
Effectiveness of Exogenous Ketone Salts in Enhancing Circulating Acetoacetate Levels—A Pilot Study in Healthy Adults



Highlights
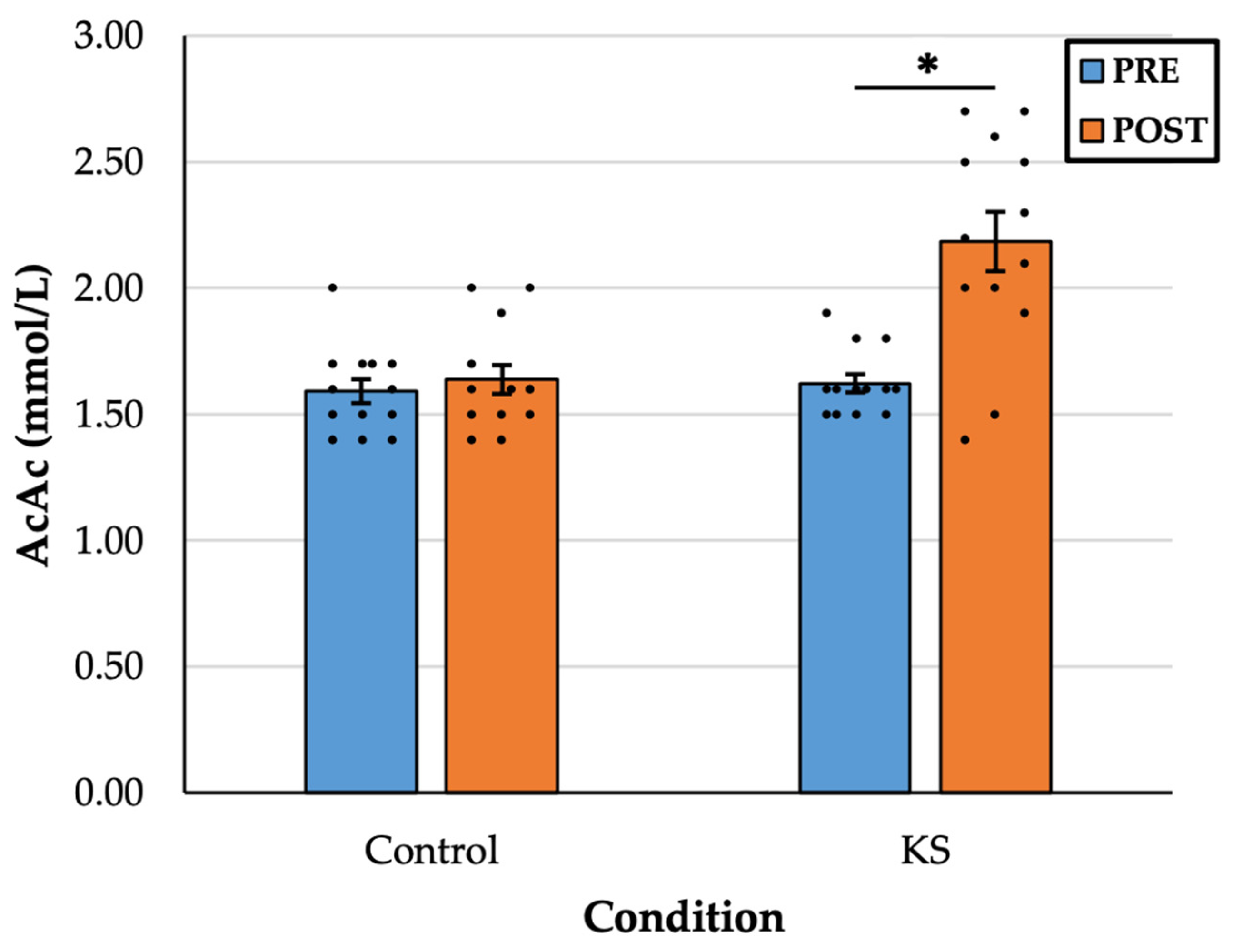
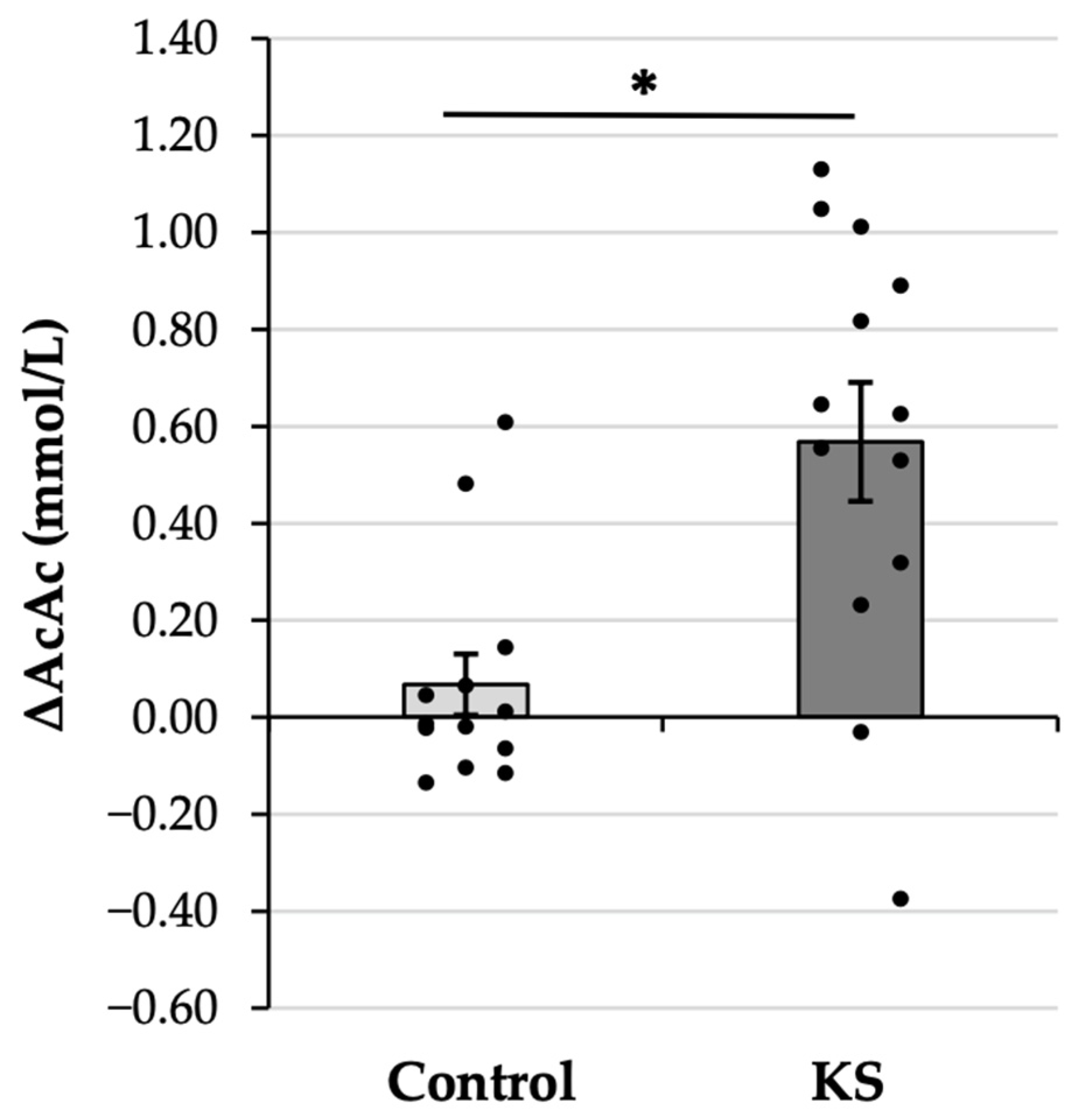
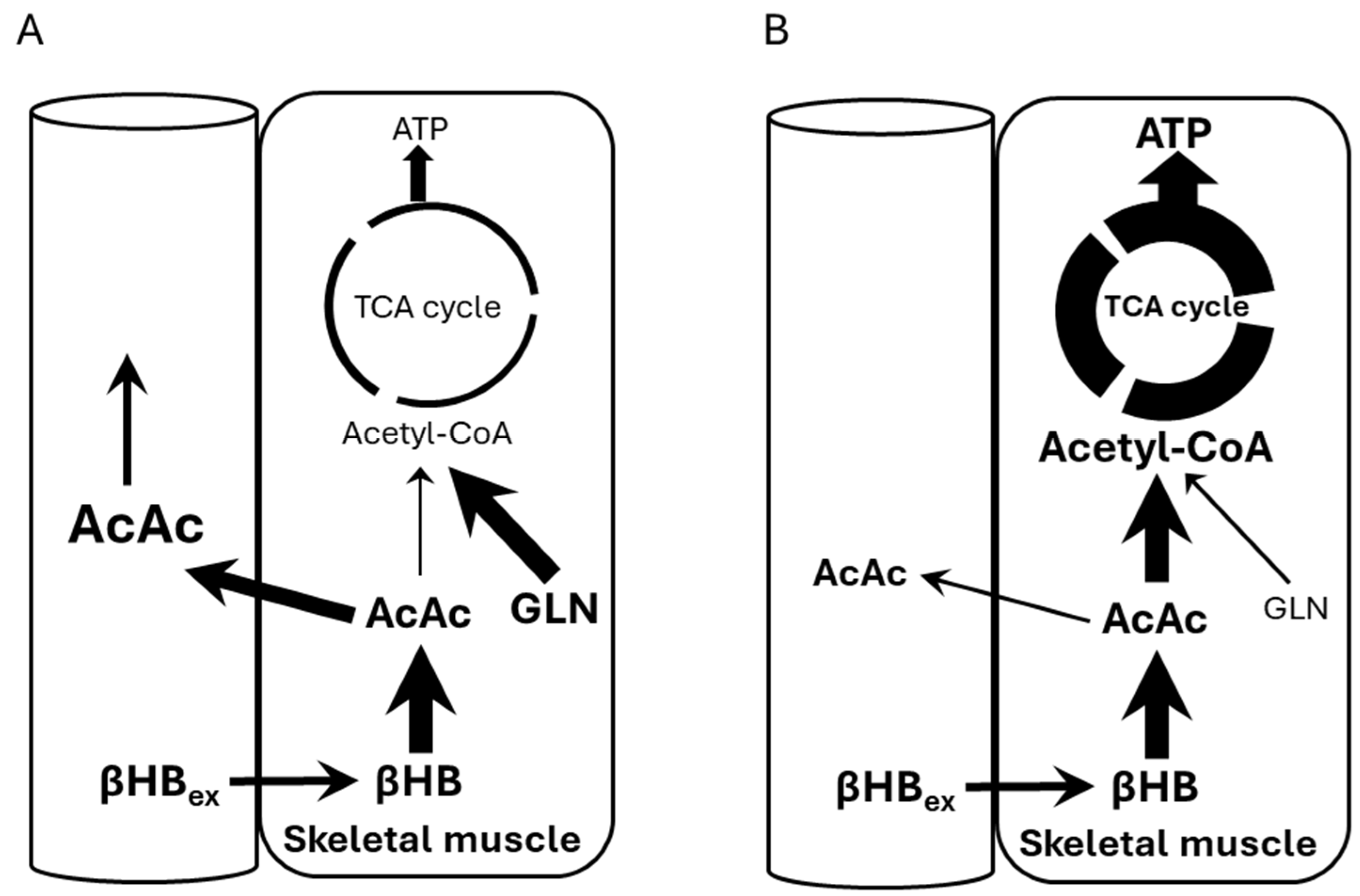
- Consuming ketone salts (KS) significantly increased plasma acetoacetate (AcAc) levels in healthy adults, an effect markedly greater than with a placebo, suggesting conversion from exogenous beta-hydroxybutyrate (βHB).
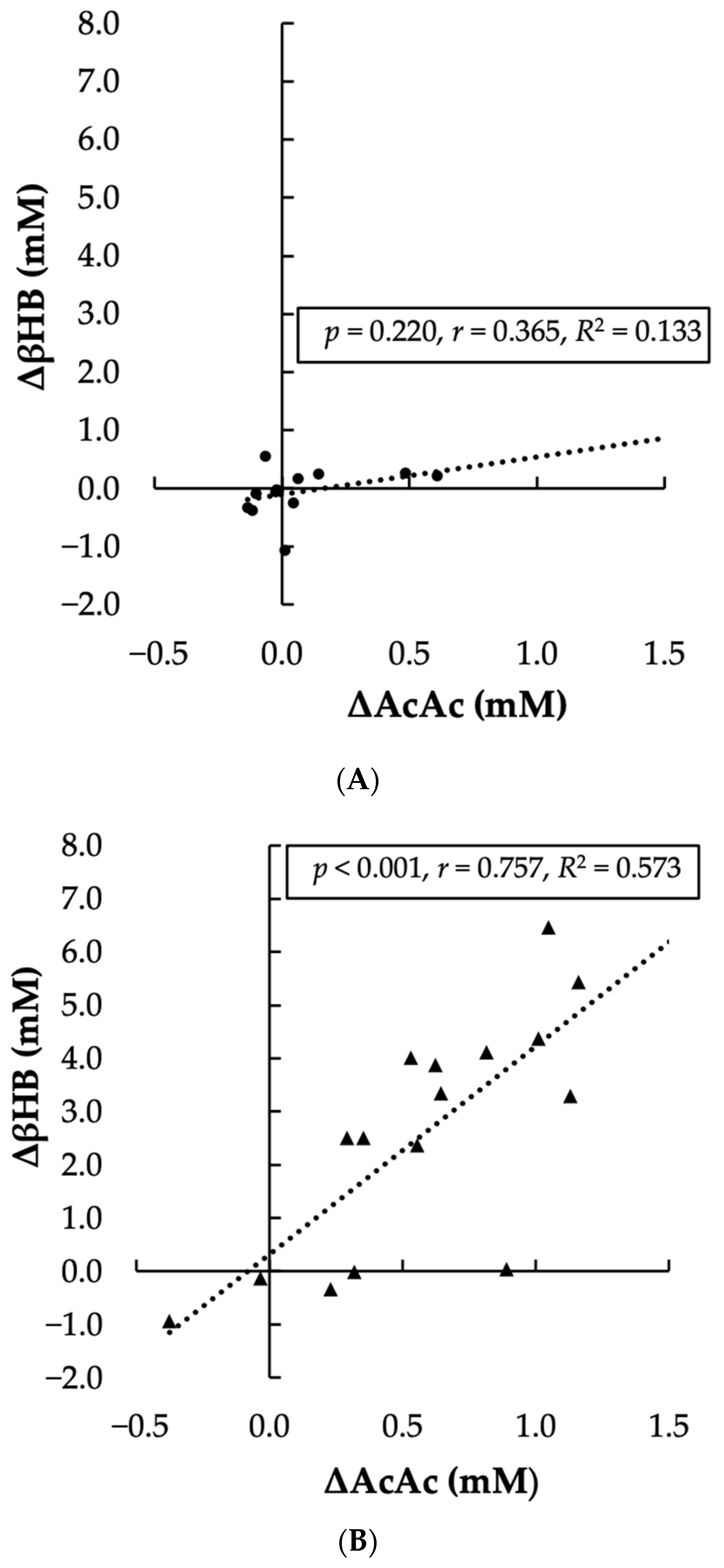
- A significant, positive, and strong correlation was observed between the change in plasma AcAc levels and the change in plasma βHB levels after KS supplementation, supporting the mechanism of conversion.
- A novel and key strength of this study is the use of gas chromatography/mass spectrometry (GC/MS) to accurately quantify plasma AcAc levels.
Abstract
1. Introduction
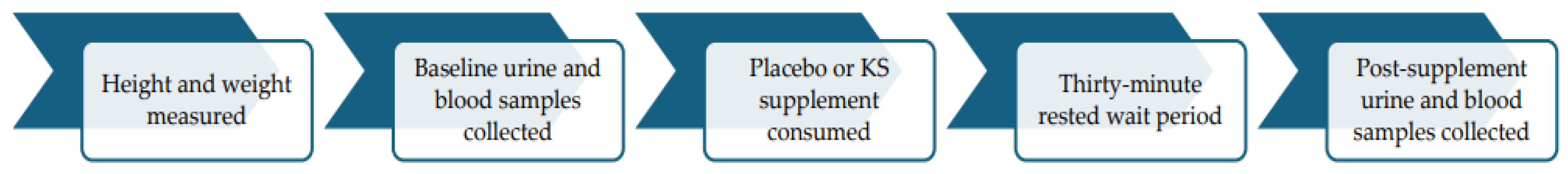
2. Materials and Methods
2.1. Experimental Design
2.2. Participants
2.3. Protocol
2.3.1. Urinary AcAc Measure
2.3.2. Plasma AcAc and βHB Measurements
2.4. Data Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kolb, H.; Kempf, K.; Röhling, M.; Lenzen-Schulte, M.; Schloot, N.C.; Martin, S. Ketone bodies: From enemy to friend and guardian angel. BMC Med. 2021, 19, 313. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Newman, J.C.; Verdin, E. β-Hydroxybutyrate: A Signaling Metabolite. Annu. Rev. Nutr. 2017, 37, 51–76. [Google Scholar] [CrossRef] [PubMed]
- Biesiekierska, M.; Strigini, M.; Śliwińska, A.; Pirola, L.; Balcerczyk, A. The Impact of Ketogenic Nutrition on Obesity and Metabolic Health: Mechanisms and Clinical Implications. Nutr Rev. 2025, nuaf010. [Google Scholar] [CrossRef] [PubMed]
- Murphy, N.E.; Carrigan, C.T.; Margolis, L.M. High-Fat Ketogenic Diets and Physical Performance: A Systematic Review. Adv. Nutr. 2021, 12, 223–233. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Fukao, T.; Lopaschuk, G.D.; Mitchell, G.A. Pathways and control of ketone body metabolism: On the fringe of lipid biochemistry. Prostaglandins Leukot. Essent. Fat. Acids 2004, 70, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Grabacka, M.; Pierzchalska, M.; Dean, M.; Reiss, K. Regulation of Ketone Body Metabolism and the Role of PPARα. Int. J. Mol. Sci. 2016, 17, 2093. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Morris, A.A. Cerebral ketone body metabolism. J. Inherit. Metab. Dis. 2005, 28, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Egan, B.; D’Agostino, D.P. Fueling Performance: Ketones Enter the Mix. Cell Metab. 2016, 24, 373–375. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, D.; Banagalee, V.; Naidoo, R. Exogenous ketone supplement ingestion and submaximal exercise response in trained, male cyclists: A randomised, double-blind study. Ann. Appl. Sport Sci. 2020, 8, S1. [Google Scholar] [CrossRef]
- Falkenhain, K.; Islam, H.; Little, J.P. Exogenous ketone supplementation: An emerging tool for physiologists with potential as a metabolic therapy. Exp. Physiol. 2023, 108, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Poff, A.M.; Koutnik, A.P.; Egan, B. Nutritional Ketosis with Ketogenic Diets or Exogenous Ketones: Features, Convergence, and Divergence. Curr. Sports Med. Rep. 2020, 19, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Walsh, J.J.; Myette-Côté, É.; Neudorf, H.; Little, J.P. Potential Therapeutic Effects of Exogenous Ketone Supplementation for Type 2 Diabetes: A Review. Curr. Pharm. Des. 2020, 26, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Newport, M.T.; VanItallie, T.B.; Kashiwaya, Y.; King, M.T.; Veech, R.L. A new way to produce hyperketonemia: Use of ketone ester in a case of Alzheimer’s disease. Alzheimers Dement. 2015, 11, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Zhang, Y.; Zhang, X.; Liu, L.; Zhou, B.; Mo, R.; Li, Y.; Li, H.; Li, F.; Tao, Y.; et al. Medium-chain triglycerides improved cognition and lipid metabolomics in mild to moderate Alzheimer’s disease patients with APOE4-/-: A double-blind, randomized, placebo-controlled crossover trial. Clin. Nutr. 2020, 39, 2092–2105. [Google Scholar] [CrossRef] [PubMed]
- Cox, P.J.; Kirk, T.; Ashmore, T.; Willerton, K.; Evans, R.; Smith, A.; Murray, A.J.; Stubbs, B.; West, J.; McLure, S.W.; et al. Nutritional Ketosis Alters Fuel Preference and Thereby Endurance Performance in Athletes. Cell Metab. 2016, 24, 256–268. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, H.; Geerts, C.; Furtos, A.; Waters, P.; Cyr, D.; Wang, S.; Mitchell, G.A. The multiple facets of acetyl-CoA metabolism: Energetics, biosynthesis, regulation, acylation and inborn errors. Mol. Genet. Metab. 2023, 138, 106966. [Google Scholar] [CrossRef] [PubMed]
- Taboulet, P.; Deconinck, N.; Thurel, A.; Haas, L.; Manamani, J.; Porcher, R.; Schmit, C.; Fontaine, J.P.; Gautier, J.F. Correlation between urine ketones (acetoacetate) and capillary blood ketones (3-beta-hydroxybutyrate) in hyperglycaemic patients. Diabetes Metab. 2007, 33, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Klocker, A.A.; Phelan, H.; Twigg, S.M.; Craig, M.E. Blood β-hydroxybutyrate vs. urine acetoacetate testing for the prevention and management of ketoacidosis in Type 1 diabetes: A systematic review. Diabet. Med. 2013, 30, 818–824. [Google Scholar] [CrossRef] [PubMed]
- Leckey, J.J.; Ross, M.L.; Quod, M.; Hawley, J.A.; Burke, L.M. Ketone Diester Ingestion Impairs Time-Trial Performance in Professional Cyclists. Front. Physiol. 2017, 8, 806. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- McCarthy, D.G.; Bostad, W.; Powley, F.J.; Little, J.P.; Richards, D.L.; Gibala, M.J. Increased cardiorespiratory stress during submaximal cycling after ketone monoester ingestion in endurance-trained adults. Appl. Physiol. Nutr. Metab. 2021, 46, 986–993. [Google Scholar] [CrossRef] [PubMed]
- Fischer, T.; Och, U.; Klawon, I.; Och, T.; Grüneberg, M.; Fobker, M.; Bordewick-Dell, U.; Marquardt, T. Effect of a Sodium and Calcium DL-β-Hydroxybutyrate Salt in Healthy Adults. J. Nutr. Metab. 2018, 2018, 9812806. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bolyard, M.L.; Graziano, C.M.; Fontaine, K.R.; Sayer, R.D.; Fisher, G.; Plaisance, E.P. Tolerability and Acceptability of an Exogenous Ketone Monoester and Ketone Monoester/Salt Formulation in Humans. Nutrients 2023, 15, 4876. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences; Routledge: London, UK, 2013. [Google Scholar]
- Blanca, M.J.; Alarcón, R.; Arnau, J.; Bono, R.; Bendayan, R. Non-normal data: Is ANOVA still a valid option? Psicothema 2017, 29, 552–557. [Google Scholar] [CrossRef] [PubMed]
- Field, A. Discovering Statistics Using SPSS for Windows; Sage: Thousand Oaks, CA, USA, 2013. [Google Scholar]
- Stubbs, B.J.; Cox, P.J.; Evans, R.D.; Santer, P.; Miller, J.J.; Faull, O.K.; Magor-Elliott, S.; Hiyama, S.; Stirling, M.; Clarke, K. On the Metabolism of Exogenous Ketones in Humans. Front. Physiol. 2017, 8, 848. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Stubbs, B.J.; Cox, P.J.; Kirk, T.; Evans, R.D.; Clarke, K. Gastrointestinal Effects of Exogenous Ketone Drinks are Infrequent, Mild, and Vary According to Ketone Compound and Dose. Int. J. Sport Nutr. Exerc. Metab. 2019, 29, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Cuenoud, B.; Hartweg, M.; Godin, J.P.; Croteau, E.; Maltais, M.; Castellano, C.A.; Carpentier, A.C.; Cunnane, S.C. Metabolism of Exogenous D-Beta-Hydroxybutyrate, an Energy Substrate Avidly Consumed by the Heart and Kidney. Front. Nutr. 2020, 7, 13. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Evans, M.; Cogan, K.E.; Egan, B. Metabolism of ketone bodies during exercise and training: Physiological basis for exogenous supplementation. J. Physiol. 2017, 595, 2857–2871. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Vice, E.; Privette, J.D.; Hickner, R.C.; Barakat, H.A. Ketone body metabolism in lean and obese women. Metabolism 2005, 54, 1542–1545. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Men (n = 7) | Women (n = 6) | Total (N = 13) |
|---|---|---|---|
| Age (years) | 22.6 ± 5.7 | 20.5 ± 1.5 | 21.6 ± 4.3 |
| Height (cm) | 186.5 ± 11.6 | 168.4 ± 6.9 | 178.2 ± 13.2 |
| Weight (kg) | 83.7 ± 13.7 | 77.1 ± 21.4 | 80.6 ± 17.2 |
| BMI (kg/m2) | 24.4 ± 5.9 | 27.2 ± 7.2 | 25.7 ± 6.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holland-Winkler, A.M.; Moore, A.R.; Bederman, I. Effectiveness of Exogenous Ketone Salts in Enhancing Circulating Acetoacetate Levels—A Pilot Study in Healthy Adults. Nutrients 2025, 17, 1665. https://doi.org/10.3390/nu17101665
Holland-Winkler AM, Moore AR, Bederman I. Effectiveness of Exogenous Ketone Salts in Enhancing Circulating Acetoacetate Levels—A Pilot Study in Healthy Adults. Nutrients. 2025; 17(10):1665. https://doi.org/10.3390/nu17101665
Chicago/Turabian StyleHolland-Winkler, A. Maleah, Andrew R. Moore, and Ilya Bederman. 2025. "Effectiveness of Exogenous Ketone Salts in Enhancing Circulating Acetoacetate Levels—A Pilot Study in Healthy Adults" Nutrients 17, no. 10: 1665. https://doi.org/10.3390/nu17101665
APA StyleHolland-Winkler, A. M., Moore, A. R., & Bederman, I. (2025). Effectiveness of Exogenous Ketone Salts in Enhancing Circulating Acetoacetate Levels—A Pilot Study in Healthy Adults. Nutrients, 17(10), 1665. https://doi.org/10.3390/nu17101665