

Human Breast Milk Exosomes: Affecting Factors, Their Possible Health Outcomes, and Future Directions in Dietetics

 , , ,
, , ,  and
and
Abstract
1. Introduction
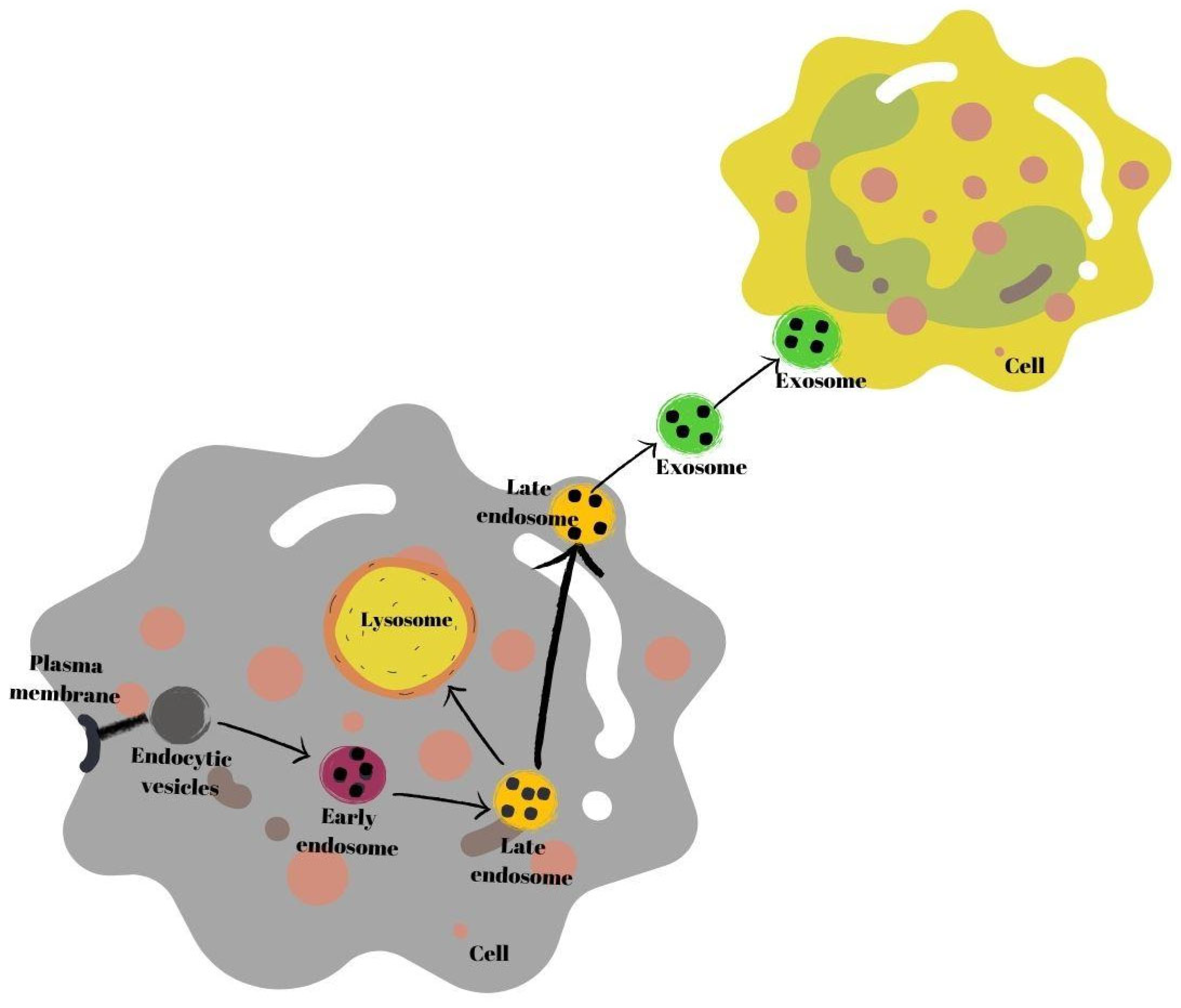
2. Exosomes Derived from Human Breast Milk
2.1. Components of Human Breast Milk Exosomes
2.1.1. Tetraspanins and Other Bioactive Proteins and Peptides
2.1.2. TGF-β
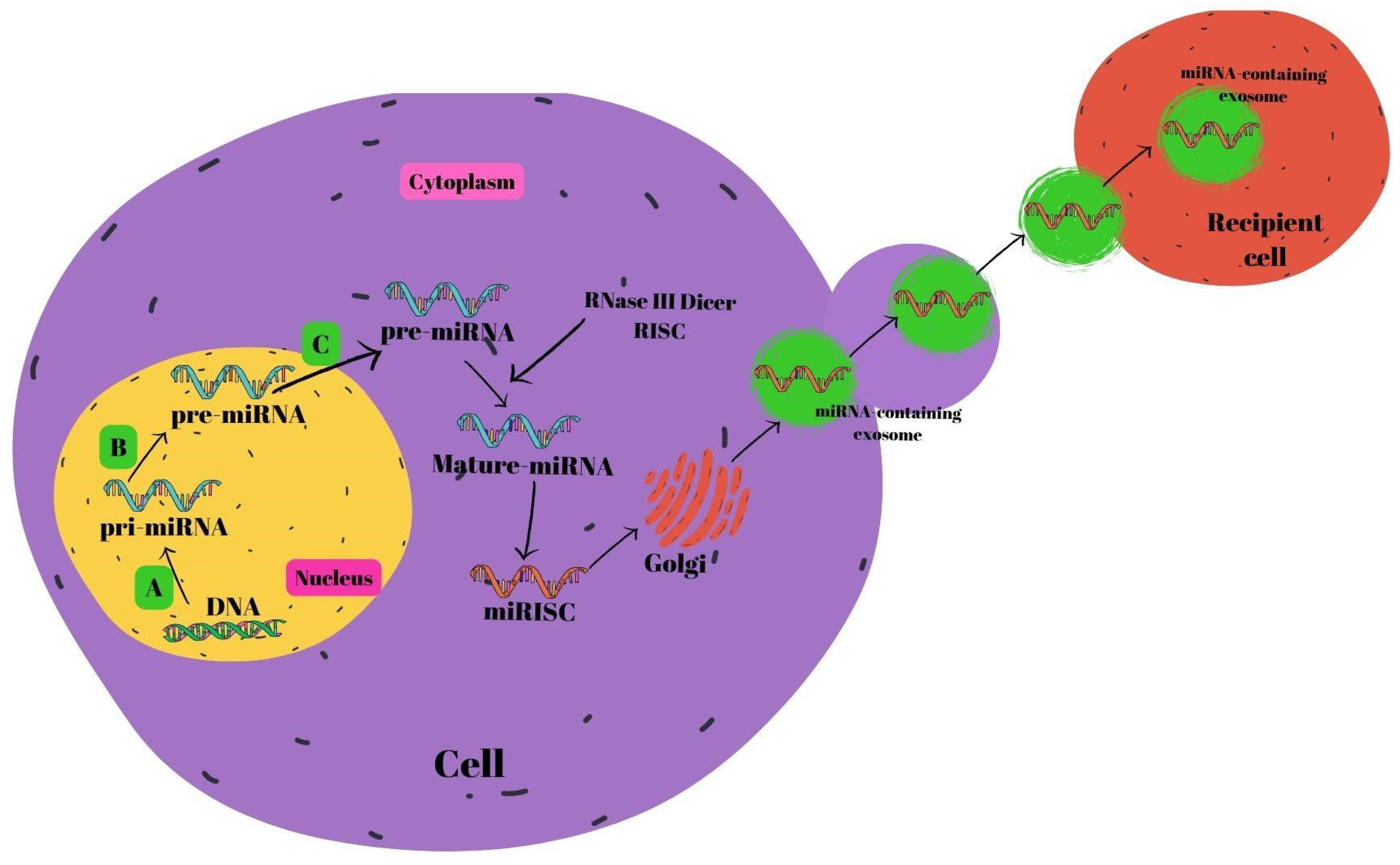
2.1.3. Noncoding RNAs
miRNAs
3. Factors Affecting Human Breast Milk Exosomes
3.1. Newborn-Associated Factors
3.1.1. Lactation Period
3.1.2. Preterm/Term Birth
3.1.3. Storage Conditions and Heat Treatment
3.1.4. Digestive System
3.2. Maternal-Associated Factors
3.2.1. Cesarean Section
3.2.2. Maternal Nutrition
3.2.3. Maternal Stress
3.2.4. Maternal Overweight and Obesity
3.2.5. Maternal Chronic Diseases
4. Possible Health Effects of Human Breast Milk Exosomes
4.1. Effects on the Immune System
4.1.1. Cell Proliferation and Inflammation
4.1.2. Immunomodulatory Function
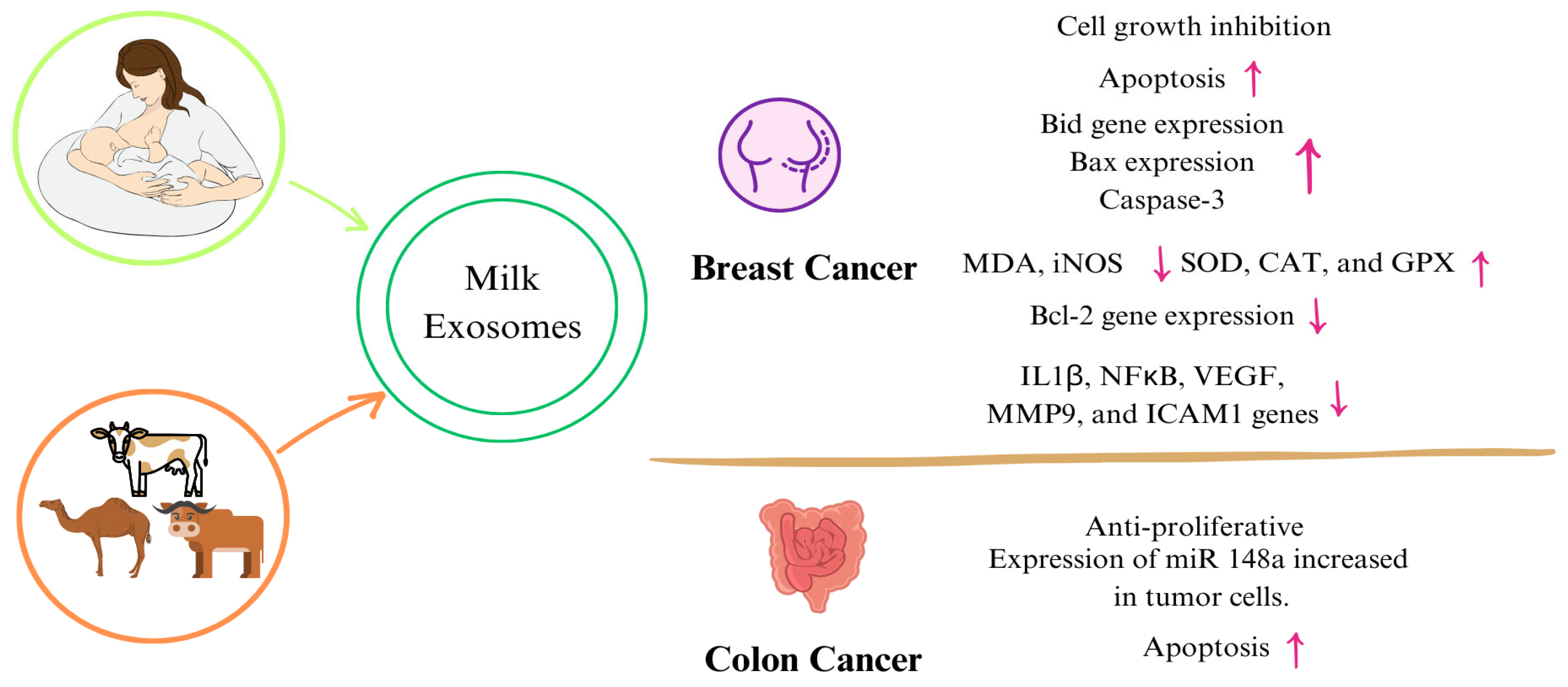
4.2. Effects on Cancer
4.3. Effects on Cardiometabolic Diseases
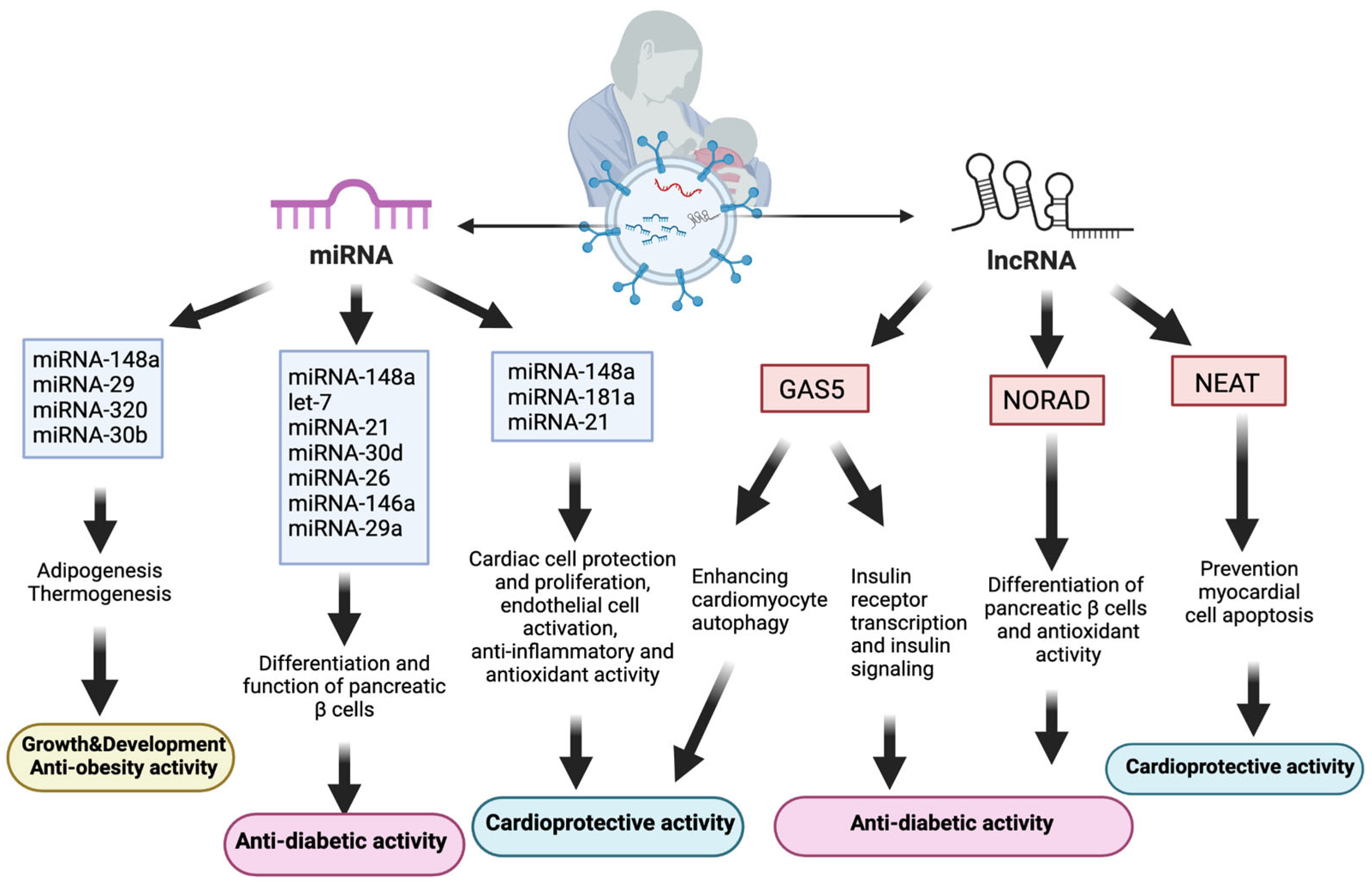
4.3.1. Effects on Obesity
4.3.2. Effects on Diabetes Mellitus
4.3.3. Effects on Cardiovascular Diseases (CVDs)
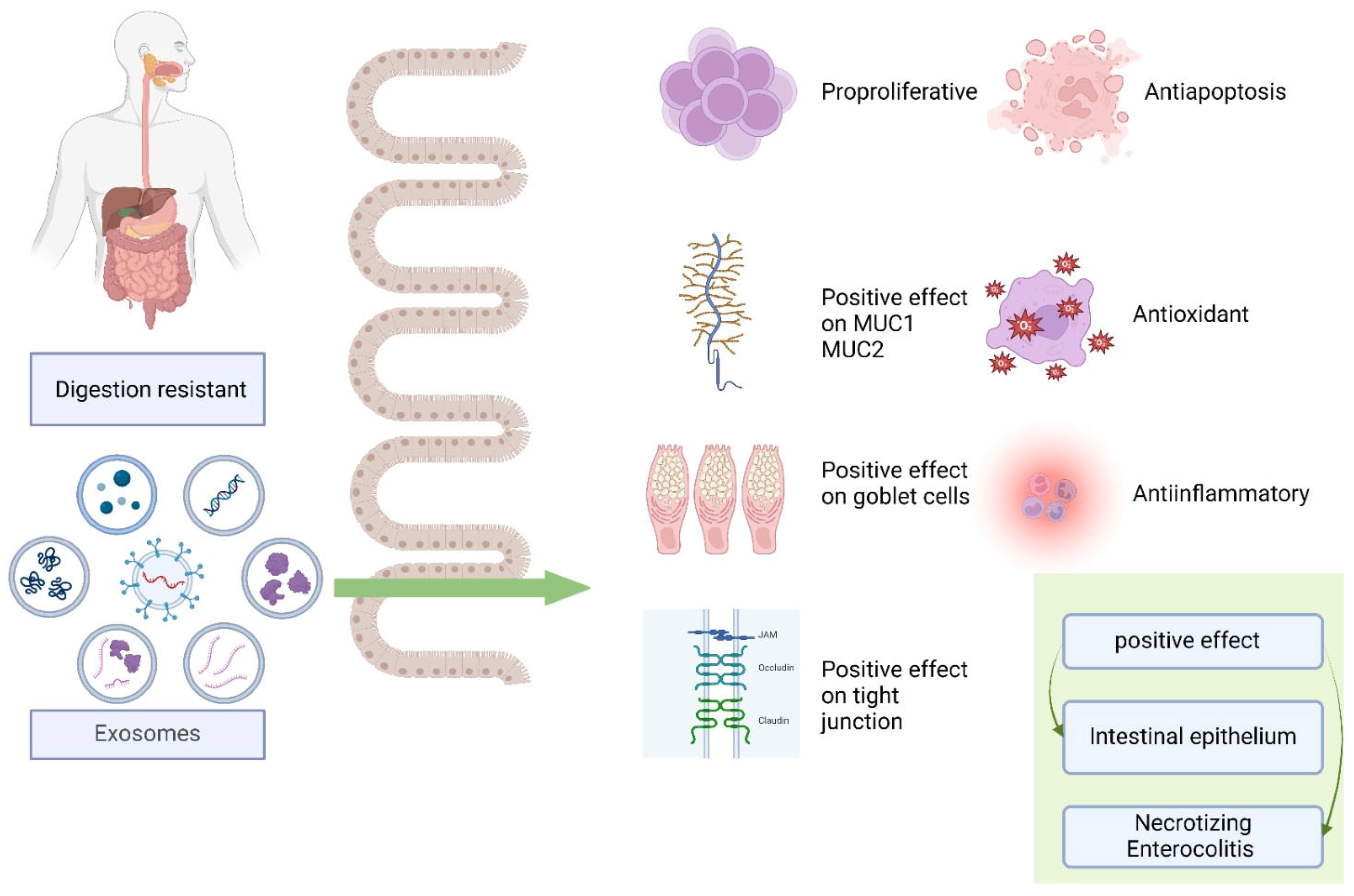
4.4. Effects on Intestinal Diseases
4.4.1. Intestinal Epithelial Function
4.4.2. Necrotizing Enterocolitis (NEC)
5. Future Directions of Human Breast Milk Exosomes in Dietetics
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kim, S.Y.; Yi, D.Y. Components of human breast milk: From macronutrient to microbiome and microRNA. Clin. Exp. Pediatr. 2020, 63, 301–309. [Google Scholar] [CrossRef]
- Hoddinott, P.; Tappin, D.; Wright, C. Breast feeding. BMJ 2008, 336, 881–887. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Implementation Guidance: Protecting, Promoting, and Supporting Breastfeeding in Facilities Providing Maternity and Newborn Services: The Revised Baby-Friendly Hospital Initiative 2018. Available online: https://www.who.int/publications/i/item/9789241513807 (accessed on 20 December 2023).
- Meek, J.Y.; Noble, L. Policy Statement: Breastfeeding and the Use of Human Milk. Pediatrics 2022, 150, e2022057988. [Google Scholar] [CrossRef]
- Agostoni, C.; Braegger, C.; Decsi, T.; Kolacek, S.; Koletzko, B.; Michaelsen, K.F.; Mihatsch, W.; Moreno, L.A.; Puntis, J.; Shamir, R.; et al. Breast-feeding: A commentary by the ESPGHAN Committee on Nutrition. J. Pediatr. Gastroenterol. Nutr. 2009, 49, 112–125. [Google Scholar] [CrossRef]
- Słyk-Gulewska, P.; Kondracka, A.; Kwaśniewska, A. MicroRNA as a new bioactive component in breast milk. Noncoding RNA Res. 2023, 8, 520–526. [Google Scholar] [CrossRef]
- Kim, K.U.; Kim, W.H.; Jeong, C.H.; Yi, D.Y.; Min, H. More than Nutrition: Therapeutic Potential of Breast Milk-Derived Exosomes in Cancer. Int. J. Mol. Sci. 2020, 21, 7327. [Google Scholar] [CrossRef]
- Koletzko, B.; Baker, S.; Cleghorn, G.; Neto, U.F.; Gopalan, S.; Hernell, O.; Hock, Q.S.; Jirapinyo, P.; Lonnerdal, B.; Pencharz, P.; et al. Global standard for the composition of infant formula: Recommendations of an ESPGHAN coordinated international expert group. J. Pediatr. Gastroenterol. Nutr. 2005, 41, 584–599. [Google Scholar] [CrossRef]
- Andreas, N.J.; Kampmann, B.; Mehring Le-Doare, K. Human breast milk: A review on its composition and bioactivity. Early Hum. Dev. 2015, 91, 629–635. [Google Scholar] [CrossRef]
- Kim, K.U.; Han, K.; Kim, J.; Kwon, D.H.; Ji, Y.W.; Yi, D.Y.; Min, H. The Protective Role of Exosome-Derived MicroRNAs and Proteins from Human Breast Milk against Infectious Agents. Metabolites 2023, 13, 635. [Google Scholar] [CrossRef]
- Melnik, B.C.; Stremmel, W.; Weiskirchen, R.; John, S.M.; Schmitz, G. Exosome-Derived MicroRNAs of Human Milk and Their Effects on Infant Health and Development. Biomolecules 2021, 11, 851. [Google Scholar] [CrossRef]
- Stephen, B.J.; Pareek, N.; Saeed, M.; Kausar, M.A.; Rahman, S.; Datta, M. Xeno-miRNA in Maternal-Infant Immune Crosstalk: An Aid to Disease Alleviation. Front. Immunol. 2020, 11, 404. [Google Scholar] [CrossRef]
- Carr, L.E.; Virmani, M.D.; Rosa, F.; Munblit, D.; Matazel, K.S.; Elolimy, A.A.; Yeruva, L. Role of Human Milk Bioactives on Infants’ Gut and Immune Health. Front. Immunol. 2021, 12, 604080. [Google Scholar] [CrossRef]
- Newburg, D.S.; Walker, W.A. Protection of the neonate by the innate immune system of developing gut and of human milk. Pediatr. Res. 2007, 61, 2–8. [Google Scholar] [CrossRef]
- Yi, D.Y.; Kim, S.Y. Human Breast Milk Composition and Function in Human Health: From Nutritional Components to Microbiome and MicroRNAs. Nutrients 2021, 13, 3094. [Google Scholar] [CrossRef]
- Zeng, B.; Chen, T.; Luo, J.Y.; Zhang, L.; Xi, Q.Y.; Jiang, Q.Y.; Sun, J.J.; Zhang, Y.L. Biological Characteristics and Roles of Noncoding RNAs in Milk-Derived Extracellular Vesicles. Adv. Nutr. 2021, 12, 1006–1019. [Google Scholar] [CrossRef]
- Hock, A.; Miyake, H.; Li, B.; Lee, C.; Ermini, L.; Koike, Y.; Chen, Y.; Määttänen, P.; Zani, A.; Pierro, A. Breast milk-derived exosomes promote intestinal epithelial cell growth. J. Pediatr. Surg. 2017, 52, 755–759. [Google Scholar] [CrossRef]
- Torregrosa Paredes, P.; Gutzeit, C.; Johansson, S.; Admyre, C.; Stenius, F.; Alm, J.; Scheynius, A.; Gabrielsson, S. Differences in exosome populations in human breast milk in relation to allergic sensitization and lifestyle. Allergy 2014, 69, 463–471. [Google Scholar] [CrossRef]
- Admyre, C.; Johansson, S.M.; Qazi, K.R.; Filén, J.J.; Lahesmaa, R.; Norman, M.; Neve, E.P.; Scheynius, A.; Gabrielsson, S. Exosomes with immune modulatory features are present in human breast milk. J. Immunol. 2007, 179, 1969–1978. [Google Scholar] [CrossRef]
- Lönnerdal, B. Summary on Human Milk Composition and Physiological Benefits. Nestle Nutr. Inst. Workshop Ser. 2019, 90, 103–105. [Google Scholar] [CrossRef]
- Chutipongtanate, S.; Morrow, A.L.; Newburg, D.S. Human Milk Extracellular Vesicles: A Biological System with Clinical Implications. Cells 2022, 11, 2345. [Google Scholar] [CrossRef]
- Couch, Y.; Buzàs, E.I.; Di Vizio, D.; Gho, Y.S.; Harrison, P.; Hill, A.F.; Lötvall, J.; Raposo, G.; Stahl, P.D.; Théry, C.; et al. A brief history of nearly EV-erything—The rise and rise of extracellular vesicles. J. Extracell. Vesicles 2021, 10, e12144. [Google Scholar] [CrossRef]
- Rashed, M.H.; Bayraktar, E.; Helal, G.K.; Abd-Ellah, M.F.; Amero, P.; Chavez-Reyes, A.; Rodriguez-Aguayo, C. Exosomes: From Garbage Bins to Promising Therapeutic Targets. Int. J. Mol. Sci. 2017, 18, 538. [Google Scholar] [CrossRef]
- Feng, X.; Chen, X.; Zheng, X.; Zhu, H.; Qi, Q.; Liu, S.; Zhang, H.; Che, J. Latest Trend of Milk Derived Exosomes: Cargos, Functions, and Applications. Front. Nutr. 2021, 8, 747294. [Google Scholar] [CrossRef]
- Gialeli, G.; Panagopoulou, O.; Liosis, G.; Siahanidou, T. Potential Epigenetic Effects of Human Milk on Infants’ Neurodevelopment. Nutrients 2023, 15, 3614. [Google Scholar] [CrossRef]
- Goyal, D.; Limesand, S.W.; Goyal, R. Epigenetic responses and the developmental origins of health and disease. J. Endocrinol. 2019, 242, T105–T119. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef]
- de la Torre Gomez, C.; Goreham, R.V.; Bech Serra, J.J.; Nann, T.; Kussmann, M. “Exosomics”—A Review of Biophysics, Biology and Biochemistry of Exosomes with a Focus on Human Breast Milk. Front. Genet. 2018, 9, 92. [Google Scholar] [CrossRef]
- Dieterich, C.M.; Felice, J.P.; O’Sullivan, E.; Rasmussen, K.M. Breastfeeding and health outcomes for the mother-infant dyad. Pediatr. Clin. N. Am. 2013, 60, 31–48. [Google Scholar] [CrossRef]
- Holme, A.M.; Sitras, V. Developmental origin of health and disease-Evidence and time for action. Acta Obstet. Gynecol. Scand. 2020, 99, 961–962. [Google Scholar] [CrossRef]
- Barker, D.J. The developmental origins of adult disease. J. Am. Coll. Nutr. 2004, 23, 588s–595s. [Google Scholar] [CrossRef]
- Shah, J.; Sims, B.; Martin, C. Therapeutic potential of human breast milk derived exosomes. J. Nanopart. Res. 2022, 24, 260. [Google Scholar] [CrossRef]
- Lässer, C.; Alikhani, V.S.; Ekström, K.; Eldh, M.; Paredes, P.T.; Bossios, A.; Sjöstrand, M.; Gabrielsson, S.; Lötvall, J.; Valadi, H. Human saliva, plasma and breast milk exosomes contain RNA: Uptake by macrophages. J. Transl. Med. 2011, 9, 9. [Google Scholar] [CrossRef]
- Le Doare, K.; Holder, B.; Bassett, A.; Pannaraj, P.S. Mother’s Milk: A Purposeful Contribution to the Development of the Infant Microbiota and Immunity. Front. Immunol. 2018, 9, 361. [Google Scholar] [CrossRef]
- Ramachandran, S.; Palanisamy, V. Horizontal transfer of RNAs: Exosomes as mediators of intercellular communication. Wiley Interdiscip. Rev. RNA 2012, 3, 286–293. [Google Scholar] [CrossRef]
- Li, C.; Ni, Y.Q.; Xu, H.; Xiang, Q.Y.; Zhao, Y.; Zhan, J.K.; He, J.Y.; Li, S.; Liu, Y.S. Roles and mechanisms of exosomal non-coding RNAs in human health and diseases. Signal. Transduct. Target Ther. 2021, 6, 383. [Google Scholar] [CrossRef]
- Koritzinsky, E.H.; Street, J.M.; Star, R.A.; Yuen, P.S. Quantification of Exosomes. J. Cell. Physiol. 2017, 232, 1587–1590. [Google Scholar] [CrossRef]
- Lin, J.; Li, J.; Huang, B.; Liu, J.; Chen, X.; Chen, X.M.; Xu, Y.M.; Huang, L.F.; Wang, X.Z. Exosomes: Novel biomarkers for clinical diagnosis. Sci. World J. 2015, 2015, 657086. [Google Scholar] [CrossRef]
- Galley, J.D.; Besner, G.E. The Therapeutic Potential of Breast Milk-Derived Extracellular Vesicles. Nutrients 2020, 12, 745. [Google Scholar] [CrossRef]
- Martin, C.; Patel, M.; Williams, S.; Arora, H.; Brawner, K.; Sims, B. Human breast milk-derived exosomes attenuate cell death in intestinal epithelial cells. Innate Immun. 2018, 24, 278–284. [Google Scholar] [CrossRef]
- Manca, S.; Upadhyaya, B.; Mutai, E.; Desaulniers, A.T.; Cederberg, R.A.; White, B.R.; Zempleni, J. Milk exosomes are bioavailable and distinct microRNA cargos have unique tissue distribution patterns. Sci. Rep. 2018, 8, 11321. [Google Scholar] [CrossRef]
- Ståhl, A.L.; Johansson, K.; Mossberg, M.; Kahn, R.; Karpman, D. Exosomes and microvesicles in normal physiology, pathophysiology, and renal diseases. Pediatr. Nephrol. 2019, 34, 11–30. [Google Scholar] [CrossRef]
- Zempleni, J.; Aguilar-Lozano, A.; Sadri, M.; Sukreet, S.; Manca, S.; Wu, D.; Zhou, F.; Mutai, E. Biological Activities of Extracellular Vesicles and Their Cargos from Bovine and Human Milk in Humans and Implications for Infants. J. Nutr. 2017, 147, 3–10. [Google Scholar] [CrossRef]
- Lässer, C. Exosomal RNA as biomarkers and the therapeutic potential of exosome vectors. Expert Opin. Biol. Ther. 2012, 12 (Suppl. S1), S189–S197. [Google Scholar] [CrossRef]
- Zeringer, E.; Li, M.; Barta, T.; Schageman, J.; Pedersen, K.W.; Neurauter, A.; Magdaleno, S.; Setterquist, R.; Vlassov, A.V. Methods for the extraction and RNA profiling of exosomes. World J. Methodol. 2013, 3, 11–18. [Google Scholar] [CrossRef]
- Şahin, Ö.N. The Part Breast Milk Plays in Epigenetic Programming. In Breastfeeding and Metabolic Programming; Şahin, Ö.N., Briana, D.D., Di Renzo, G.C., Eds.; Springer International Publishing: Cham, Switzerland, 2023; pp. 131–147. [Google Scholar]
- Dai, J.; Su, Y.; Zhong, S.; Cong, L.; Liu, B.; Yang, J.; Tao, Y.; He, Z.; Chen, C.; Jiang, Y. Exosomes: Key players in cancer and potential therapeutic strategy. Signal. Transduct. Target Ther. 2020, 5, 145. [Google Scholar] [CrossRef]
- Geddes, D.; Kakulas, F. 7 Human Milk: Bioactive Components and Their Effects on the Infant and Beyond. In Breastfeeding and Breast Milk—From Biochemistry to Impact; Family Larson-Rosenquist Foundation, Ed.; Georg Thieme Verlag KG: Stuttgart, Germany, 2018. [Google Scholar]
- Giovanazzi, A.; van Herwijnen, M.J.C.; Kleinjan, M.; van der Meulen, G.N.; Wauben, M.H.M. Surface protein profiling of milk and serum extracellular vesicles unveils body fluid-specific signatures. Sci. Rep. 2023, 13, 8758. [Google Scholar] [CrossRef]
- Weiskirchen, R.; Schröder, S.K.; Weiskirchen, S.; Buhl, E.M.; Melnik, B. Isolation of Bovine and Human Milk Extracellular Vesicles. Biomedicines 2023, 11, 2715. [Google Scholar] [CrossRef]
- Vaswani, K.; Mitchell, M.D.; Holland, O.J.; Qin Koh, Y.; Hill, R.J.; Harb, T.; Davies, P.S.W.; Peiris, H. A Method for the Isolation of Exosomes from Human and Bovine Milk. J. Nutr. Metab. 2019, 2019, 5764740. [Google Scholar] [CrossRef]
- Dong, P.; Zhang, Y.; Yan, D.Y.; Wang, Y.; Xu, X.; Zhao, Y.C.; Xiao, T.T. Protective Effects of Human Milk-Derived Exosomes on Intestinal Stem Cells Damaged by Oxidative Stress. Cell Transplant. 2020, 29, 963689720912690. [Google Scholar] [CrossRef]
- Liao, Y.; Du, X.; Li, J.; Lönnerdal, B. Human milk exosomes and their microRNAs survive digestion in vitro and are taken up by human intestinal cells. Mol. Nutr. Food. Res 2017, 61, 82. [Google Scholar] [CrossRef]
- Wang, X.; Yan, X.; Zhang, L.; Cai, J.; Zhou, Y.; Liu, H.; Hu, Y.; Chen, W.; Xu, S.; Liu, P.; et al. Identification and Peptidomic Profiling of Exosomes in Preterm Human Milk: Insights into Necrotizing Enterocolitis Prevention. Mol. Nutr. Food. Res. 2019, 63, e1801247. [Google Scholar] [CrossRef]
- Qin, W.; Tsukasaki, Y.; Dasgupta, S.; Mukhopadhyay, N.; Ikebe, M.; Sauter, E.R. Exosomes in Human Breast Milk Promote EMT. Clin. Cancer. Res. 2016, 22, 4517–4524. [Google Scholar] [CrossRef]
- Rubio, M.; Bustamante, M.; Hernandez-Ferrer, C.; Fernandez-Orth, D.; Pantano, L.; Sarria, Y.; Piqué-Borras, M.; Vellve, K.; Agramunt, S.; Carreras, R.; et al. Circulating miRNAs, isomiRs and small RNA clusters in human plasma and breast milk. PLoS ONE 2018, 13, e0193527. [Google Scholar] [CrossRef]
- Hashemian, S.M.; Pourhanifeh, M.H.; Fadaei, S.; Velayati, A.A.; Mirzaei, H.; Hamblin, M.R. Non-coding RNAs and Exosomes: Their Role in the Pathogenesis of Sepsis. Mol. Ther. Nucleic Acids 2020, 21, 51–74. [Google Scholar] [CrossRef]
- Hammond, S.M. An overview of microRNAs. Adv. Drug Deliv. Rev. 2015, 87, 3–14. [Google Scholar] [CrossRef]
- Wahid, F.; Shehzad, A.; Khan, T.; Kim, Y.Y. MicroRNAs: Synthesis, mechanism, function, and recent clinical trials. Biochim. Biophys. Acta 2010, 1803, 1231–1243. [Google Scholar] [CrossRef]
- Zhang, J.; Li, S.; Li, L.; Li, M.; Guo, C.; Yao, J.; Mi, S. Exosome and exosomal microRNA: Trafficking, sorting, and function. Genom. Proteom. Bioinform. 2015, 13, 17–24. [Google Scholar] [CrossRef]
- Xu, L.; Yang, B.F.; Ai, J. MicroRNA transport: A new way in cell communication. J. Cell. Physiol. 2013, 228, 1713–1719. [Google Scholar] [CrossRef]
- Yu, X.; Odenthal, M.; Fries, J.W. Exosomes as miRNA Carriers: Formation-Function-Future. Int. J. Mol. Sci. 2016, 17, 2028. [Google Scholar] [CrossRef]
- Suzuki, H.I. Roles of MicroRNAs in Disease Biology. JMA J. 2023, 6, 104–113. [Google Scholar] [CrossRef]
- Paul, P.; Chakraborty, A.; Sarkar, D.; Langthasa, M.; Rahman, M.; Bari, M.; Singha, R.S.; Malakar, A.K.; Chakraborty, S. Interplay between miRNAs and human diseases. J. Cell. Physiol. 2018, 233, 2007–2018. [Google Scholar] [CrossRef]
- Manna, I.; De Benedittis, S.; Quattrone, A.; Maisano, D.; Iaccino, E.; Quattrone, A. Exosomal miRNAs as Potential Diagnostic Biomarkers in Alzheimer’s Disease. Pharmaceuticals 2020, 13, 243. [Google Scholar] [CrossRef]
- Oliveira, S.R.; Dionísio, P.A.; Correia Guedes, L.; Gonçalves, N.; Coelho, M.; Rosa, M.M.; Amaral, J.D.; Ferreira, J.J.; Rodrigues, C.M.P. Circulating Inflammatory miRNAs Associated with Parkinson’s Disease Pathophysiology. Biomolecules 2020, 10, 945. [Google Scholar] [CrossRef]
- Khodakarimi, S.; Zarebkohan, A.; Kahroba, H.; Omrani, M.; Sepasi, T.; Mohaddes, G.; Beyrampour-Basmenj, H.; Ebrahimi, A.; Ebrahimi-Kalan, A. The role of miRNAs in the regulation of autophagy in autoimmune diseases. Life Sci. 2021, 287, 119726. [Google Scholar] [CrossRef]
- Ren, F.; Gao, W.C.; Ke, Z.P.; Xu, Y.; Liu, Y. Identification of six miRNAs serving as predictive biomarkers in coronary artery disease. J. Cell. Biochem. 2019, 120, 1932–1942. [Google Scholar] [CrossRef]
- Brandao, B.B.; Lino, M.; Kahn, C.R. Extracellular miRNAs as mediators of obesity-associated disease. J. Physiol. 2022, 600, 1155–1169. [Google Scholar] [CrossRef]
- Zou, C. Advances in the study of miRNAs in chronic kidney disease with cardiovascular complications. Front. Physiol. 2023, 14, 1283597. [Google Scholar] [CrossRef]
- Alotaibi, F. Exosomal microRNAs in cancer: Potential biomarkers and immunotherapeutic targets for immune checkpoint molecules. Front. Genet. 2023, 14, 1052731. [Google Scholar] [CrossRef]
- Joseph, D.; Grover, B.; Telias, M. Biomarker potential of vitreous microRNA in retinal disease: A meta-analysis. medRxiv 2024. [Google Scholar] [CrossRef]
- Tingö, L.; Ahlberg, E.; Johansson, L.; Pedersen, S.A.; Chawla, K.; Sætrom, P.; Cione, E.; Simpson, M.R. Non-Coding RNAs in Human Breast Milk: A Systematic Review. Front. Immunol. 2021, 12, 725323. [Google Scholar] [CrossRef]
- Benmoussa, A.; Provost, P. Milk MicroRNAs in Health and Disease. Compr. Rev. Food Sci. Food Saf. 2019, 18, 703–722. [Google Scholar] [CrossRef]
- Golan-Gerstl, R.; Elbaum Shiff, Y.; Moshayoff, V.; Schecter, D.; Leshkowitz, D.; Reif, S. Characterization and biological function of milk-derived miRNAs. Mol. Nutr. Food Res 2017, 61, 1700009. [Google Scholar] [CrossRef]
- Leiferman, A.; Shu, J.; Upadhyaya, B.; Cui, J.; Zempleni, J. Storage of Extracellular Vesicles in Human Milk, and MicroRNA Profiles in Human Milk Exosomes and Infant Formulas. J. Pediatr. Gastroenterol. Nutr. 2019, 69, 235–238. [Google Scholar] [CrossRef]
- Reif, S.; Elbaum-Shiff, Y.; Koroukhov, N.; Shilo, I.; Musseri, M.; Golan-Gerstl, R. Cow and Human Milk-Derived Exosomes Ameliorate Colitis in DSS Murine Model. Nutrients 2020, 12, 2589. [Google Scholar] [CrossRef]
- Yang, L.; Hu, R.; Li, J.; Mo, X.; Xu, L.; Shen, N.; Sheng, W.; Li, Y. Exosomal microRNAs in Human Breast Milk: Potential Effect on Neonatal Breast Milk Jaundice. 2020. Available online: https://assets-eu.researchsquare.com/files/rs-77051/v1/1e1f2b7e-03a2-4d87-afd0-52637c0a77a1.pdf?c=1631857657 (accessed on 14 July 2024).
- Hatmal, M.M.; Al-Hatamleh, M.A.I.; Olaimat, A.N.; Alshaer, W.; Hasan, H.; Albakri, K.A.; Alkhafaji, E.; Issa, N.N.; Al-Holy, M.A.; Abderrahman, S.M.; et al. Immunomodulatory Properties of Human Breast Milk: MicroRNA Contents and Potential Epigenetic Effects. Biomedicines 2022, 10, 1219. [Google Scholar] [CrossRef]
- Xi, Y.; Jiang, X.; Li, R.; Chen, M.; Song, W.; Li, X. The levels of human milk microRNAs and their association with maternal weight characteristics. Eur. J. Clin. Nutr. 2016, 70, 445–449. [Google Scholar] [CrossRef]
- Shiff, Y.E.; Reif, S.; Marom, R.; Shiff, K.; Reifen, R.; Golan-Gerstl, R. MiRNA-320a is less expressed and miRNA-148a more expressed in preterm human milk compared to term human milk. J. Funct. Foods 2019, 57, 68–74. [Google Scholar] [CrossRef]
- Freiría-Martínez, L.; Iglesias-Martínez-Almeida, M.; Rodríguez-Jamardo, C.; Rivera-Baltanás, T.; Comís-Tuche, M.; Rodrígues-Amorím, D.; Fernández-Palleiro, P.; Blanco-Formoso, M.; Diz-Chaves, Y.; González-Freiria, N.; et al. Human Breast Milk microRNAs, Potential Players in the Regulation of Nervous System. Nutrients 2023, 15, 3284. [Google Scholar] [CrossRef]
- Alsaweed, M.; Lai, C.T.; Hartmann, P.E.; Geddes, D.T.; Kakulas, F. Human Milk Cells Contain Numerous miRNAs that May Change with Milk Removal and Regulate Multiple Physiological Processes. Int. J. Mol. Sci. 2016, 17, 956. [Google Scholar] [CrossRef]
- Yan, X.; Liu, L.; Yao, S.; Chen, Y.; Yu, Q.; Jiang, C.; Chen, W.; Chen, X.; Han, S. LncRNA and mRNA profiles of human milk-derived exosomes and their possible roles in protecting against necrotizing enterocolitis. Food. Funct. 2022, 13, 12953–12965. [Google Scholar] [CrossRef]
- Mourtzi, N.; Siahanidou, T.; Tsifintaris, M.; Karamichali, E.; Tasiopoulou, A.; Sertedaki, A.; Pesmatzoglou, M.; Kapetanaki, A.; Liosis, G.; Baltatzis, G.; et al. lncRNA NORAD is consistently detected in breastmilk exosomes and its expression is downregulated in mothers of preterm infants. Int. J. Mol. Med. 2021, 48, 5049. [Google Scholar] [CrossRef]
- Garcia, C.; Duan, R.D.; Brévaut-Malaty, V.; Gire, C.; Millet, V.; Simeoni, U.; Bernard, M.; Armand, M. Bioactive compounds in human milk and intestinal health and maturity in preterm newborn: An overview. Cell. Mol. Biol. 2013, 59, 108–131. [Google Scholar]
- Carney, M.C.; Tarasiuk, A.; DiAngelo, S.L.; Silveyra, P.; Podany, A.; Birch, L.L.; Paul, I.M.; Kelleher, S.; Hicks, S.D. Metabolism-related microRNAs in maternal breast milk are influenced by premature delivery. Pediatr. Res. 2017, 82, 226–236. [Google Scholar] [CrossRef]
- Clerc, P.; Coll Constans, M.G.; Lulka, H.; Broussaud, S.; Guigné, C.; Leung-Theung-Long, S.; Perrin, C.; Knauf, C.; Carpéné, C.; Pénicaud, L.; et al. Involvement of cholecystokinin 2 receptor in food intake regulation: Hyperphagia and increased fat deposition in cholecystokinin 2 receptor-deficient mice. Endocrinology 2007, 148, 1039–1049. [Google Scholar] [CrossRef]
- Kahn, S.; Liao, Y.; Du, X.; Xu, W.; Li, J.; Lönnerdal, B. Exosomal MicroRNAs in Milk from Mothers Delivering Preterm Infants Survive In Vitro Digestion and Are Taken Up by Human Intestinal Cells. Mol. Nutr. Food Res. 2018, 62, e1701050. [Google Scholar] [CrossRef]
- Zhou, Y.; Yu, Z.; Wang, X.; Chen, W.; Liu, Y.; Zhang, Y.; Yin, J.; Han, S. Exosomal circRNAs contribute to intestinal development via the VEGF signalling pathway in human term and preterm colostrum. Aging 2021, 13, 11218–11233. [Google Scholar] [CrossRef]
- Miyake, H.; Lee, C.; Chusilp, S.; Bhalla, M.; Li, B.; Pitino, M.; Seo, S.; O’Connor, D.L.; Pierro, A. Human breast milk exosomes attenuate intestinal damage. Pediatr. Surg. Int. 2020, 36, 155–163. [Google Scholar] [CrossRef]
- Howard, K.M.; Jati Kusuma, R.; Baier, S.R.; Friemel, T.; Markham, L.; Vanamala, J.; Zempleni, J. Loss of miRNAs during processing and storage of cow’s (Bos taurus) milk. J. Agric. Food Chem. 2015, 63, 588–592. [Google Scholar] [CrossRef]
- Smyczynska, U.; Bartlomiejczyk, M.A.; Stanczak, M.M.; Sztromwasser, P.; Wesolowska, A.; Barbarska, O.; Pawlikowska, E.; Fendler, W. Impact of processing method on donated human breast milk microRNA content. PLoS ONE 2020, 15, e0236126. [Google Scholar] [CrossRef]
- Wang, H.; Wu, D.; Sukreet, S.; Delaney, A.; Belfort, M.B.; Zempleni, J. Quantitation of Exosomes and Their MicroRNA Cargos in Frozen Human Milk. JPGN Rep. 2022, 3, e172. [Google Scholar] [CrossRef]
- Kirchner, B.; Pfaffl, M.W.; Dumpler, J.; von Mutius, E.; Ege, M.J. microRNA in native and processed cow’s milk and its implication for the farm milk effect on asthma. J. Allergy Clin. Immunol. 2016, 137, 1893–1895.e1813. [Google Scholar] [CrossRef]
- Kleinjan, M.; van Herwijnen, M.J.; Libregts, S.F.; van Neerven, R.J.; Feitsma, A.L.; Wauben, M.H. Regular Industrial Processing of Bovine Milk Impacts the Integrity and Molecular Composition of Extracellular Vesicles. J. Nutr. 2021, 151, 1416–1425. [Google Scholar] [CrossRef]
- Lu, L.; Han, C.; Wang, M.; Du, H.; Chen, N.; Gao, M.; Wang, N.; Qi, D.; Bai, W.; Yin, J.; et al. Study on the preparation of bovine milk exosomes and the stability of lyophilized powder. bioRxiv 2023. [Google Scholar] [CrossRef]
- Kosaka, N.; Izumi, H.; Sekine, K.; Ochiya, T. microRNA as a new immune-regulatory agent in breast milk. Silence 2010, 1, 7. [Google Scholar] [CrossRef]
- Melnik, B.C. Milk exosomal miRNAs: Potential drivers of AMPK-to-mTORC1 switching in β-cell de-differentiation of type 2 diabetes mellitus. Nutr. Metab. 2019, 16, 85. [Google Scholar] [CrossRef]
- Weil, P.P.; Reincke, S.; Hirsch, C.A.; Giachero, F.; Aydin, M.; Scholz, J.; Jönsson, F.; Hagedorn, C.; Nguyen, D.N.; Thymann, T.; et al. Uncovering the gastrointestinal passage, intestinal epithelial cellular uptake, and AGO2 loading of milk miRNAs in neonates using xenomiRs as tracers. Am. J. Clin. Nutr. 2023, 117, 1195–1210. [Google Scholar] [CrossRef]
- Mirza, A.H.; Kaur, S.; Nielsen, L.B.; Størling, J.; Yarani, R.; Roursgaard, M.; Mathiesen, E.R.; Damm, P.; Svare, J.; Mortensen, H.B.; et al. Breast Milk-Derived Extracellular Vesicles Enriched in Exosomes from Mothers with Type 1 Diabetes Contain Aberrant Levels of microRNAs. Front. Immunol. 2019, 10, 2543. [Google Scholar] [CrossRef]
- Gutman-Ido, E.; Reif, S.; Musseri, M.; Schabes, T.; Golan-Gerstl, R. Oxytocin Regulates the Expression of Selected Colostrum-derived microRNAs. J. Pediatr. Gastroenterol. Nutr. 2022, 74, e8–e15. [Google Scholar] [CrossRef]
- Chiba, T.; Kooka, A.; Kowatari, K.; Yoshizawa, M.; Chiba, N.; Takaguri, A.; Fukushi, Y.; Hongo, F.; Sato, H.; Wada, S. Expression profiles of hsa-miR-148a-3p and hsa-miR-125b-5p in human breast milk and infant formulae. Int. Breastfeed J. 2022, 17, 1. [Google Scholar] [CrossRef]
- Munch, E.M.; Harris, R.A.; Mohammad, M.; Benham, A.L.; Pejerrey, S.M.; Showalter, L.; Hu, M.; Shope, C.D.; Maningat, P.D.; Gunaratne, P.H.; et al. Transcriptome profiling of microRNA by Next-Gen deep sequencing reveals known and novel miRNA species in the lipid fraction of human breast milk. PLoS ONE 2013, 8, e50564. [Google Scholar] [CrossRef]
- Lukasik, A.; Brzozowska, I.; Zielenkiewicz, U.; Zielenkiewicz, P. Detection of Plant miRNAs Abundance in Human Breast Milk. Int. J. Mol. Sci 2017, 19, 37. [Google Scholar] [CrossRef]
- Zamanillo, R.; Sánchez, J.; Serra, F.; Palou, A. Breast Milk Supply of MicroRNA Associated with Leptin and Adiponectin Is Affected by Maternal Overweight/Obesity and Influences Infancy BMI. Nutrients 2019, 11, 2589. [Google Scholar] [CrossRef]
- Kaeffer, B. Human Breast Milk miRNAs: Their Diversity and Potential for Preventive Strategies in Nutritional Therapy. Int. J. Mol. Sci. 2023, 24, 6106. [Google Scholar] [CrossRef]
- Pomar, C.A.; Castro, H.; Picó, C.; Serra, F.; Palou, A.; Sánchez, J. Cafeteria Diet Consumption during Lactation in Rats, rather than Obesity Per Se, alters miR-222, miR-200a, and miR-26a Levels in Milk. Mol. Nutr. Food Res. 2019, 63, e1800928. [Google Scholar] [CrossRef]
- Susiloretni, K.A.; Smith, E.R.; Suparmi; Marsum; Agustina, R.; Shankar, A.H. The psychological distress of parents is associated with reduced linear growth of children: Evidence from a nationwide population survey. PLoS ONE 2021, 16, e0246725. [Google Scholar] [CrossRef]
- Bozack, A.K.; Colicino, E.; Rodosthenous, R.; Bloomquist, T.R.; Baccarelli, A.A.; Wright, R.O.; Wright, R.J.; Lee, A.G. Associations between maternal lifetime stressors and negative events in pregnancy and breast milk-derived extracellular vesicle microRNAs in the programming of intergenerational stress mechanisms (PRISM) pregnancy cohort. Epigenetics 2021, 16, 389–404. [Google Scholar] [CrossRef]
- Leghi, G.E.; Netting, M.J.; Middleton, P.F.; Wlodek, M.E.; Geddes, D.T.; Muhlhausler, A.B.S. The impact of maternal obesity on human milk macronutrient composition: A systematic review and meta-analysis. Nutrients 2020, 12, 934. [Google Scholar] [CrossRef]
- Shah, K.B.; Chernausek, S.D.; Garman, L.D.; Pezant, N.P.; Plows, J.F.; Kharoud, H.K.; Demerath, E.W.; Fields, D.A. Human Milk Exosomal MicroRNA: Associations with Maternal Overweight/Obesity and Infant Body Composition at 1 Month of Life. Nutrients 2021, 13, 91. [Google Scholar] [CrossRef]
- Kupsco, A.; Prada, D.; Valvi, D.; Hu, L.; Petersen, M.S.; Coull, B.; Grandjean, P.; Weihe, P.; Baccarelli, A.A. Human milk extracellular vesicle miRNA expression and associations with maternal characteristics in a population-based cohort from the Faroe Islands. Sci. Rep. 2021, 11, 5840. [Google Scholar] [CrossRef]
- Cho, Y.E.; Vorn, R.; Chimenti, M.; Crouch, K.; Shaoshuai, C.; Narayanaswamy, J.; Harken, A.; Schmidt, R.; Gill, J.; Lee, H. Extracellular vesicle miRNAs in breast milk of obese mothers. Front. Nutr. 2022, 9, 976886. [Google Scholar] [CrossRef]
- Shah, K.B.; Fields, D.A.; Pezant, N.P.; Kharoud, H.K.; Gulati, S.; Jacobs, K.; Gale, C.A.; Kharbanda, E.O.; Nagel, E.M.; Demerath, E.W.; et al. Gestational Diabetes Mellitus Is Associated with Altered Abundance of Exosomal MicroRNAs in Human Milk. Clin. Ther. 2022, 44, 172–185.e171. [Google Scholar] [CrossRef]
- Zheng, Z.; Mo, J.; Lin, F.; Wang, J.; Chen, J.; Luo, H.; Liu, Y.; Su, C.; Gu, X.; Xiong, F.; et al. Milk Exosomes from Gestational Diabetes Mellitus (GDM) and Healthy Parturient Exhibit Differential miRNAs Profiles and Distinct Regulatory Bioactivity on Hepatocyte Proliferation. Mol. Nutr. Food. Res. 2023, 67, e2300005. [Google Scholar] [CrossRef]
- Riquelme, I.; Tapia, O.; Leal, P.; Sandoval, A.; Varga, M.G.; Letelier, P.; Buchegger, K.; Bizama, C.; Espinoza, J.A.; Peek, R.M.; et al. miR-101-2, miR-125b-2 and miR-451a act as potential tumor suppressors in gastric cancer through regulation of the PI3K/AKT/mTOR pathway. Cell. Oncol. 2016, 39, 23–33. [Google Scholar] [CrossRef]
- Chen, W.; Wang, X.; Yan, X.; Yu, Z.; Zhang, J.; Han, S. The emerging role of exosomes in the pathogenesis, prognosis and treatment of necrotizing enterocolitis. Am. J. Transl. Res. 2020, 12, 7020–7033. [Google Scholar]
- Mosca, F.; Giannì, M.L. Human milk: Composition and health benefits. Pediatr. Med. Chir. 2017, 39, 155. [Google Scholar] [CrossRef]
- Zonneveld, M.I.; van Herwijnen, M.J.C.; Fernandez-Gutierrez, M.M.; Giovanazzi, A.; de Groot, A.M.; Kleinjan, M.; van Capel, T.M.M.; Sijts, A.; Taams, L.S.; Garssen, J.; et al. Human milk extracellular vesicles target nodes in interconnected signalling pathways that enhance oral epithelial barrier function and dampen immune responses. J. Extracell. Vesicles 2021, 10, e12071. [Google Scholar] [CrossRef]
- Carrillo-Lozano, E.; Sebastián-Valles, F.; Knott-Torcal, C. Circulating microRNAs in Breast Milk and Their Potential Impact on the Infant. Nutrients 2020, 12, 66. [Google Scholar] [CrossRef]
- Lonnerdal, B.; Du, X.; Liao, Y.; Li, J. Human milk exosomes resist digestion in vitro and are internalized by human intestinal cells. FASEB J. 2015, 29, 121-3. [Google Scholar] [CrossRef]
- Chen, W.; Chen, X.; Qian, Y.; Wang, X.; Zhou, Y.; Yan, X.; Yu, B.; Yao, S.; Yu, Z.; Zhu, J.; et al. Lipidomic Profiling of Human Milk Derived Exosomes and Their Emerging Roles in the Prevention of Necrotizing Enterocolitis. Mol. Nutr. Food Res. 2021, 65, e2000845. [Google Scholar] [CrossRef]
- Reif, S.; Elbaum Shiff, Y.; Golan-Gerstl, R. Milk-derived exosomes (MDEs) have a different biological effect on normal fetal colon epithelial cells compared to colon tumor cells in a miRNA-dependent manner. J. Transl. Med. 2019, 17, 325. [Google Scholar] [CrossRef]
- Jiang, R.; Lönnerdal, B. Milk-Derived miR-22-3p Promotes Proliferation of Human Intestinal Epithelial Cells (HIECs) by Regulating Gene Expression. Nutrients 2022, 14, 4901. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, Y.; Xu, G.; Liu, L.; Li, H.; Li, Y.; Yin, J.; Wang, X.; Yu, Z. Human breast milk-derived exosomes through inhibiting AT II cell apoptosis to prevent bronchopulmonary dysplasia in rat lung. J. Cell. Mol. Med. 2022, 26, 4169–4182. [Google Scholar] [CrossRef]
- Azaryan, E.; Karbasi, S.; Saharkhiz, M.; Hanafi-Bojd, M.Y.; Zarban, A.; Emadian Razavi, F.; Naseri, M. Effect of HM-Exos on the migration and inflammatory response of LPS-exposed dental pulp stem cells. BMC Oral Health 2023, 23, 95. [Google Scholar] [CrossRef]
- Gao, R.; Zhang, R.; Qian, T.; Peng, X.; He, W.; Zheng, S.; Cao, Y.; Pierro, A.; Shen, C. A comparison of exosomes derived from different periods breast milk on protecting against intestinal organoid injury. Pediatr. Surg. Int. 2019, 35, 1363–1368. [Google Scholar] [CrossRef]
- He, S.; Liu, G.; Zhu, X. Human breast milk-derived exosomes may help maintain intestinal epithelial barrier integrity. Pediatr. Res. 2021, 90, 366–372. [Google Scholar] [CrossRef]
- Guo, M.M.; Zhang, K.; Zhang, J.H. Human Breast Milk-Derived Exosomal miR-148a-3p Protects Against Necrotizing Enterocolitis by Regulating p53 and Sirtuin 1. Inflammation 2022, 45, 1254–1268. [Google Scholar] [CrossRef]
- Näslund, T.I.; Paquin-Proulx, D.; Paredes, P.T.; Vallhov, H.; Sandberg, J.K.; Gabrielsson, S. Exosomes from breast milk inhibit HIV-1 infection of dendritic cells and subsequent viral transfer to CD4+ T cells. Aids 2014, 28, 171–180. [Google Scholar] [CrossRef]
- Na, R.S.; E, G.X.; Sun, W.; Sun, X.W.; Qiu, X.Y.; Chen, L.P.; Huang, Y.F. Expressional analysis of immune-related miRNAs in breast milk. Genet. Mol. Res. 2015, 14, 11371–11376. [Google Scholar] [CrossRef]
- Vélez-Ixta, J.M.; Benítez-Guerrero, T.; Aguilera-Hernández, A.; Martínez-Corona, H.; Corona-Cervantes, K.; Juárez-Castelán, C.J.; Rangel-Calvillo, M.N.; García-Mena, J. Detection and Quantification of Immunoregulatory miRNAs in Human Milk and Infant Milk Formula. BioTech 2022, 11, 11. [Google Scholar] [CrossRef]
- Zhou, Q.; Li, M.; Wang, X.; Li, Q.; Wang, T.; Zhu, Q.; Zhou, X.; Wang, X.; Gao, X.; Li, X. Immune-related microRNAs are abundant in breast milk exosomes. Int. J. Biol. Sci. 2012, 8, 118–123. [Google Scholar] [CrossRef]
- Kuo, G.; Wu, C.Y.; Yang, H.Y. MiR-17-92 cluster and immunity. J. Formos. Med. Assoc. 2019, 118, 2–6. [Google Scholar] [CrossRef]
- Ventura, A.; Young, A.G.; Winslow, M.M.; Lintault, L.; Meissner, A.; Erkeland, S.J.; Newman, J.; Bronson, R.T.; Crowley, D.; Stone, J.R.; et al. Targeted deletion reveals essential and overlapping functions of the miR-17 through 92 family of miRNA clusters. Cell 2008, 132, 875–886. [Google Scholar] [CrossRef]
- Zhang, X.; Smith, S.M.; Wang, X.; Zhao, B.; Wu, L.; Hu, X. Three paralogous clusters of the miR-17~92 family of microRNAs restrain IL-12-mediated immune defense. Cell. Mol. Immunol. 2021, 18, 1751–1760. [Google Scholar] [CrossRef]
- Ahlberg, E.; Martí, M.; Govindaraj, D.; Severin, E.; Duchén, K.; Jenmalm, M.C.; Tingö, L. Immune-related microRNAs in breast milk and their relation to regulatory T cells in breastfed children. Pediatr. Allergy Immunol. 2023, 34, e13952. [Google Scholar] [CrossRef]
- Torrez Lamberti, M.F.; Parker, L.A.; Gonzalez, C.F.; Lorca, G.L. Pasteurization of human milk affects the miRNA cargo of EVs decreasing its immunomodulatory activity. Sci. Rep. 2023, 13, 10057. [Google Scholar] [CrossRef]
- Zahoor, M.A.; Yao, X.D.; Henrick, B.M.; Verschoor, C.P.; Abimiku, A.; Osawe, S.; Rosenthal, K.L. Expression profiling of human milk derived exosomal microRNAs and their targets in HIV-1 infected mothers. Sci. Rep. 2020, 10, 12931. [Google Scholar] [CrossRef]
- Amitay, E.L.; Dubnov Raz, G.; Keinan-Boker, L. Breastfeeding, Other Early Life Exposures and Childhood Leukemia and Lymphoma. Nutr. Cancer 2016, 68, 968–977. [Google Scholar] [CrossRef]
- Amitay, E.L.; Keinan-Boker, L. Breastfeeding and Childhood Leukemia Incidence: A Meta-analysis and Systematic Review. JAMA Pediatr. 2015, 169, e151025. [Google Scholar] [CrossRef]
- Gong, Q.Q.; Quan, D.D.; Guo, C.; Zhang, C.; Zhang, Z.J. Association between maternal breastfeeding and risk of systemic neoplasms of offspring. Ital. J. Pediatr. 2022, 48, 98. [Google Scholar] [CrossRef]
- Fan, D.; Xia, Q.; Lin, D.; Ma, Y.; Rao, J.; Liu, L.; Tang, H.; Xu, T.; Li, P.; Chen, G.; et al. Role of breastfeeding on maternal and childhood cancers: An umbrella review of meta-analyses. J. Glob. Health 2023, 13, 04067. [Google Scholar] [CrossRef]
- Martin, R.M.; Gunnell, D.; Owen, C.G.; Smith, G.D. Breast-feeding and childhood cancer: A systematic review with metaanalysis. Int. J. Cancer 2005, 117, 1020–1031. [Google Scholar] [CrossRef]
- Chowdhury, R.; Sinha, B.; Sankar, M.J.; Taneja, S.; Bhandari, N.; Rollins, N.; Bahl, R.; Martines, J. Breastfeeding and maternal health outcomes: A systematic review and meta-analysis. Acta Paediatr. 2015, 104, 96–113. [Google Scholar] [CrossRef]
- Collaborative Group on Hormonal Factors in Breast Cancer. Breast cancer and breastfeeding: Collaborative reanalysis of individual data from 47 epidemiological studies in 30 countries, including 50302 women with breast cancer and 96973 women without the disease. Lancet 2002, 360, 187–195. [Google Scholar] [CrossRef]
- Badawy, A.A.; El-Magd, M.A.; AlSadrah, S.A. Therapeutic Effect of Camel Milk and Its Exosomes on MCF7 Cells In Vitro and In Vivo. Integr. Cancer Ther. 2018, 17, 1235–1246. [Google Scholar] [CrossRef]
- Martino, E.; Balestrieri, A.; Mele, L.; Sardu, C.; Marfella, R.; D’Onofrio, N.; Campanile, G.; Balestrieri, M.L. Milk Exosomal miR-27b Worsen Endoplasmic Reticulum Stress Mediated Colorectal Cancer Cell Death. Nutrients 2022, 14, 81. [Google Scholar] [CrossRef]
- Ramezani, R.; Mohammadian, M.; Hosseini, E.S.; Zare, M. The effect of bovine milk lactoferrin-loaded exosomes (exoLF) on human MDA-MB-231 breast cancer cell line. BMC. Complement. Med. Ther. 2023, 23, 228. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, Y.; Dang, H.; Wu, X. MicroRNA-148a-3p inhibits the proliferation of cervical cancer cells by regulating the expression levels of DNMT1 and UTF1. Oncol. Lett. 2021, 22, 617. [Google Scholar] [CrossRef]
- Qiao, H.; Wang, N.; Guan, Q.L.; Xie, P.; Li, X.K. miR-22-3p Suppresses Cell Proliferation and Migration of Gastric Cancer by Targeting ENO1. Altern. Ther. Health Med. 2023, 29, 278–283. [Google Scholar]
- Zheng, B.; Liang, L.; Wang, C.; Huang, S.; Cao, X.; Zha, R.; Liu, L.; Jia, D.; Tian, Q.; Wu, J.; et al. MicroRNA-148a suppresses tumor cell invasion and metastasis by downregulating ROCK1 in gastric cancer. Clin. Cancer Res. 2011, 17, 7574–7583. [Google Scholar] [CrossRef]
- Munagala, R.; Aqil, F.; Jeyabalan, J.; Agrawal, A.K.; Mudd, A.M.; Kyakulaga, A.H.; Singh, I.P.; Vadhanam, M.V.; Gupta, R.C. Exosomal formulation of anthocyanidins against multiple cancer types. Cancer Lett. 2017, 393, 94–102. [Google Scholar] [CrossRef]
- Liu, F.; Lv, D.; Wang, L.; Feng, X.; Zhang, R.; Liu, W.; Han, W. Breastfeeding and overweight/obesity among children and adolescents: A cross-sectional study. BMC Pediatr. 2022, 22, 347. [Google Scholar] [CrossRef]
- Ross, M.G.; Desai, M. Developmental programming of offspring obesity, adipogenesis, and appetite. Clin. Obstet. Gynecol. 2013, 56, 529–536. [Google Scholar] [CrossRef]
- Harder, T.; Bergmann, R.; Kallischnigg, G.; Plagemann, A. Duration of breastfeeding and risk of overweight: A meta-analysis. Am. J. Epidemiol. 2005, 162, 397–403. [Google Scholar] [CrossRef]
- Ortega-García, J.A.; Kloosterman, N.; Alvarez, L.; Tobarra-Sánchez, E.; Cárceles-Álvarez, A.; Pastor-Valero, R.; López-Hernández, F.A.; Sánchez-Solis, M.; Claudio, L. Full breastfeeding and obesity in children: A prospective study from birth to 6 years. Child. Obes. 2018, 14, 327–337. [Google Scholar] [CrossRef]
- Yan, J.; Liu, L.; Zhu, Y.; Huang, G.; Wang, P.P. The association between breastfeeding and childhood obesity: A meta-analysis. BMC Public Health 2014, 14, 1267. [Google Scholar] [CrossRef]
- Yun, B.; Kim, Y.; Park, D.J.; Oh, S. Comparative analysis of dietary exosome-derived microRNAs from human, bovine and caprine colostrum and mature milk. J. Anim. Sci. Technol. 2021, 63, 593–602. [Google Scholar] [CrossRef]
- Melnik, B.C.; Schmitz, G. MicroRNAs: Milk’s epigenetic regulators. Best. Pract. Res. Clin. Endocrinol. Metab. 2017, 31, 427–442. [Google Scholar] [CrossRef]
- Xu, Q.; Jiang, Y.; Yin, Y.; Li, Q.; He, J.; Jing, Y.; Qi, Y.T.; Xu, Q.; Li, W.; Lu, B.; et al. A regulatory circuit of miR-148a/152 and DNMT1 in modulating cell transformation and tumor angiogenesis through IGF-IR and IRS1. J. Mol. Cell Biol. 2013, 5, 3–13. [Google Scholar] [CrossRef]
- Abbas, M.A.; Al-Saigh, N.N.; Saqallah, F.G. Regulation of adipogenesis by exosomal milk miRNA. Rev. Endocr. Metab. Disord. 2023, 24, 297–316. [Google Scholar] [CrossRef]
- Ma, L.; Gilani, A.; Yi, Q.; Tang, L. MicroRNAs as Mediators of Adipose Thermogenesis and Potential Therapeutic Targets for Obesity. Biology 2022, 11, 1657. [Google Scholar] [CrossRef]
- Banday, M.Z.; Sameer, A.S.; Nissar, S. Pathophysiology of diabetes: An overview. Avicenna J. Med. 2020, 10, 174–188. [Google Scholar] [CrossRef]
- Cerf, M.E. Beta cell dysfunction and insulin resistance. Front. Endocrinol 2013, 4, 37. [Google Scholar] [CrossRef]
- Butler, P.C.; Meier, J.J.; Butler, A.E.; Bhushan, A. The replication of beta cells in normal physiology, in disease and for therapy. Nat. Clin. Pract. Endocrinol. Metab. 2007, 3, 758–768. [Google Scholar] [CrossRef]
- Wang, H.L.; Wang, L.; Zhao, C.Y.; Lan, H.Y. Role of TGF-Beta Signaling in Beta Cell Proliferation and Function in Diabetes. Biomolecules 2022, 12, 373. [Google Scholar] [CrossRef]
- Lund-Blix, N.A.; Dydensborg Sander, S.; Størdal, K.; Nybo Andersen, A.M.; Rønningen, K.S.; Joner, G.; Skrivarhaug, T.; Njølstad, P.R.; Husby, S.; Stene, L.C. Infant Feeding and Risk of Type 1 Diabetes in Two Large Scandinavian Birth Cohorts. Diabetes Care 2017, 40, 920–927. [Google Scholar] [CrossRef]
- Owen, C.G.; Martin, R.M.; Whincup, P.H.; Smith, G.D.; Cook, D.G. Does breastfeeding influence risk of type 2 diabetes in later life? A quantitative analysis of published evidence. Am. J. Clin. Nutr. 2006, 84, 1043–1054. [Google Scholar] [CrossRef]
- Melnik, B.C.; Schmitz, G. Milk Exosomal microRNAs: Postnatal Promoters of β Cell Proliferation but Potential Inducers of β Cell De-Differentiation in Adult Life. Int. J. Mol. Sci. 2022, 23, 1503. [Google Scholar] [CrossRef]
- Chen, R.; Yang, H.; Dai, J.; Zhang, M.; Lu, G.; Zhang, M.; Yu, H.; Zheng, M.; He, Q. The biological functions of maternal-derived extracellular vesicles during pregnancy and lactation and its impact on offspring health. Clin. Nutr 2023, 42, 493–504. [Google Scholar] [CrossRef]
- Gao, Y.; Zhang, R.; Dai, S.; Zhang, X.; Li, X.; Bai, C. Role of TGF-β/Smad Pathway in the Transcription of Pancreas-Specific Genes during Beta Cell Differentiation. Front. Cell Dev. Biol. 2019, 7, 351. [Google Scholar] [CrossRef]
- Alharbi, K.S. GAS5: A pivotal lncRNA in diabetes mellitus pathogenesis and management. Pathol. Res. Pract. 2024, 253, 154955. [Google Scholar] [CrossRef]
- Carter, G.; Miladinovic, B.; Patel, A.A.; Deland, L.; Mastorides, S.; Patel, N.A. Circulating long noncoding RNA GAS5 levels are correlated to prevalence of type 2 diabetes mellitus. BBA. Clin. 2015, 4, 102–107. [Google Scholar] [CrossRef]
- Giacco, F.; Brownlee, M. Oxidative stress and diabetic complications. Circ. Res. 2010, 107, 1058–1070. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Cardiovascular Diseases (CVDs). Fact Sheets. Available online: https://www.who.int/news-room/fact-sheets/detail/cardiovascular-diseases-(cvds) (accessed on 13 July 2024).
- Frąk, W.; Wojtasińska, A.; Lisińska, W.; Młynarska, E.; Franczyk, B.; Rysz, J. Pathophysiology of Cardiovascular Diseases: New Insights into Molecular Mechanisms of Atherosclerosis, Arterial Hypertension, and Coronary Artery Disease. Biomedicines 2022, 10, 1938. [Google Scholar] [CrossRef]
- Hajar, R. Risk Factors for Coronary Artery Disease: Historical Perspectives. Heart Views 2017, 18, 109–114. [Google Scholar] [CrossRef]
- El-Khuffash, A.; Lewandowski, A.J.; Jain, A.; Hamvas, A.; Singh, G.K.; Levy, P.T. Cardiac Performance in the First Year of Age among Preterm Infants Fed Maternal Breast Milk. JAMA Netw. Open 2021, 4, e2121206. [Google Scholar] [CrossRef]
- Umer, A.; Hamilton, C.; Edwards, R.A.; Cottrell, L.; Giacobbi, P., Jr.; Innes, K.; John, C.; Kelley, G.A.; Neal, W.; Lilly, C. Association between Breastfeeding and Childhood Cardiovascular Disease Risk Factors. Matern Child. Health J. 2019, 23, 228–239. [Google Scholar] [CrossRef]
- Yang, F.; Janszky, I.; Roos, N.; Li, J.; László, K.D. Prenatal Exposure to Severe Stress and Risks of Ischemic Heart Disease and Stroke in Offspring. JAMA Netw. Open 2023, 6, e2349463. [Google Scholar] [CrossRef]
- Ravelli, A.C.; van der Meulen, J.H.; Osmond, C.; Barker, D.J.; Bleker, O.P. Infant feeding and adult glucose tolerance, lipid profile, blood pressure, and obesity. Arch. Dis. Child. 2000, 82, 248–252. [Google Scholar] [CrossRef]
- El-Khuffash, A.; Jain, A.; Lewandowski, A.J.; Levy, P.T. Preventing disease in the 21st century: Early breast milk exposure and later cardiovascular health in premature infants. Pediatr. Res. 2020, 87, 385–390. [Google Scholar] [CrossRef]
- Neves, K.B.; Rios, F.J.; Sevilla-Montero, J.; Montezano, A.C.; Touyz, R.M. Exosomes and the cardiovascular system: Role in cardiovascular health and disease. J. Physiol. 2023, 601, 4923–4936. [Google Scholar] [CrossRef]
- Chen, D.; Zhang, M. GAS5 regulates diabetic cardiomyopathy via miR-221-3p/p27 axis-associated autophagy. Mol. Med. Rep. 2021, 23, 11774. [Google Scholar] [CrossRef]
- Chen, H.; Xia, W.; Hou, M. LncRNA-NEAT1 from the competing endogenous RNA network promotes cardioprotective efficacy of mesenchymal stem cell-derived exosomes induced by macrophage migration inhibitory factor via the miR-142-3p/FOXO1 signaling pathway. Stem. Cell Res. Ther 2020, 11, 31. [Google Scholar] [CrossRef]
- Liu, D.; Xing, R.; Zhang, Q.; Tian, X.; Qi, Y.; Song, H.; Liu, Y.; Yu, H.; Zhang, X.; Jing, Q.; et al. The CREG1-FBXO27-LAMP2 axis alleviates diabetic cardiomyopathy by promoting autophagy in cardiomyocytes. Exp. Mol. Med. 2023, 55, 2025–2038. [Google Scholar] [CrossRef]
- Sindi, H.A.; Russomanno, G.; Satta, S.; Abdul-Salam, V.B.; Jo, K.B.; Qazi-Chaudhry, B.; Ainscough, A.J.; Szulcek, R.; Jan Bogaard, H.; Morgan, C.C.; et al. Therapeutic potential of KLF2-induced exosomal microRNAs in pulmonary hypertension. Nat. Commun. 2020, 11, 1185. [Google Scholar] [CrossRef]
- Yin, Q.; Wang, P.; Wu, X. MicroRNA -148 alleviates cardiac dysfunction, immune disorders and myocardial apoptosis in myocardial ischemia-reperfusion (MI/R) injury by targeting pyruvate dehydrogenase kinase (PDK4). Bioengineered 2021, 12, 5552–5565. [Google Scholar] [CrossRef]
- Liu, J.; Wu, J.; Li, L.; Li, T.; Wang, J. The Role of Exosomal Non-Coding RNAs in Coronary Artery Disease. Front. Pharmacol. 2020, 11, 603104. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, L.; Li, Y.; Chen, L.; Wang, X.; Guo, W.; Zhang, X.; Qin, G.; He, S.H.; Zimmerman, A.; et al. Exosomes/microvesicles from induced pluripotent stem cells deliver cardioprotective miRNAs and prevent cardiomyocyte apoptosis in the ischemic myocardium. Int. J. Cardiol. 2015, 192, 61–69. [Google Scholar] [CrossRef]
- Rani, P.; Vashisht, M.; Golla, N.; Shandilya, S.; Onteru, S.K.; Singh, D. Milk miRNAs encapsulated in exosomes are stable to human digestion and permeable to intestinal barrier in vitro. J. Funct. Foods 2017, 34, 431–439. [Google Scholar] [CrossRef]
- Menon, R.; Debnath, C.; Lai, A.; Guanzon, D.; Bhatnagar, S.; Kshetrapal, P.; Sheller-Miller, S.; Salomon, C. Protein Profile Changes in Circulating Placental Extracellular Vesicles in Term and Preterm Births: A Longitudinal Study. Endocrinology 2020, 161, bqaa009. [Google Scholar] [CrossRef]
- Jiang, L.; Fei, H.; Jin, X.; Liu, X.; Yang, C.; Li, C.; Chen, J.; Yang, A.; Zhu, J.; Wang, H.; et al. Extracellular Vesicle-Mediated Secretion of HLA-E by Trophoblasts Maintains Pregnancy by Regulating the Metabolism of Decidual NK Cells. Int. J. Biol. Sci. 2021, 17, 4377–4395. [Google Scholar] [CrossRef]
- Wolf, T.; Baier, S.R.; Zempleni, J. The Intestinal Transport of Bovine Milk Exosomes Is Mediated by Endocytosis in Human Colon Carcinoma Caco-2 Cells and Rat Small Intestinal IEC-6 Cells. J. Nutr. 2015, 145, 2201–2206. [Google Scholar] [CrossRef]
- Ross, M.; Atalla, H.; Karrow, N.; Mallard, B.A. The bioactivity of colostrum and milk exosomes of high, average, and low immune responder cows on human intestinal epithelial cells. J. Dairy Sci. 2021, 104, 2499–2510. [Google Scholar] [CrossRef]
- Gao, H.N.; Hu, H.; Wen, P.C.; Lian, S.; Xie, X.L.; Song, H.L.; Yang, Z.N.; Ren, F.Z. Yak milk-derived exosomes alleviate lipopolysaccharide-induced intestinal inflammation by inhibiting PI3K/AKT/C3 pathway activation. J. Dairy Sci. 2021, 104, 8411–8424. [Google Scholar] [CrossRef]
- Wang, L.; Gao, R.; Li, B.; Alganabi, M.; He, W.; Shen, C.; Zhu, H.; Pierro, A. Human breast milk-derived exosomes protect against intestinal ischemia and reperfusion injury in neonatal rats. J. Pediatr. Surg. 2022, 57, 1264–1268. [Google Scholar] [CrossRef]
- Fitzgibbons, S.C.; Ching, Y.; Yu, D.; Carpenter, J.; Kenny, M.; Weldon, C.; Lillehei, C.; Valim, C.; Horbar, J.D.; Jaksic, T. Mortality of necrotizing enterocolitis expressed by birth weight categories. J. Pediatr. Surg. 2009, 44, 1072–1075; discussion 1075–1076. [Google Scholar] [CrossRef]
- Zeng, R.; Wang, J.; Zhuo, Z.; Luo, Y.; Sha, W.; Chen, H. Stem cells and exosomes: Promising candidates for necrotizing enterocolitis therapy. Stem Cell Res. Ther. 2021, 12, 323. [Google Scholar] [CrossRef]
- Chang, X.; Song, Y.H.; Xia, T.; He, Z.X.; Zhao, S.B.; Wang, Z.J.; Gu, L.; Li, Z.S.; Xu, C.; Wang, S.L.; et al. Macrophage-derived exosomes promote intestinal mucosal barrier dysfunction in inflammatory bowel disease by regulating TMIGD1 via mircroRNA-223. Int. Immunopharmacol. 2023, 121, 110447. [Google Scholar] [CrossRef]
- Cerasani, J.; Ceroni, F.; De Cosmi, V.; Mazzocchi, A.; Morniroli, D.; Roggero, P.; Mosca, F.; Agostoni, C.; Giannì, M.L. Human Milk Feeding and Preterm Infants’ Growth and Body Composition: A Literature Review. Nutrients 2020, 12, 1155. [Google Scholar] [CrossRef]
- Quigley, M.; Embleton, N.D.; McGuire, W. Formula versus donor breast milk for feeding preterm or low birth weight infants. Cochrane Database Syst. Rev. 2018, 6, Cd002971. [Google Scholar] [CrossRef]
- Hu, X.; Zhang, R.; Liang, H.; An, J.; Yang, Y.; Huo, J.; Chen, Z.; Quan, W.; Jiang, L.; Li, C.; et al. Comparison and Investigation of Exosomes from Human Amniotic Fluid Stem Cells and Human Breast Milk in Alleviating Neonatal Necrotizing Enterocolitis. Stem. Cell Rev. Rep. 2023, 19, 754–766. [Google Scholar] [CrossRef]
- Pisano, C.; Galley, J.; Elbahrawy, M.; Wang, Y.; Farrell, A.; Brigstock, D.; Besner, G.E. Human Breast Milk-Derived Extracellular Vesicles in the Protection Against Experimental Necrotizing Enterocolitis. J. Pediatr. Surg. 2020, 55, 54–58. [Google Scholar] [CrossRef]
- Cornick, S.; Tawiah, A.; Chadee, K. Roles and regulation of the mucus barrier in the gut. Tissue Barriers 2015, 3, e982426. [Google Scholar] [CrossRef]
- Liu, D.; Xu, Y.; Feng, J.; Yu, J.; Huang, J.; Li, Z. Mucins and Tight Junctions are Severely Altered in Necrotizing Enterocolitis Neonates. Am. J. Perinatol. 2021, 38, 1174–1180. [Google Scholar] [CrossRef]
- Tong, L.; Hao, H.; Zhang, X.; Zhang, Z.; Lv, Y.; Zhang, L.; Yi, H. Oral Administration of Bovine Milk-Derived Extracellular Vesicles Alters the Gut Microbiota and Enhances Intestinal Immunity in Mice. Mol. Nutr. Food Res. 2020, 64, e1901251. [Google Scholar] [CrossRef]
- Paradis, T.; Bègue, H.; Basmaciyan, L.; Dalle, F.; Bon, F. Tight Junctions as a Key for Pathogens Invasion in Intestinal Epithelial Cells. Int. J. Mol. Sci. 2021, 22, 2506. [Google Scholar] [CrossRef]
- Chiba, T.; Takaguri, A.; Kooka, A.; Kowatari, K.; Yoshizawa, M.; Fukushi, Y.; Hongo, F.; Sato, H.; Fujisawa, M.; Wada, S.; et al. Suppression of milk-derived miR-148a caused by stress plays a role in the decrease in intestinal ZO-1 expression in infants. Clin. Nutr. 2022, 41, 2691–2698. [Google Scholar] [CrossRef]
- Shang, J.; Ning, J.; Bai, X.; Cao, X.; Yue, X.; Yang, M. Identification and analysis of miRNAs expression profiles in human, bovine, and donkey milk exosomes. Int. J. Biol. Macromol 2023, 252, 126321. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Maternal-Associated Factor | Mechanism of Action | Reference |
|---|---|---|
| Mode of birth | ||
| Cesarean section | Change in miRNA expression levels | [105] |
| Greater expression of miRNA-320 than miRNA-148a in colostrum from mothers not receiving exogenous oxytocin | [102] | |
| Lower levels of miRNA-148a and miRNA-125b in transition and mature human breast milk | [106] | |
| Vaginal birth | Increased in miRNA-148a and miRNA-30 levels, and decreased miRNA-320 levels in colostrum due to increased levels of exogenous oxytocin | [105] |
| Maternal nutrition | ||
| High-fat or high-carbohydrate diets with similar energy and protein | Increased expression of miRNA-67 and miRNA-27 | [107] |
| Association with miRNA-148a-5p and miRNA-146b-5p levels related with maternal weight | [83,86,109] | |
| Animals fed the obesogenic diet model | Increased levels of miRNA-222, and decreased levels of miRNA-200 and miRNA-26 | [111] |
| Maternal stress (psychological distress) | Interaction with epigenetic regulation of pathways such as fatty acid metabolism, steroid biosynthesis, and the Hippo signaling pathway | [113] |
| Maternal weight | ||
| Overweight/obesity | Decreased levels of miRNA-148a and miRNA-30b which are linked to infant anthropometric measurements in the breast milk of overweight/obese women | [115] |
| Normal body weight | Decreased levels of leptin, adiponectin, and partially different miRNA levels (miR-17, miR-103, miR-181a, miR-let7c, miR-222, miR-146b) in mothers with normal body weight | [109] |
| Obesity | Changes in 19 miRNAs, including miRNA-575, miRNA-630, miRNA-642a-3p, and miRNA-652-5p, related with neurological diseases and psychological disorders in the breast milk of obese women | [114] |
| Maternal chronic stress | ||
| Gestational diabetes mellitus | Decreased levels of miRNA-148a, miRNA-30b, miRNA-let-7a, and miRNA-let-7d levels in mothers with GDM, correlated positively with maternal obesity | [112,115] |
| T1DM | Altered levels of several miRNAs related with proinflammatory cytokine production in the breast milk of mothers with T1DM | [101] |
| Type of Milk and Exosome | Cancer Type | Possible Mechanism | Reference |
|---|---|---|---|
| Human breast milk exosome TGF β2 | MCF7 breast cancer cells and MCF10A fibrocystic normal cells (derived from woman with benign breast disease) | TGF β2 dose-dependent (high dose); Affects cell morphology and actin cytoskeleton Increases SMA and vimentin levels and decreases E-cadherin levels Promotes EMT | [55] |
| Human breast milk exosomes | LS123 colon cancer cells | Milk-derived exosomes can penetrate both malignant and normal cells, and change the mRNA expression profiles of those cells Following incubation, the miRNA-148a expression was upregulated, decreasing tumor cell proliferation | [124] |
| Camel milk exosomes | MCF7 breast cancer cells | Inhibited MCF7 cell proliferation Tumor weight decreased Tumor DNA damage increased Higher caspase-3 activity, increased Bax, and downregulated Bcl2 gene expression all pointed to cancer cell death via apoptosis Inhibition of malondialdehyde levels and inducible nitric oxide synthase mRNA levels; higher levels of catalase, superoxide dismutase, and glutathione peroxidase in tumor tissues Decreased NF-κB, IL-1β, MMP9, VEGF, and ICAM1 gene expression levels Reduced metastasis and angiogenesis in tumor tissues | [148] |
| Buffalo milk exosomal miRNA-27b | HCT116 and HT-29 colorectal cancer cell | miRNA-27b transfection increased cytotoxic effects Lysosome accumulation and the amount of reactive oxygen species in the mitochondria were increased in miRNA-27b+ cells miRNA-27b promoted mitochondrial stress and apoptotic death | [149] |
| Bovine milk lactoferrin-loaded exosomes (exoLF) | MDA-MB-231 breast cancer cell line | Cytotoxic on cancer cell while normal mesenchymal stem cells remained viable A decrease in anti-apoptotic protein Bcl-2 levels and an increase in the pro-apoptotic protein Bid levels following the exoLF therapy | [150] |
| Bovine milk exosomes | A549 and H1299 lung cancer HCT116 colon cancer PC3 and DU145 prostate cancer MDA-MB-231 and MCF7 breast cancer PANC1 and Mia PaCa2 pancreatic cancer OVCA432 ovarian cancer | Inhibits the proliferation of certain types of human cancer cells (antiproliferative effect) | [154] |
| Anthocyanidin-loaded exosomes (ExoAnthos) | A549 and H1299 lung cancer HCT116 colon cancer PC3 and DU145 prostate cancer MDA-MB-231 and MCF7 breast cancer PANC1 and Mia PaCa2 pancreatic cancer OVCA432 ovarian cancer | Antiproliferative effects ExoAnthos exhibited a dose-dependent inhibition of TNFα-induced and NF-κB activity in breast (MCF7) and lung (H1299) cancer cells | [154] |
| Type of Exosome | Cell Culture or Experimental Animal | Possible Mechanism | Reference |
|---|---|---|---|
| Resistance to digestion | |||
| Human milk miRNA 22-3-p | In vitro human intestinal crypt-like cells (HIEC) | Exosomes remained viable in in vitro digestion and absorbed by ICs. Overall, 288 mature miRNAs were isolated from samples of exosomes in the intestinal epithelium. Hsa-miRNA-22-3p was the most prevalent miRNA. Both digested and undigested exosomes entered the cells of the human intestinal crypt. | [53] |
| miRNA bovine milk formula | In vitro HIEC-6 In vivo newborn piglets | Milk-derived miRNAs survive gastrointestinal passage in newborns, accumulating in preterm piglets’ ICs after enteral feeding with bovine colostrum/formula. In piglets, supplementing colostrum with cel-miR-39-5p/-3p raised blood concentrations of cel-miR-39-3p and AGO2 loading in ICs. This shows that miRNA signaling could be transmitted vertically from milk to the newborn digestive tract. | [100] |
| Sahiwal cow-derived exosomal miRNA | In vitro Caco-2 cell | Exosomes isolated from in vitro digested milk and their respective water controls (miRNA-182-5p, miRNA-148a, miRNA-25, miRNA-21, and miRNA-2478) were among the immune-related miRNAs that did not significantly differ. Milk exosomal miRNAs can cross the intestinal barrier meaning these are resistant to digestion. Transepithelial migration via the Caco-2 monolayer. | [193] |
| Holstein cow-derived exosomal protein Colostrum Milk | In vitro Caco-2 cell | The study examined the expression of exosome surface indicators in the colostrum and milk of H, A, and L immune responder cows, revealing their ability to be absorbed by human intestinal epithelial cells. Co-incubation with colostrum and milk exosomes from H responder cows resulted in significantly higher metabolic activity compared to L responder exosomes. Milk exosomes, unlike colostrum exosomes from L responders, did not activate the caspase 3 pathway in Caco-2 cells, as evidenced by significantly lower caspase 3 activity, an indicator of apoptosis. | [197] |
| Protective effects on epithelial function | |||
| Human breast milk exosomes | In vitro ISCs | The vitality of ISCs was significantly improved by adding exosomes to ISCs exposed to H2O2. Marked upregulation of the Wnt/b-catenin axis genes Cyclin D1, c-Myc, and Axin2 mRNA expression in ISCs treated with exosomes (p < 0.05 for all). The inclusion of carnosic acid, a specific Wnt/b-catenin signaling inhibitor, markedly decreased the viability of the cells. ISC was protected from oxidative stress damage. | [52] |
| Human breast milk exosomes Colostrum Transition milk Ripe milk | In vivo C57BL/6 mice pups Ex vivo intestinal organoid model | Intestinal organoids shrank in size when exposed to LPS, which also promoted intestinal regeneration and inflammation by TNF-α and TLR4 expression upregulation. Human-milk-derived exosomes protected the intestinal organoids from LPS-induced epithelial damage. Colostrum was more effective protective agent, suppressed LPS-induced injury and reduced inflammation (TNF-α and TLR4 expression reduction) than other term milks. | [128] |
| Holstein cow exosomal proteins Colostrum Milk | In vitro Caco-2 cell culture | Exosomes from cow colostrum and milk preserved Caco-2 metabolic function and did not harm these cells. Colostrum suppressed the activity of the apoptosis marker caspase 3. | [197] |
| Exosomal proteins yak vs. cow milk | In vitro IEC-6 | The proteomics examination found 58 higher and 334 lower expressed proteins in yak milk exosomes compared to cow milk exosomes. Compared with cow milk exosomes, yak milk exosomes more efficiently activate the PI3K/AKT/C3 signaling pathway, thereby increasing IEC-6 survival and decreasing intestinal inflammation incidence and severity. Milk exosomes achieve this effect in two ways, which are attenuated LPS-induced intestinal inflammation and decreased inflammatory cytokine levels | [198] |
| Human breast milk exosomes | In vivo Sprague–Dawley rats | Exosomes administration decreased the damage caused by intestinal IR. In rats with IR, exosome injection resulted in a considerable TNF-α downregulation. Exosomes promoted intestinal regeneration, which decreased IR-mediated damage. Ki67 protein expression was significantly elevated with exosomes, suggesting enhanced IEC proliferation. Exosomes mitigated IR-induced intestinal damage by boosting intestinal regeneration and lowering mucosal inflammation. | [199] |
| Possible effects on NEC | |||
| Human breast milk exosomes miRNA125b miRNA148B | In vitro IECs | miRNA125b suppressed p53, which is an apoptosis-inducing agent. miRNA148b was associated to IEC development and survival. Exosomes prevented IECs from destruction. Exosomes supported gastrointestinal and immune system development. | [40] |
| Human breast milk exosomes miRNA 22-3-p | In vitro HIEC | In the intestinal epithelium, 288 mature miRNAs from all 24 exosome samples were detected. Exosomes provided oxidative stress resistance for human intestinal crypt-like cells. | [53] |
| Human breast milk exosomes Exosomal peptides Preterm milk Term milk | In vitro FHC In vivo Sprague–Dawley rats | Compared with term exosome application, preterm exosome application significantly increased FHC proliferation and cell migration. In the in vivo study, three proteins were upregulated about the topic. Two of them with immunomodulatory, anti-inflammatory and antimicrobial properties were lactoferrin and one of them with potentially regenerating the damaged intestinal mucosa properties was one lactoadherin MFGE8. | [54] |
| Human breast milk exosomes Raw milk Pasteurized milk | In vitro Ex vivo C57BL/6 mice | Administration of exosomes from raw and pasteurized human breast milk during NEC decreased IL-6 expression and MPO activity, and increased goblet cell count. MUC2 expression was elevated by both milk, with no discernible difference in MUC2 expression between the two. NEC-induced inflammatory response was equally attenuated by raw and pasteurized human breast milk exosomes. Protected goblet cells and mucosa production. | [91] |
| Human breast milk exosomes miRNA-148a-3p | In vitro Normal colonic epithelial cells Colonic tumor cells | Exosomes in normal cells downregulated PTEN, a miRNA-148a target, inhibiting proliferation and DNMT1 expression, inducing alterations associated with epithelial mesenchymal transition. Exosomes did not exhibit this effect on tumour cells. | [124] |
| Human breast milk exosomes Preterm milk Term milk | In vitro Caco-2 cell line In vivo C57BL/6 mice | LPS damage severely damaged the intestinal mucosa in pups, whereas human-milk-derived exosomes offered protection. NEC and NEC + milk without exosomes showed higher proinflammatory cytokine IL-6β and TNF-α levels. NEC + milk without exosomes showed lower ZO-1, claudin-1, and occludin levels. | [129] |
| Human breast milk exosomes miRNA-148a-3p | In vitro Small IEC line (IEC-6) In vivo C57BL/6 mice | By reducing target p53 expression, miRNA-148a-3p increased SIRT1 levels. The in vivo use of miRNA-148a-3p agomir showed a similar protective effect as human breast milk exosomes. | [130] |
| Human breast milk exosomes AFSC exosomes | In vitro IEC-6 rat crypt epithelial cells IEC-8 rat ileal epithelial cells In vivo C57BL/6 mice pups | Human AFSC exosomes vs. Human breast milk exosomes (in vitro): AFSC exosomes selectively controlled IEC migration, whereas human breast milk exosomes preferentially suppressed the inflammatory response of IECs. Human breast milk exosomes (in vivo): The number of ileal crypts was markedly recovered in vivo following human milk exosome administration compared with that using AFSC exosomes. | [205] |
| Human breast milk exosomes | In vitro IEC-6 rat small intestinal epithelial cells In vivo Sprague–Dawley rats | Intravenous or enteral exosome therapy significantly decreased the frequency and intensity of experimental NEC (29% vs. 11.9%) (in vivo), while safeguarding IECs from damage by anti-apoptopic and pro-proliferative properties (in vitro). | [206] |
| Human breast milk exosomes mir-148a | In vitro Caco-2 cell In vivo CD1 mice | Caco-2 cells showed that ZO-1 protein levels were markedly increased, whereas DNMT1 protein levels were significantly decreased upon miRNA-148a overexpression (in vitro). With an upward trend in DNMT1 levels in infant intestines, stress-induced suppression of miRNA-148a expression in mothers’ milk may result in a decrease in intestinal ZO-1 concentration (in vivo). | [211] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Çelik, E.; Cemali, Ö.; Şahin, T.Ö.; Deveci, G.; Biçer, N.Ç.; Hirfanoğlu, İ.M.; Ağagündüz, D.; Budán, F. Human Breast Milk Exosomes: Affecting Factors, Their Possible Health Outcomes, and Future Directions in Dietetics. Nutrients 2024, 16, 3519. https://doi.org/10.3390/nu16203519
Çelik E, Cemali Ö, Şahin TÖ, Deveci G, Biçer NÇ, Hirfanoğlu İM, Ağagündüz D, Budán F. Human Breast Milk Exosomes: Affecting Factors, Their Possible Health Outcomes, and Future Directions in Dietetics. Nutrients. 2024; 16(20):3519. https://doi.org/10.3390/nu16203519
Chicago/Turabian StyleÇelik, Elif, Özge Cemali, Teslime Özge Şahin, Gülsüm Deveci, Nihan Çakır Biçer, İbrahim Murat Hirfanoğlu, Duygu Ağagündüz, and Ferenc Budán. 2024. "Human Breast Milk Exosomes: Affecting Factors, Their Possible Health Outcomes, and Future Directions in Dietetics" Nutrients 16, no. 20: 3519. https://doi.org/10.3390/nu16203519
APA StyleÇelik, E., Cemali, Ö., Şahin, T. Ö., Deveci, G., Biçer, N. Ç., Hirfanoğlu, İ. M., Ağagündüz, D., & Budán, F. (2024). Human Breast Milk Exosomes: Affecting Factors, Their Possible Health Outcomes, and Future Directions in Dietetics. Nutrients, 16(20), 3519. https://doi.org/10.3390/nu16203519