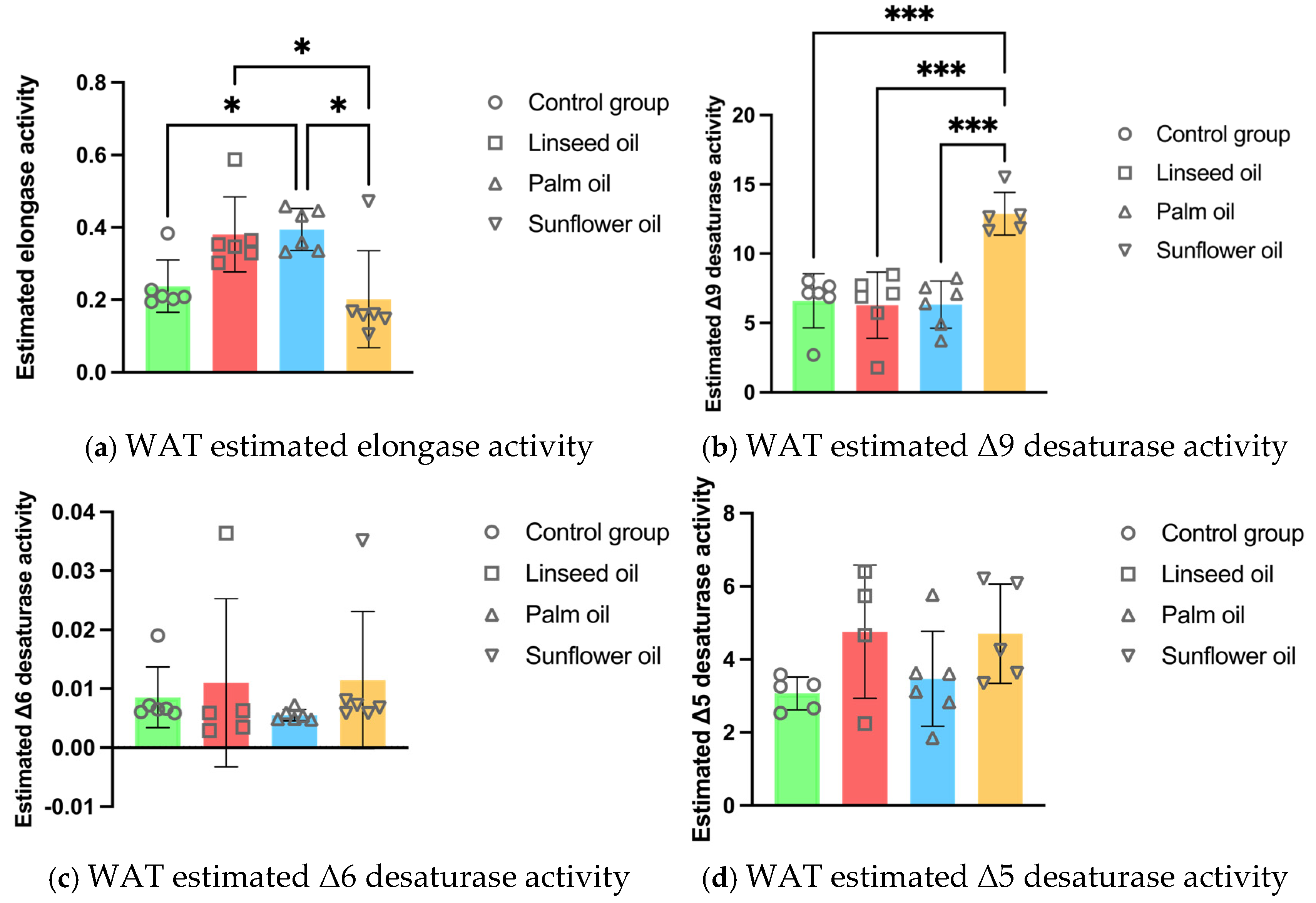
3.6. White Adipose Tissue Enzyme Activity Estimations
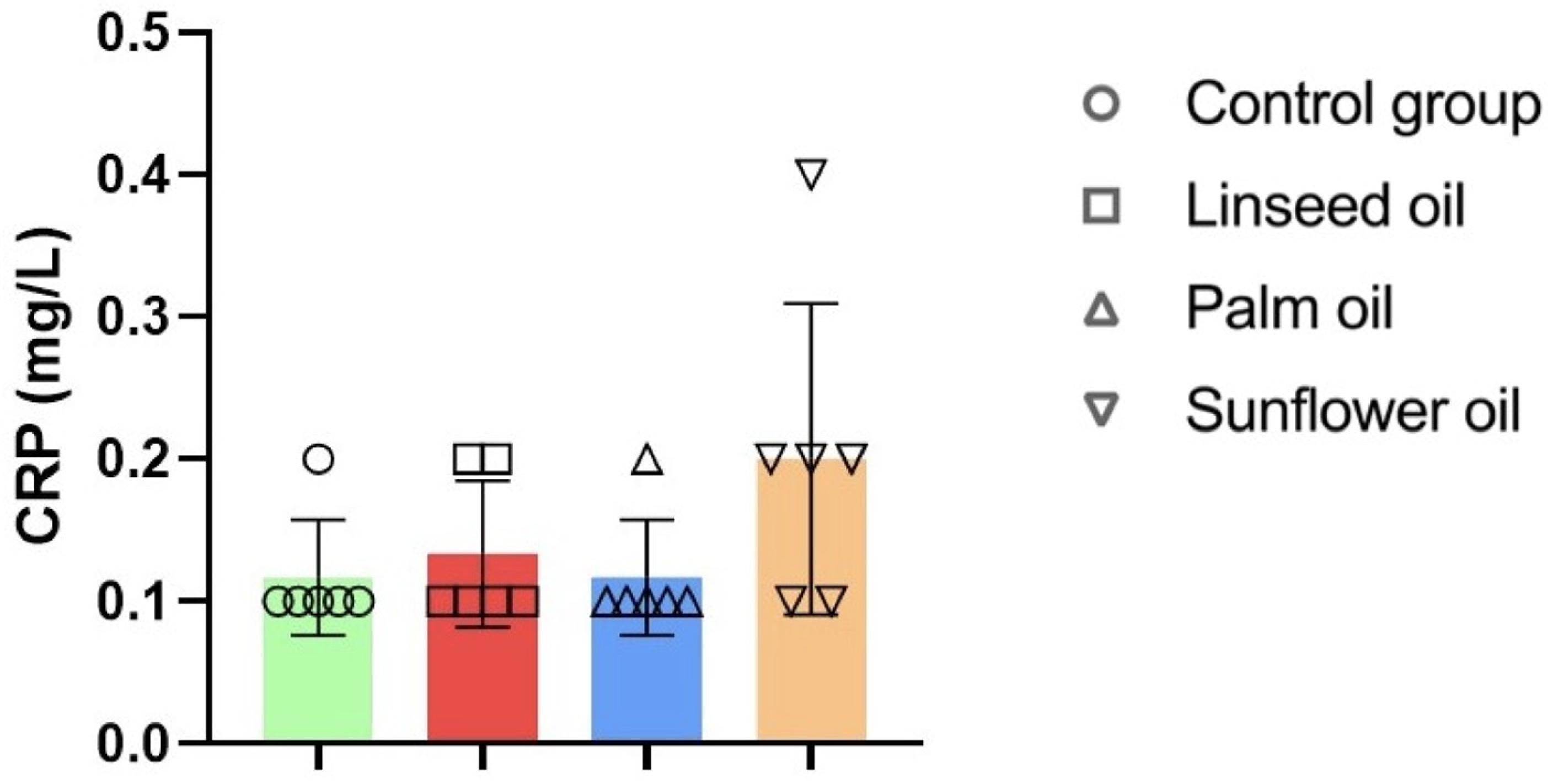
3.7. Serum Lipid Profile, Cardiovascular Indices, and C-Reactive Protein
3.8. Serum Lipid Correlations with Fatty Acid Distribution
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| EFA | Essential fatty acid |
| LA | Linoleic acid (18:2, n-6) |
| ALA | Alpha-linolenic acid (18:3, n-3) |
| PUFA | Polyunsaturated fatty acid |
| EPA | Eicosapentaenoic acid (20:5, n-3) |
| DHA | Docosahexaenoic acid (22:6, n-3) |
| SFA | Saturated fatty acid |
| PA | Palmitic acid (16:0) |
| STA | Stearic acid (18:0) |
| MUFA | Monounsaturated fatty acid |
| OA | Oleic acid (18:1, n-9) |
| PL | Phospholipid |
| AA | Arachidonic acid (20:4, n-6) |
| PLFA | Phospholipid fatty acid |
| CKD | Chronic kidney disease |
| WAT | White adipose tissue |
| BHT | Butylated hydroxytoluene |
| TLC | Thin-layer chromatography |
| GLC | Gas–liquid chromatography |
| EA | Elongase activity |
| D9D | Δ9 Desaturase activity |
| D6D | Δ6 Desaturase activity |
| GLA | Gamma-linolenic acid (18:3, n-6) |
| D5D | Δ5 Desaturase activity |
| DGLA | Dihomo-gamma-linolenic acid (20:3, n-6) |
| TC | Total cholesterol |
| HDL | High-density lipoprotein |
| LDL | Low-density lipoprotein |
| TG | Triglyceride |
| CRP | C-reactive protein |
| AI | Atherogenic Index |
| AC | Atherogenic Coefficient |
| CRR | Cardiac Risk Ratio |
| CPI | Cardioprotective Index |
| non-HDL-c | non-HDL cholesterol |
| SD | Standard deviation |
| DPA | Docosapentaenoic acid (22:5, n-3) |
| POA | Palmitoleic acid (16:1, n-7) |
| VA | Vaccenic acid (18:1, n-7) |
| UI | Unsaturation index |
References
- Wang, X.; Song, R.; Clinchamps, M.; Dutheil, F. New Insights into High-Fat Diet with Chronic Diseases. Nutrients 2023, 15, 4031. [Google Scholar] [CrossRef] [PubMed]
- Wilson, B.A.; Pollard, R.D.; Ferguson, D.S. Nutriential Hazards: Macronutrients: Essential Fatty Acids. In Encyclopedia of Food Safety, 1st ed.; Motarjemi, Y., Ed.; Academic Press: Cambridge, MA, USA, 2014; Volume 3, pp. 95–102. [Google Scholar] [CrossRef]
- Al-Madhagy, S.; Ashmawy, N.S.; Mamdouh, A.; Eldahshan, O.A.; Farag, M.A. A Comprehensive Review of the Health Benefits of Flaxseed Oil in Relation to Its Chemical Composition and Comparison with Other Omega-3-Rich Oils. Eur. J. Med. Res. 2023, 28, 240. [Google Scholar] [CrossRef] [PubMed]
- Mancini, A.; Imperlini, E.; Nigro, E.; Montagnese, C.; Daniele, A.; Orrù, S.; Buono, P. Biological and Nutritional Properties of Palm Oil and Palmitic Acid: Effects on Health. Molecules 2015, 20, 17339–17361. [Google Scholar] [CrossRef] [PubMed]
- Akkaya, M.R. Prediction of Fatty Acid Composition of Sunflower Seeds by Near-Infrared Reflectance Spectroscopy. J. Food Sci. Technol. 2018, 55, 2318–2325. [Google Scholar] [CrossRef]
- Ventura, R.; Martínez-Ruiz, I.; Hernández-Álvarez, M.I. Phospholipid Membrane Transport and Associated Diseases. Biomedicines 2022, 10, 1201. [Google Scholar] [CrossRef]
- Hąc-Wydro, K.; Wydro, P. The Influence of Fatty Acids on Model Cholesterol/Phospholipid Membranes. Chem. Phys. Lipids 2007, 150, 66–81. [Google Scholar] [CrossRef]
- Wang, B.; Tontonoz, P. Phospholipid Remodeling in Physiology and Disease. Annu. Rev. Physiol. 2019, 81, 165–188. [Google Scholar] [CrossRef]
- Clemente-Suárez, V.J.; Beltrán-Velasco, A.I.; Redondo-Flórez, L.; Martín-Rodríguez, A.; Tornero-Aguilera, J.F. Global Impacts of Western Diet and Its Effects on Metabolism and Health: A Narrative Review. Nutrients 2023, 15, 2749. [Google Scholar] [CrossRef]
- Innes, J.K.; Calder, P.C. Omega-6 Fatty Acids and Inflammation. Prostaglandins Leukot. Essent. Fat. Acids 2018, 132, 41–48. [Google Scholar] [CrossRef]
- Calder, P.C. Omega-3 Fatty Acids and Inflammatory Processes: From Molecules to Man. Biochem. Soc. Trans. 2017, 45, 1105–1115. [Google Scholar] [CrossRef]
- Gonzalez-Becerra, K.; Barron-Cabrera, E.; Muñoz-Valle, J.F.; Torres-Castillo, N.; Rivera-Valdes, J.J.; Rodriguez-Echevarria, R.; Martinez-Lopez, E. A Balanced Dietary Ratio of n-6:n-3 Polyunsaturated Fatty Acids Exerts an Effect on Total Fatty Acid Profile in RBCs and Inflammatory Markers in Subjects with Obesity. Healthcare 2023, 11, 2333. [Google Scholar] [CrossRef] [PubMed]
- Fenton, J.I.; Gurzell, E.A.; Davidson, E.A.; Harris, W.S. Red Blood Cell PUFAs Reflect the Phospholipid PUFA Composition of Major Organs. Prostaglandins Leukot. Essent. Fat. Acids 2016, 112, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Tu, W.C.; Mühlhäusler, B.S.; Yelland, L.N.; Gibson, R.A. Correlations between Blood and Tissue Omega-3 LCPUFA Status Following Dietary ALA Intervention in Rats. Prostaglandins Leukot. Essent. Fat. Acids 2013, 88, 53–60. [Google Scholar] [CrossRef]
- Melo, H.M.; Santos, L.E.; Ferreira, S.T. Diet-Derived Fatty Acids, Brain Inflammation, and Mental Health. Front. Neurosci. 2019, 13, 265. [Google Scholar] [CrossRef] [PubMed]
- Custers; Emma, E.M.; Kiliaan; Amanda, J. Dietary Lipids from Body to Brain. Prog. Lipid Res. 2022, 85, 101144. [Google Scholar] [CrossRef]
- Sánchez-Alegría, K.; Arias, C. Functional Consequences of Brain Exposure to Saturated Fatty Acids: From Energy Metabolism and Insulin Resistance to Neuronal Damage. Endocrinol. Diabetes Metab. 2022, 6, e386. [Google Scholar] [CrossRef]
- Dighriri, I.M.; Alsubaie, A.M.; Hakami, F.M.; Hamithi, D.M.; Alshekh, M.M.; Khobrani, F.A.; Dalak, F.E.; Hakami, A.A.; Alsueaadi, E.H.; Alsaawi, L.S.; et al. Effects of Omega-3 Polyunsaturated Fatty Acids on Brain Functions: A Systematic Review. Cureus 2022, 14, e30091. [Google Scholar] [CrossRef]
- De Boer, I.H.; Utzschneider, K.M. The Kidney’s Role in Systemic Metabolism—Still Much to Learn. Nephrol. Dial. Transplant. 2017, 32, 588–590. [Google Scholar] [CrossRef]
- Rangel Silvares, R.; Nunes Goulart da Silva Pereira, E.; Eduardo Ilaquita Flores, E.; Lino Rodrigues, K.; Ribeiro Silva, A.; Gonçalves-de-Albuquerque, C.F.; Daliry, A. High-Fat Diet-Induced Kidney Alterations in Rats with Metabolic Syndrome: Endothelial Dysfunction and Decreased Antioxidant Defense. Diabetes Metab. Syndr. Obes. Targets Ther. 2019, 12, 1773–1781. [Google Scholar] [CrossRef]
- Sun, Y.; Ge, X.; Li, X.; He, J.; Wei, X.; Du, J.; Sun, J.; Li, X.; Xun, Z.; Liu, W.; et al. High-Fat Diet Promotes Renal Injury by Inducing Oxidative Stress and Mitochondrial Dysfunction. Cell Death Dis. 2020, 11, 914. [Google Scholar] [CrossRef]
- Kawai, T.; Autieri, M.V.; Scalia, R. Adipose Tissue Inflammation and Metabolic Dysfunction in Obesity. Am. J. Physiol.-Cell Physiol. 2021, 320, C375–C391. [Google Scholar] [CrossRef] [PubMed]
- Piché, M.-E.; Tchernof, A.; Després, J.-P. Obesity Phenotypes, Diabetes, and Cardiovascular Diseases. Circ. Res. 2020, 126, 1477–1500. [Google Scholar] [CrossRef] [PubMed]
- Kahn, C.R.; Wang, G.; Lee, K.Y. Altered Adipose Tissue and Adipocyte Function in the Pathogenesis of Metabolic Syndrome. J. Clin. Investig. 2019, 129, 3990–4000. [Google Scholar] [CrossRef]
- Popović, T.; Nenadović, A.; Stanković, A.; Debeljak Martačić, J.; Ranković, S.; Kovačević, S.; Nešović Ostojić, J.; Ilić, A.; Milašin, J.; De Luka, S.; et al. Liver Phospholipid Fatty Acid Composition in Response to Chronic High-Fat Diets. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2024, 1869, 159526. [Google Scholar] [CrossRef]
- Schreyer, S.A.; Wilson, D.L.; LeBoeuf, R.C. C57BL/6 Mice Fed High Fat Diets as Models for Diabetes-Accelerated Atherosclerosis. Atherosclerosis 1998, 136, 17–24. [Google Scholar] [CrossRef]
- Funkat, A.; Massa, C.M.; Jovanovska, V.; Proietto, J.; Andrikopoulos, S. Metabolic Adaptations of Three Inbred Strains of Mice (C57BL/6, DBA/2, and 129T2) in Response to a High-Fat Diet. J. Nutr. 2004, 134, 3264–3269. [Google Scholar] [CrossRef]
- Dutta, S.; Sengupta, P. Men and Mice: Relating Their Ages. Life Sci. 2016, 152, 244–248. [Google Scholar] [CrossRef]
- Arifin, W.N.; Zahiruddin, W.M. Sample Size Calculation in Animal Studies Using Resource Equation Approach. Malays. J. Med. Sci. 2017, 24, 101–105. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A Simple Method for the Isolation and Purification of Total Lipids from Animal Tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Daneshmand, R.; Kurl, S.; Tuomainen, T.-P.; Virtanen, J.K. Associations of Estimated Δ-5-Desaturase and Δ-6-Desaturase Activities with Stroke Risk Factors and Risk of Stroke: The Kuopio Ischaemic Heart Disease Risk Factor Study. Br. J. Nutr. 2017, 117, 582–590. [Google Scholar] [CrossRef]
- Domínguez-López, I.; Arancibia-Riveros, C.; Tresserra-Rimbau, A.; Castro-Barquero, S.; Casas, R.; Vázquez-Ruiz, Z.; Ros, E.; Fitó, M.; Estruch, R.; López-Sabater, M.C.; et al. Relationship between Estimated Desaturase Enzyme Activity and Metabolic Syndrome in a Longitudinal Study. Front. Nutr. 2022, 9, 991277. [Google Scholar] [CrossRef] [PubMed]
- Oršolić, N.; Landeka Jurčević, I.; Đikić, D.; Rogić, D.; Odeh, D.; Balta, V.; Perak Junaković, E.; Terzić, S.; Jutrić, D. Effect of Propolis on Diet-Induced Hyperlipidemia and Atherogenic Indices in Mice. Antioxidants 2019, 8, 156. [Google Scholar] [CrossRef] [PubMed]
- Gimenez, M.S.; Oliveros, L.B.; Gomez, N.N. Nutritional Deficiencies and Phospholipid Metabolism. Int. J. Mol. Sci. 2011, 12, 2408–2433. [Google Scholar] [CrossRef] [PubMed]
- Bacle, A.; Kadri, L.; Khoury, S.; Ferru-Clément, R.; Faivre, J.-F.; Cognard, C.; Bescond, J.; Krzesiak, A.; Contzler, H.; Delpech, N.; et al. A Comprehensive Study of Phospholipid Fatty Acid Rearrangements in Metabolic Syndrome: Correlations with Organ Dysfunction. Dis. Models Mech. 2020, 13, dmm043927. [Google Scholar] [CrossRef]
- Paiva, K.M.; Oliveira, R.F.; Helena, L.; Madruga, A.; Sousa, G.; Freire, K.F.; De, J.; Araújo, L.; Lopes, R. Physical Exercise and Flaxseed Oil Supplementation Influence the Glial Plasticity in the Rat Hippocampus. Acta Neurobiol. Exp. 2023, 82, 448–461. [Google Scholar] [CrossRef]
- Butler, M.J.; Mackey-Alfonso, S.E.; Massa, N.; Baskin, K.K.; Barrientos, R.M. Dietary Fatty Acids Differentially Impact Phagocytosis, Inflammatory Gene Expression, and Mitochondrial Respiration in Microglial and Neuronal Cell Models. Front. Cell. Neurosci. 2023, 17, 1227241. [Google Scholar] [CrossRef]
- Wysoczański, T.; Sokoła-Wysoczańska, E.; Pękala, J.; Lochyński, S.; Czyż, K.; Bodkowski, R.; Herbinger, G.; Patkowska-Sokoła, B.; Librowski, T. Omega-3 Fatty Acids and Their Role in Central Nervous System—A Review. Curr. Med. Chem. 2016, 23, 816–831. [Google Scholar] [CrossRef]
- Fisette, A.; Sergi, D.; Breton-Morin, A.; Descôteaux, S.; Martinoli, M.-G. New Insights on the Role of Bioactive Food Derivatives in Neurodegeneration and Neuroprotection. Curr. Pharm. Des. 2022, 28, 3068–3081. [Google Scholar] [CrossRef]
- Liu, B.; Zhang, Y.; Yang, Z.; Liu, M.; Zhang, C.; Zhao, Y.; Song, C. ω-3 DPA Protected Neurons from Neuroinflammation by Balancing Microglia M1/M2 Polarizations through Inhibiting NF-ΚB/MAPK P38 Signaling and Activating Neuron-BDNF-PI3K/AKT Pathways. Mar. Drugs 2021, 19, 587. [Google Scholar] [CrossRef]
- Song, J.; Kim, Y.-S.; Lee, D.H.; Lee, S.H.; Park, H.J.; Lee, D.; Kim, H. Neuroprotective Effects of Oleic Acid in Rodent Models of Cerebral Ischaemia. Sci. Rep. 2019, 9, 10732. [Google Scholar] [CrossRef]
- Wang, X.; Lin, H.; Gu, Y. Multiple Roles of Dihomo-γ-Linolenic Acid against Proliferation Diseases. Lipids Health Dis. 2012, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- Mustonen, A.-M.; Nieminen, P. Dihomo-γ-Linolenic Acid (20:3n-6)—Metabolism, Derivatives, and Potential Significance in Chronic Inflammation. Int. J. Mol. Sci. 2023, 24, 2116. [Google Scholar] [CrossRef] [PubMed]
- Chang, Z.; Yang, M.; Ji, H. Molecular Characterization and Functional Analysis of Apoptosis-Inducing Factor (AIF) in Palmitic Acid-Induced Apoptosis in Ctenopharyngodon Idellus Kidney (CIK) Cells. Fish Physiol. Biochem. 2021, 47, 213–224. [Google Scholar] [CrossRef]
- Gillingham, L.G.; Harris-Janz, S.; Jones, P.J.H. Dietary Monounsaturated Fatty Acids Are Protective against Metabolic Syndrome and Cardiovascular Disease Risk Factors. Lipids 2011, 46, 209–228. [Google Scholar] [CrossRef]
- Gwon, D.; Hwang, T.; Ro, J.-Y.; Kang, Y.-J.; Jeong, J.; Kim, D.-K.; Lim, K.; Kim, D.; Choi, D.; Kim, J.-J. High Endogenous Accumulation of ω-3 Polyunsaturated Fatty Acids Protect against Ischemia-Reperfusion Renal Injury through AMPK-Mediated Autophagy in Fat-1 Mice. Int. J. Mol. Sci. 2017, 18, 2081. [Google Scholar] [CrossRef]
- Ajami, M.; Davoodi, S.H.; Habibey, R.; Namazi, N.; Soleimani, M.; Pazoki-Toroudi, H. Effect of DHA+EPA on Oxidative Stress and Apoptosis Induced by Ischemia-Reperfusion in Rat Kidneys. Fundam. Clin. Pharmacol. 2012, 27, 593–602. [Google Scholar] [CrossRef]
- Ashtiyani, S.C.; Najafi, H.; Kabirinia, K.; Vahedi, E.; Jamebozorky, L. Oral Omega-3 Fatty Acid for Reduction of Kidney Dysfunction Induced by Reperfusion Injury in Rats. Iran. J. Kidney Dis. 2012, 6, 275–283. [Google Scholar]
- Liput, K.P.; Lepczyński, A.; Ogłuszka, M.; Nawrocka, A.; Poławska, E.; Grzesiak, A.; Ślaska, B.; Pareek, C.S.; Czarnik, U.; Pierzchała, M. Effects of Dietary N–3 and N–6 Polyunsaturated Fatty Acids in Inflammation and Cancerogenesis. Int. J. Mol. Sci. 2021, 22, 6965. [Google Scholar] [CrossRef]
- Shioda, R.; Jo-Watanabe, A.; Lee-Okada, H.-C.; Yasukawa, K.; Okuno, T.; Suzuki, Y.; Yokomizo, T. Dietary Intake of N-3 Polyunsaturated Fatty Acids Alters the Lipid Mediator Profile of the Kidney but Does Not Attenuate Renal Insufficiency. Biochem. Biophys. Res. Commun. 2021, 582, 49–56. [Google Scholar] [CrossRef]
- Rund, K.M.; Peng, S.; Greite, R.; Claaßen, C.; Nolte, F.; Oger, C.; Galano, J.-M.; Balas, L.; Durand, T.; Chen, R.; et al. Dietary Omega-3 PUFA Improved Tubular Function after Ischemia Induced Acute Kidney Injury in Mice but Did Not Attenuate Impairment of Renal Function. Prostaglandins Other Lipid Mediat. 2019, 146, 106386. [Google Scholar] [CrossRef]
- Liu, K.; Acharjee, A.; Hinz, C.; Liggi, S.; Murgia, A.; Denes, J.; Gulston, M.K.; Wang, X.; Chu, Y.; West, J.A.; et al. Consequences of Lipid Remodeling of Adipocyte Membranes Being Functionally Distinct from Lipid Storage in Obesity. J. Proteome Res. 2020, 19, 3919–3935. [Google Scholar] [CrossRef] [PubMed]
- Holzer, R.G.; Park, E.-J.; Li, N.; Tran, H.; Chen, M.; Choi, C.; Solinas, G.; Karin, M. Saturated Fatty Acids Induce C-Src Clustering within Membrane Subdomains, Leading to JNK Activation. Cell 2011, 147, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Solinas, G.; Naugler, W.; Galimi, F.; Lee, M.-S.; Karin, M. Saturated Fatty Acids Inhibit Induction of Insulin Gene Transcription by JNK-Mediated Phosphorylation of Insulin-Receptor Substrates. Proc. Natl. Acad. Sci. USA 2006, 103, 16454–16459. [Google Scholar] [CrossRef] [PubMed]
- Palmgren, H.; Petkevicius, K.; Bartesaghi, S.; Ahnmark, A.; Ruiz, M.; Nilsson, R.; Löfgren, L.; Glover, M.S.; Andréasson, A.-C.; Andersson, L.; et al. Elevated Adipocyte Membrane Phospholipid Saturation Does Not Compromise Insulin Signaling. Diabetes 2022, 72, 1350–1363. [Google Scholar] [CrossRef]
- Ravaut, G.; Légiot, A.; Bergeron, K.-F.; Mounier, C. Monounsaturated Fatty Acids in Obesity-Related Inflammation. Int. J. Mol. Sci. 2020, 22, 330. [Google Scholar] [CrossRef]
- Ralston, J.C.; Metherel, A.H.; Stark, K.D.; Mutch, D.M. SCD1 Mediates the Influence of Exogenous Saturated and Monounsaturated Fatty Acids in Adipocytes: Effects on Cellular Stress, Inflammatory Markers and Fatty Acid Elongation. J. Nutr. Biochem. 2016, 27, 241–248. [Google Scholar] [CrossRef]
- Martins, B.C.; Ribeiro, S.; Teixeira, S.; Peixoto, T.C.; Lisboa, P.C.; Martins, F.F.; Souza-Mello, V.; Daleprane, J.B. Consumption of Interesterified Palm Oil Leads Inflammation of White Adipose Tissue and Triggers Metabolic Disturbances in Mice on a High-Fat Diet. Sci. Rep. 2024, 14, 12530. [Google Scholar] [CrossRef]
- Li, R.; Cao, C.; Zheng, Z.; Yang, X.; Tan, C.P.; Xu, Y.; Liu, Y. Palm Oil Consumption and Its Repercussion on Endogenous Fatty Acids Distribution. Food Funct. 2021, 12, 2020–2031. [Google Scholar] [CrossRef]
- Saavedra-Peña, R.D.M.; Taylor, N.; Flannery, C.; Rodeheffer, M.S. Estradiol Cycling Drives Female Obesogenic Adipocyte Hyperplasia. Cell Rep. 2023, 42, 112390. [Google Scholar] [CrossRef]
- Tóth, M.E.; Dukay, B.; Péter, M.; Balogh, G.; Szűcs, G.; Zvara, Á.; Szebeni, G.J.; Hajdu, P.; Sárközy, M.; Puskás, L.G.; et al. Male and Female Animals Respond Differently to High-Fat Diet and Regular Exercise Training in a Mouse Model of Hyperlipidemia. Int. J. Mol. Sci. 2021, 22, 4198. [Google Scholar] [CrossRef]
- Walker, C.G.; Browning, L.M.; Mander, A.P.; Madden, J.; West, A.L.; Calder, P.C.; Jebb, S.A. Age and Sex Differences in the Incorporation of EPA and DHA into Plasma Fractions, Cells and Adipose Tissue in Humans. Br. J. Nutr. 2013, 111, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Seike, M.; Ashida, H.; Yamashita, Y. Dietary Flaxseed Oil Induces Production of Adiponectin in Visceral Fat and Prevents Obesity in Mice. Nutr. Res. 2023, 121, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Baranowski, M.; Enns, J.; Blewett, H.; Yakandawala, U.; Zahradka, P.; Taylor, C.G. Dietary Flaxseed Oil Reduces Adipocyte Size, Adipose Monocyte Chemoattractant Protein-1 Levels and T-Cell Infiltration in Obese, Insulin-Resistant Rats. Cytokine 2012, 59, 382–391. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, D.; Lavie, C.J.; Elagizi, A.; Milani, R.V. Update on Omega-3 Polyunsaturated Fatty Acids on Cardiovascular Health. Nutrients 2022, 14, 5146. [Google Scholar] [CrossRef]
- Parikh, M.; Pierce, G.N. Dietary Flaxseed: What We Know and Don’t Know about Its Effects on Cardiovascular Disease. Can. J. Physiol. Pharmacol. 2019, 97, 75–81. [Google Scholar] [CrossRef]
- Dupasquier, C.M.C.; Dibrov, E.; Kneesh, A.L.; Cheung, P.K.M.; Lee, K.G.Y.; Alexander, H.K.; Yeganeh, B.K.; Moghadasian, M.H.; Pierce, G.N. Dietary Flaxseed Inhibits Atherosclerosis in the LDL Receptor-Deficient Mouse in Part through Antiproliferative and Anti-Inflammatory Actions. Am. J. Physiol.-Heart Circ. Physiol. 2007, 293, H2394–H2402. [Google Scholar] [CrossRef]
- Ismail, S.R.; Maarof, S.K.; Siedar, A.S.; Ali, A. Systematic Review of Palm Oil Consumption and the Risk of Cardiovascular Disease. PLoS ONE 2018, 13, e0193533. [Google Scholar] [CrossRef]
- Cai, H.; Zhang, J.; Liu, C.; Le, T.N.; Lu, Y.; Feng, F.; Zhao, M. High-Fat Diet-Induced Decreased Circulating Bile Acids Contribute to Obesity Associated with Gut Microbiota in Mice. Foods 2024, 13, 699. [Google Scholar] [CrossRef]
- Malesza, I.J.; Malesza, M.; Walkowiak, J.; Mussin, N.; Walkowiak, D.; Aringazina, R.; Bartkowiak-Wieczorek, J.; Mądry, E. High-Fat, Western-Style Diet, Systemic Inflammation, and Gut Microbiota: A Narrative Review. Cells 2021, 10, 3164. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid | Control Group (%) | Linseed Oil (%) | Palm Oil (%) | Sunflower Oil (%) |
|---|---|---|---|---|
| SFA | 45.34 ± 1.35 | 42.04 ± 5.7 | 46.15 ± 1.15 | 45.64 ± 0.84 |
| PA (16:0) | 21.68 ± 0.71 | 21.28 ± 4.38 | 22.1 ± 0.56 | 22.31 ± 0.71 |
| STA (18:0) | 23.67 ± 0.68 | 20.76 ± 3.03 a | 24.05 ± 0.63 bb | 23.33 ± 0.27 b |
| MUFA | 22.91 ± 0.74 | 23.88 ± 4.99 | 22.4 ± 0.91 | 22.62 ± 0.73 |
| POA (16:1, n-7) | 0.48 ± 0.04 | 0.55 ± 0.12 | 0.44 ± 0.02 | 0.48 ± 0.02 |
| OA (18:1, n-9) | 18.46 ± 0.61 | 19.35 ± 3.72 | 18.21 ± 0.76 | 18.44 ± 0.61 |
| VA (18:1, n-7) | 3.97 ± 0.11 | 3.73 ± 0.76 | 3.75 ± 0.16 | 3.69 ± 0.17 |
| PUFA | 31.75 ± 1.97 | 32.9 ± 4.39 | 31.45 ± 1.87 | 31.74 ± 0.55 |
| n-6 | 14.06 ± 0.54 | 14.7 ± 1.49 | 14.22± 2.5 | 14.55 ± 0.19 |
| LA (18:2, n-6) | 0.73 ± 0.04 | 1.29 ± 0.25 aaa | 1.31 ± 0.18 aaa | 0.89 ± 0.23 b,cc |
| GLA (18:3, n-6) | 0.51 ± 0.07 | 1.04 ± 1.38 | 0.51 ± 0.05 | 0.47 ± 0.06 |
| DGLA (20:3, n-6) | 0.37 ± 0.009 | 0.58 ± 0.09 aa | 0.34 ± 0.15 bbb | 0.36 ± 0.01 bb |
| AA (20:4, n-6) | 9.27 ± 0.36 | 6.56 ± 3.98 | 8.94 ± 2.64 | 9.84 ± 0.33 |
| DTA (22:4, n-6) | 3.17 ± 0.35 | 3.06 ± 0.94 | 3.12 ± 0.28 | 2.99 ± 0.41 |
| n-3 | 17.69 ± 2.19 | 16.42 ± 3.59 | 17.23 ± 0.76 | 17.19 ± 0.39 |
| ALA (18:3, n-3) | 3.48 ± 2.55 | 1.85 ± 0.99 | 2.25 ± 0.23 | 2.16 ± 0.37 |
| EPA (20:5, n-3) | 0.08 ± 0.04 | 0.16 ± 0.03 aa | 0.09 ± 0.04 b | 0.06 ± 0.03 bb |
| DPA (22:5, n-3) | 0.14 ± 0.006 | 0.58 ± 0.1 aa | 0.11 ± 0.01 bb | 0.28 ± 0.41 |
| DHA (22:6, n-3) | 13.99 ± 0.42 | 13.82 ± 3.06 | 14.77 ± 0.75 | 14.68 ± 0.32 |
| n-6/n-3 | 0.8 ± 0.09 | 0.89 ± 0.17 | 0.83 ± 0.17 | 0.85 ± 0.01 |
| EPA/AA | 0.008 ± 0.004 | 0.06 ± 0.08 | 0.01 ± 0.01 | 0.006 ± 0.003 |
| UI | 172.3 ± 4.44 | 161.3 ± 39.39 | 172.2 ± 6.11 | 174.5 ± 2.17 |
| n-3 index | 3.25 ± 0.36 | 3.22 ± 0.95 | 3.21 ± 0.28 | 3.05 ± 0.42 |
| Fatty Acid | Control Group (%) | Linseed Oil (%) | Palm Oil (%) | Sunflower Oil (%) |
|---|---|---|---|---|
| SFA | 39.82 ± 3.2 | 33.7 ± 3.9 a | 33.44 ± 3.3 a | 37.18 ± 2.57 |
| PA (16:0) | 24.21 ± 2.17 | 18.23 ± 2.26 aaa | 17.96± 1.14 aaa | 25.02 ± 2.34 bbb,ccc |
| STA (18:0) | 15.61 ± 1.2 | 15.39 ± 2.24 | 15.48 ± 2.42 | 12.17 ± 1.67 a,b,c |
| MUFA | 23.78 ± 0.89 | 22.19 ± 3.11 | 21.01 ± 2.88 | 30.65 ± 7.39 a,b,cc |
| POA (16:1, n-7) | 2.03 ± 0.31 | 1.39 ± 0.28 a | 0.79 ± 0.23 aaa | 1.67 ± 0.51 cc |
| OA (18:1, n-9) | 19.67 ± 0.58 | 20.18 ± 3.12 | 18.91 ± 2.75 | 27.43 ± 7.12 a,b,cc |
| VA (18:1, n-7) | 2.08 ± 0.11 | 1.31 ± 0.36 aaa | 1.29 ± 0.14 aaa | 1.54 ± 0.26 aa |
| PUFA | 36.4 ± 3.42 | 44.11 ± 2.91 a | 45.55 ± 1.62 aa | 31.33 ± 5.53 bbb,ccc |
| n-6 | 30.08 ± 2.2 | 28.89 ± 2.96 | 39.76 ± 1.6 aaa,bbb | 25.08 ± 3.83 a,ccc |
| LA (18:2, n-6) | 16.52 ± 0.99 | 19.18 ± 1.95 | 26.03 ± 3.82 aaa,bbb | 13.89 ± 1.23 bb,ccc |
| GLA (18:3, n-6) | 0.37 ± 0.07 | 0.46 ± 0.14 | 0.35 ± 0.05 | 0.3 ± 0.01 b |
| DGLA (20:3, n-6) | 0.54 ± 0.05 | 0.65± 0.18 | 0.57 ± 0.11 | 0.49 ± 0.12 |
| AA (20:4, n-6) | 11.98 ± 01.53 | 8.01 ± 3.29 | 12.14 ± 2.43 | 9.81 ± 3.83 |
| DTA (22:4, n-6) | 0.66 ± 0.06 | 0.54 ± 0.26 | 0.67 ± 0.14 | 0.58 ± 0.13 |
| n-3 | 6.32 ± 1.23 | 15.22 ± 4.85 aaa | 5.78 ± 1.65 bbb | 5.64 ± 1.67 bbb |
| ALA (18:3, n-3) | 0.41 ± 0.04 | 5.76 ± 4.49 aa | 0.23 ± 0.11 bb | 0.27 ± 0.05 bb |
| EPA (20:5, n-3) | 0.08 ± 0.04 | 1.5 ± 0.87 aaa | 0.04 ± 0.07 bbb | 0.27 ± 0.51 bb |
| DPA (22:5, n-3) | 0.28 ± 0.05 | 1.64 ± 1.36 a | 0.21 ± 0.08 bb | 0.27 ± 0.19 b |
| DHA (22:6, n-3) | 5.55 ± 1.12 | 5.57 ± 1.22 | 5.30 ± 1.65 | 4.79 ± 1.65 |
| n-6/n-3 | 4.88 ± 0.74 | 2.2 ± 1.16 a | 7.34 ± 2.05 a,bbb | 4.72 ± 0.78 b,c |
| EPA/AA | 0.006 ± 0.003 | 0.22 ± 0.12 aa | 0.003 ± 0.006 bb | 0.05 ± 0.11 b |
| UI | 146.5 ± 14.10 | 145.9 ± 50.45 | 160.8 ± 11.64 | 134.6 ± 20.97 |
| n-3 index | 5.63 ± 1.16 | 7.08 ± 0.93 | 5.33 ± 1.67 | 5.06 ± 1.49 |
| Fatty Acid | Control Group (%) | Linseed Oil (%) | Palm Oil (%) | Sunflower Oil (%) |
|---|---|---|---|---|
| SFA | 28.58 ± 3.51 | 21.6 ± 8.56 | 20.56 ± 6.67 | 31.54 ± 4.12 b,c |
| PA (16:0) | 23.03 ± 1.47 | 15.38 ± 4.59 aa | 14.75 ± 4.67 aa | 26.25 ± 1.88 bbb,ccc |
| STA (18:0) | 5.55 ± 2.13 | 6.22 ± 4.01 | 5.82 ± 2.15 | 5.28 ± 3.49 |
| MUFA | 39.68 ± 3.41 | 35.22 ± 3.47 | 36.17 ± 2.22 | 53.14 ± 1.31 aaa,bbb,ccc |
| POA (16:1, n-7) | 4.47 ± 0.79 | 2.54 ± 0.57 aaa | 1.41 ± 0.43 aaa | 2.82 ± 0.97 aa,c |
| OA (18:1, n-9) | 33.18 ± 3.29 | 31.42 ± 3.24 | 33.92 ± 1.95 | 44.35 ± 10.85 a,bb,c |
| VA (18:1, n-7) | 2.02 ± 0.14 | 1.27 ± 0.08 aaa | 0.84 ± 0.37 aaa,b | 1.28 ± 0.15 aaa,cc |
| PUFA | 31.84 ± 1.34 | 43.17 ± 5.67 | 48.27 ± 13.67 a | 16.86 ± 0.61 a,bbb,ccc |
| n-6 | 29.98 ± 1.27 | 21.41 ± 1.02 | 47.05 ± 13.44 aa,bbb | 15.83 ± 0.64 a,ccc |
| LA (18:2, n-6) | 27.5 ± 3.8 | 20.25 ± 1.29 | 45.65 ± 13.43 aa,bbb | 15.36 ± 1.01 a,ccc |
| GLA (18:3, n-6) | 0.16 ± 0.007 | 0.26 ± 0.02 aa | 0.18 ± 0.09 b | 0.17 ± 0.02 b |
| DGLA (20:3, n-6) | 0.22 ± 0.08 | 0.21 ± 0.25 | 0.24 ± 0.05 | 0.18 ± 0.21 |
| AA (20:4, n-6) | 0.57 ± 0.04 | 0.41 ± 0.18 | 0.82 ± 0.27 b | 0.49 ± 0.21 |
| DTA (22:4, n-6) | 0.11 ± 0.06 | 0.17 ± 0.18 | 0.16 ± 0.09 | 0.14 ± 0.13 |
| n-3 | 1.89 ± 0.12 | 21.53 ± 5.98 aaa | 1.21 ± 0.32 bbb | 1.02 ± 0.28 bbb |
| ALA (18:3, n-3) | 1.42 ± 0.42 | 20.45 ± 6.85 aaa | 0.97 ± 0.28 bbb | 0.71 ± 0.33 bbb |
| EPA (20:5, n-3) | 0.06 ± 0.06 | 0.42 ± 0.61 | ND | 0.11 ± 0.13 |
| DPA (22:5, n-3) | 0.09 ± 0.08 | 0.27 ± 0.17 | ND | 0.11 ± 0.13 |
| DHA (22:6, n-3) | 0.34 ± 0.32 | 0.39 ± 0.19 | 0.24 ± 0.09 | 0.22 ± 0.14 |
| n-6/n-3 | 16.15 ± 0.82 | 0.89 ± 0.03 aa | 39.41 ± 9 aaa,bbb | 16.34 ± 3.93 bb,ccc |
| EPA/AA | 0.04 ± 0.006 | 0.44 ± 0.13 aa | ND | 0.009 ± 0.009 bb |
| UI | 105.0 ± 9.60 | 148.0 ± 15.65 aaa | 137.0 ± 25.67 a | 87.97 ± 5.98 bbb,ccc |
| n-3 index | 0.37 ± 0.38 | 0.81 ± 0.78 | 0.24 ± 0.09 | 0.50 ± 0.67 |
| Parameters | Control Group | Linseed Oil | Palm Oil | Sunflower Oil |
|---|---|---|---|---|
| TC (mmol/L) | 2.00 ± 0.18 | 2.37 ± 0.37 | 2.50 ± 0.54 | 2.45 ± 0.47 |
| LDL (mmol/L) | 0.45 ± 0.08 | 0.23 ± 0.13 a | 0.55 ± 0.16 bbb | 0.59 ± 0.09 bbb |
| HDL (mmol/L) | 1.03 ± 0.10 | 1.45 ± 0.32 a | 1.40 ± 0.25 | 1.25 ± 0.30 |
| non-HDL-c (mmol/L) | 0.96 ± 0.09 | 0.92 ± 0.16 | 1.11 ± 0.27 | 1.15 ± 0.20 |
| TG (mmol/L) | 1.12 ± 0.14 | 1.51 ± 0.29 | 1.23 ± 0.29 | 1.25 ± 0.38 |
| AI | 0.03 ± 0.05 | 0.02± 0.09 | −0.06 ± 0.04 | −0.01 ± 0.06 |
| AC | 0.93 ± 0.06 | 0.66 ± 0.20 a | 0.79 ± 0.05 | 0.94 ± 0.15 bb |
| CRR | 1.90 ± 0.06 | 1.65 ± 0.21 a | 1.78 ± 0.07 | 1.95 ± 0.14 bb |
| CPI | 2.34 ± 0.41 | 9.07 ± 4.93 aaa | 2.62 ± 0.44 bb | 2.16 ± 0.59 bbb |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nenadovic, A.; Kovacevic, S.; Stankovic, A.; Popovic, T.; Debeljak Martacic, J.; Rankovic, S.; De Luka, S.R.; Milasin, J.; Nesovic Ostojic, J. Organ-Specific Responses to Chronic High-Fat Diets in Mice: Insights into Phospholipid Fatty Acid Distribution. Nutrients 2025, 17, 821. https://doi.org/10.3390/nu17050821
Nenadovic A, Kovacevic S, Stankovic A, Popovic T, Debeljak Martacic J, Rankovic S, De Luka SR, Milasin J, Nesovic Ostojic J. Organ-Specific Responses to Chronic High-Fat Diets in Mice: Insights into Phospholipid Fatty Acid Distribution. Nutrients. 2025; 17(5):821. https://doi.org/10.3390/nu17050821
Chicago/Turabian StyleNenadovic, Aleksandra, Sanjin Kovacevic, Anica Stankovic, Tamara Popovic, Jasmina Debeljak Martacic, Slavica Rankovic, Silvio R. De Luka, Jelena Milasin, and Jelena Nesovic Ostojic. 2025. "Organ-Specific Responses to Chronic High-Fat Diets in Mice: Insights into Phospholipid Fatty Acid Distribution" Nutrients 17, no. 5: 821. https://doi.org/10.3390/nu17050821
APA StyleNenadovic, A., Kovacevic, S., Stankovic, A., Popovic, T., Debeljak Martacic, J., Rankovic, S., De Luka, S. R., Milasin, J., & Nesovic Ostojic, J. (2025). Organ-Specific Responses to Chronic High-Fat Diets in Mice: Insights into Phospholipid Fatty Acid Distribution. Nutrients, 17(5), 821. https://doi.org/10.3390/nu17050821