

Beneficial Effects of Butyrate on Kidney Disease


Abstract
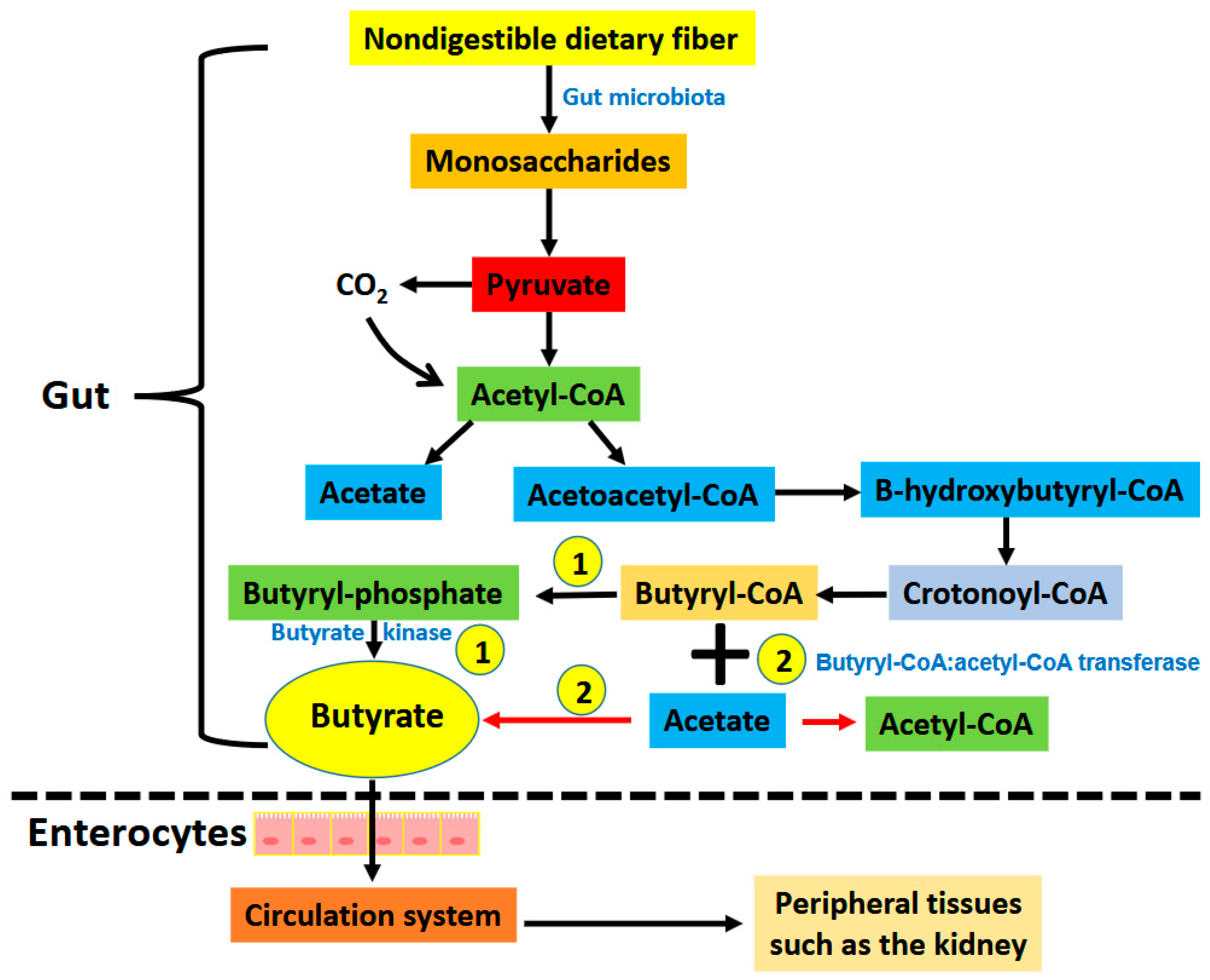
1. Introduction
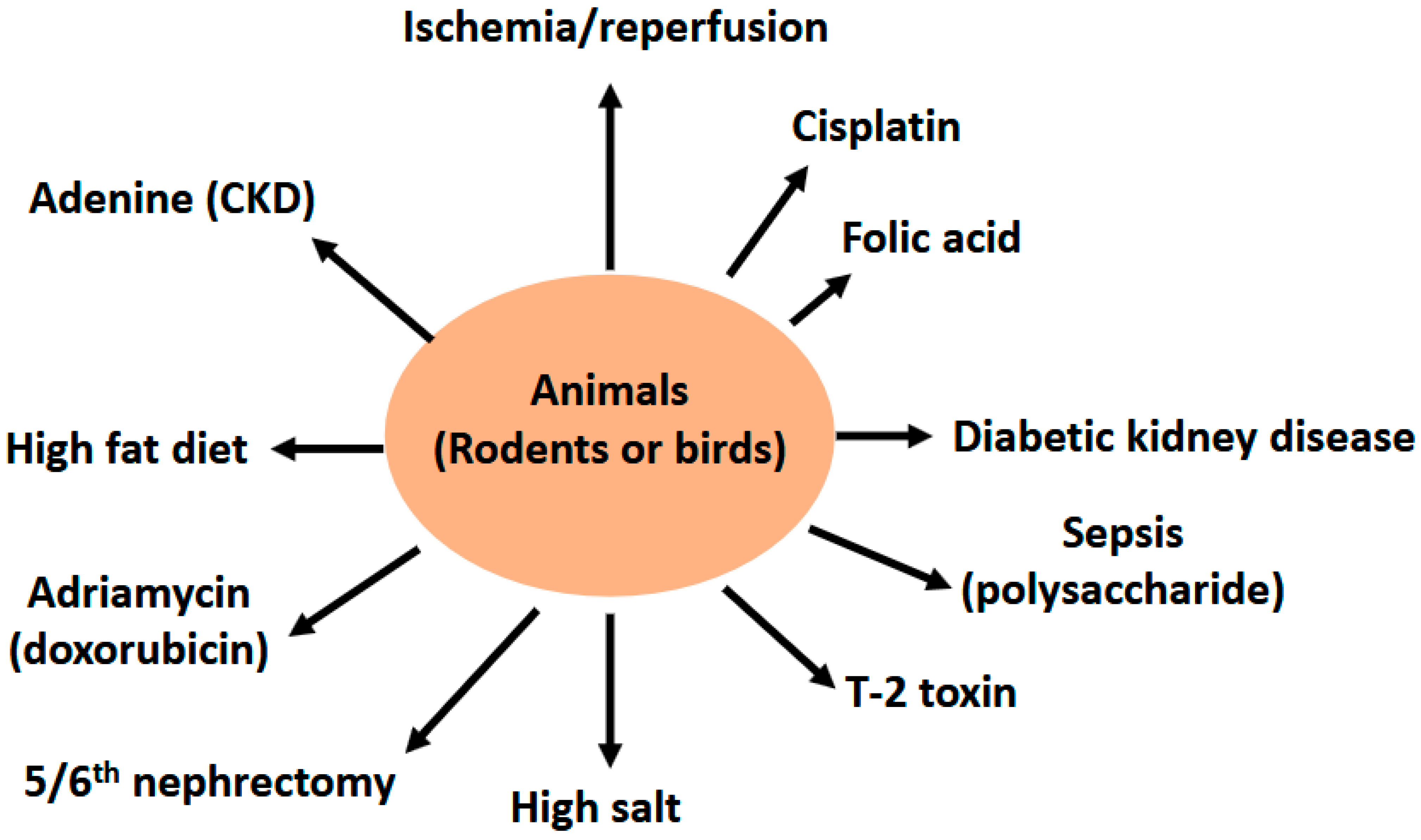
2. Renoprotective Effects of Butyrate on Renal Injuries
2.1. Butyrate and Renal Ischemia—Reperfusion Injury
2.2. Butyrate and Cisplatin—Induced Kidney Injury
2.3. Butyrate and Folic Acid—Induced Kidney Injury
2.4. Butyrate and Lipopolysaccharide (LPS)—Induced Kidney Injury
2.5. Butyrate and Renal Failure Induced by 5/6th Nephrectomy (Nx)
2.6. Butyrate and Diabetic Kidney Disease (DKD)
2.7. Butyrate and High Fat Diet (HFD) Induced Glomerulopathy
2.8. Butyrate and Adenine—Induced Chronic Kidney Disease (CKD)
2.9. Butyrate and Salt—Induced Hypertension and Renal Damage
2.10. Butyrate and Adriamycin—Induced Nephropathy
2.11. Butyrate and Contrast—Induced Nephropathy
2.12. Butyrate and T-2 Toxin Induced Renal Injury
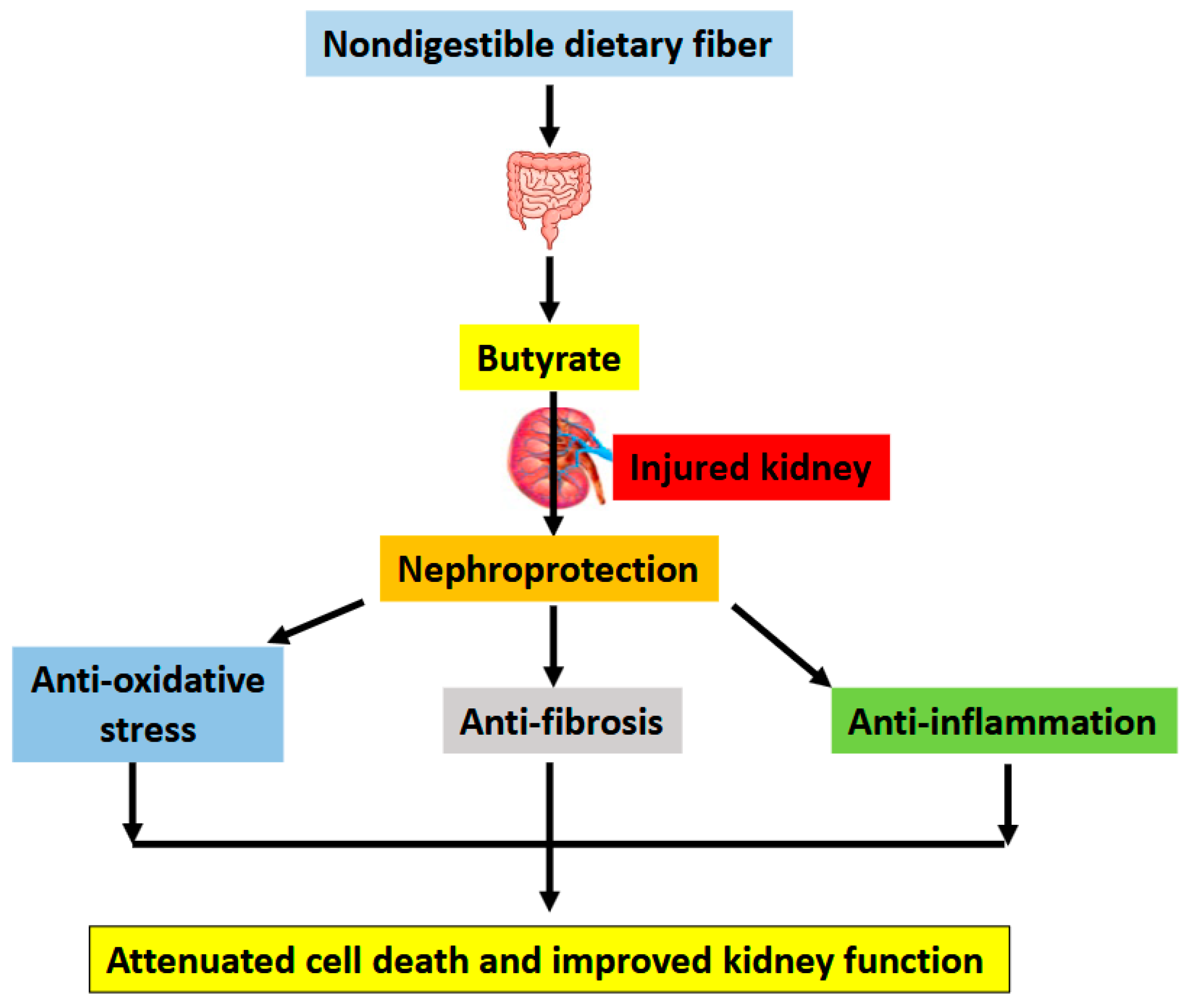
3. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Snyder, R. What You Must Know About Kidney Disease; Square One Publisher: New York, NY, USA, 2010. [Google Scholar]
- Dipiro, J.T.; Talbet, R.L.; Yee, G.C.; Matzke, G.R.; Wells, B.G.; Posey, L.M. Pharmacotherapy: A Pathophysiological Approach, 9th ed.; McGraw-Hill Education: New York, NY, USA, 2014. [Google Scholar]
- Kaneko, K.; Soty, M.; Zitoun, C.; Duchampt, A.; Silva, M.; Philippe, E.; Gautier-Stein, A.; Rajas, F.; Mithieux, G. The role of kidney in the inter-organ coordination of endogenous glucose production during fasting. Mol. Metab. 2018, 16, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Sumida, K.D.; Garrett, J.H.; McJilton, W.T.; Hevener, A.L.; Donovan, C.M. Effect of endurance training and fasting on renal gluconeogenic enzymes in the rat. Int. J. Sport Nutr. Exerc. Metab. 2004, 14, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Kamt, S.F.; Liu, J.; Yan, L.J. Renal-Protective Roles of Lipoic Acid in Kidney Disease. Nutrients 2023, 15, 1732. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.J. NADH/NAD(+) Redox Imbalance and Diabetic Kidney Disease. Biomolecules 2021, 11, 730. [Google Scholar] [CrossRef]
- Serna, J.D.C.; Amaral, A.G.; Caldeira da Silva, C.C.; Munhoz, A.C.; Vilas-Boas, E.A.; Menezes-Filho, S.L.; Kowaltowski, A.J. Regulation of kidney mitochondrial function by caloric restriction. Am. J. Physiol. Ren. Physiol. 2022, 323, F92–F106. [Google Scholar] [CrossRef]
- Liu, H.; Yan, L.-J. The Role of Ketone Bodies in Various Animal Models of Kidney Disease. Endocrines 2023, 4, 236–249. [Google Scholar] [CrossRef]
- Voroneanu, L.; Burlacu, A.; Brinza, C.; Covic, A.; Balan, G.G.; Nistor, I.; Popa, C.; Hogas, S.; Covic, A. Gut Microbiota in Chronic Kidney Disease: From Composition to Modulation towards Better Outcomes—A Systematic Review. J. Clin. Med. 2023, 12, 1948. [Google Scholar] [CrossRef]
- Vernocchi, P.; Del Chierico, F.; Putignani, L. Gut Microbiota Metabolism and Interaction with Food Components. Int. J. Mol. Sci. 2020, 21, 3688. [Google Scholar] [CrossRef]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the gut microbiota in nutrition and health. BMJ 2018, 361, k2179. [Google Scholar] [CrossRef]
- Li, C.; Chen, X.; Yao, J.; Zha, W.; Li, M.; Shen, J.; Jiang, H.; Tian, P. Curcumin modulated gut microbiota and alleviated renal fibrosis in 5/6 nephrectomy-induced chronic kidney disease rats. PLoS ONE 2025, 20, e0314029. [Google Scholar] [CrossRef]
- Chen, Y.; Li, H.; Lai, F.; Min, T.; Wu, H.; Zhan, Q. The Influence and Mechanisms of Natural Plant Polysaccharides on Intestinal Microbiota-Mediated Metabolic Disorders. Foods 2024, 13, 3882. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Chang, C.I.; Hou, C.Y.; Chang-Chien, G.P.; Lin, S.; Hsu, C.N. Dietary Resveratrol Butyrate Monoester Supplement Improves Hypertension and Kidney Dysfunction in a Young Rat Chronic Kidney Disease Model. Nutrients 2023, 15, 635. [Google Scholar] [CrossRef] [PubMed]
- Mosterd, C.M.; Kanbay, M.; van den Born, B.J.H.; van Raalte, D.H.; Rampanelli, E. Intestinal microbiota and diabetic kidney diseases: The Role of microbiota and derived metabolites inmodulation of renal inflammation and disease progression. Best Pract. Res. Clin. Endocrinol. Metab. 2021, 35, 101484. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Xu, S.; Zhang, B.; Sun, X. Ramulus Mori (Sangzhi) Alkaloids Alleviate Diabetic Nephropathy through Improving Gut Microbiota Disorder. Nutrients 2024, 16, 2346. [Google Scholar] [CrossRef]
- Liu, J.; Tan, Y.; Cheng, H.; Zhang, D.; Feng, W.; Peng, C. Functions of Gut Microbiota Metabolites, Current Status and Future Perspectives. Aging Dis. 2022, 13, 1106–1126. [Google Scholar] [CrossRef]
- Liu, H.; Diep, T.N.; Wang, Y.; Wang, Y.; Yan, L.J. Diabetic Kidney Disease: Contribution of Phenyl Sulfate Derived from Dietary Tyrosine upon Gut Microbiota Catabolism. Biomolecules 2024, 14, 1153. [Google Scholar] [CrossRef]
- Otten, B.M.J.; Sthijns, M.; Troost, F.J. A Combination of Acetate, Propionate, and Butyrate Increases Glucose Uptake in C2C12 Myotubes. Nutrients 2023, 15, 946. [Google Scholar] [CrossRef]
- Du, Y.; He, C.; An, Y.; Huang, Y.; Zhang, H.; Fu, W.; Wang, M.; Shan, Z.; Xie, J.; Yang, Y.; et al. The Role of Short Chain Fatty Acids in Inflammation and Body Health. Int. J. Mol. Sci. 2024, 25, 7379. [Google Scholar] [CrossRef]
- Lee, C.C.; Kono, T.; Syed, F.; Weaver, S.A.; Sohn, P.; Wu, W.; Chang, G.; Liu, J.; Slak Rupnik, M.; Evans-Molina, C. Sodium butyrate prevents cytokine-induced beta-cell dysfunction through restoration of stromal interaction molecule 1 expression and activation of store-operated calcium entry. FASEB J. 2024, 38, e23853. [Google Scholar] [CrossRef]
- Peng, K.; Dong, W.; Luo, T.; Tang, H.; Zhu, W.; Huang, Y.; Yang, X. Butyrate and obesity: Current research status and future prospect. Front. Endocrinol. 2023, 14, 1098881. [Google Scholar] [CrossRef]
- Gao, Y.; Yao, Q.; Meng, L.; Wang, J.; Zheng, N. Double-side role of short chain fatty acids on host health via the gut-organ axes. Anim. Nutr. 2024, 18, 322–339. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Umar, S.; Rust, B.; Lazarova, D.; Bordonaro, M. Secondary Bile Acids and Short Chain Fatty Acids in the Colon: A Focus on Colonic Microbiome, Cell Proliferation, Inflammation, and Cancer. Int. J. Mol. Sci. 2019, 20, 1214. [Google Scholar] [CrossRef] [PubMed]
- Mayorga-Ramos, A.; Barba-Ostria, C.; Simancas-Racines, D.; Guaman, L.P. Protective role of butyrate in obesity and diabetes: New insights. Front. Nutr. 2022, 9, 1067647. [Google Scholar] [CrossRef]
- Cheng, X.; Zhou, T.; He, Y.; Xie, Y.; Xu, Y.; Huang, W. The role and mechanism of butyrate in the prevention and treatment of diabetic kidney disease. Front. Microbiol. 2022, 13, 961536. [Google Scholar] [CrossRef] [PubMed]
- Watchon, M.; Robinson, K.J.; Luu, L.; An, Y.; Yuan, K.C.; Plenderleith, S.K.; Cheng, F.; Don, E.K.; Nicholson, G.A.; Lee, A.; et al. Treatment with sodium butyrate induces autophagy resulting in therapeutic benefits for spinocerebellar ataxia type 3. FASEB J. 2024, 38, e23429. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, P.; Gamage, H.; Laird, A.S. Butyrate as a potential therapeutic agent for neurodegenerative disorders. Neurochem. Int. 2024, 176, 105745. [Google Scholar] [CrossRef]
- Pant, K.; Venugopal, S.K.; Lorenzo Pisarello, M.J.; Gradilone, S.A. The Role of Gut Microbiome-Derived Short-Chain Fatty Acid Butyrate in Hepatobiliary Diseases. Am. J. Pathol. 2023, 193, 1455–1467. [Google Scholar] [CrossRef]
- Mohamed Elfadil, O.; Mundi, M.S.; Abdelmagid, M.G.; Patel, A.; Patel, N.; Martindale, R. Butyrate: More Than a Short Chain Fatty Acid. Curr. Nutr. Rep. 2023, 12, 255–262. [Google Scholar] [CrossRef]
- Recharla, N.; Geesala, R.; Shi, X.Z. Gut Microbial Metabolite Butyrate and Its Therapeutic Role in Inflammatory Bowel Disease: A Literature Review. Nutrients 2023, 15, 2275. [Google Scholar] [CrossRef]
- Choy, W.H.; Adler, A.; Morgan-Lang, C.; Gough, E.K.; Hallam, S.J.; Manges, A.R.; Chew, B.H.; Penniston, K.; Miller, A.; Lange, D. Deficient butyrate metabolism in the intestinal microbiome is a potential risk factor for recurrent kidney stone disease. Urolithiasis 2024, 52, 38. [Google Scholar] [CrossRef]
- Huang, P.; Cao, J.; Chen, J.; Luo, Y.; Gong, X.; Wu, C.; Wang, Y. Crosstalk between gut microbiota and renal ischemia/reperfusion injury. Front. Cell. Infect. Microbiol. 2022, 12, 1015825. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Yang, X.; Jian, Y.; Liu, J.; Ke, X.; Chen, S.; Yang, D.; Yang, D. SIRT3 deficiency exacerbates early-stage fibrosis after ischaemia-reperfusion-induced AKI. Cell. Signal. 2022, 93, 110284. [Google Scholar] [CrossRef]
- Park, E.J.; Dusabimana, T.; Je, J.; Jeong, K.; Yun, S.P.; Kim, H.J.; Kim, H.; Park, S.W. Honokiol Protects the Kidney from Renal Ischemia and Reperfusion Injury by Upregulating the Glutathione Biosynthetic Enzymes. Biomedicines 2020, 8, 352. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hou, D.; Chen, X.; Zhu, J.; Zhang, R.; Sun, W.; Li, P.; Tian, Y.; Kong, X. Hydralazine protects against renal ischemia-reperfusion injury in rats. Eur. J. Pharmacol. 2019, 843, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Wu, M.; Zhou, P.; Huang, C.; Ye, C.; Wang, L. The long non-coding RNA MALAT1 is increased in renal ischemia-reperfusion injury and inhibits hypoxia-induced inflammation. Ren. Fail. 2018, 40, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Li, Q.; Liu, S.; An, X.; Huang, Z.; Zhang, B.; Yuan, Y.; Xing, C. Protective effect of hyperoside against renal ischemia-reperfusion injury via modulating mitochondrial fission, oxidative stress, and apoptosis. Free Radic Res. 2019, 53, 727–736. [Google Scholar] [CrossRef]
- Zhu, Y.B.; Zhang, Y.P.; Zhang, J.; Zhang, Y.B. Evaluation of Vitamin C Supplementation on Kidney Function and Vascular Reactivity Following Renal Ischemic Injury in Mice. Kidney Blood Press. Res. 2016, 41, 460–470. [Google Scholar] [CrossRef]
- Zhuang, S.; Lu, B.; Daubert, R.A.; Chavin, K.D.; Wang, L.; Schnellmann, R.G. Suramin promotes recovery from renal ischemia/reperfusion injury in mice. Kidney Int. 2009, 75, 304–311. [Google Scholar] [CrossRef]
- Silver, S.A.; Cardinal, H.; Colwell, K.; Burger, D.; Dickhout, J.G. Acute kidney injury: Preclinical innovations, challenges, and opportunities for translation. Can. J. Kidney Health Dis. 2015, 2, 30. [Google Scholar] [CrossRef]
- Huang, R.; Zhang, C.; Xiang, Z.; Lin, T.; Ling, J.; Hu, H. Role of mitochondria in renal ischemia-reperfusion injury. FEBS J. 2024, 291, 5365–5378. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhang, Z.; Zhang, N. Protective Effects of Butyrate on Renal Ischemia-Reperfusion Injury in Rats. Urol. Int. 2019, 102, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhou, C.; Chen, Y.; He, X.; Gao, F.; Xue, D. Quantitative increase in short-chain fatty acids, especially butyrate protects kidney from ischemia/reperfusion injury. J. Investig. Med. 2022, 70, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Ma, X. Gut microbial sodium butyrate alleviates renal ischemia-reperfusion injury by regulating HES1/PPARalpha. Mol. Immunol. 2022, 150, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, M.O.; Ahmed, M.M.; Arshad, S.; Javaid, U.; Khan, I.A.; Manzoor, M.; Andleeb, S.; Riaz, R.; Munawar, S.H.; Manzoor, Z.; et al. Nephroprotective Effects of Alhagi camelorum against Cisplatin-Induced Nephrotoxicity in Albino Wistar Rats. Molecules 2022, 27, 941. [Google Scholar] [CrossRef] [PubMed]
- Iskander, A.; Yan, L.J. Cisplatin-Induced Kidney Toxicity: Potential Roles of Major NAD(+)-Dependent Enzymes and Plant-Derived Natural Products. Biomolecules 2022, 12, 1078. [Google Scholar] [CrossRef]
- Chen, W.J.; Chen, Y.T.; Ko, J.L.; Chen, J.Y.; Zheng, J.Y.; Liao, J.W.; Ou, C.C. Butyrate modulates gut microbiota and anti-inflammatory response in attenuating cisplatin-induced kidney injury. Biomed. Pharmacother. 2024, 181, 117689. [Google Scholar] [CrossRef]
- Favero, C.; Pintor-Chocano, A.; Sanz, A.; Ortiz, A.; Sanchez-Nino, M.D. Butyrate promotes kidney resilience through a coordinated kidney protective response in tubular cells. Biochem. Pharmacol. 2024, 224, 116203. [Google Scholar] [CrossRef]
- Yan, L.J. Folic acid-induced animal model of kidney disease. Anim. Model. Exp. Med. 2021, 4, 329–342. [Google Scholar] [CrossRef]
- Johnson, B.E.; Haritha, C.V.; Mathesh, K.; Vamadevan, B.; Sharma, A.; Aneesha, V.A.; Jadhav, S.E.; Parida, S.; Singh, T.U.; Lingaraju, M.C. Weekly administration of betulinic acid prevents development of chronic renal failure from acute renal failure in folic acid-induced mouse model of kidney injury. Naunyn Schmiedebergs Arch. Pharmacol. 2025. [Google Scholar] [CrossRef]
- Santos, S.; Bosch, R.J.; Ortega, A.; Largo, R.; Fernandez-Agullo, T.; Gazapo, R.; Egido, J.; Esbrit, P. Up-regulation of parathyroid hormone-related protein in folic acid-induced acute renal failure. Kidney Int. 2001, 60, 982–995. [Google Scholar] [CrossRef]
- Schubert, G.E. Folic acid-induced acute renal failure in the the rat: Morphological studies. Kidney Int. Suppl. 1976, 6, S46–S50. [Google Scholar] [PubMed]
- Corte-Iglesias, V.; Saiz, M.L.; Andrade-Lopez, A.C.; Salazar, N.; Bernet, C.R.; Martin-Martin, C.; Borra, J.M.; Lozano, J.J.; Aransay, A.M.; Diaz-Corte, C.; et al. Propionate and butyrate counteract renal damage and progression to chronic kidney disease. Nephrol. Dial. Transplant. 2024, 40, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Song, Y.; Zhao, M.; Yi, Z.; Zeng, Q. Protective effects of edaravone, a free radical scavenger, on lipopolysaccharide-induced acute kidney injury in a rat model of sepsis. Int. Urol. Nephrol. 2015, 47, 1745–1752. [Google Scholar] [CrossRef]
- Peerapornratana, S.; Manrique-Caballero, C.L.; Gomez, H.; Kellum, J.A. Acute kidney injury from sepsis: Current concepts, epidemiology, pathophysiology, prevention and treatment. Kidney Int. 2019, 96, 1083–1099. [Google Scholar] [CrossRef] [PubMed]
- Doi, K.; Leelahavanichkul, A.; Yuen, P.S.; Star, R.A. Animal models of sepsis and sepsis-induced kidney injury. J. Clin. Investig. 2009, 119, 2868–2878. [Google Scholar] [CrossRef]
- Dou, X.; Yan, D.; Ma, Z.; Gao, N.; Shan, A. Sodium butyrate alleviates LPS-induced kidney injury via inhibiting TLR2/4 to regulate rBD2 expression. J. Food Biochem. 2022, 46, e14126. [Google Scholar] [CrossRef]
- Tian, X.; Yuan, L.; Zeng, Y. Butyrate attenuates SA-AKI by inhibiting pyroptosis via the STING-GSDMD axis. Biochem. Biophys. Res. Commun. 2025, 743, 151143. [Google Scholar] [CrossRef]
- Khan, M.; Farooqi, S.; Mitchell, K.L.; Chowdhury, S.K.R.; Cabrera-Ayala, M.; Huang, J.; Wallace, D.C.; Weiss, S.L. Effect of sodium butyrate on kidney and liver mitochondrial dysfunction in a lipopolysaccharide mouse model. FASEB J. 2024, 38, e70228. [Google Scholar] [CrossRef]
- Banu, K.; Lin, Q.; Basgen, J.M.; Planoutene, M.; Wei, C.; Reghuvaran, A.C.; Tian, X.; Shi, H.; Garzon, F.; Garzia, A.; et al. AMPK mediates regulation of glomerular volume and podocyte survival. JCI Insight 2021, 6, e150004. [Google Scholar] [CrossRef]
- Han, B.; Zhang, X.; Wang, L.; Yuan, W. Dysbiosis of Gut Microbiota Contributes to Uremic Cardiomyopathy via Induction of IFNgamma-Producing CD4(+) T Cells Expansion. Microbiol. Spectr. 2023, 11, e0310122. [Google Scholar] [CrossRef]
- Lauriola, M.; Farre, R.; Dejongh, S.; de Loor, H.; Evenepoel, P.; Masereeuw, R.; Zadora, W.; Meijers, B. Dietary protein intake and the tubular handling of indoxyl sulfate. Nephrol. Dial. Transplant. 2024, gfae220. [Google Scholar] [CrossRef]
- Shobeiri, N.; Adams, M.A.; Holden, R.M. Vascular calcification in animal models of CKD: A review. Am. J. Nephrol. 2010, 31, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.; Krieg, R.; Massey, H.D.; Carl, D.; Ghosh, S.; Gehr, T.W.B.; Ghosh, S.S. Sodium butyrate ameliorates insulin resistance and renal failure in CKD rats by modulating intestinal permeability and mucin expression. Nephrol. Dial. Transplant. 2019, 34, 783–794. [Google Scholar] [CrossRef]
- Wei, P.Z.; Szeto, C.C. Mitochondrial dysfunction in diabetic kidney disease. Clin. Chim. Acta 2019, 496, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Lee, E.Y. Excessively Enlarged Mitochondria in the Kidneys of Diabetic Nephropathy. Antioxidants 2021, 10, 741. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, Y.; Wang, Y.; Yan, L.-J. Rodent Models of Streptozotocin-Induced Diabetes as Suitable Paradigms for Studying Diabetic Kidney Disease. Free Radic. Antioxid. 2024, 14, 32–33. [Google Scholar] [CrossRef]
- Zhou, T.; Xu, H.; Cheng, X.; He, Y.; Ren, Q.; Li, D.; Xie, Y.; Gao, C.; Zhang, Y.; Sun, X.; et al. Sodium Butyrate Attenuates Diabetic Kidney Disease Partially via Histone Butyrylation Modification. Mediators Inflamm. 2022, 2022, 7643322. [Google Scholar] [CrossRef]
- Du, Y.; Tang, G.; Yuan, W. Suppression of HDAC2 by sodium butyrate alleviates apoptosis of kidney cells in db/db mice and HG-induced NRK-52E cells. Int. J. Mol. Med. 2020, 45, 210–222. [Google Scholar] [CrossRef]
- Dong, W.; Jia, Y.; Liu, X.; Zhang, H.; Li, T.; Huang, W.; Chen, X.; Wang, F.; Sun, W.; Wu, H. Sodium butyrate activates NRF2 to ameliorate diabetic nephropathy possibly via inhibition of HDAC. J. Endocrinol. 2017, 232, 71–83. [Google Scholar] [CrossRef]
- Gu, J.; Huang, W.; Zhang, W.; Zhao, T.; Gao, C.; Gan, W.; Rao, M.; Chen, Q.; Guo, M.; Xu, Y.; et al. Sodium butyrate alleviates high-glucose-induced renal glomerular endothelial cells damage via inhibiting pyroptosis. Int. Immunopharmacol. 2019, 75, 105832. [Google Scholar] [CrossRef]
- Du, Y.; Yang, Y.T.; Tang, G.; Jia, J.S.; Zhu, N.; Yuan, W.J. Butyrate alleviates diabetic kidney disease by mediating the miR-7a-5p/P311/TGF-beta1 pathway. FASEB J. 2020, 34, 10462–10475. [Google Scholar] [CrossRef] [PubMed]
- Ye, K.; Zhao, Y.; Huang, W.; Zhu, Y. Sodium butyrate improves renal injury in diabetic nephropathy through AMPK/SIRT1/PGC-1alpha signaling pathway. Sci. Rep. 2024, 14, 17867. [Google Scholar] [CrossRef]
- Yu, Y.; Jia, Y.Y.; Li, H.J. Sodium butyrate improves mitochondrial function and kidney tissue injury in diabetic kidney disease via the AMPK/PGC-1alpha pathway. Ren. Fail. 2023, 45, 2287129. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Xi, Y.; Yan, M.; Sun, C.; Tang, J.; Dong, X.; Yang, Z.; Wu, L. Lactiplantibacillus plantarum NKK20 Increases Intestinal Butyrate Production and Inhibits Type 2 Diabetic Kidney Injury through PI3K/Akt Pathway. J. Diabetes Res. 2023, 2023, 8810106. [Google Scholar] [CrossRef]
- Avgoustou, E.; Tzivaki, I.; Diamantopoulou, G.; Zachariadou, T.; Avramidou, D.; Dalopoulos, V.; Skourtis, A. Obesity-Related Chronic Kidney Disease: From Diagnosis to Treatment. Diagnostics 2025, 15, 169. [Google Scholar] [CrossRef]
- Jakhotia, S.; Kavvuri, R.; Raviraj, S.; Baishya, S.; Pasupulati, A.K.; Reddy, G.B. Obesity-related glomerulopathy is associated with elevated WT1 expression in podocytes. Int. J. Obes. 2024, 48, 1080–1091. [Google Scholar] [CrossRef]
- Lei, L.; Zhu, T.; Cui, T.J.; Liu, Y.; Hocher, J.G.; Chen, X.; Zhang, X.M.; Cai, K.W.; Deng, Z.Y.; Wang, X.H.; et al. Renoprotective effects of empagliflozin in high-fat diet-induced obesity-related glomerulopathy by regulation of gut-kidney axis. Am. J. Physiol. Cell. Physiol. 2024, 327, C994–C1011. [Google Scholar] [CrossRef]
- Shi, Y.; Xing, L.; Zheng, R.; Luo, X.; Yue, F.; Xiang, X.; Qiu, A.; Xie, J.; Russell, R.; Zhang, D. Butyrate attenuates high-fat diet-induced glomerulopathy through GPR43-Sirt3 pathway. Br. J. Nutr. 2024, 133, 1–10. [Google Scholar] [CrossRef]
- Liu, X.; Huang, S.; Wang, F.; Zheng, L.; Lu, J.; Chen, J.; Li, S. Huangqi-Danshen Decoction Ameliorates Adenine-Induced Chronic Kidney Disease by Modulating Mitochondrial Dynamics. Evid. Based Complement Altern. Med. 2019, 2019, 9574045. [Google Scholar] [CrossRef]
- Diwan, V.; Brown, L.; Gobe, G.C. Adenine-induced chronic kidney disease in rats. Nephrology 2018, 23, 5–11. [Google Scholar] [CrossRef]
- Tian, X.; Zeng, Y.; Tu, Q.; Jiao, Y.; Yao, S.; Chen, Y.; Sun, L.; Xia, Q.; Luo, Y.; Yuan, L.; et al. Butyrate alleviates renal fibrosis in CKD by regulating NLRP3-mediated pyroptosis via the STING/NF-kappaB/p65 pathway. Int. Immunopharmacol. 2023, 124, 111010. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Jose, A.; Alonzo-Palma, N.; Malik, T.; Shankaranarayanan, D.; Regunathan-Shenk, R.; Raj, D.S. Butyrate producing microbiota are reduced in chronic kidney diseases. Sci. Rep. 2021, 11, 23530. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.N.; Hsu, C.N.; Hou, C.Y.; Chen, S.Y.; Tain, Y.L. Resveratrol Butyrate Esters Reduce Hypertension in a Juvenile Rat Model of Chronic Kidney Disease Exacerbated by Microplastics. Nutrients 2024, 16, 4076. [Google Scholar] [CrossRef] [PubMed]
- Nagata, D.; Hishida, E. Elucidating the complex interplay between chronic kidney disease and hypertension. Hypertens. Res. 2024, 47, 3409–3422. [Google Scholar] [CrossRef]
- Ameer, O.Z. Hypertension in chronic kidney disease: What lies behind the scene. Front. Pharmacol. 2022, 13, 949260. [Google Scholar] [CrossRef]
- Qian, Q. Salt, water and nephron: Mechanisms of action and link to hypertension and chronic kidney disease. Nephrology 2018, 23 (Suppl. S4), 44–49. [Google Scholar] [CrossRef]
- Bovee, D.M.; Cuevas, C.A.; Zietse, R.; Danser, A.H.J.; Mirabito Colafella, K.M.; Hoorn, E.J. Salt-sensitive hypertension in chronic kidney disease: Distal tubular mechanisms. Am. J. Physiol. Ren. Physiol. 2020, 319, F729–F745. [Google Scholar] [CrossRef]
- Carresi, C.; Cardamone, A.; Coppoletta, A.R.; Caminiti, R.; Macri, R.; Lorenzo, F.; Scarano, F.; Mollace, R.; Guarnieri, L.; Ruga, S.; et al. The protective effect of Bergamot Polyphenolic Fraction on reno-cardiac damage induced by DOCA-salt and unilateral renal artery ligation in rats. Biomed. Pharmacother. 2024, 171, 116082. [Google Scholar] [CrossRef]
- Efentakis, P.; Symeonidi, L.; Gianniou, D.D.; Mikropoulou, E.V.; Giardoglou, P.; Valakos, D.; Vatsellas, G.; Tsota, M.; Kostomitsopoulos, N.; Smyrnioudis, I.; et al. Antihypertensive Potential of Pistacia lentiscus var. Chia: Molecular Insights and Therapeutic Implications. Nutrients 2024, 16, 2152. [Google Scholar] [CrossRef]
- Wu, C.; Chen, Z.; Zhang, L.; Zhu, Y.; Deng, M.; Huang, C.; Liu, Y.; Zhu, Q.; Wang, L. Sodium butyrate ameliorates deoxycorticosterone acetate/salt-induced hypertension and renal damage by inhibiting the MR/SGK1 pathway. Hypertens. Res. 2021, 44, 168–178. [Google Scholar] [CrossRef]
- Mohamad, R.H.; El-Bastawesy, A.M.; Zekry, Z.K.; Al-Mehdar, H.A.; Al-Said, M.G.; Aly, S.S.; Sharawy, S.M.; El-Merzabani, M.M. The role of Curcuma longa against doxorubicin (adriamycin)-induced toxicity in rats. J. Med. Food 2009, 12, 394–402. [Google Scholar] [CrossRef]
- Zhang, Q.; Wu, G.; Guo, S.; Liu, Y.; Liu, Z. Effects of tristetraprolin on doxorubicin (adriamycin)-induced experimental kidney injury through inhibiting IL-13/STAT6 signal pathway. Am. J. Transl. Res. 2020, 12, 1203–1221. [Google Scholar] [PubMed]
- Li, J.; Wang, P.Y.; Long, N.A.; Zhuang, J.; Springer, D.A.; Zou, J.; Lin, Y.; Bleck, C.K.E.; Park, J.H.; Kang, J.G.; et al. p53 prevents doxorubicin cardiotoxicity independently of its prototypical tumor suppressor activities. Proc. Natl. Acad. Sci. USA 2019, 116, 19626–19634. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, C.; Santos, R.X.; Cardoso, S.; Correia, S.; Oliveira, P.J.; Santos, M.S.; Moreira, P.I. Doxorubicin: The good, the bad and the ugly effect. Curr. Med. Chem. 2009, 16, 3267–3285. [Google Scholar] [CrossRef] [PubMed]
- Mahzari, S.; Hosseinian, S.; Hadjzadeh, M.A.; Mohebbati, R.; Noshahr, Z.S.; Rad, A.K. Kidney dysfunction and oxidative stress in doxorubicin-induced nephrotic rat: Protective role of sesame oil. Saudi J. Kidney Dis. Transpl. 2021, 32, 1243–1252. [Google Scholar] [CrossRef]
- Avagimyan, A.; Pogosova, N.; Kakturskiy, L.; Sheibani, M.; Challa, A.; Kogan, E.; Fogacci, F.; Mikhaleva, L.; Vandysheva, R.; Yakubovskaya, M.; et al. Doxorubicin-related cardiotoxicity: Review of fundamental pathways of cardiovascular system injury. Cardiovasc. Pathol. 2024, 73, 107683. [Google Scholar] [CrossRef]
- Zhou, Y.F.; Fu, Y.; Lai, Z.Q.; Xu, H.L.; Shen, N.; Long, J.; Zhang, H.; Dong, Y.F. Heat shock protein 22 alleviates doxorubicin-induced kidney injury by suppressing oxidative stress and apoptosis. Sci. Rep. 2024, 14, 23527. [Google Scholar] [CrossRef]
- Suleimani, Y.A.; Maskari, R.A.; Ali, B.H.; Ali, H.; Manoj, P.; Al-Khamiyasi, A.; Abdelrahman, A.M. Nephroprotective effects of diminazene on doxorubicin-induced acute kidney injury in rats. Toxicol. Rep. 2023, 11, 460–468. [Google Scholar] [CrossRef]
- Sanajou, D.; Nazari Soltan Ahmad, S.; Hosseini, V.; Kalantary-Charvadeh, A.; Marandi, Y.; Roshangar, L.; Bahrambeigi, S.; Mesgari-Abbasi, M. β-Lapachone protects against doxorubicin-induced nephrotoxicity via NAD+/AMPK/NF-κB in mice. Naunyn Schmiedebergs Arch. Pharmacol. 2019, 392, 633–640. [Google Scholar] [CrossRef]
- Felizardo, R.J.F.; de Almeida, D.C.; Pereira, R.L.; Watanabe, I.K.M.; Doimo, N.T.S.; Ribeiro, W.R.; Cenedeze, M.A.; Hiyane, M.I.; Amano, M.T.; Braga, T.T.; et al. Gut microbial metabolite butyrate protects against proteinuric kidney disease through epigenetic- and GPR109a-mediated mechanisms. FASEB J. 2019, 33, 11894–11908. [Google Scholar] [CrossRef]
- Colclough-Davy, R.; Eaton, A.; Perry, M.J. Evaluating Kidney Function in Patients Receiving Contrast Media. Radiol. Technol. 2022, 94, 124–128. [Google Scholar] [PubMed]
- Hisamune, R.; Yamakawa, K.; Umemura, Y.; Ushio, N.; Mochizuki, K.; Inokuchi, R.; Doi, K.; Takasu, A. Association Between IV Contrast Media Exposure and Acute Kidney Injury in Patients Requiring Emergency Admission: A Nationwide Observational Study in Japan. Crit. Care Explor. 2024, 6, e1142. [Google Scholar] [CrossRef] [PubMed]
- Caglic, I.; Sushentsev, N.; Colarieti, A.; Warren, A.Y.; Shah, N.; Lamb, B.W.; Barrett, T. Value of the capsular enhancement sign on dynamic contrast-enhanced prostate multiparametric MRI for the detection of extracapsular extension. Eur. J. Radiol. 2022, 150, 110275. [Google Scholar] [CrossRef] [PubMed]
- Kusirisin, P.; Apaijai, N.; Noppakun, K.; Kuanprasert, S.; Chattipakorn, S.C.; Chattipakorn, N. Circulating mitochondrial dysfunction as an early biomarker for contrast media-induced acute kidney injury in chronic kidney disease patients. J. Cell. Mol. Med. 2023, 27, 2059–2070. [Google Scholar] [CrossRef]
- Andreucci, M.; Faga, T.; Pisani, A.; Sabbatini, M.; Michael, A. Acute kidney injury by radiographic contrast media: Pathogenesis and prevention. BioMed Res. Int. 2014, 2014, 362725. [Google Scholar] [CrossRef]
- Seeliger, E.; Lenhard, D.C.; Persson, P.B. Contrast media viscosity versus osmolality in kidney injury: Lessons from animal studies. BioMed Res. Int. 2014, 2014, 358136. [Google Scholar] [CrossRef]
- Jiang, Z.; Zhang, J.; Lu, Y. Protective Effects and Mechanisms of Rosuvastatin on Acute Kidney Injury Induced by Contrast Media in Rats. Int. J. Nephrol. 2020, 2020, 3490641. [Google Scholar] [CrossRef]
- Naziroglu, M.; Yoldas, N.; Uzgur, E.N.; Kayan, M. Role of contrast media on oxidative stress, Ca2+ signaling and apoptosis in kidney. J. Membr. Biol. 2013, 246, 91–100. [Google Scholar] [CrossRef]
- Machado, R.A.; Constantino Lde, S.; Tomasi, C.D.; Rojas, H.A.; Vuolo, F.S.; Vitto, M.F.; Cesconetto, P.A.; de Souza, C.T.; Ritter, C.; Dal-Pizzol, F. Sodium butyrate decreases the activation of NF-kappaB reducing inflammation and oxidative damage in the kidney of rats subjected to contrast-induced nephropathy. Nephrol. Dial. Transplant. 2012, 27, 3136–3140. [Google Scholar] [CrossRef]
- Yan, L.J.; Orr, W.C.; Sohal, R.S. Identification of oxidized proteins based on sodium dodecyl sulfate-polyacrylamide gel electrophoresis, immunochemical detection, isoelectric focusing, and microsequencing. Anal. Biochem. 1998, 263, 67–71. [Google Scholar] [CrossRef]
- Yan, L.J.; Lodge, J.K.; Traber, M.G.; Packer, L. Apolipoprotein B carbonyl formation is enhanced by lipid peroxidation during copper-mediated oxidation of human low-density lipoproteins. Arch. Biochem. Biophys. 1997, 339, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.J.; Lodge, J.K.; Traber, M.G.; Matsugo, S.; Packer, L. Comparison between copper-mediated and hypochlorite-mediated modifications of human low density lipoproteins evaluated by protein carbonyl formation. J. Lipid Res. 1997, 38, 992–1001. [Google Scholar] [CrossRef]
- He, Y.; Zhu, X.; Song, H.; Liu, Y.; Cao, C. Sodium butyrate alleviates T-2 toxin-induced liver toxicity and renal toxicity in quails by modulating oxidative stress-related Nrf2 signaling pathway, inflammation, and CYP450 enzyme system. J. Food Sci. 2024, 89, 8036–8053. [Google Scholar] [CrossRef] [PubMed]
- Xiang, M.; Qiao, L.; Han, Q.; Zha, Y.; Sui, X.; Wang, Q. Effects of Supplementation With Different Specificities of Dietary Fiber on Health-Related Indicators in Adults With Overweight or Obesity: A Systematic Review and Meta-analysis of Randomized Controlled Trials. Nutr. Rev. 2025, nuae193. [Google Scholar] [CrossRef]
- Li, Y.; Han, M.; Song, J.; Liu, S.; Wang, Y.; Su, X.; Wei, K.; Xu, Z.; Li, H.; Wang, Z. The prebiotic effects of soluble dietary fiber mixture on renal anemia and the gut microbiota in end-stage renal disease patients on maintenance hemodialysis: A prospective, randomized, placebo-controlled study. J. Transl. Med. 2022, 20, 599. [Google Scholar] [CrossRef]
- Wong, J.M. Gut microbiota and cardiometabolic outcomes: Influence of dietary patterns and their associated components. Am J. Clin. Nutr. 2014, 100 (Suppl. S1), 369S–377S. [Google Scholar] [CrossRef]
- Wong, J.; Piceno, Y.M.; DeSantis, T.Z.; Pahl, M.; Andersen, G.L.; Vaziri, N.D. Expansion of urease- and uricase-containing, indole- and p-cresol-forming and contraction of short-chain fatty acid-producing intestinal microbiota in ESRD. Am. J. Nephrol. 2014, 39, 230–237. [Google Scholar] [CrossRef]
- Zuo, Z.; Pang, W.; Sun, W.; Lu, B.; Zou, L.; Zhang, D.; Wang, Y. Metallothionein-Kidney Bean Polyphenol Complexes Showed Antidiabetic Activity in Type 2 Diabetic Rats by Improving Insulin Resistance and Regulating Gut Microbiota. Foods 2023, 12, 3139. [Google Scholar] [CrossRef]
- Yang, Y.; Luo, J.; Wang, S.; Yang, D.; Chen, S.; Wang, Q.; Zhou, A. A water-soluble polysaccharide from finger citron ameliorates diabetes mellitus via gut microbiota-GLP-1 pathway in high-fat diet and streptozotocin-induced diabetic mice. Int. J. Biol. Macromol. 2025, 300, 140126. [Google Scholar] [CrossRef]
- Singh, I.; Anand, S.; Gowda, D.J.; Kamath, A.; Singh, A.K. Caloric restriction mimetics improve gut microbiota: A promising neurotherapeutics approach for managing age-related neurodegenerative disorders. Biogerontology 2024, 25, 899–922. [Google Scholar] [CrossRef]
- Estrela, G.R.; Wasinski, F.; Batista, R.O.; Hiyane, M.I.; Felizardo, R.J.; Cunha, F.; de Almeida, D.C.; Malheiros, D.M.; Camara, N.O.; Barros, C.C.; et al. Caloric Restriction Is More Efficient than Physical Exercise to Protect from Cisplatin Nephrotoxicity via PPAR-Alpha Activation. Front. Physiol. 2017, 8, 116. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.H.; Ao, Q.G.; Cheng, Q.L. Caloric restriction inhibits renal artery ageing by reducing endothelin-1 expression. Ann. Transl. Med. 2021, 9, 979. [Google Scholar] [CrossRef] [PubMed]
- Koutnik, A.P.; Klein, S.; Robinson, A.T.; Watso, J.C. Efficacy and Safety of Long-term Ketogenic Diet Therapy in a Patient With Type 1 Diabetes. JCEM Case Rep. 2024, 2, luae102. [Google Scholar] [CrossRef] [PubMed]
- Athinarayanan, S.J.; Roberts, C.G.P.; Vangala, C.; Shetty, G.K.; McKenzie, A.L.; Weimbs, T.; Volek, J.S. The case for a ketogenic diet in the management of kidney disease. BMJ Open Diabetes Res. Care 2024, 12, e004101. [Google Scholar] [CrossRef]
- Favero, C.; Ortiz, A.; Sanchez-Nino, M.D. Probiotics for kidney disease. Clin. Kidney J. 2022, 15, 1981–1986. [Google Scholar] [CrossRef]
- Ighodaro, I.; Eric, O.K.; Adebayo, O. Interactions of PPAR alpha and GLUT4 in DOCA/salt-induced renal injury in mice. Niger. J. Physiol. Sci. 2013, 28, 127–133. [Google Scholar]
- Ding, R.; Xiao, Z.; Jiang, Y.; Yang, Y.; Ji, Y.; Bao, X.; Xing, K.; Zhou, X.; Zhu, S. Calcitriol ameliorates damage in high-salt diet-induced hypertension: Evidence of communication with the gut-kidney axis. Exp. Biol. Med. 2022, 247, 624–640. [Google Scholar] [CrossRef]
- Tan, W.; Chen, Y.; An, P.; Wang, A.; Chu, M.; Shi, L.; Hou, X.; Tang, B.; Zhang, X.; Li, Z. Sodium butyrate-induced histone hyperacetylation up-regulating WT1 expression in porcine kidney fibroblasts. Biotechnol. Lett. 2015, 37, 1195–1202. [Google Scholar] [CrossRef]
- Cai, K.; Ma, Y.; Cai, F.; Huang, X.; Xiao, L.; Zhong, C.; Ren, P.; Luo, Q.; Chen, J.; Han, F. Changes of gut microbiota in diabetic nephropathy and its effect on the progression of kidney injury. Endocrine 2022, 76, 294–303. [Google Scholar] [CrossRef]
- Yan, L.J.; Allen, D.C. Cadmium-Induced Kidney Injury: Oxidative Damage as a Unifying Mechanism. Biomolecules 2021, 11, 1575. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Approach | Reference |
|---|---|
| Consuming more fruits and vegetables or specific dietary fiber | [117,118,119,120] |
| Ingesting natural products, e.g., polysaccharides and polyphenols | [121,122] |
| Caloric restriction/dietary restriction | [7,123,124,125] |
| Ketogenic diet | [126,127] |
| Taking prebiotics or probiotics | [118,128] |
| Cutting back on salt | [129,130] |
| Exogenous sodium butyrate | [131,132] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diep, T.N.; Liu, H.; Yan, L.-J. Beneficial Effects of Butyrate on Kidney Disease. Nutrients 2025, 17, 772. https://doi.org/10.3390/nu17050772
Diep TN, Liu H, Yan L-J. Beneficial Effects of Butyrate on Kidney Disease. Nutrients. 2025; 17(5):772. https://doi.org/10.3390/nu17050772
Chicago/Turabian StyleDiep, Tram N., Haoxin Liu, and Liang-Jun Yan. 2025. "Beneficial Effects of Butyrate on Kidney Disease" Nutrients 17, no. 5: 772. https://doi.org/10.3390/nu17050772
APA StyleDiep, T. N., Liu, H., & Yan, L.-J. (2025). Beneficial Effects of Butyrate on Kidney Disease. Nutrients, 17(5), 772. https://doi.org/10.3390/nu17050772