
Comprehensive Review of Functional and Nutraceutical Properties of Craterellus cornucopioides (L.) Pers.


 ,
,  ,
,  , ,
, ,  and
and Highlights
- This review focuses on the wild mushroom species Craterellus cornucopioides (L.) Pers. (Cantharellaceae) and presents its morphology, chemical profile, and medicinal po-tential.
- Also known as the black trumpet, it is renowned for its excellent versatility and unique taste.
- Beyond its gastronomic value, recent scientific data have documented its nutraceuti-cal potential as a source of bioactive compounds with pharmaceutical and therapeu-tic significance.
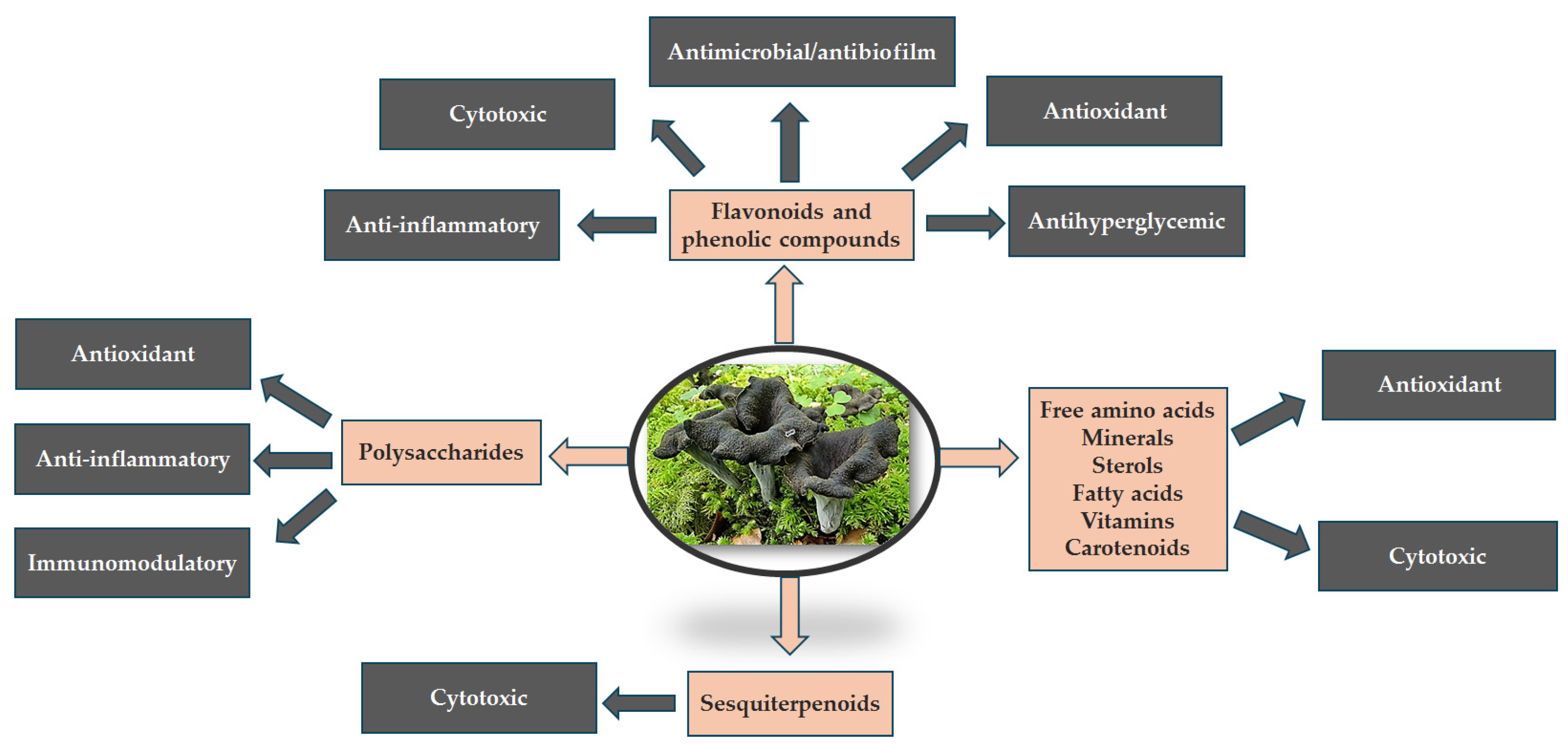
- The present study emphasizes both the distinct attributes of this edible mushroom re-lated to the nutritional value supported by macro- and micronutrients (glucides, pro-teins, amino acids, fatty acids, vitamins, minerals) as well as the functional properties (immunomodulatory, anti-inflammatory, antimutagenic, cytotoxic, antimicrobial properties) provided by the biologically active compounds from the classes of poly-phenols (flavonoids, phenolic acids), terpenoids (sesquiterpenoids, sterols), and pol-ysaccharides.
Abstract
1. Introduction
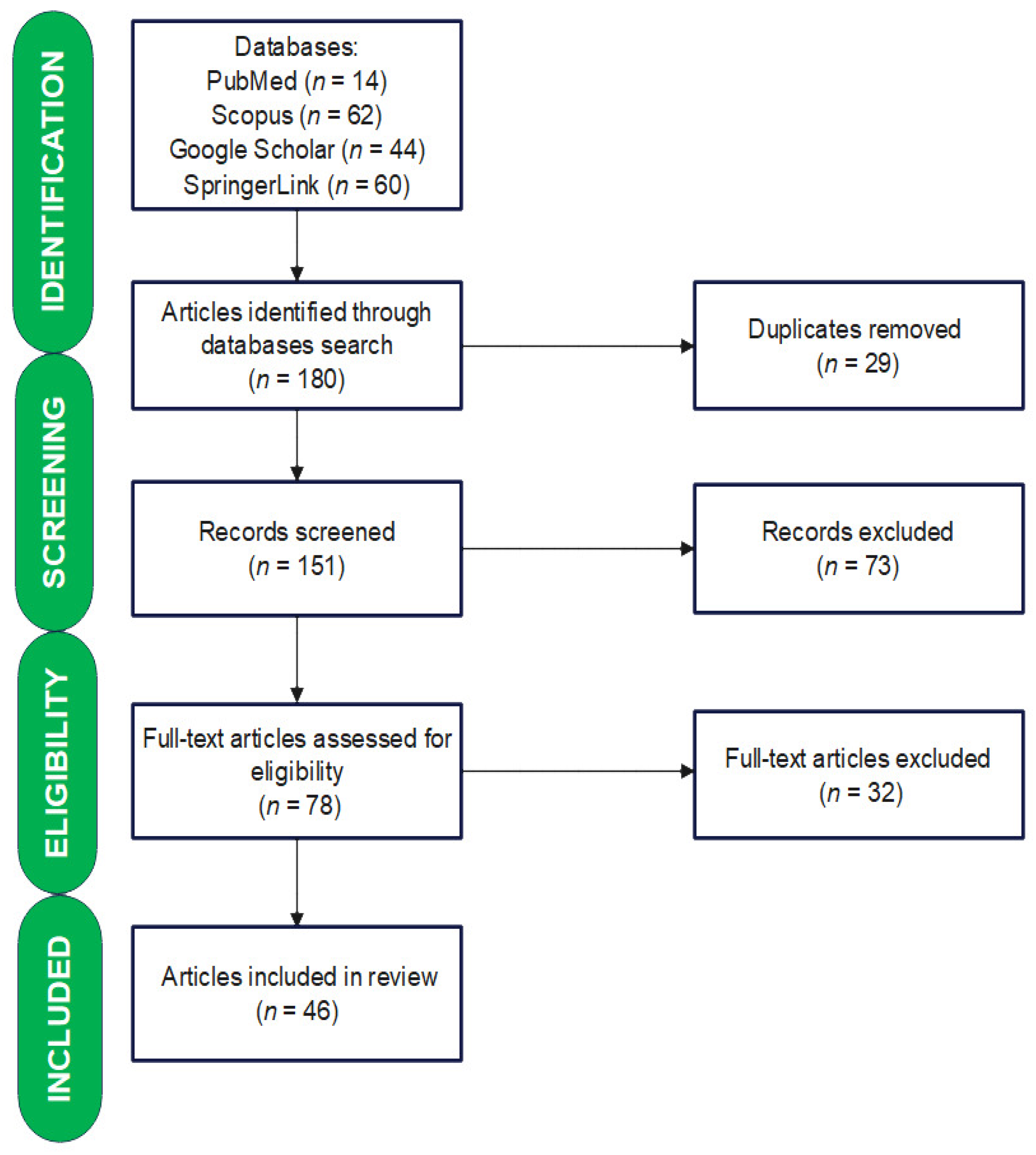
2. Materials and Methods
3. Results
3.1. Chemical Composition
3.2. Functional Activities
3.2.1. Immunomodulatory Activity
3.2.2. Anti-Inflammatory Activity
3.2.3. Antimutagenic Effects

{kind=link}
{kind=link}
{kind=link}
| Functional Activity | Type of Study | Mechanism (s) | Extracts or Tested Vegetal Products | Reference |
|---|---|---|---|---|
| Antimicrobial | In vitro | Biofilm inhibition | Extracts | [43] |
| In vitro | Inhibitory activity against reference strains of Staphylococcus aureus, B. subtilis, B. cereus, Escherichia coli, Proteus mirabilis, Aspergillus niger, Candida albicans, Penicillium italicum, Mucor mucedo, Trichoderma viride | Acetone extract | [8] | |
| In vitro | Antibacterial activity on all tested microorganisms—reference strains of B. subtilis, E. faecalis, B. licheniformis, A. tumefaciens, E. coli, S. aureus | Water and methanol extracts | [19] | |
| Anti-inflammatory | ||||
| Immunomodulatory | In vitro | RAW264.7 cell activation with enhanced phagocytosis | Isolated polysaccharides | [28] |
| In vivo mice models | Enhanced immunoregulatory activity Significantly increased spleen and thymus weight indices of the BALB/c mice models Synergistic effects on the T- or B-lymphocyte proliferation induced by ConA or LPS, respectively Enhanced natural killer (NK) cell activity Significantly increased phagocytic activity of peritoneal macrophages in immunosuppressive mice | Isolated polysaccharides | [27] | |
| In vivo | Protective function against immunosuppression induced by cyclophosphamide Immunoregulatory activity in immunosuppression BALB/c mice model Macrophage activation via enhanced production of cytokines (IL-2, IL-6, and IL-8) | Isolated polysaccharides | [28] | |
| Antioxidant | In vivo simulated digestion model | Reduced DPPH-radical-scavenging activity, reducing power, and metal chelating activity due to the processing methods | Processed products (steaming, boiling, frying, and microwaving) | [17] |
| Free radical scavenging, superoxide anion scavenging, and reducing power | Acetone extract | [8] | ||
| In vitro | Strong scavenging abilities on DPPH and ABTS radicals. Oxidative hemolysis induced by AAPH in mice erythrocytes was effectively reversed by incubation with CCPP-1 Protective effect against AAPH-induced oxidative stress in erythrocytes | Polysaccharide fraction (CCPP-1) | [13] | |
| In vitro | Free-radical-scavenging effect | Extracts | [43] | |
| In vitro | DPPH-radical-scavenging activities enhanced with elevated concentrations (methanolic extracts > water extracts) | Water and methanol extracts | [15,19] | |
| Cytotoxic | In vitro | Against MCF-7 breast cancer cell line | Extracts | [43] |
| In vitro | Moderate cytotoxic effect on cancer cell lines (human epithelial carcinoma HeLa, human lung carcinoma A549, human colon carcinoma LS174); HeLa cell was most sensitive No cytotoxic effect on normal cells | Acetone extract | [8] | |
| In vitro | Some degree of cytotoxicity over HepG2 cell line Cytotoxic effects of the extracts increased with elevated concentrations Methanolic extracts had the lowest IC50 values, and thus, methanolic extracts > water extracts | Water and methanol extracts | [19] | |
| In vitro | Cyclohexane and dichloromethane extracts expressed significant cytotoxic activity against human epithelial cervical cancer cells (HeLa), adenocarcinomic human alveolar basal epithelial cells (A549), colorectal cancer cells (LS174), and normal MRC-5 human embryonic lung fibroblast cells | Cyclohexane and dichloromethane extracts | [15] | |
| Angiotensin-converting enzyme (ACE)-inhibitory | In vitro | Strong and dose-dependent ACE-inhibitory activity found only for the aqueous extract (IC50 = 0.74 μg/mL) | Aqueous and methanol extracts | [15] |
3.2.4. Antioxidant Activity
3.2.5. Cytotoxic Activity
3.2.6. Antimicrobial Activity
4. Discussion
5. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Badalyan, S.M.; Barkhudaryan, A.; Rapior, S. Recent Progress in Research on the Pharmacological Potential of Mushrooms and Prospects for Their Clinical Application; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar] [CrossRef]
- Kaliyaperumal, M.; Kezo, K.; Gunaseelan, S. A Global Overview of Edible Mushrooms. In Biology of Macrofungi, Fungal Biology; Singh, B.P., Ed.; Springer Nature: Cham, Switzerland, 2018; pp. 15–56. [Google Scholar] [CrossRef]
- Chatterjee, S.; Sarma, M.K.; Deb, U.; Steinhauser, G.; Walther, C.; Gupta, D.K. Mushrooms: From Nutrition to Mycoremediation. Environ. Sci. Pollut. Res. 2017, 24, 19480–19493. [Google Scholar] [CrossRef]
- Dai, X.; Sharma, M.; Chen, J. (Eds.) Fungi in Sustainable Food Production; Springer Nature: Cham, Switzerland, 2016. [Google Scholar]
- Beluhan, S.; Ranogajec, A. Chemical Composition and Non-Volatile Components of Croatian Wild Edible Mushrooms. Food Chem. 2011, 124, 1076–1082. [Google Scholar] [CrossRef]
- Butkhup, L.; Samappito, W.; Jorjong, S. Evaluation of Bioactivities and Phenolic Contents of Wild Edible Mushrooms from Northeastern Thailand. Food Sci. Biotechnol. 2018, 27, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Egbuna, C.; Dable-Tupas, G. Functional Foods and Nutraceuticals—Bioactive Components, Formulations and Innovations; Springer Nature: Cham, Switzerland, 2024. [Google Scholar] [CrossRef]
- Kosanić, M.; Ranković, B.; Stanojković, T.; Radović-Jakovljević, M.; Ćirić, A.; Grujičić, D.; Milošević-Djordjević, O. Craterellus Cornucopioides Edible Mushroom as Source of Biologically Active Compounds. Nat. Prod. Commun. 2019, 14, 1934578X19843610. [Google Scholar] [CrossRef]
- Boruah, T.A.; Parashar, P.B.; Devi, H.D.; Nath, A.K.E.; Barman, A.F.; Barman, H. Craterellus cornucopioides (L.)/Craterellus odoratus (Schwein.) Fr. (Black Chanterelle). In Phytochemistry and Nutritional Composition of Significant Wild Medicinal and Edible Mushrooms: Traditional Uses and Pharmacology; Sharma, A., Bhardwaj, G., Nayik, G.A., Eds.; Royal Society of Chemistry: London, UK, 2023; pp. 142–164. [Google Scholar]
- Læssøe, T.; Petersen, J.H. Fungi of Temperate Europe; Princeton University Press: Princeton, NJ, USA, 2019; Volume 1. [Google Scholar]
- Costea, T.; Hudiţă, A.; Olaru, O.T.; Gălăţeanu, B.; Gîrd, C.E.; Mocanu, M.M. Chemical Composition, Antioxidant Activity and Cytotoxic Effects of Romanian Craterellus cornucopioides (L) Pers. Mushroom. Farmacia 2020, 68, 340–347. [Google Scholar] [CrossRef]
- Ozuna-Valencia, K.H.; Moreno-Robles, A.L.; Rodriguez-Felix, F.; Moreno-Vasquez, M.J.; Barreras-Urbina, C.G.; Madera-Santana, T.J.; Ruiz-Cruz, S.; Bernal-Mercado, A.-T.; Armenta-Villegas, L.; Tapia-Hernandez, J.A. Black Trumpet (Craterellus cornucopioides). In Mushrooms: Nutraceuticals and Functional Foods; Pandita, D., Pandita, A., Eds.; CRC Press: Boca Raton, FL, USA, 2023. [Google Scholar]
- Yang, W.W.; Wang, L.M.; Gong, L.L.; Lu, Y.M.; Pan, W.J.; Wang, Y.; Zhang, W.N.; Chen, Y. Structural Characterization and Antioxidant Activities of a Novel Polysaccharide Fraction from the Fruiting Bodies of Craterellus Cornucopioides. Int. J. Biol. Macromol. 2018, 117, 473–482. [Google Scholar] [CrossRef]
- Guo, M.Z.; Meng, M.; Duan, S.Q.; Feng, C.C.; Wang, C.L. Structure Characterization, Physicochemical Property and Immunomodulatory Activity on RAW264.7 Cells of a Novel Triple-Helix Polysaccharide from Craterellus cornucopioides. Int. J. Biol. Macromol. 2019, 126, 796–804. [Google Scholar] [CrossRef]
- Radović, J.; Leković, A.; Tačić, A.; Dodevska, M.; Stanojković, T.; Marinković, T.; Jelić, Č.; Kundaković-Vasović, T. Black Trumpet, Craterellus cornucopioides (L.) Pers.: Culinary Mushroom with Angiotensin Converting Enzyme Inhibitory and Cytotoxic Activity. Polish J. Food Nutr. Sci. 2022, 72, 171–181. [Google Scholar] [CrossRef]
- Savoie, J.M.; Largeteau, M.L. Production of Edible Mushrooms in Forests: Trends in Development of a Mycosilviculture. Appl. Microbiol. Biotechnol. 2011, 89, 971–979. [Google Scholar] [CrossRef]
- Liu, Y.; Duan, X.; Zhang, M.; Li, C.; Zhang, Z.; Liu, A.; Hu, B.; He, J.; Wu, D.; Chen, H.; et al. Cooking Methods Effect on the Nutrients, Bioaccessibility and Antioxidant Activity of Craterellus cornucopioides. Lwt 2020, 131, 109768. [Google Scholar] [CrossRef]
- Guo, M.; Meng, M.; Zhao, J.; Wang, X.; Wang, C. Immunomodulatory Effects of the Polysaccharide from Craterellus cornucopioides via Activating the TLR4-NFκB Signaling Pathway in Peritoneal Macrophages of BALB/c Mice. Int. J. Biol. Macromol. 2020, 160, 871–879. [Google Scholar] [CrossRef]
- Kol, S.; Bostanci, A.; Kocabaş, A.; Uzun, Y.; Sadi, G. Cell Growth Inhibitory Potential of Craterellus cornucopioides (L.) Pers. Together with Antioxidant and Antimicrobial Properties. Anatol. J. Bot. 2018, 2, 60–64. [Google Scholar] [CrossRef]
- Palacios, I.; Lozano, M.; Moro, C.; D’Arrigo, M.; Rostagno, M.A.; Martínez, J.A.; García-Lafuente, A.; Guillamón, E.; Villares, A. Antioxidant Properties of Phenolic Compounds Occurring in Edible Mushrooms. Food Chem. 2011, 128, 674–678. [Google Scholar] [CrossRef]
- Liu, Y.T.; Sun, J.; Luo, Z.Y.; Rao, S.Q.; Su, Y.J.; Xu, R.R.; Yang, Y.J. Chemical Composition of Five Wild Edible Mushrooms Collected from Southwest China and Their Antihyperglycemic and Antioxidant Activity. Food Chem. Toxicol. 2012, 50, 1238–1244. [Google Scholar] [CrossRef] [PubMed]
- Vamanu, E.; Nita, S. Biological Activity of Fluidized Bed Ethanol Extracts from Several Edible Mushrooms. Food Sci. Biotechnol. 2014, 23, 1483–1490. [Google Scholar] [CrossRef]
- Vasdekis, E.P.; Karkabounas, A.; Giannakopoulos, I.; Savvas, D.; Lekka, M.E. Screening of Mushrooms Bioactivity: Piceatannol Was Identified as a Bioactive Ingredient in the Order Cantharellales. Eur. Food Res. Technol. 2018, 244, 861–871. [Google Scholar] [CrossRef]
- Liu, Y.; Duan, X.; Zhang, M.; Li, C.; Zhang, Z.; Hu, B.; Liu, A.; Li, Q.; Chen, H.; Tang, Z.; et al. Extraction, Structure Characterization, Carboxymethylation and Antioxidant Activity of Acidic Polysaccharides from Craterellus cornucopioides. Ind. Crops Prod. 2021, 159, 113079. [Google Scholar] [CrossRef]
- Liu, Y.; Duan, X.; Duan, S.; Li, C.; Hu, B.; Liu, A.; Wu, Y.; Wu, H.; Chen, H.; Wu, W.; et al. Effects of In Vitro Digestion and Fecal Fermentation on the Stability and Metabolic Behavior of Polysaccharides from Craterellus cornucopioides. Food Funct. 2020, 11, 6899–6910. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.J.; Gong, L.L.; Li, Y.Y.; Zhou, Z.B.; Yang, W.W.; Wan, C.X.; Zhang, W.N. Anti-Inflammatory Effect of a Polysaccharide Fraction from Craterellus cornucopioides in LPS-Stimulated Macrophages. J. Food Biochem. 2021, 45, e13842. [Google Scholar] [CrossRef]
- Guo, M.Z.; Meng, M.; Feng, C.C.; Wang, X.; Wang, C.L. A Novel Polysaccharide Obtained from Craterellus cornucopioides Enhances Immunomodulatory Activity in Immunosuppressive Mice Models via Regulation of the TLR4-NF-ΚB Pathway. Food Funct. 2019, 10, 4792–4801. [Google Scholar] [CrossRef]
- Zhang, C.; Shu, Y.; Li, Y.; Guo, M. Extraction and Immunomodulatory Activity of the Polysaccharide Obtained from Craterellus cornucopioides. Front. Nutr. 2022, 9, 1017431. [Google Scholar] [CrossRef]
- Guo, H.; Diao, Q.P.; Hou, D.Y.; Li, Z.H.; Zhou, Z.Y.; Feng, T.; Liu, J.K. Sesquiterpenoids from Cultures of the Edible Mushroom Craterellus cornucopioides. Phytochem. Lett. 2017, 21, 114–117. [Google Scholar] [CrossRef]
- Guo, H.; Diao, Q.P.; Zhang, B.; Feng, T. Two New Illudane Sesquiterpenoids and One New Menthane Monoterpene from Cultures of Craterellus cornucopioides. J. Asian Nat. Prod. Res. 2019, 21, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Zhou, Z.Y.; Liu, J.K. Three New Keto Esters from Cultures of the Basidiomycete Craterellus cornucopioides. Chin. J. Nat. Med. 2010, 8, 88–90. [Google Scholar] [CrossRef]
- Magnus, V.; Laćan, G.; Aplin, R.T.; Thaller, V. Glycerol Tridehydrocrepenynate from the Basidiomycete Craterellus cornucopioides. Phytochemistry 1989, 28, 3047–3050. [Google Scholar] [CrossRef]
- Watanabe, F.; Schwarz, J.; Takenaka, S.; Miyamoto, E.; Ohishi, N.; Nelle, E.; Hochstrasser, R.; Yabuta, Y. Characterization of Vitamin B12 Compounds in the Wild Edible Mushrooms Black Trumpet (Craterellus cornucopioides) and Golden Chanterelle (Cantharellus cibarius). J. Nutr. Sci. Vitaminol. 2012, 58, 438–441. [Google Scholar] [CrossRef] [PubMed]
- Dospatliev, L.; Petkova, Z.; Antova, G.; Angelova-Romova, M.; Ivanova, M.; Mustafa, S. Proximate Composition of Wild Edible Mushrooms from the Batak Mountain, Bulgaria. J. Microbiol. Biotechnol. Food Sci. 2023, 12, e4718. [Google Scholar] [CrossRef]
- Villares, A.; Mateo-Vivaracho, L.; García-Lafuente, A.; Guillamón, E. Storage Temperature and UV-Irradiation Influence on the Ergosterol Content in Edible Mushrooms. Food Chem. 2014, 147, 252–256. [Google Scholar] [CrossRef]
- Sesli, E.; Tuzen, M.; Soylak, M. Evaluation of Trace Metal Contents of Some Wild Edible Mushrooms from Black Sea Region, Turkey. J. Hazard. Mater. 2008, 160, 462–467. [Google Scholar] [CrossRef]
- Sarikurkcu, C.; Akata, I.; Guven, G.; Tepe, B. Metal Concentration and Health Risk Assessment of Wild Mushrooms Collected from the Black Sea Region of Turkey. Environ. Sci. Pollut. Res. 2020, 27, 26419–26441. [Google Scholar] [CrossRef]
- Vetter, J. Lithium Content of Some Common Edible Wild-Growing Mushrooms. Food Chem. 2005, 90, 31–37. [Google Scholar] [CrossRef]
- Ouali, Z.; Chaar, H.; Venturella, G.; Cirlincione, F.; Gargano, M.L.; Jaouani, A. Chemical Composition and Nutritional Value of Nine Wild Edible Mushrooms from Northwestern Tunisia. Ital. J. Mycol. 2023, 52, 32–49. [Google Scholar] [CrossRef]
- O’Callaghan, Y.C.; O’Brien, N.M.; Kenny, O.; Harrington, T.; Brunton, N.; Smyth, T.J. Anti-Inflammatory Effects of Wild Irish Mushroom Extracts in RAW264.7 Mouse Macrophage Cells. J. Med. Food 2015, 18, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Grüter, A.; Friederich, U.; Würgler, F.E. Antimutagenic Effects of Mushrooms. Mutat. Res. Fundam. Mol. Mech. Mutagen. 1990, 231, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, D.; Sharma, A.; Talukder, G. Plant Extracts as Modulators of Genotoxic Effects. Bot. Rev. 1996, 62, 275–300. [Google Scholar] [CrossRef]
- Dinçer, E.; Işık, H.; Hepokur, C.; Tutar, U.; Çelik, C. Cytotoxic, Antioxidant, Antibiofilm, and Antimicrobial Activities of Mushroom Species from Turkey. Int. J. Med. Mushrooms 2023, 25, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, M.; Dospatliev, L.; Papazov, P. Application of ICP-OES Method of Determination of K, P, Mg, Na and Ca in Nine Wild Edible Mushrooms with a Chemometric Approach. Comptes Rendus L’Acad. Bulg. Sci. 2019, 72, 182–190. [Google Scholar] [CrossRef]
- Costa-Silva, F.; Marques, G.; Matos, C.C.; Barros, A.I.R.N.A.; Nunes, F.M. Selenium Contents of Portuguese Commercial and Wild Edible Mushrooms. Food Chem. 2011, 126, 91–96. [Google Scholar] [CrossRef]
- Sánchez, C. Bioactives from Mushroom and Their Application. In Food Bioactives: Extraction and Biotechnology Applications; Springer International Publishing: Cham, Switzerland, 2017; pp. 23–57. [Google Scholar] [CrossRef]



| Class of Compounds | Metabolite (s) | Extract/Vegetal Product | Reference |
|---|---|---|---|
| Flavonoids and phenolic compounds | Ferulic acid, Gallic acid, p-hydroxybenzoic acid, Protocatechuic acid, Chlorogenic acid, Caffeic acid, Syringic acid, Rosmarinic acid Pyrogallol, Catechin, Epicatechin, Epigallocatechin gallate Rutin, Quercetin, t-resveratrol, piceatannol | Methanolic extract Ethanolic extract Aqueous extract | [8,11,15,19,20,21,22,23] |
| Polysaccharides | CCPs-1, CCPs-2 (acidic polysaccharides with different contents of uronic acid, rhamnose, fucose, arabinose, xylose, mannose, glucose, and galactose) CCPP-1 (a water-soluble polysaccharide, a heteropolysaccharide consisting of mannose, glucose, xylose, arabinose, and fructose) | Aqueous extract after removal of pigments and lipids with ethanol | [13,14,18,24,25,26,27,28] |
| Sesquiterpenoids | Craterellins A–C Gymnomitr-3-en-10β,15-diol Illudin F, illudin M, illudin T Illudalenol | Cultures of the mushroom Ethyl acetate extract | [29,30] |
| Keto esters | 4-oxohex-1,6-diyl diacetate, 4-oxohex-5-enyl acetate, 6-hydroxy-4-oxohexyl acetate | Ethyl acetate extract | [31] |
| Triacylglycerol | Glycerol tri-dehydrocrepenynate (octadeca-9Z, 14Z-dien-12-ynoate) | Chloroforme + methanol extract after removal of lipases with isopropanol | [32] |
| Water soluble vitamins | Vitamins C, B1, B2, B3 and B6 Vitamin B12 | Methanolic extract Aqueous extract | [15,33] |
| Fat-soluble vitamins | Vitamins A, D3 and E | Cyclohexane and Dichloromethane extract | [15] |
| Free amino acids | Threonine, Methionine, Valine, Leucine/isoleucine, Phenylalanine (essential amino acids) Arginine, Serine, Aspartic acid, Glutamic acid, Glycine, Alanine, Tyrosine, Proline (non-essential amino acids) | Dried mushroom | [15,21] |
| Fatty acids | C 15:0, C 16:0, C 16:1, C 17:0, C 18:0, C 18:1n9c, C 18:2n6c, C 20:0, C 20:5n3, C 22:0, C 22:6n3, C 24:0, C 24:1n15c, SFAs, MUFAs, PUFAs | Cyclohexane extract | [15,21,34] |
| Sterols | Ergosterol, brassicasterol, campesterol, stigmasterol, β-sitosterol | Petroleum ether extract Chloroforme + methanol extract | [21,34,35] |
| Carotenoids | β-carotene and lycopene | Methanolic extract | [19] |
| Minerals | Fe, Zn, K, Na, Ca, Mn, Cu, Mg, Pb, Cd, As, P, Li | Fresh mushroom | [21,36,37,38,39] |
| Class of Compounds | Functional Activity | Edible Mushrooms Comparatively Tested | Reference |
|---|---|---|---|
| Flavonoids and phenolic compounds Free amino acids Fatty acids Sterols Minerals | α-glucosidase-inhibitory activity | Clitocybe maxima > C. cornucopioides > Stropharia rugoso-annulata > Laccaria amethystea > Catathelasma ventricosum (ethanolic extract) Stropharia rugoso-annulata > C. cornucopioides > Clitocybe maxima > Laccaria amethystea > Catathelasma ventricosum (ethyl acetate extract) | [21] |
| α-amylase-inhibitory activity | C. cornucopioides > Clitocybe maxima > Laccaria amethystea > Stropharia rugoso-annulata > Catathelasma ventricosum (ethanolic extract) Clitocybe maxima > Stropharia rugoso-annulata > C. cornucopioides > Laccaria amethystea > Catathelasma ventricosum (ethyl acetate extract) | ||
| Antioxidant—DPPH assay | Clitocybe maxima > C. cornucopioides > Stropharia rugoso-annulata > Laccaria amethystea > Catathelasma ventricosum (ethanolic extract) C. cornucopioides > Stropharia rugoso-annulata > Clitocybe maxima > Laccaria amethystea > Catathelasma ventricosum (ethyl acetate extract) | ||
| Antioxidant—ferrous-ion-chelating-activity assay | Stropharia rugoso-annulata > C. cornucopioides = Catathelasma ventricosum > Laccaria amethystea > Clitocybe maxima (ethanolic extract) Clitocybe maxima > Laccaria amethystea > Stropharia rugoso-annulata > C. cornucopioides > Catathelasma ventricosum (ethyl acetate extract) | ||
| Antioxidant—Reducing-power assay | Clitocybe maxima > Catathelasma ventricosum > Stropharia rugoso-annulata > C. cornucopioides > Laccaria amethystea (ethanolic extract) Stropharia rugoso-annulata > Catathelasma ventricosum > Clitocybe maxima > C. cornucopioides > Laccaria amethystea (ethyl acetate extract) | ||
| Flavonoids and phenolic compounds | Antioxidant—DPPH, ABTS assay | Pleurotus ostreatus > C. cornucopioides > Agaricus bisporus > Tuber melanosporum > Marasmius oreades (fluidized bed ethanol extracts) | [22] |
| Antioxidant—ferrous-ion-chelating-activity assay | Tuber melanosporum > C. cornucopioides > Marasmius oreades > Agaricus bisporus > Pleurotus ostreatus (fluidized bed ethanol extracts) | ||
| Antioxidant—inhibition-of-erythrocyte-hemolysis assay | Marasmius oreades > Tuber melanosporum > Pleurotus ostreatus > C. cornucopioides > Agaricus bisporus (fluidized bed ethanol extracts) | ||
| Anti-inflammatory—xanthine oxidase activity assay | Tuber melanosporum> Pleurotus ostreatus > C. cornucopioides > Agaricus bisporus (fluidized bed ethanol extracts) | ||
| Anti-inflammatory—lipoxygenase inhibition assay | C. cornucopioides > Agaricus bisporus > Tuber melanosporum> Pleurotus ostreatus (fluidized bed ethanol extracts) | ||
| Flavonoids and phenolic compounds | Cytotoxic | C. cornucopioides = Cantharellus cibarius > Cantharellus cinereus > Hydnum repandum > Calocybe gambosa (methanolic extracts) | [23] |
| Flavonoids and phenolic compounds | Antioxidant—inhibition-of-linoleic-acid-oxidation assay | Cantharellus cibarius > C. cornucopioides > Lactarius deliciosus > Calocybe gambosa > Hygrosphorus marzuolus > Boletus edulis > Pleurotus ostreatus > Agaricus bisporus (methanolic extract) | [20] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bumbu, M.-G.; Niculae, M.; Ielciu, I.; Hanganu, D.; Oniga, I.; Benedec, D.; Nechita, M.-A.; Nechita, V.-I.; Marcus, I. Comprehensive Review of Functional and Nutraceutical Properties of Craterellus cornucopioides (L.) Pers. Nutrients 2024, 16, 831. https://doi.org/10.3390/nu16060831
Bumbu M-G, Niculae M, Ielciu I, Hanganu D, Oniga I, Benedec D, Nechita M-A, Nechita V-I, Marcus I. Comprehensive Review of Functional and Nutraceutical Properties of Craterellus cornucopioides (L.) Pers. Nutrients. 2024; 16(6):831. https://doi.org/10.3390/nu16060831
Chicago/Turabian StyleBumbu, Mariana-Gabriela, Mihaela Niculae, Irina Ielciu, Daniela Hanganu, Ilioara Oniga, Daniela Benedec, Mihaela-Ancuța Nechita, Vlad-Ionuț Nechita, and Ioan Marcus. 2024. "Comprehensive Review of Functional and Nutraceutical Properties of Craterellus cornucopioides (L.) Pers." Nutrients 16, no. 6: 831. https://doi.org/10.3390/nu16060831
APA StyleBumbu, M.-G., Niculae, M., Ielciu, I., Hanganu, D., Oniga, I., Benedec, D., Nechita, M.-A., Nechita, V.-I., & Marcus, I. (2024). Comprehensive Review of Functional and Nutraceutical Properties of Craterellus cornucopioides (L.) Pers. Nutrients, 16(6), 831. https://doi.org/10.3390/nu16060831