
Effect of Sodium Butyrate Supplementation on Type 2 Diabetes—Literature Review



Abstract
1. Pathogenesis of Type 2 Diabetes
2. Global Trends and Demographic Patterns of T2D
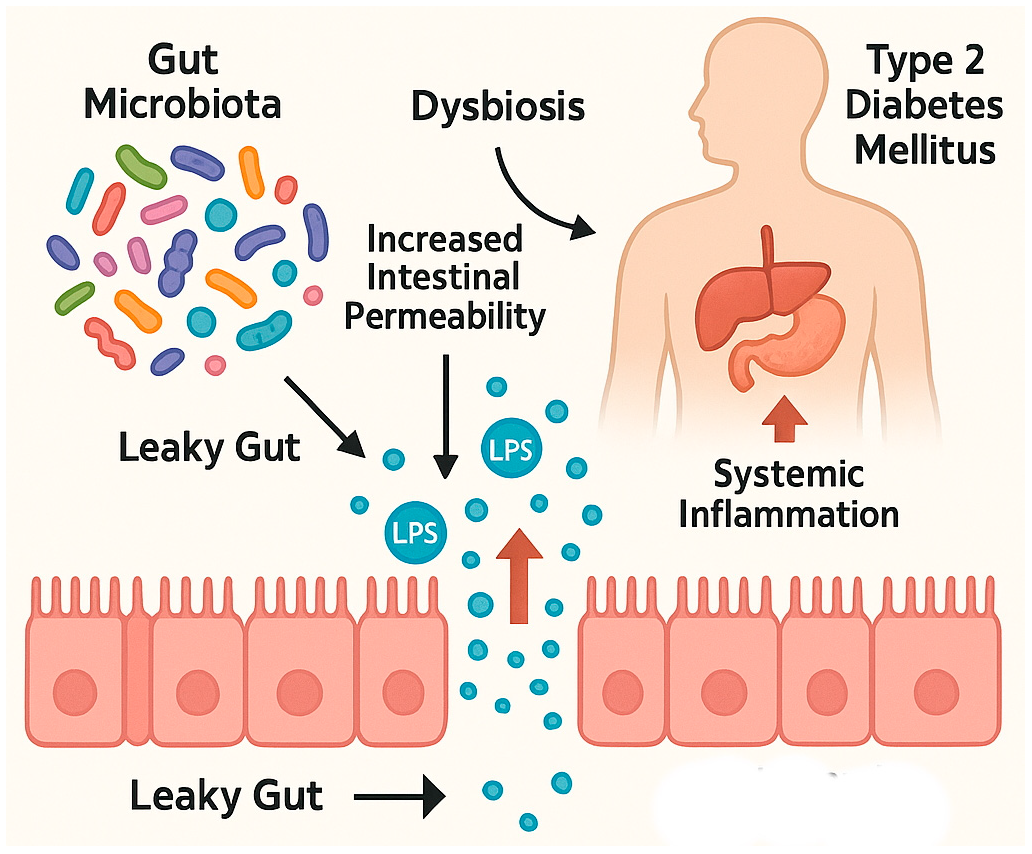
3. Disruption of the Intestinal Barrier and Its Implications in Type 2 Diabetes Mellitus
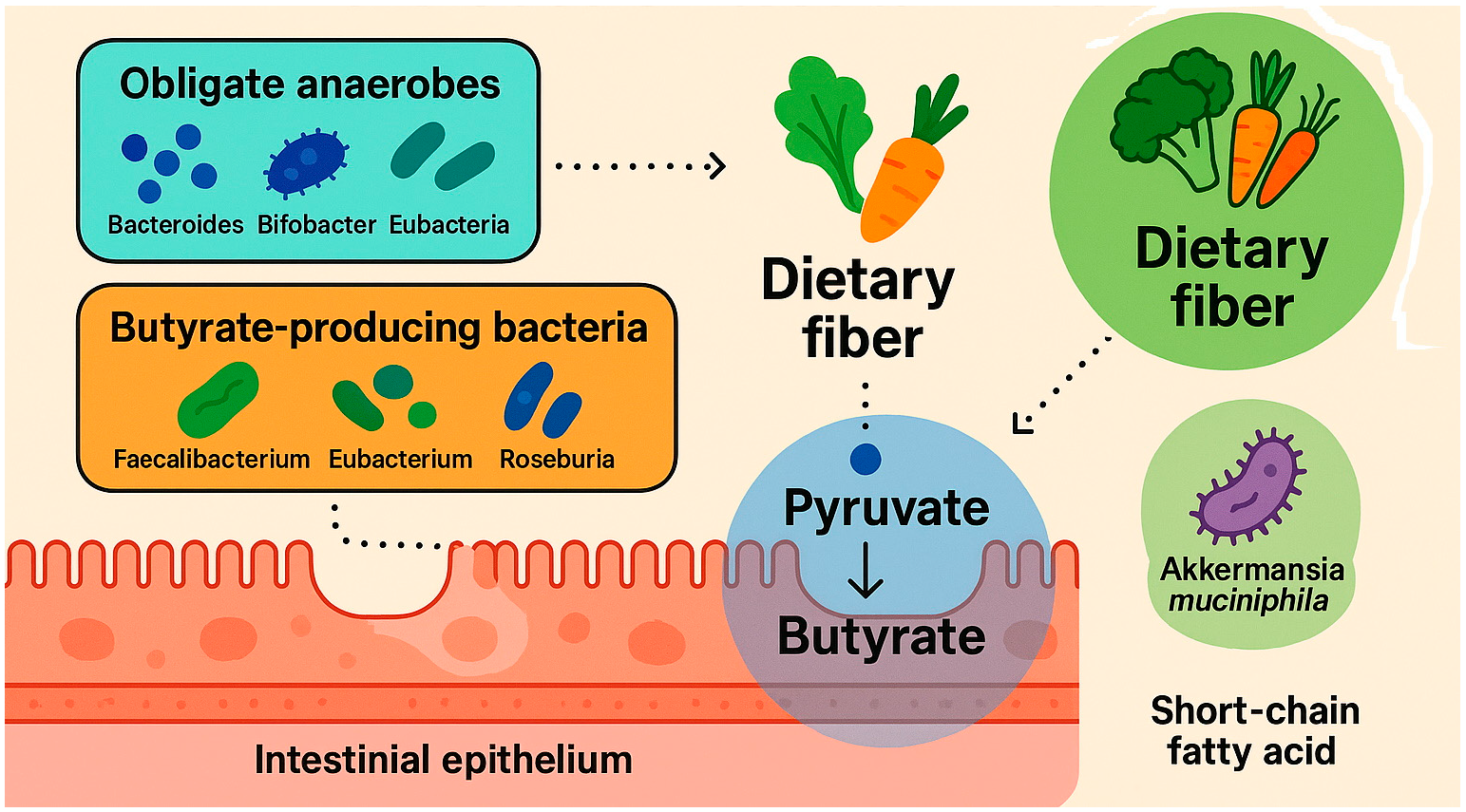
4. Mechanism of Butyrate Formation in the Gastrointestinal Tract
5. Forms, Dosages, and Safety of Sodium Butyrate Supplementation
6. Effect of Butyrate on the Intestinal Microbiota
7. Effect of Sodium Butyrate in Type 2 Diabetes: A Study Overview

{kind=link}
{kind=link}
| References | Model | Study Groups | Intervention | Survey Results |
| S. Khan et al. (2016) [69] | Animals | Sprague-Dawley rats with type 2 diabetes (n = 34) | NaB doses of 200 and 400 mg/kg twice daily or control metformin 150 mg/kg twice; for 10 weeks | Butyrate significantly reduced plasma glucose, HbA1c, insulin resistance, gluconeogenesis, and dyslipidaemia comparably to metformin. Butyrate alleviated micro- and macro-vesicular steatosis of the liver and fat deposition in brown adipose tissue and white adipose tissue (adipocyte hypertrophy). Butyrate also reduced damage to pancreatic beta cells. |
| Y. Hu et al. (2018) [75] | Animals | Sprague-Dawley rats with type 2 diabetes (n = 40) | NaB (500 mg/kg/d) was injected intraperitoneally; for 6 weeks | NaB improved insulin resistance and β-cell function and reduced β-cell apoptosis in rats with T2DM. NaB alleviated hyperglycaemia, lowered TC and LDL-c levels, prevented weight loss, and increased glucose tolerance. NaB ameliorated diabetes-induced islet histological changes and functional damage and alleviated β-cell apoptosis. |
| Ty-Hua Xu et al. (2018) [70] | Animals | 7-week-old male db/db mice (type 2 diabetes; n = 24) | (1) model group (2) intervention group (sodium butyrate 0.5 g/kg/day); (3) Metformin 0.15 g/kg/day control group); | Butyrate significantly reduced blood HbA1c, inflammatory cytokines, and LPS levels in db/db mice. NaB reduced inflammatory cell infiltration and increased intestinal integrity and intercellular adhesion molecules. NaBut had the effect of reducing the Firmicutes:Bacteroidetes ratio. In Caco-2 cells, butyrate significantly promoted cell proliferation, enhanced the cells’ ability to counteract oxidative stress, reduced the secretion of inflammatory cytokines, and preserved the single-cell integrity of the epithelium. |
| N. Roshanravan et al. (2017) [76] | People | 60 patients with type 2 diabetes; age 30–55 years; BMI 27 kg/m2–35 kg/m2 | 600 mg/d NaBut (group A), 10 g/d HP inulin (group B), 600 mg/d NaBut + 10 g/d HP inulin (Group C); placebo (group D); for 45 days | Reduction in diastolic blood pressure (in groups A, B, C) compared with the placebo group. Sodium butyrate + inulin intervention reduced fasting blood glucose levels and waist-to-hip ratio. Waist circumference (in groups B and C) decreased significantly after the intervention. Increase in GLP1 peptide levels (in groups A and C) compared to the placebo group. |
| N. Roshanravan et al. (2018) [72] | People | 60 patients with type 2 diabetes; age 30–55 years; BMI 27 kg/m2–35 kg/m2 | 600 mg/d NaBut (group A), 10 g/d HP inulin (group B), 600 mg/d NaBut + 10 g/d HP inulin (Group C); placebo (group D); for 45 days | A. muciniphila increased significantly after supplementation with HP inulin and (separately) butyrate. Increased expression of microRNA-375 was observed after supplementation with butyrate and butyrate + inulin compared to the placebo group. |
| N. Roshanravan et al. (2020) [71] | People | 60 patients with type 2 diabetes; age 30–55 years; BMI 27 kg/m2–35 kg/m2 | 600 mg/d NaBut (Group A), 10 g/d HP inulin (group B), 600 mg/d NaBut + 10 g/d HP inulin (Group C); placebo (group D); for 45 days | Butyrate relatively decreased the expression levels of TLR2/4, NF-κB1, caspase-1, NLRP3, IL-1β, and IL-18. Butyrate + inulin—increased fold change in miR-146a and miR-9 compared to the placebo group. Change in total antioxidant capacity and superoxide dismutase were significantly increased after butyrate and concomitant butyrate + inulin. |
| Z. Khosravi et al. (2022) [74] | People | 42 patients with type 2 diabetes | NaBut (n = 21) (600 mg/d) or placebo (n = 21); for 6 weeks | Butyrate significantly reduced systolic and diastolic blood pressure. BS2hpp decreased significantly in the intervention group and the placebo group. NaBut significantly increased insulin levels, total cholesterol, low-density lipoprotein cholesterol and reduced NO levels. |
| P. Panufnik et al. (2024) [46] | People | 52 patients with type 2 diabetes and abdominal pain | Butyrate at a dose of 1.5 g/day (n = 29) or placebo (n = 23); for 12 weeks | Alleviation of gastrointestinal signs and symptoms. Slight but significant improvements in BMI and HbA1C levels. A significant decrease in the incidence of SIBO. |
8. Conclusions
Funding
Conflicts of Interest
References
- Onyango, E.M.; Onyango, B.M. The Rise of Noncommunicable Diseases in Kenya: An Examination of the Time Trends and Contribution of the Changes in Diet and Physical Inactivity. J. Epidemiol. Glob. Health 2018, 8, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ruze, R.; Liu, T.; Zou, X.; Song, J.; Chen, Y.; Xu, R.; Yin, X.; Xu, Q. Obesity and Type 2 Diabetes Mellitus: Connections in Epidemiology, Pathogenesis, and Treatments. Front. Endocrinol. 2023, 14, 1161521. [Google Scholar] [CrossRef]
- Khan, R.M.M.; Chua, Z.J.Y.; Tan, J.C.; Yang, Y.; Liao, Z.; Zhao, Y. From Pre-Diabetes to Diabetes: Diagnosis, Treatments and Translational Research. Medicina 2019, 55, 546. [Google Scholar] [CrossRef]
- Konsekwencje Ekonomiczne Otyłości | Cukrzyca | Amerykańskie Stowarzyszenie Diabetologiczne. Available online: https://diabetesjournals.org/diabetes/article/56/11/2668/13593/Economic-Consequences-of-the-Obese (accessed on 27 December 2024).
- Balkau, B.; Mhamdi, L.; Oppert, J.-M.; Nolan, J.; Golay, A.; Porcellati, F.; Laakso, M.; Ferrannini, E. Physical Activity and Insulin Sensitivity. Diabetes 2008, 57, 2613–2618. [Google Scholar] [CrossRef]
- Chang, A.M.; Halter, J.B. Aging and Insulin Secretion. Am. J. Physiol.-Endocrinol. Metab. 2003, 284, E7–E12. [Google Scholar] [CrossRef]
- Larsson, S.C.; Permert, J.; Håkansson, N.; Näslund, I.; Bergkvist, L.; Wolk, A. Overall Obesity, Abdominal Adiposity, Diabetes and Cigarette Smoking in Relation to the Risk of Pancreatic Cancer in Two Swedish Population-Based Cohorts. Br. J. Cancer 2005, 93, 1310–1315. [Google Scholar] [CrossRef]
- Yun, J.E.; Kimm, H.; Choi, Y.J.; Jee, S.H.; Huh, K.B. Smoking Is Associated with Abdominal Obesity, Not Overall Obesity, in Men with Type 2 Diabetes. J. Prev. Med. Public Health 2012, 45, 316–322. [Google Scholar] [CrossRef]
- Knott, C.; Bell, S.; Britton, A. Alcohol Consumption and the Risk of Type 2 Diabetes: A Systematic Review and Dose-Response Meta-Analysis of More Than 1.9 Million Individuals From 38 Observational Studies. Diabetes Care 2015, 38, 1804–1812. [Google Scholar] [CrossRef]
- Doria, A.; Patti, M.-E.; Kahn, C.R. The Emerging Genetic Architecture of Type 2 Diabetes. Cell Metab. 2008, 8, 186–200. [Google Scholar] [CrossRef]
- Khan, M.A.B.; Hashim, M.J.; King, J.K.; Govender, R.D.; Mustafa, H.; Al Kaabi, J. Epidemiology of Type 2 Diabetes—Global Burden of Disease and Forecasted Trends. J. Epidemiol. Glob. Health 2020, 10, 107–111. [Google Scholar] [CrossRef]
- Kautzky-Willer, A.; Harreiter, J.; Pacini, G. Sex and Gender Differences in Risk, Pathophysiology and Complications of Type 2 Diabetes Mellitus. Endocr. Rev. 2016, 37, 278–316. [Google Scholar] [CrossRef] [PubMed]
- Cioana, M.; Deng, J.; Nadarajah, A.; Hou, M.; Qiu, Y.; Chen, S.S.J.; Rivas, A.; Toor, P.P.; Banfield, L.; Thabane, L.; et al. Global Prevalence of Diabetic Retinopathy in Pediatric Type 2 Diabetes. JAMA Netw. Open 2023, 6, e231887. [Google Scholar] [CrossRef] [PubMed]
- Dabelea, D.; Stafford, J.M.; Mayer-Davis, E.J.; D’Agostino, R.; Dolan, L.; Imperatore, G.; Linder, B.; Lawrence, J.M.; Marcovina, S.M.; Mottl, A.K.; et al. Association of Type 1 Diabetes vs. Type 2 Diabetes Diagnosed During Childhood and Adolescence with Complications During Teenage Years and Young Adulthood. JAMA 2017, 317, 825–835. [Google Scholar] [CrossRef]
- Ogunrinola, G.A.; Oyewale, J.O.; Oshamika, O.O.; Olasehinde, G.I. The Human Microbiome and Its Impacts on Health. Int. J. Microbiol. 2020, 2020, 8045646. [Google Scholar] [CrossRef]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What Is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef]
- Zhang, Z.; Tian, T.; Chen, Z.; Liu, L.; Luo, T.; Dai, J. Characteristics of the Gut Microbiome in Patients with Prediabetes and Type 2 Diabetes. PeerJ 2021, 9, e10952. [Google Scholar] [CrossRef]
- Gaike, A.H.; Paul, D.; Bhute, S.; Dhotre, D.P.; Pande, P.; Upadhyaya, S.; Reddy, Y.; Sampath, R.; Ghosh, D.; Chandraprabha, D.; et al. The Gut Microbial Diversity of Newly Diagnosed Diabetics but Not of Prediabetics Is Significantly Different from That of Healthy Nondiabetics. mSystems 2020, 5, e00578-19. [Google Scholar] [CrossRef]
- Shanahan, F.; Ghosh, T.S.; O’Toole, P.W. The Healthy Microbiome—What Is the Definition of a Healthy Gut Microbiome? Gastroenterology 2021, 160, 483–494. [Google Scholar] [CrossRef]
- Hollister, E.B.; Gao, C.; Versalovic, J. Compositional and Functional Features of the Gastrointestinal Microbiome and Their Effects on Human Health. Gastroenterology 2014, 146, 1449–1458. [Google Scholar] [CrossRef]
- Snelson, M.; de Pasquale, C.; Ekinci, E.I.; Coughlan, M.T. Gut Microbiome, Prebiotics, Intestinal Permeability and Diabetes Complications. Best Pract. Res. Clin. Endocrinol. Metab. 2021, 35, 101507. [Google Scholar] [CrossRef]
- Gurung, M.; Li, Z.; You, H.; Rodrigues, R.; Jump, D.B.; Morgun, A.; Shulzhenko, N. Role of Gut Microbiota in Type 2 Diabetes Pathophysiology. eBioMedicine 2020, 51, 102590. [Google Scholar] [CrossRef] [PubMed]
- Dimba, N.R.; Mzimela, N.; Khathi, A. Improved Gut Health May Be a Potential Therapeutic Approach for Managing Prediabetes: A Literature Review. Biomedicines 2024, 12, 1275. [Google Scholar] [CrossRef]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D.; et al. A Metagenome-Wide Association Study of Gut Microbiota in Type 2 Diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef]
- Hu, R.; Yuan, Y.; Liu, C.; Zhou, J.; Ji, L.; Jiang, G. New Insights into the Links between Anti-Diabetes Drugs and Gut Microbiota. Endocr. Connect. 2020, 10, R36–R42. [Google Scholar] [CrossRef]
- Sadagopan, A.; Mahmoud, A.; Begg, M.; Tarhuni, M.; Fotso, M.; Gonzalez, N.A.; Sanivarapu, R.R.; Osman, U.; Latha Kumar, A.; Mohammed, L. Understanding the Role of the Gut Microbiome in Diabetes and Therapeutics Targeting Leaky Gut: A Systematic Review. Cureus 2023, 15, e41559. [Google Scholar] [CrossRef]
- Iatcu, C.O.; Steen, A.; Covasa, M. Gut Microbiota and Complications of Type-2 Diabetes. Nutrients 2021, 14, 166. [Google Scholar] [CrossRef]
- Gomes, J.M.G.; de Assis Costa, J.; de Cássia Gonçalves Alfenas, R. Metabolic Endotoxemia and Diabetes Mellitus: A Systematic Review. Metabolism 2017, 68, 133–144. [Google Scholar] [CrossRef]
- De Santis, S.; Cavalcanti, E.; Mastronardi, M.; Jirillo, E.; Chieppa, M. Nutritional Keys for Intestinal Barrier Modulation. Front. Immunol. 2015, 6, 612. [Google Scholar] [CrossRef]
- Veres-Székely, A.; Szász, C.; Pap, D.; Szebeni, B.; Bokrossy, P.; Vannay, Á. Zonulin as a Potential Therapeutic Target in Microbiota-Gut-Brain Axis Disorders: Encouraging Results and Emerging Questions. Int. J. Mol. Sci. 2023, 24, 7548. [Google Scholar] [CrossRef]
- Fasano, A. All Disease Begins in the (Leaky) Gut: Role of Zonulin-Mediated Gut Permeability in the Pathogenesis of Some Chronic Inflammatory Diseases. F1000Research 2020, 9, 69. [Google Scholar] [CrossRef]
- Cani, P.D.; Possemiers, S.; Van de Wiele, T.; Guiot, Y.; Everard, A.; Rottier, O.; Geurts, L.; Naslain, D.; Neyrinck, A.; Lambert, D.M.; et al. Changes in Gut Microbiota Control Inflammation in Obese Mice through a Mechanism Involving GLP-2-Driven Improvement of Gut Permeability. Gut 2009, 58, 1091–1103. [Google Scholar] [CrossRef] [PubMed]
- Martínez-López, Y.E.; Esquivel-Hernández, D.A.; Sánchez-Castañeda, J.P.; Neri-Rosario, D.; Guardado-Mendoza, R.; Resendis-Antonio, O. Type 2 Diabetes, Gut Microbiome, and Systems Biology: A Novel Perspective for a New Era. Gut Microbes 2022, 14, 2111952. [Google Scholar] [CrossRef] [PubMed]
- Scheithauer, T.P.M.; Rampanelli, E.; Nieuwdorp, M.; Vallance, B.A.; Verchere, C.B.; van Raalte, D.H.; Herrema, H. Gut Microbiota as a Trigger for Metabolic Inflammation in Obesity and Type 2 Diabetes. Front. Immunol. 2020, 11, 571731. [Google Scholar] [CrossRef] [PubMed]
- Gasaly, N.; Hermoso, M.A.; Gotteland, M. Butyrate and the Fine-Tuning of Colonic Homeostasis: Implication for Inflammatory Bowel Diseases. Int. J. Mol. Sci. 2021, 22, 3061. [Google Scholar] [CrossRef]
- Lagier, J.-C.; Hugon, P.; Khelaifia, S.; Fournier, P.-E.; La Scola, B.; Raoult, D. The Rebirth of Culture in Microbiology through the Example of Culturomics to Study Human Gut Microbiota. Clin. Microbiol. Rev. 2015, 28, 237–264. [Google Scholar] [CrossRef]
- Louis, P.; Flint, H.J. Formation of Propionate and Butyrate by the Human Colonic Microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef]
- Louis, P.; Flint, H.J. Diversity, Metabolism and Microbial Ecology of Butyrate-Producing Bacteria from the Human Large Intestine. FEMS Microbiol. Lett. 2009, 294, 1–8. [Google Scholar] [CrossRef]
- Vital, M.; Howe, A.C.; Tiedje, J.M. Revealing the Bacterial Butyrate Synthesis Pathways by Analyzing (Meta)Genomic Data. mBio 2014, 5, e00889-14. [Google Scholar] [CrossRef]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front. Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef]
- Belzer, C.; Chia, L.W.; Aalvink, S.; Chamlagain, B.; Piironen, V.; Knol, J.; de Vos, W.M. Microbial Metabolic Networks at the Mucus Layer Lead to Diet-Independent Butyrate and Vitamin B12 Production by Intestinal Symbionts. mBio 2017, 8, e00770-17. [Google Scholar] [CrossRef]
- Liu, H.; Wang, J.; He, T.; Becker, S.; Zhang, G.; Li, D.; Ma, X. Butyrate: A Double-Edged Sword for Health? Adv. Nutr. 2018, 9, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Facchin, S.; Vitulo, N.; Calgaro, M.; Buda, A.; Romualdi, C.; Pohl, D.; Perini, B.; Lorenzon, G.; Marinelli, C.; D’Incà, R.; et al. Microbiota Changes Induced by Microencapsulated Sodium Butyrate in Patients with Inflammatory Bowel Disease. Neurogastroenterol. Motil. 2020, 32, e13914. [Google Scholar] [CrossRef] [PubMed]
- Verhaar, B.J.H.; Wijdeveld, M.; Wortelboer, K.; Rampanelli, E.; Levels, J.H.M.; Collard, D.; Cammenga, M.; Nageswaran, V.; Haghikia, A.; Landmesser, U.; et al. Effects of Oral Butyrate on Blood Pressure in Patients with Hypertension: A Randomized, Placebo-Controlled Trial. Hypertension 2024, 81, 2124–2136. [Google Scholar] [CrossRef]
- Lewandowski, K.; Kaniewska, M.; Karłowicz, K.; Rosołowski, M.; Rydzewska, G. The Effectiveness of Microencapsulated Sodium Butyrate at Reducing Symptoms in Patients with Irritable Bowel Syndrome. Przegląd Gastroenterol. 2022, 17, 28–34. [Google Scholar] [CrossRef]
- Panufnik, P.; Wiecek, M.; Szwarc, P.; Kaniewska, M.; Lewandowski, K.; Franek, E.; Rydzewska, G. 610-P: Effect of Butyrate on GI Signs, SIBO, and Diabetes Control—Randomized, Placebo-Controlled Study in Patients with Type 2 Diabetes. Diabetes 2024, 73, 610-P. [Google Scholar] [CrossRef]
- Coppola, S.; Nocerino, R.; Paparo, L.; Bedogni, G.; Calignano, A.; Di Scala, C.; de Giovanni di Santa Severina, A.F.; De Filippis, F.; Ercolini, D.; Berni Canani, R. Therapeutic Effects of Butyrate on Pediatric Obesity. JAMA Netw. Open 2022, 5, e2244912. [Google Scholar] [CrossRef]
- Singh, V.; Lee, G.; Son, H.; Koh, H.; Kim, E.S.; Unno, T.; Shin, J.-H. Butyrate Producers, “The Sentinel of Gut”: Their Intestinal Significance with and beyond Butyrate, and Prospective Use as Microbial Therapeutics. Front. Microbiol. 2023, 13, 1103836. [Google Scholar] [CrossRef]
- Pan, L.-L.; Niu, W.; Fang, X.; Liang, W.; Li, H.; Chen, W.; Zhang, H.; Bhatia, M.; Sun, J. Clostridium Butyricum Strains Suppress Experimental Acute Pancreatitis by Maintaining Intestinal Homeostasis. Mol. Nutr. Food Res. 2019, 63, e1801419. [Google Scholar] [CrossRef]
- Gonçalves, P.; Martel, F. Regulation of Colonic Epithelial Butyrate Transport: Focus on Colorectal Cancer. Porto Biomed. J. 2016, 1, 83–91. [Google Scholar] [CrossRef]
- Li, M.; van Esch, B.C.A.M.; Wagenaar, G.T.M.; Garssen, J.; Folkerts, G.; Henricks, P.A.J. Pro- and Anti-Inflammatory Effects of Short Chain Fatty Acids on Immune and Endothelial Cells. Eur. J. Pharmacol. 2018, 831, 52–59. [Google Scholar] [CrossRef]
- Carretta, M.D.; Quiroga, J.; López, R.; Hidalgo, M.A.; Burgos, R.A. Participation of Short-Chain Fatty Acids and Their Receptors in Gut Inflammation and Colon Cancer. Front. Physiol. 2021, 12, 662739. [Google Scholar] [CrossRef] [PubMed]
- Vagnerová, K.; Hudcovic, T.; Vodička, M.; Ergang, P.; Klusoňová, P.; Petr Hermanová, P.; Šrůtková, D.; Pácha, J. The Effect of Oral Butyrate on Colonic Short-Chain Fatty Acid Transporters and Receptors Depends on Microbial Status. Front. Pharmacol. 2024, 15, 1341333. [Google Scholar] [CrossRef]
- Karaki, S.; Mitsui, R.; Hayashi, H.; Kato, I.; Sugiya, H.; Iwanaga, T.; Furness, J.B.; Kuwahara, A. Short-Chain Fatty Acid Receptor, GPR43, Is Expressed by Enteroendocrine Cells and Mucosal Mast Cells in Rat Intestine. Cell Tissue Res. 2006, 324, 353–360. [Google Scholar] [CrossRef]
- Thangaraju, M.; Cresci, G.A.; Liu, K.; Ananth, S.; Gnanaprakasam, J.P.; Browning, D.D.; Mellinger, J.D.; Smith, S.B.; Digby, G.J.; Lambert, N.A.; et al. GPR109A Is a G-Protein-Coupled Receptor for the Bacterial Fermentation Product Butyrate and Functions as a Tumor Suppressor in Colon. Cancer Res. 2009, 69, 2826–2832. [Google Scholar] [CrossRef]
- Cherbut, C.; Ferrier, L.; Rozé, C.; Anini, Y.; Blottière, H.; Lecannu, G.; Galmiche, J.P. Short-Chain Fatty Acids Modify Colonic Motility through Nerves and Polypeptide YY Release in the Rat. Am. J. Physiol. 1998, 275, G1415–G1422. [Google Scholar] [CrossRef]
- Velázquez, O.C.; Lederer, H.M.; Rombeau, J.L. Butyrate and the Colonocyte. Production, Absorption, Metabolism, and Therapeutic Implications. Adv. Exp. Med. Biol. 1997, 427, 123–134. [Google Scholar]
- Zhu, L.; Han, J.; Li, L.; Wang, Y.; Li, Y.; Zhang, S. Claudin Family Participates in the Pathogenesis of Inflammatory Bowel Diseases and Colitis-Associated Colorectal Cancer. Front. Immunol. 2019, 10, 1441. [Google Scholar] [CrossRef]
- Chen, G.; Ran, X.; Li, B.; Li, Y.; He, D.; Huang, B.; Fu, S.; Liu, J.; Wang, W. Sodium Butyrate Inhibits Inflammation and Maintains Epithelium Barrier Integrity in a TNBS-Induced Inflammatory Bowel Disease Mice Model. eBioMedicine 2018, 30, 317–325. [Google Scholar] [CrossRef]
- Wang, R.X.; Lee, J.S.; Campbell, E.L.; Colgan, S.P. Microbiota-Derived Butyrate Dynamically Regulates Intestinal Homeostasis through Regulation of Actin-Associated Protein Synaptopodin. Proc. Natl. Acad. Sci. USA 2020, 117, 11648–11657. [Google Scholar] [CrossRef]
- Liu, L.; Li, L.; Min, J.; Wang, J.; Wu, H.; Zeng, Y.; Chen, S.; Chu, Z. Butyrate Interferes with the Differentiation and Function of Human Monocyte-Derived Dendritic Cells. Cell. Immunol. 2012, 277, 66–73. [Google Scholar] [CrossRef]
- Lee, C.; Kim, B.G.; Kim, J.H.; Chun, J.; Im, J.P.; Kim, J.S. Sodium Butyrate Inhibits the NF-Kappa B Signaling Pathway and Histone Deacetylation, and Attenuates Experimental Colitis in an IL-10 Independent Manner. Int. Immunopharmacol. 2017, 51, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Skrzydło-Radomańska, B. Application of Butyric Acid Preparations in Clinical Practice. Lekarz POZ 2019, 5, 67–74. [Google Scholar]
- Siddiqui, M.T.; Han, Y.; Shapiro, D.; West, G.; Fiocchi, C.; Cresci, G.A.M. The Postbiotic Butyrate Mitigates Gut Mucosal Disruption Caused by Acute Ethanol Exposure. Int. J. Mol. Sci. 2024, 25, 1665. [Google Scholar] [CrossRef]
- Canani, R.B.; Terrin, G.; Cirillo, P.; Castaldo, G.; Salvatore, F.; Cardillo, G.; Coruzzo, A.; Troncone, R. Butyrate as an Effective Treatment of Congenital Chloride Diarrhea. Gastroenterology 2004, 127, 630–634. [Google Scholar] [CrossRef]
- Malago, J.J.; Sangu, C.L. Intraperitoneal Administration of Butyrate Prevents the Severity of Acetic Acid Colitis in Rats. J. Zhejiang Univ. Sci. B 2015, 16, 224–234. [Google Scholar] [CrossRef]
- Roychowdhury, S.; Glueck, B.; Han, Y.; Mohammad, M.A.; Cresci, G.A.M. A Designer Synbiotic Attenuates Chronic-Binge Ethanol-Induced Gut-Liver Injury in Mice. Nutrients 2019, 11, 97. [Google Scholar] [CrossRef]
- Wang, S.; Yuan, M.; Zhang, L.; Zhu, K.; Sheng, C.; Zhou, F.; Xu, Z.; Liu, Q.; Liu, Y.; Lu, J.; et al. Sodium Butyrate Potentiates Insulin Secretion from Rat Islets at the Expense of Compromised Expression of β Cell Identity Genes. Cell Death Dis. 2022, 13, 67. [Google Scholar] [CrossRef]
- Khan, S.; Jena, G. Sodium Butyrate Reduces Insulin-Resistance, Fat Accumulation and Dyslipidemia in Type-2 Diabetic Rat: A Comparative Study with Metformin. Chem.-Biol. Interact. 2016, 254, 124–134. [Google Scholar] [CrossRef]
- Xu, Y.-H.; Gao, C.-L.; Guo, H.-L.; Zhang, W.-Q.; Huang, W.; Tang, S.-S.; Gan, W.-J.; Xu, Y.; Zhou, H.; Zhu, Q. Sodium butyrate supplementation ameliorates diabetic inflammation in db/db mice. J. Endocrinol. 2018, 238, 231–244. [Google Scholar] [CrossRef]
- Roshanravan, N.; Alamdari, N.M.; Jafarabadi, M.A.; Mohammadi, A.; Shabestari, B.R.; Nasirzadeh, N.; Asghari, S.; Mansoori, B.; Akbarzadeh, M.; Ghavami, A.; et al. Effects of Oral Butyrate and Inulin Supplementation on Inflammation-Induced Pyroptosis Pathway in Type 2 Diabetes: A Randomized, Double-Blind, Placebo-Controlled Trial. Cytokine 2020, 131, 155101. [Google Scholar] [CrossRef]
- Roshanravan, N.; Mahdavi, R.; Jafarabadi, M.A.; Alizadeh, E.; Ghavami, A.; Saadat, Y.R.; Alamdari, N.M.; Dastouri, M.R.; Alipour, S.; Ostadrahimi, A. The Effects of Sodium Butyrate and High-Performance Inulin Supplementation on the Promotion of Gut Bacterium Akkermansia muciniphila Growth and Alterations in miR-375 and KLF5 Expression in Type 2 Diabetic Patients: A Randomized, Double-Blind, Placebo-Controlled Trial. Eur. J. Integr. Med. 2018, 18, 1–7. [Google Scholar] [CrossRef]
- Li, J.; Lin, S.; Vanhoutte, P.M.; Woo, C.W.; Xu, A. Akkermansia Muciniphila Protects Against Atherosclerosis by Preventing Metabolic Endotoxemia-Induced Inflammation in Apoe−/− Mice. Circulation 2016, 133, 2434–2446. [Google Scholar] [CrossRef] [PubMed]
- Khosravi, Z.; Hadi, A.; Tutunchi, H.; Asghari-Jafarabadi, M.; Naeinie, F.; Roshanravan, N.; Ostadrahimi, A.; Fadel, A. The Effects of Butyrate Supplementation on Glycemic Control, Lipid Profile, Blood Pressure, Nitric Oxide Level and Glutathione Peroxidase Activity in Type 2 Diabetic Patients: A Randomized Triple -Blind, Placebo-Controlled Trial. Clin. Nutr. ESPEN 2022, 49, 79–85. [Google Scholar] [CrossRef]
- Hu, Y.; Liu, J.; Yuan, Y.; Chen, J.; Cheng, S.; Wang, H.; Xu, Y. Sodium Butyrate Mitigates Type 2 Diabetes by Inhibiting PERK-CHOP Pathway of Endoplasmic Reticulum Stress. Environ. Toxicol. Pharmacol. 2018, 64, 112–121. [Google Scholar] [CrossRef]
- Roshanravan, N.; Mahdavi, R.; Alizadeh, E.; Jafarabadi, M.A.; Hedayati, M.; Ghavami, A.; Alipour, S.; Alamdari, N.M.; Barati, M.; Ostadrahimi, A. Effect of Butyrate and Inulin Supplementation on Glycemic Status, Lipid Profile and Glucagon-Like Peptide 1 Level in Patients with Type 2 Diabetes: A Randomized Double-Blind, Placebo-Controlled Trial. Horm. Metab. Res. 2017, 49, 886–891. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krauze, W.; Busz, N.; Pikuła, W.; Maternowska, M.; Prowans, P.; Maciejewska-Markiewicz, D. Effect of Sodium Butyrate Supplementation on Type 2 Diabetes—Literature Review. Nutrients 2025, 17, 1753. https://doi.org/10.3390/nu17111753
Krauze W, Busz N, Pikuła W, Maternowska M, Prowans P, Maciejewska-Markiewicz D. Effect of Sodium Butyrate Supplementation on Type 2 Diabetes—Literature Review. Nutrients. 2025; 17(11):1753. https://doi.org/10.3390/nu17111753
Chicago/Turabian StyleKrauze, Wiktoria, Nikola Busz, Weronika Pikuła, Martyna Maternowska, Piotr Prowans, and Dominika Maciejewska-Markiewicz. 2025. "Effect of Sodium Butyrate Supplementation on Type 2 Diabetes—Literature Review" Nutrients 17, no. 11: 1753. https://doi.org/10.3390/nu17111753
APA StyleKrauze, W., Busz, N., Pikuła, W., Maternowska, M., Prowans, P., & Maciejewska-Markiewicz, D. (2025). Effect of Sodium Butyrate Supplementation on Type 2 Diabetes—Literature Review. Nutrients, 17(11), 1753. https://doi.org/10.3390/nu17111753