
Stachydrine Showing Metabolic Changes in Mice Exposed to House Dust Mites Ameliorates Allergen-Induced Inflammation

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Chronic HDM-Induced Airway Inflammation Mouse Model
2.3. AHR Measurement
2.4. Bronchoalveolar Lavage Fluid (BALF) Collection and Immune Cell Analysis
2.5. Serum IgE and Cytokine Measurement
2.6. Histological Analysis of Lung Tissue
2.7. Metabolite Analysis
2.8. Cell Culture and HDM Exposure
2.9. Western Blot Analysis
2.10. Statistical Analysis
3. Results
3.1. HDM Challenge Induces Chronic Airway Inflammation
3.2. Inflammatory Mediator Expression
3.3. Metabolite Profile Analysis in Mouse Serum
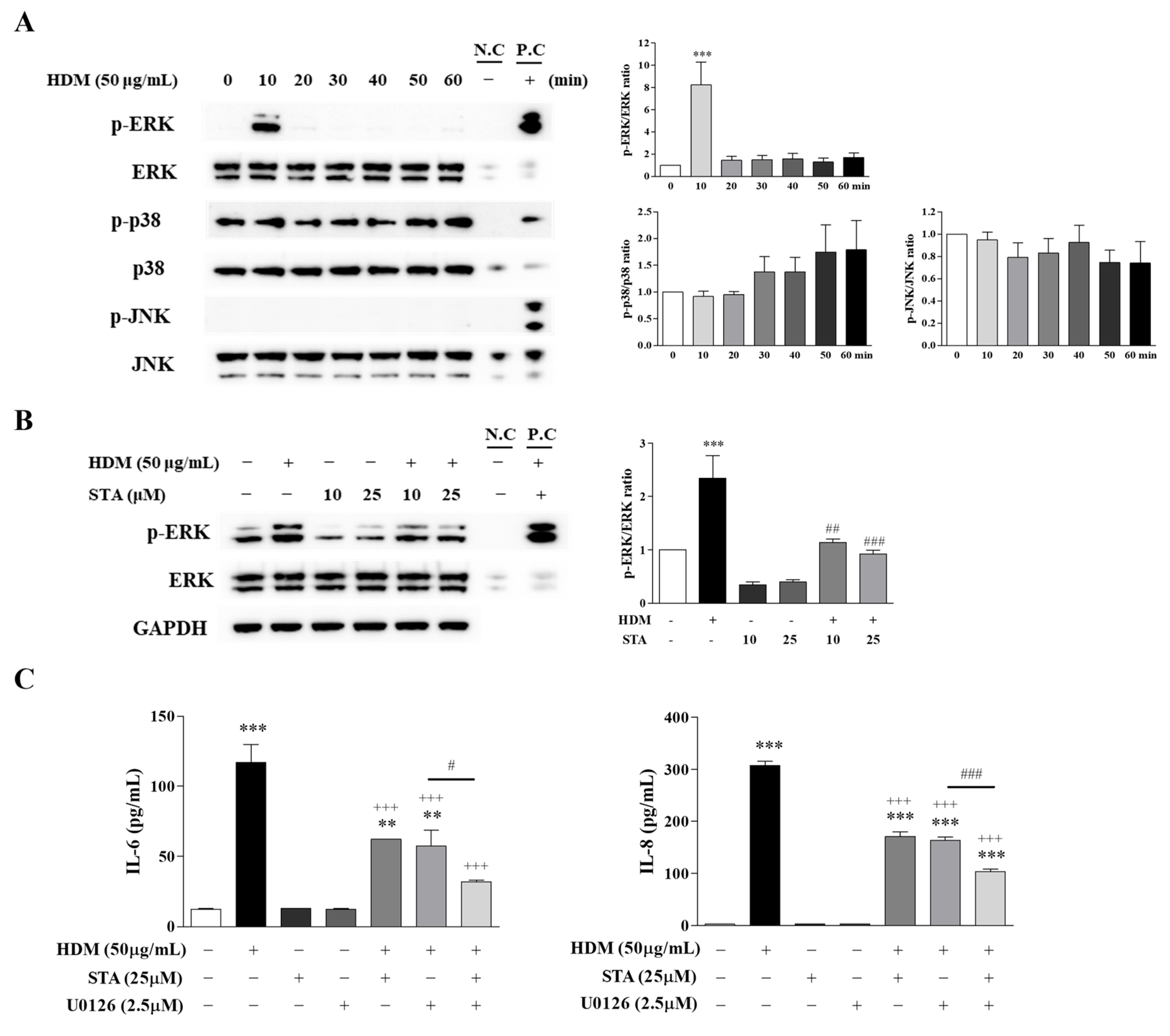
3.4. Effect of STA on HDM-Induced Inflammation in BEAS-2B Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| STA | Stachydrine |
| HDM | House dust mite |
| IL | Interleukin |
| ERK | Extracellular signal-regulated kinase |
References
- Kim, M.S.; Cho, K.A.; Cho, Y.J.; Woo, S.Y. Effects of interleukin-9 blockade on chronic airway inflammation in murine asthma models. Allergy Asthma Immunol. Res. 2013, 5, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Carroll, O.R.; Pillar, A.L.; Brown, A.C.; Feng, M.; Chen, H.; Donovan, C. Advances in respiratory physiology in mouse models of experimental asthma. Front Physiol. 2023, 14, 1099719. [Google Scholar] [CrossRef] [PubMed]
- Porpodis, K.; Tsiouprou, I.; Apostolopoulos, A.; Ntontsi, P.; Fouka, E.; Papakosta, D.; Vliagoftis, H.; Domvri, K. Eosinophilic asthma, phenotypes-endotypes and current biomarkers of choice. J. Pers. Med. 2022, 12, 1093. [Google Scholar] [CrossRef] [PubMed]
- Svenningsen, S.; Nair, P. Asthma endotypes and an overview of targeted therapy for asthma. Front. Med. 2017, 4, 158. [Google Scholar] [CrossRef]
- Ito, K.; Herbert, C.; Siegle, J.S.; Vuppusetty, C.; Hansbro, N.; Thomas, P.S.; Foster, P.S.; Barnes, P.J.; Kumar, R.K. Steroid-resistant neutrophilic inflammation in a mouse model of an acute exacerbation of asthma. Am. J. Respir. Cell Mol. Biol. 2008, 39, 543–550. [Google Scholar] [CrossRef]
- Rodriguez-Coira, J.; Villaseñor, A.; Izquierdo, E.; Huang, M.; Barker-Tejeda, T.C.; Radzikowska, U.; Sokolowska, M.; Barber, D. The importance of metabolism for immune homeostasis in allergic diseases. Front. Immunol. 2021, 12, 692004. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, T.; Zhao, X.; Wang, J.; Wang, Q. Plasma metabolites and gut microbiota are associated with T cell imbalance in BALB/c model of eosinophilic asthma. Front. Pharmacol. 2022, 13, 819747. [Google Scholar] [CrossRef]
- Choi, Y.; Kim, M.; Kim, S.J.; Yoo, H.J.; Kim, S.H.; Park, H.S. Metabolic shift favoring C18:0 ceramide accumulation in obese asthma. Allergy 2020, 75, 2858–2866. [Google Scholar] [CrossRef]
- Shao, Z.; Lu, J.; Zhang, C.; Zeng, G.; Chen, B.; Liang, H.; Wu, A.; Zhang, X.; Wang, X. Stachydrine ameliorates the progression of intervertebral disc degeneration via the PI3K/Akt/NF-κB signaling pathway: In vitro and in vivo studies. Food Funct. 2020, 11, 10864–10875. [Google Scholar] [CrossRef]
- Xie, X.; Zhang, Z.; Wang, X.; Luo, Z.; Lai, B.; Xiao, L.; Wang, N. Stachydrine protects eNOS uncoupling and ameliorates endothelial dysfunction induced by homocysteine. Mol. Med. 2018, 24, 10. [Google Scholar] [CrossRef]
- He, Z.; Li, P.; Liu, P.; Xu, P. Exploring stachydrine: From natural occurrence to biological activities and metabolic pathways. Front. Plant Sci. 2024, 15, 1442879. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yang, A.; Wu, Y.; Guan, W.; Xiong, B.; Peng, X.; Wei, X.; Chen, C.; Liu, Z. Stachydrine ameliorates carbon tetrachloride-induced hepatic fibrosis by inhibiting inflammation, oxidative stress and regulating MMPs/TIMPs system in rats. Biomed. Pharmacother. 2018, 97, 1586–1594. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Shan, X.; Chen, H.; Li, Z.; Zhao, P.; Zhang, C.; Guo, W.; Xu, M.; Lu, R. Stachydrine ameliorates cardiac fibrosis through inhibition of angiotensin II/transformation growth factor β1 fibrogenic axis. Front. Pharmacol. 2019, 10, 538. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Zhang, X.; Larsson, L. Metabolomic profiling of respiratory muscles and lung in response to long-term controlled mechanical ventilation. Front. Cell Dev. Biol. 2022, 10, 849973. [Google Scholar] [CrossRef]
- Gandhi, V.D.; Vliagoftis, H. Airway epithelium interactions with aeroallergens: Role of secreted cytokines and chemokines in innate immunity. Front. Immunol. 2015, 6, 147. [Google Scholar] [CrossRef]
- Jakwerth, C.A.; Ordovas-Montanes, J.; Blank, S.; Schmidt-Weber, C.B.; Zissler, U.M. Role of respiratory epithelial cells in allergic diseases. Cells 2022, 11, 1387. [Google Scholar] [CrossRef]
- Böhm, L.; Maxeiner, J.; Meyer-Martin, H.; Reuter, S.; Finotto, S.; Klein, M.; Schild, H.; Schmitt, E.; Bopp, T.; Taube, C. IL-10 and regulatory T cells cooperate in allergen-specific immunotherapy to ameliorate allergic asthma. J. Immunol. 2015, 194, 887–897. [Google Scholar] [CrossRef]
- Lambrecht, B.N.; Hammad, H.; Fahy, J.V. The cytokines of asthma. Immunity 2019, 50, 975–991. [Google Scholar] [CrossRef]
- Lin, S.C.; Shi, L.S.; Ye, Y.L. Advanced molecular knowledge of therapeutic drugs and natural products focusing on inflammatory cytokines in asthma. Cells 2019, 8, 685. [Google Scholar] [CrossRef]
- Fallon, P.G.; Schwartz, C. The high and lows of type 2 asthma and mouse models. J. Allergy Clin. Immunol. 2020, 145, 496–498. [Google Scholar] [CrossRef]
- Branchett, W.J.; Stölting, H.; Oliver, R.A.; Walker, S.A.; Puttur, F.; Gregory, L.G.; Gabryšová, L.; Wilson, M.S.; O’Garra, A.; Lloyd, C.M. A T cell-myeloid IL-10 axis regulates pathogenic IFN-γ-dependent immunity in a mouse model of type 2-low asthma. J. Allergy Clin. Immunol. 2020, 145, 666–678.e9. [Google Scholar] [CrossRef] [PubMed]
- Margelidon-Cozzolino, V.; Tsicopoulos, A.; Chenivesse, C.; de Nadai, P. Role of Th17 cytokines in airway remodeling in asthma and therapy perspectives. Front. Allergy 2022, 3, 806391. [Google Scholar] [CrossRef] [PubMed]
- Maltby, S.; Tay, H.L.; Yang, M.; Foster, P.S. Mouse models of severe asthma: Understanding the mechanisms of steroid resistance, tissue remodelling and disease exacerbation. Respirology 2017, 22, 874–885. [Google Scholar] [CrossRef] [PubMed]
- Padrón-Morales, J.; García-Solaesa, V.; Isidoro-García, M.; Hernández-Hernández, L.; García-Sánchez, A.; Hincapié-López, G.; Lorente-Toledano, F.; Dávila, I.; Sanz, C. Implications of cytokine genes in allergic asthma. Allergol. Immunopathol. 2014, 42, 603–608. [Google Scholar] [CrossRef]
- Radzikowska, U.; Baerenfaller, K.; Cornejo-Garcia, J.A.; Karaaslan, C.; Barletta, E.; Sarac, B.E.; Zhakparov, D.; Villaseñor, A.; Eguiluz-Gracia, I.; Mayorga, C.; et al. Omics technologies in allergy and asthma research: An EAACI position paper. Allergy 2022, 77, 2888–2908. [Google Scholar] [CrossRef]
- Reinke, S.N.; Gallart-Ayala, H.; Gómez, C.; Checa, A.; Fauland, A.; Naz, S.; Kamleh, M.A.; Djukanović, R.; Hinks, T.S.C.; Wheelock, C.E. Metabolomics analysis identifies different metabotypes of asthma severity. Eur. Respir. J. 2017, 49, 1601740. [Google Scholar] [CrossRef]
- Wang, C.; Jiang, S.; Zhang, S.; Ouyang, Z.; Wang, G.; Wang, F. Research progress of metabolomics in asthma. Metabolites 2021, 11, 567. [Google Scholar] [CrossRef]
- Shin, H.K.; Bang, Y.J. Aromatic Amino Acid Metabolites: Molecular Messengers Bridging Immune-Microbiota Communication. Immune Netw. 2025, 25, e10. [Google Scholar] [CrossRef]
- Liao, L.; Tang, Y.; Li, B.; Tang, J.; Xu, H.; Zhao, K.; Zhang, X. Stachydrine, a potential drug for the treatment of cardiovascular system and central nervous system diseases. Biomed. Pharmacother. 2023, 161, 114489. [Google Scholar] [CrossRef]
- Zeng, H.; Xu, D.; Song, Y.; Tian, S.; Qiao, J.; Li, Z.; Zhao, L.; Shi, H.; Zhou, Y.; Li, S.; et al. Synthesis, characterization and anti-breast cancer activities of stachydrine derivatives. Eur. J. Med. Chem. 2023, 259, 115679. [Google Scholar] [CrossRef]
- Zhai, Z.; Mu, T.; Zhao, L.; Zhu, D.; Zhong, X.; Li, Y.; Liang, C.; Li, W.; Zhou, Q. Stachydrine represses the proliferation and enhances cell cycle arrest and apoptosis of breast cancer cells via PLA2G2A/DCN axis. Chem. Biol. Drug Des. 2024, 103, e14429. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Zhou, Y.; Wang, M.; Guo, C.; Cao, Z.; Zhang, R.; Peng, C. A review of pharmacological and pharmacokinetic properties of stachydrine. Pharmacol. Res. 2020, 155, 104755. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Sun, L.; Qiu, Y.; Zhu, W.; Hu, K.; Mao, J. Protective effect of stachydrine against cerebral ischemia-reperfusion injury by reducing inflammation and apoptosis through P65 and JAK2/STAT3 signaling pathway. Front. Pharmacol. 2020, 11, 64. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Zhou, C.; Zhang, W.; Wang, W.; He, B.; Hu, B.; Jiang, G.; Wang, Y.; Hong, J.; Li, S.; et al. Stachydrine prevents LPS-induced bone loss by inhibiting osteoclastogenesis via NF-κB and Akt signalling. J. Cell. Mol. Med. 2019, 23, 6730–6743. [Google Scholar] [CrossRef]
- Shi, Q.; Liu, Z.; Yang, Y.; Geng, P.; Zhu, Y.Y.; Zhang, Q.; Bai, F.; Bai, G. Identification of anti-asthmatic compounds in Pericarpium citri reticulatae and evaluation of their synergistic effects. Acta Pharmacol. Sin. 2009, 30, 567–575. [Google Scholar] [CrossRef]
- Smith, K.G.; Kamdar, A.A.; Stark, J.M. Chapter 8, Lung defenses: Intrinsic, innate, and adaptive. In Kendig’s Disorders of the Respiratory Tract in Children, 9th ed.; Wilmot, R.W., Bush, A., Deterding, R.R., Ratjen, F., Sly, P., Zar, H., Li, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 120–133.e2. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Name | PubChem CID | 14 PBS vs. 14 Der.p | 26 PBS vs. 26 Der.p | 35 PBS vs. 35 Der.p | Up/Down | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ratio | p-Value | Ratio | p-Value | Ratio | p-Value | ||||||||
| 1-Methyladenosine | 27476 | 1.2 | 0.034 | * | 1.0 | 0.503 | 1.2 | 0.446 | ↑ | - | - | ||
| 2-Hydroxybutyric acid | 440864 | 1.6 | 0.015 | * | 0.8 | 0.156 | 1.1 | 0.515 | ↑ | - | - | ||
| Dihydroxyacetone phosphate | 668 | 1.6 | 0.016 | * | 0.7 | 0.424 | 0.9 | 0.483 | ↑ | - | - | ||
| Fructose 6-phosphate | 603 | 2.0 | 0.005 | ** | 0.6 | 0.371 | 0.7 | 0.111 | ↑ | - | - | ||
| Gluconic acid | 10690 | 1.4 | 0.041 | * | 0.9 | 0.236 | 1.1 | 0.434 | ↑ | - | - | ||
| Glucosamine 6-sulfuric acid | 72361 | 2.2 | 0.024 | * | <1 | N.A. | N.A. | N.A. | ↑ | - | - | ||
| Glucose 1-phosphate | 65533 | 1.7 | 0.025 | * | 0.6 | N.A. | 0.5 | 0.155 | ↑ | - | - | ||
| Glucose 6-phosphate | 5958 | 1.8 | 0.001 | ** | 0.5 | 0.296 | 0.6 | 0.146 | ↑ | - | - | ||
| N,N-Dimethylglycine | 673 | 1.2 | 0.025 | * | 0.9 | 0.215 | 1.0 | 0.971 | ↑ | - | - | ||
| Nicotinamide | 936 | 1.3 | 0.012 | * | 0.6 | 0.157 | 1.0 | 0.676 | ↑ | - | - | ||
| Ribose 5-phosphate | 439167 | 1.8 | 0.017 | * | 0.6 | 0.280 | 0.8 | 0.588 | ↑ | - | - | ||
| Ribulose 5-phosphate | 439184 | 1.5 | 0.012 | * | 0.6 | 0.170 | 0.9 | 0.693 | ↑ | - | - | ||
| S-Methylglutathione | 115260 | 1.4 | 0.038 | * | 1.0 | N.A. | 0.9 | 0.546 | ↑ | - | - | ||
| Sedoheptulose 7-phosphate | 165007 | 2.1 | 0.013 | * | 0.7 | 0.539 | 0.7 | 0.261 | ↑ | - | - | ||
| Serotonin | 5202 | 1.2 | 0.010 | ** | 1.0 | 0.851 | 0.7 | 0.266 | ↑ | - | - | ||
| Succinic acid | 1110 | 1.6 | 0.006 | ** | 0.7 | 0.469 | 1.0 | 0.866 | ↑ | - | - | ||
| Uracil | 1174 | 1.8 | 0.031 | * | 0.6 | 0.131 | 0.8 | 0.211 | ↑ | - | - | ||
| Xanthosine | 64959 | 2.0 | 0.044 | * | 0.9 | 0.826 | 0.7 | 0.437 | ↑ | - | - | ||
| Anserine_divalent | 112072 | 0.7 | 0.038 | * | 0.8 | 0.293 | 1.2 | 0.505 | ↓ | - | - | ||
| Citrulline | 9750 | 0.8 | 0.002 | ** | 0.9 | 0.214 | 1.1 | 0.507 | ↓ | - | - | ||
| N-Acetylglycine | 10972 | 0.8 | 0.011 | * | 0.9 | 0.365 | 1.1 | 0.622 | ↓ | - | - | ||
| N-Acetyltryptophan | 439917 | 0.6 | 0.040 | * | 0.6 | 0.089 | 1.1 | 0.774 | ↓ | - | - | ||
| γ-Glu-Asn | 131801686 | 0.8 | 0.005 | ** | 0.9 | 0.538 | 1.1 | 0.236 | ↓ | - | - | ||
| γ-Glu-Trp | 3038501 | 0.7 | 0.048 | * | 0.9 | 0.305 | 1.0 | 0.519 | ↓ | - | - | ||
| 1-Methylhistidine 3-Methylhistidine | 92105 64969 | 1.0 | 0.972 | 0.8 | 0.026 | * | 1.0 | 0.788 | - | ↓ | - | ||
| 3-Phosphoglyceric acid | 439183 | 1.2 | 0.378 | 0.5 | 0.040 | * | 1.0 | 0.816 | - | ↓ | - | ||
| Carnosine | 439224 | 0.9 | 0.604 | 0.7 | 0.032 | * | 0.9 | 0.792 | - | ↓ | - | ||
| Ile | 791 | 1.0 | 0.913 | 0.8 | 0.018 | * | 1.0 | 0.848 | - | ↓ | - | ||
| Leu | 857 | 0.9 | 0.336 | 0.8 | 0.042 | * | 0.8 | 0.200 | - | ↓ | - | ||
| Met | 876 | 1.0 | 0.387 | 0.9 | 0.038 | * | 1.0 | 0.837 | - | ↓ | - | ||
| N6,N6,N6-Trimethyllysine | 440120 | 1.0 | 0.774 | 0.7 | 0.030 | * | 1.0 | 0.787 | - | ↓ | - | ||
| SDMA | 169148 | 1.1 | 0.108 | 0.7 | 0.015 | * | 1.0 | 0.847 | - | ↓ | - | ||
| Tyr | 1153 | 1.0 | 0.535 | 0.8 | 0.038 | * | 0.9 | 0.594 | - | ↓ | - | ||
| XA0035 | 0.9 | 0.276 | 0.9 | 0.026 | * | 1.1 | 0.629 | - | ↓ | - | |||
| XC0001 | 0.9 | 0.528 | 0.8 | 0.012 | * | 0.9 | 0.691 | - | ↓ | - | |||
| XC0029 Stachydrine | 0 115244 | 1.0 | 0.702 | 0.8 | 0.007 | ** | 0.8 | 0.276 | - | ↓ | - | ||
| γ-Glu-Arg_divalent | 20719180 | 1.0 | 0.954 | 0.9 | 0.048 | * | 1.1 | 0.321 | - | ↓ | - | ||
| γ-Glu-Ser | 22844748 | 1.0 | 0.470 | 0.9 | 0.025 | * | 1.2 | 0.139 | - | ↓ | - | ||
| Butyric acid Isobutyric acid | 264 6590 | 1.4 | 0.229 | 1.0 | 0.983 | 1.3 | 0.022 | * | - | - | ↑ | ||
| N-Acetylserine | 65249 | 1.0 | 0.920 | 0.8 | 0.145 | 1.5 | 0.038 | * | - | - | ↑ | ||
| XA0012 | 1.1 | 0.584 | 0.8 | 0.361 | 1.4 | 0.048 | * | - | - | ↑ | |||
| XA0019 | 0.9 | 0.513 | 1.2 | 0.632 | 1.9 | 0.039 | * | - | - | ↑ | |||
| Trimethylamine N-oxide | 1145 | 1.3 | 0.289 | 0.9 | 0.243 | 0.7 | 0.049 | * | - | - | ↓ | ||
| XC0040 | 1.3 | 0.289 | 0.9 | 0.243 | 0.7 | 0.049 | * | - | - | ↓ | |||
| 2-Aminoisobutyric acid 2-Aminobutyric acid | 6119 6657 | 1.2 | 0.024 | * | 0.7 | 0.041 | * | 1.0 | 0.802 | ↑ | ↓ | - | |
| Ethyl glucuronide | 18392195 | 1.3 | 0.039 | * | N.A. | N.A. | 0.6 | 0.037 | * | ↑ | - | ↓ | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Do, J.-H.; Hong, J.Y.; Jang, J.-H.; Jeong, K.-T.; Kim, S.H.; Lee, H.-J. Stachydrine Showing Metabolic Changes in Mice Exposed to House Dust Mites Ameliorates Allergen-Induced Inflammation. Nutrients 2025, 17, 2015. https://doi.org/10.3390/nu17122015
Do J-H, Hong JY, Jang J-H, Jeong K-T, Kim SH, Lee H-J. Stachydrine Showing Metabolic Changes in Mice Exposed to House Dust Mites Ameliorates Allergen-Induced Inflammation. Nutrients. 2025; 17(12):2015. https://doi.org/10.3390/nu17122015
Chicago/Turabian StyleDo, Ji-Hye, Jung Yeon Hong, Ji-Hye Jang, Kyu-Tae Jeong, Seung Hyun Kim, and Hye-Ja Lee. 2025. "Stachydrine Showing Metabolic Changes in Mice Exposed to House Dust Mites Ameliorates Allergen-Induced Inflammation" Nutrients 17, no. 12: 2015. https://doi.org/10.3390/nu17122015
APA StyleDo, J.-H., Hong, J. Y., Jang, J.-H., Jeong, K.-T., Kim, S. H., & Lee, H.-J. (2025). Stachydrine Showing Metabolic Changes in Mice Exposed to House Dust Mites Ameliorates Allergen-Induced Inflammation. Nutrients, 17(12), 2015. https://doi.org/10.3390/nu17122015