
Effect of Akkermansia muciniphila on GLP-1 and Insulin Secretion


Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Cell Preparation and Protein Quantification
2.1.1. Insulin Study
- Treatment Formulation
- Cell Line Preparation, Culture Conditions, and Insulin Measurement
2.1.2. GLP-1 Study
- Treatment Formulation
- Cell Line Preparation, Culture Conditions, and GLP-1 Measurement
2.2. Data Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yadav, D.; Mishra, M.; Tiwari, A.; Bisen, P.S.; Goswamy, H.M.; Prasad, G.B.K.S. Prevalence of Dyslipidemia and Hypertension in Indian Type 2 Diabetic Patients with Metabolic Syndrome and its Clinical Significance. Osong Public Health Res. Perspect. 2014, 5, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.S.; Hossain, K.S.; Das, S.; Kundu, S.; Adegoke, E.O.; Rahman, M.A.; Hannan, M.A.; Uddin, M.J.; Pang, M.G. Role of Insulin in Health and Disease: An Update. Int. J. Mol. Sci. 2021, 22, 6403. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Aronoff, S.L.; Berkowitz, K.; Shreiner, B.; Want, L. Glucose Metabolism and Regulation: Beyond Insulin and Glucagon. Diabetes Spectr. 2004, 17, 183–190. [Google Scholar] [CrossRef]
- Nakrani, M.N.; Wineland, R.H.; Anjum, F. Physiology, Glucose Metabolism. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2025. Available online: https://www.ncbi.nlm.nih.gov/books/NBK560599/ (accessed on 14 May 2025).
- Zheng, Z.; Zong, Y.; Ma, Y.; Tian, Y.; Pang, Y.; Zhang, C.; Gao, J. Glucagon-like peptide-1 receptor: Mechanisms and advances in therapy. Sig. Transduct. Target. Ther. 2024, 9, 234. [Google Scholar] [CrossRef] [PubMed]
- Kansakar, U.; Nieves Garcia, C.; Santulli, G.; Gambardella, J.; Mone, P.; Jankauskas, S.S.; Lombardi, A. Exogenous Ketones in Cardiovascular Disease and Diabetes: From Bench to Bedside. J. Clin. Med. 2024, 13, 7391. [Google Scholar] [CrossRef] [PubMed]
- Mone, P.; Lombardi, A.; Kansakar, U.; Varzideh, F.; Jankauskas, S.S.; Pansini, A.; Marzocco, S.; De Gennaro, S.; Famiglietti, M.; Macina, G. Empagliflozin Improves the MicroRNA Signature of Endothelial Dysfunction in Patients with Heart Failure with Preserved Ejection Fraction and Diabetes. J. Pharmacol. Exp. Ther. 2023, 384, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Pellegrino, A.; Coppola, G.; Santopaolo, F.; Gasbarrini, A.; Ponziani, F.R. Role of Akkermansia in Human Diseases: From Causation to Therapeutic Properties. Nutrients 2023, 15, 1815. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Li, Z.; Hu, G.; Zhu, L.; Sun, Z.; Jiang, Y.; Gao, M.J.; Zhan, X. Study of growth, metabolism, and morphology of Akkermansia muciniphila with an in vitro advanced bionic intestinal reactor. BMC Microbiol. 2021, 21, 61. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Creely, S.J.; McTernan, P.G.; Kusminski, C.M.; Fisher, M.; Da Silva, N.F.; Khanolkar, M.; Evans, M.; Harte, A.L.; Kumar, S. Lipopolysaccharide activates an innate immune system response in human adipose tissue in obesity and type 2 diabetes. Am. J. Physiol.-Endocrinol. Metab. 2007, 292, E740–E747. [Google Scholar] [CrossRef] [PubMed]
- Khondkaryan, L.; Margaryan, S.; Poghosyan, D.; Manukyan, G. Impaired Inflammatory Response to LPS in Type 2 Diabetes Mellitus. Int. J. Inflam. 2018, 2018, 2157434. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Fu, Y.J.; Xu, B.; Huang, S.W.; Luo, X.; Deng, X.L.; Luo, S.; Liu, C.; Wang, Q.; Chen, J.Y.; Zhou, L. Baicalin prevents LPS-induced activation of TLR4/NF-κB p65 pathway and inflammation in mice via inhibiting the expression of CD14. Acta Pharmacol. Sin. 2021, 42, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Jinjie, L.; Ge, Y.; Qihe, Z.; Zhuo, L.; Xin, J.; Ying, X. Function of Akkermansia muciniphila in type 2 diabetes and related diseases. Frontiers in Microbiology. Front. Microbiol. 2023, 14, 1172400. [Google Scholar] [CrossRef]
- Drucker, D.J. Mechanisms of Action and Therapeutic Application of Glucagon-like Peptide-1. Cell Metab. 2018, 27, 740–756. [Google Scholar] [CrossRef] [PubMed]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: A proof-of-concept exploratory study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Li, Q.; Cheng, L.; Buch, H.; Zhang, F. Akkermansia muciniphila is a promising probiotic. Microb. Biotechnol. 2019, 12, 1109–1125. [Google Scholar] [CrossRef] [PubMed]
- Plovier, H.; Everard, A.; Druart, C.; Depommier, C.; Van Hul, M.; Geurts, L.; Chilloux, J.; Ottman, N.; Duparc, T.; Lichtenstein, L. A purified membrane protein from Akkermansia muciniphila or the pasteurized bacterium improves metabolism in obese and diabetic mice. Nat. Med. 2017, 23, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.S.; Cho, C.H.; Yun, M.S.; Jang, S.J.; You, H.J.; Kim, J.H.; Han, D.; Cha, K.H.; Moon, S.H.; Lee, K.; et al. Akkermansia muciniphila secretes a glucagon-like peptide-1-inducing protein that improves glucose homeostasis and ameliorates metabolic disease in mice. Nat. Microbiol. 2021, 6, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Chelakkot, C.; Choi, Y.; Kim, D.K.; Park, H.T.; Ghim, J.; Kwon, Y.; Jeon, J.; Kim, M.S.; Jee, Y.K.; Gho, Y.S. Akkermansia muciniphila-derived extracellular vesicles influence gut permeability through the regulation of tight junctions. Exp. Mol. Med. 2018, 50, e450. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; de Vos, W.M. Next-generation beneficial microbes: The case of Akkermansia muciniphila. Nat. Rev. Microbiol. 2017, 8, 1765. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
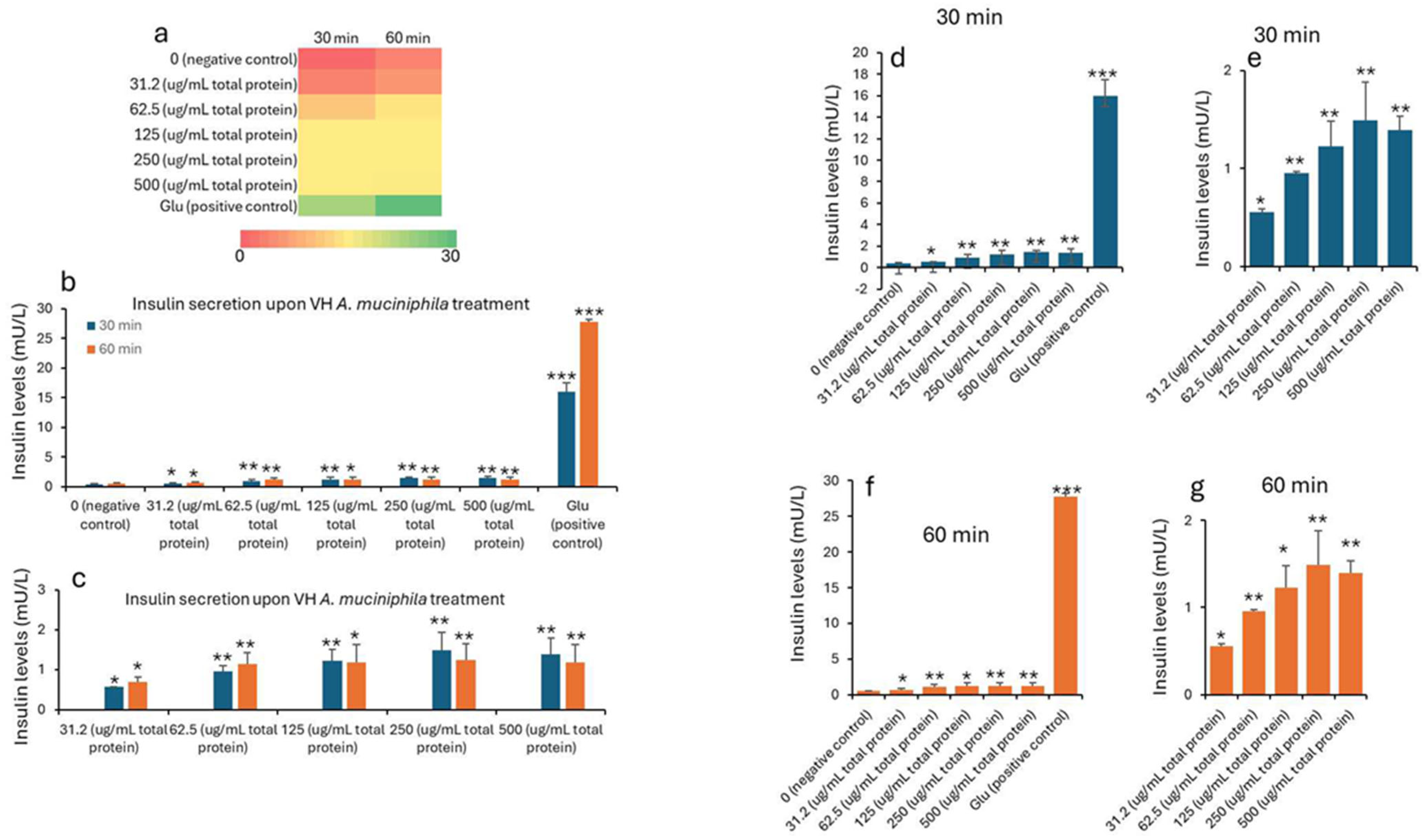
| Treatment (µg/mL) * | 0 min (Baseline) | 30 min | 60 min | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | SEM | Mean | SD | SEM | p Values | Mean | SD | SEM | p Values | |
| 0 (negative control) | 0.25 | 0.20 | 0.07 | 0.41 | 0.03 | 0.01 | 0.57 | 0.01 | 0.00 | ||
| 31.2 (μg/mL total protein) | 0.18 | 0.06 | 0.02 | 0.56 | 0.02 | 0.01 | 0.027 | 0.68 | 0.14 | 0.05 | 0.0182 |
| 62.5 (μg/mL total protein) | 0.95 | 0.25 | 0.08 | 0.009 | 1.14 | 0.29 | 0.10 | 0.0057 | |||
| 125 (μg/mL total protein) | 1.23 | 0.39 | 0.13 | 0.008 | 1.18 | 0.45 | 0.15 | 0.0158 | |||
| 250 (μg/mL total protein) | 1.49 | 0.14 | 0.05 | 0.0004 | 1.25 | 0.40 | 0.13 | 0.0092 | |||
| 500 (μg/mL total protein) | 1.39 | 0.35 | 0.12 | 0.0039 | 1.17 | 0.45 | 0.15 | 0.0153 | |||
| Glu (positive control) | 15.9 | 1.51 | 0.50 | 2.8 × 10−5 | 27.7 | 0.45 | 0.15 | 4.08 × 10−7 | |||
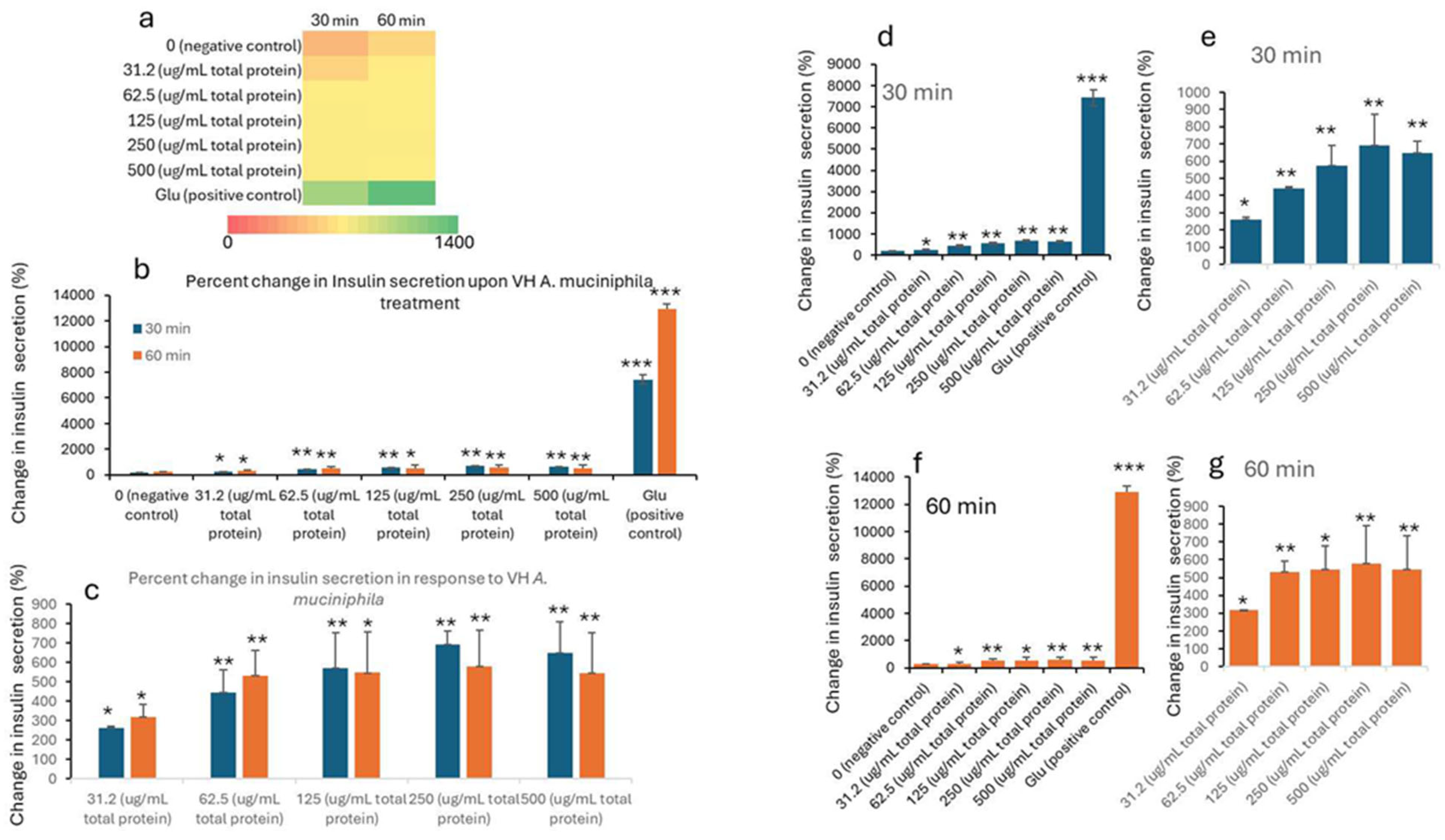
| Treatment (µg/mL) * | 0 min (Baseline) | 30 min | 60 min | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | SEM | Mean | SD | SEM | p Values | Mean | SD | SEM | p Values | |
| 0 (negative control) | 117.42 | 91.57 | 30.52 | 191.34 | 13.52 | 4.51 | 263.71 | 2.48 | 0.83 | ||
| 31.2 (μg/mL total protein) | 82.58 | 27.84 | 9.28 | 260.39 | 8.51 | 2.84 | 0.027 | 317.28 | 64.06 | 21.35 | 0.0182 |
| 62.5 (μg/mL total protein) | 443.78 | 116.81 | 38.94 | 0.009 | 530.95 | 132.97 | 44.32 | 0.0057 | |||
| 125 (μg/mL total protein) | 572.27 | 180.74 | 60.25 | 0.008 | 546.95 | 210.22 | 70.07 | 0.0158 | |||
| 250 (μg/mL total protein) | 693.30 | 67.18 | 22.39 | 0.0004 | 579.84 | 187.53 | 62.51 | 0.0092 | |||
| 500 (μg/mL total protein) | 647.64 | 162.44 | 54.15 | 0.0039 | 546.33 | 207.60 | 69.20 | 0.0153 | |||
| Glu (positive control) | 7434.4 | 701.2 | 233.7 | 2.8 × 10−5 | 12,898.6 | 415.49 | 138.5 | 4.08 × 10−7 | |||
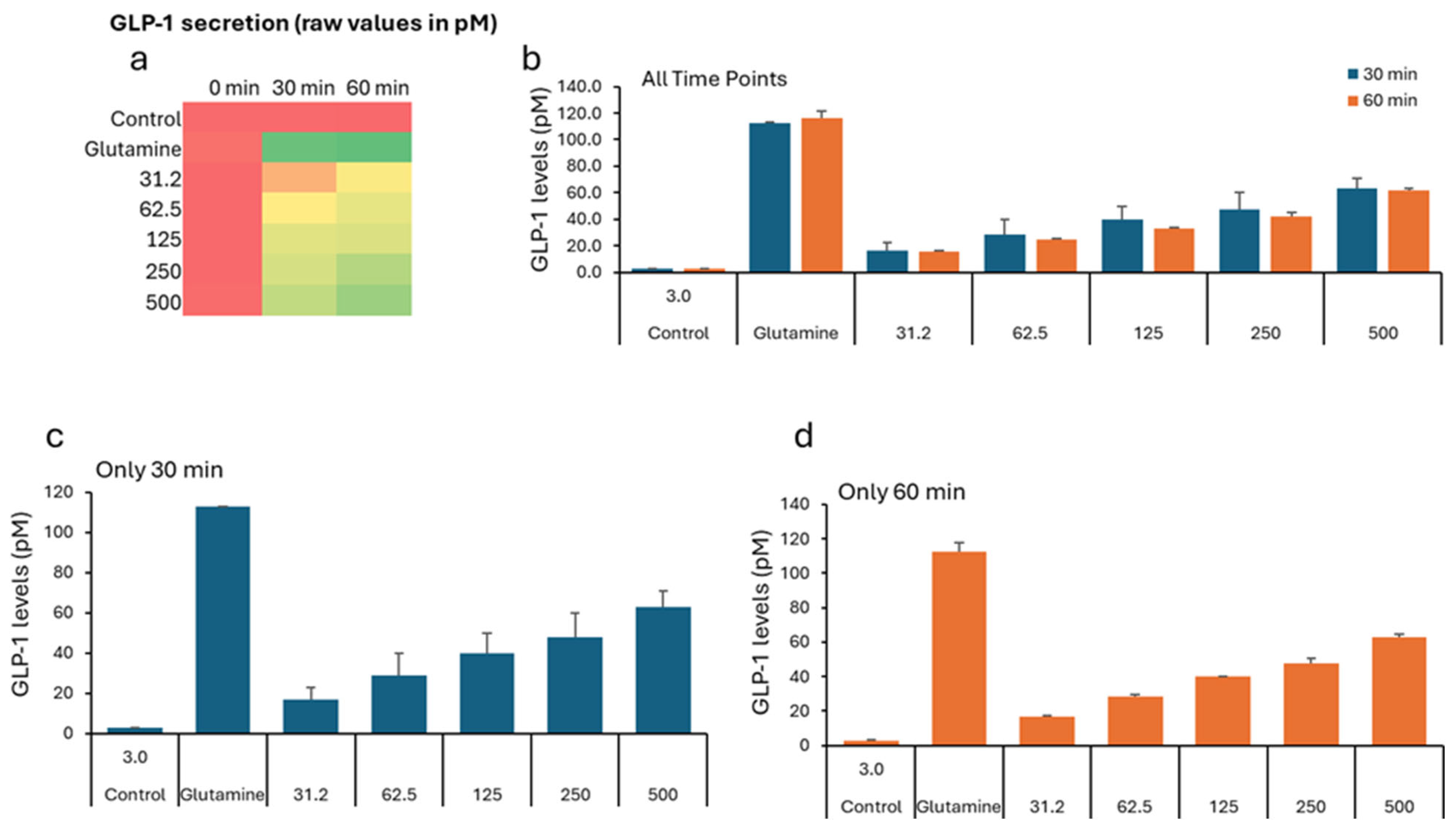
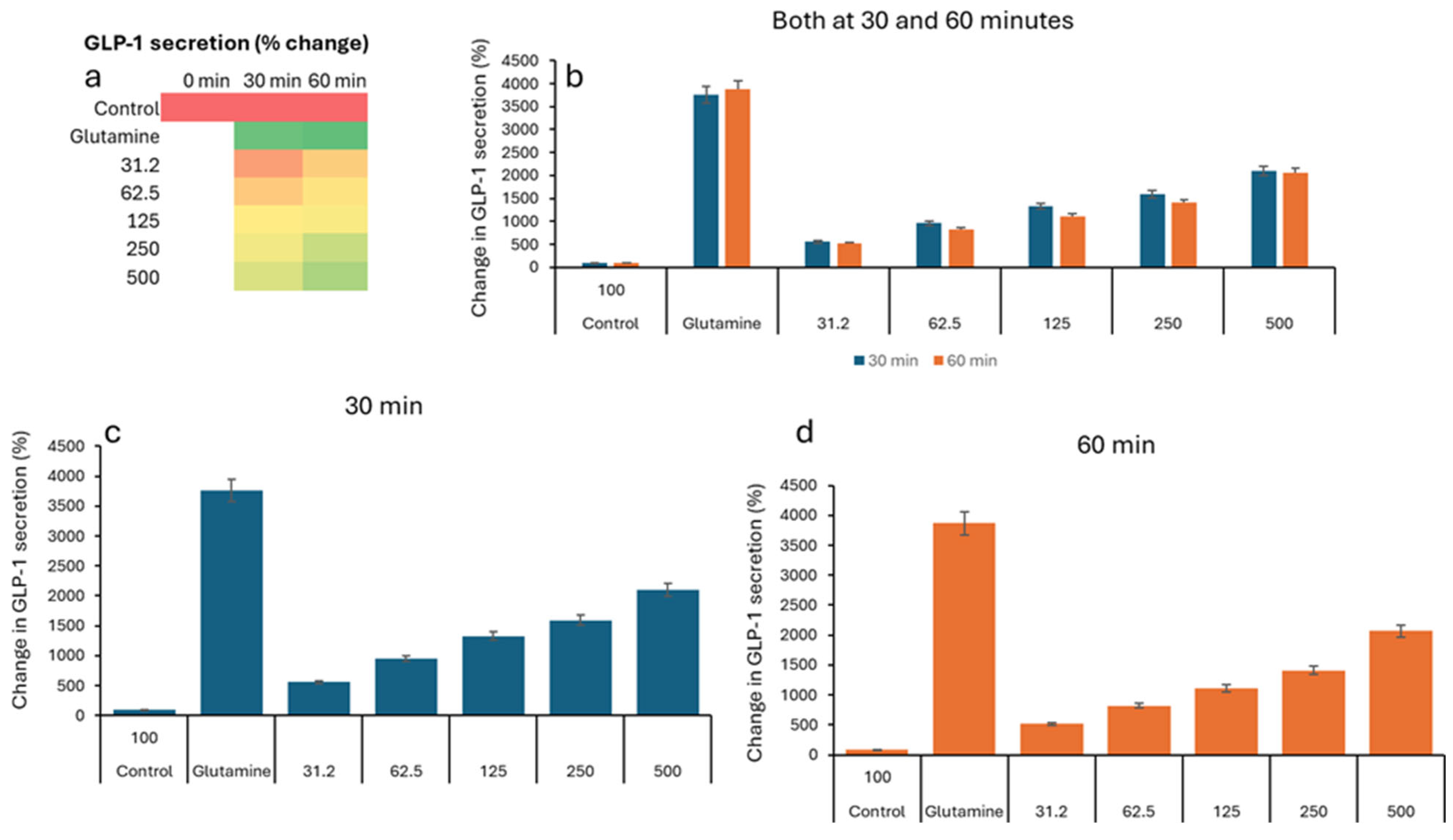
| Treatment (µg/mL) * | 0 min (Baseline) | 30 min | 60 min | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | SEM | Mean | SD | SEM | Mean | SD | SEM | |
| 0 (Negative Control) | 2.82 | 0.89 | 0.30 | 2.76 | 0.25 | 0.08 | 2.59 | 0.46 | 0.15 |
| 31.2 | 2.42 | 0.35 | 0.12 | 25.78 | 3.45 | 1.15 | 45.66 | 4.74 | 1.58 |
| 62.5 | 2.32 | 0.20 | 0.07 | 43.79 | 6.85 | 2.28 | 54.93 | 10.85 | 3.62 |
| 125 | 2.49 | 0.46 | 0.15 | 58.03 | 11.26 | 3.75 | 60.53 | 9.22 | 3.07 |
| 250 | 2.40 | 0.17 | 0.06 | 62.92 | 2.79 | 0.93 | 78.46 | 11.77 | 3.92 |
| 500 | 3.45 | 0.35 | 0.12 | 72.10 | 7.74 | 2.58 | 89.57 | 8.58 | 2.86 |
| Positive Control (Glutamine-10 mM) | 5.07 | 2.21 | 0.74 | 112.63 | 0.48 | 0.16 | 115.93 | 5.23 | 1.74 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arukha, A.P.; Nayak, S.; Swain, D.M. Effect of Akkermansia muciniphila on GLP-1 and Insulin Secretion. Nutrients 2025, 17, 2516. https://doi.org/10.3390/nu17152516
Arukha AP, Nayak S, Swain DM. Effect of Akkermansia muciniphila on GLP-1 and Insulin Secretion. Nutrients. 2025; 17(15):2516. https://doi.org/10.3390/nu17152516
Chicago/Turabian StyleArukha, Ananta Prasad, Subhendu Nayak, and Durga Madhab Swain. 2025. "Effect of Akkermansia muciniphila on GLP-1 and Insulin Secretion" Nutrients 17, no. 15: 2516. https://doi.org/10.3390/nu17152516
APA StyleArukha, A. P., Nayak, S., & Swain, D. M. (2025). Effect of Akkermansia muciniphila on GLP-1 and Insulin Secretion. Nutrients, 17(15), 2516. https://doi.org/10.3390/nu17152516