Manganese: From Soil to Human Health—A Comprehensive Overview of Its Biological and Environmental Significance



Abstract
1. Introduction
2. The Manganese Cycle
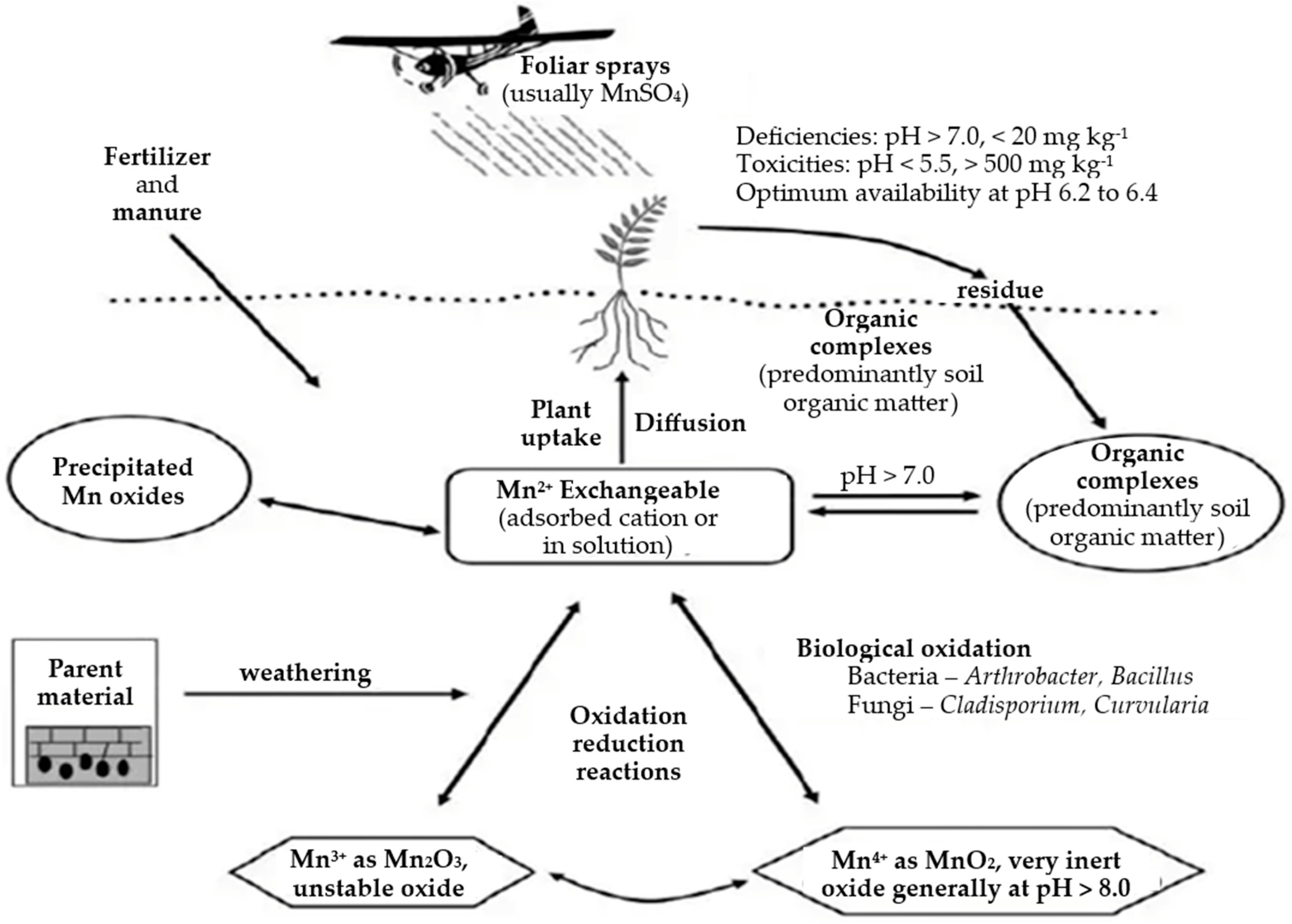
2.1. Manganese in Environment
2.1.1. Manganese Levels in Soil
2.1.2. Manganese in Water
3. Manganese in Plants
3.1. Plant Manganese Transport and Accumulation
3.2. Manganese’s Biochemical Role in Plants
3.3. Manganese Deficiency in Plants

3.4. Manganese Toxicity
3.5. Manganese’s Impact on Agricultural Production
3.6. Interactions of Manganese with Other Elements
3.7. Manganese Availability as a Function of Organic Manure and Manganese Fertilizer Applications
4. Manganese in Human Nutrition
4.1. Manganese in the Human Diet
4.2. Manganese in Food
Sources of Manganese
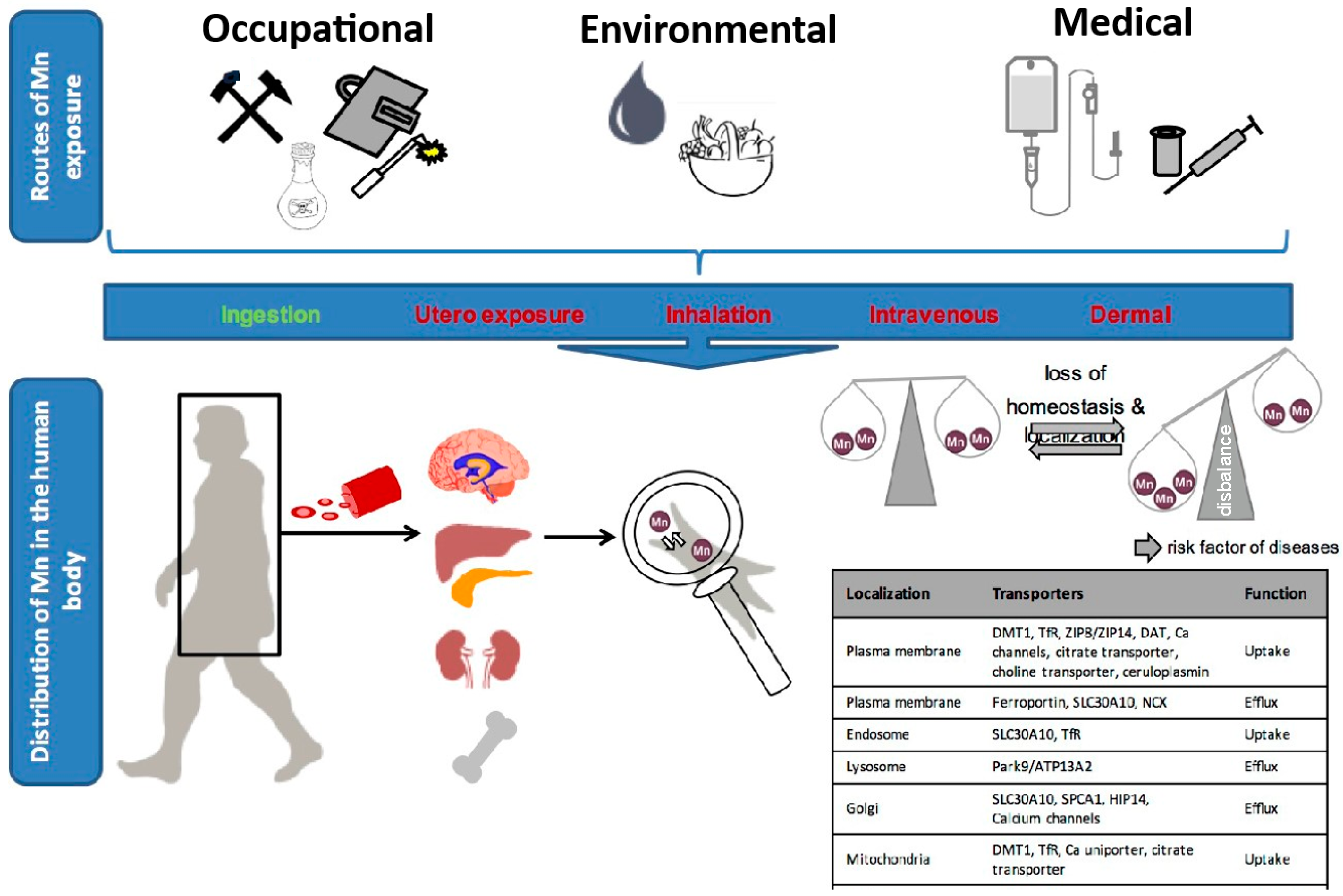
4.3. Human Manganese Uptake and Distribution
4.4. Manganese Exposure and Absorption Pathways
4.5. Ingestion
4.6. Prenatal Exposure
4.7. Inhalation
4.8. Intravenous and Cutaneous Exposure
5. Pharmacokinetics of Manganese
5.1. Manganism, a Neurodegenerative Condition
5.2. Manganese Toxicity Symptoms
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yokel, R.A. Manganese flux across the blood–brain barrier. NeuroMolecular Med. 2009, 11, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Dey, S.; Tripathy, B.; Kumar, M.S.; Das, A.P. Ecotoxicological consequences of manganese mining pollutants and their biological remediation. Environ. Chem. Ecotoxicol. 2023, 5, 55–61. [Google Scholar] [CrossRef]
- Graham, R.D.; Webb, M.J. Micronutrients and disease resistance and tolerance in plants. Micronutr. Agric. 2018, 4, 329–370. [Google Scholar] [CrossRef]
- Huber, D.M.; Graham, R.D. The role of nutrition in crop resistance and tolerance to disease. In Mineral Nutrition of Crops: Fundamental Mechanisms and Implications; Rengel, Z., Ed.; Food Products Press: New York, NY, USA, 1999; pp. 205–226. [Google Scholar]
- Heckman, J.R.; Clarke, B.B.; Murphy, J.A. Optimizing manganese fertilization for the suppression of take-all patch disease on creeping bentgrass. Crop. Sci. 2003, 43, 1395–1398. [Google Scholar] [CrossRef]
- Eskandari, S.; Sharifnabi, B. Foliar spray time affects the efficacy of applied manganese on enhancing cucumber resistance to Podosphaera fuliginea. Sci. Hortic. 2020, 261, 108780. [Google Scholar] [CrossRef]
- Khoshru, B.; Mitra, D.; Nosratabad, A.F.; Reyhanitabar, A.; Mandal, L.; Farda, B.; Djebaili, R.; Pellegrini, M.; Guerra-Sierra, B.E.; Senapati, A.; et al. Enhancing manganese availability for plants through microbial potential: A sustainable approach for improving soil health and food security. Bacteria 2023, 2, 129–141. [Google Scholar] [CrossRef]
- Rengel, Z.; Cakmak, I.; White, J.P. (Eds.) Marschner’s Mineral Nutrition of Plants; Elsevier Science Publishing: Amsterdam, The Netherlands, 2022. [Google Scholar] [CrossRef]
- Diedrick, K. Manganese fertility in soybean production. Pioneer Hi-Bred Agron. Sci. 2010, 20, 23–34. [Google Scholar]
- Millaleo, R.; Reyes-Diaz, M.; Ivanov, A.; Mora, M.; Alberdi, M. Manganese as essential and toxic element for plants: Transport, accumulation and resistance mechanisms. J. Soil Sci. Plant Nutr. 2010, 10, 470–481. [Google Scholar] [CrossRef]
- Moss, A.J. Use of vitamin and mineral supplements in the United States: Current uses, types of products, and nutrients. In Advance Data, Vital and Health Statistics of the National Center for Health Statistics; National Center for Health Statistics: Hyattsville, MD, USA, 1989; No. 174. [Google Scholar]
- Bornhorst, J.; Wehe, C.A.; Huwel, S.; Karst, U.; Galla, H.J.; Schwerdtle, T. Impact of manganese on and transfer across blood-brain and blood-cerebrospinal fluid barrier in vitro. J. Biol. Chem. 2012, 287, 17140–17151. [Google Scholar] [CrossRef]
- Rubin, L.L.; Staddon, J.M. The cell biology of the blood-brain barrier. Annu. Rev. Neurosci. 1999, 22, 11–28. [Google Scholar] [CrossRef]
- Yoon, M.; Schroeter, J.D.; Nong, A.; Taylor, M.D.; Dorman, D.C.; Andersen, M.E.; Clewell, H.J. Physiologically based pharmacokinetic modeling of fetal and neonatal manganese exposure in humans: Describing manganese homeostasis during development. Toxicol. Sci. 2011, 122, 297–316. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, S.K.; Bräuer, S.L. 7. Microbial diversity and manganese cycling: A review of manganese-oxidizing microbial cave communities. In Microbial Life of Cave Systems; De Gruyter: Göttingen, Germany, 2015; pp. 137–160. [Google Scholar] [CrossRef]
- Anonymous. 2024. Available online: https://www.slideserve.com/keren/manganese-cycle (accessed on 20 February 2024).
- Carmichael, S.K.; Zorn, B.T.; Santelli, C.M.; Roble, L.A.; Carmichael, M.J.; Bräuer, S.L. Nutrient input influences fungal community composition and size and can stimulate manganese (II) oxidation in caves. Environ. Microbiol. Rep. 2015, 7, 592–605. [Google Scholar] [CrossRef] [PubMed]
- Paula, C.C.; Bichuette, M.E.; Seleghim, M.H. Nutrient availability in tropical caves influences the dynamics of microbial biomass. MicrobiologyOpen 2020, 9, e1044. [Google Scholar] [CrossRef] [PubMed]
- Malakouti, M.J.; Tehrani, M.H. Effect of Micronutrients on the Yield and Quality of Agricultural Products: Micro-Nutrients with Macro-Effects; Tarbiat Modares University: Tehran, Iran, 1999. [Google Scholar]
- Schulte, E.E.; Kelling, K.A. Soil and Applied Manganese. Understanding Plant Nutrients; A2526; University of Wisconsin-Madison and University of Wisconsin-Extension: Madison, WI, USA, 1999. [Google Scholar]
- Bean, E.L. Potable water–quality goals. J. AWWA 1974, 66, 221–230. [Google Scholar] [CrossRef]
- Ginige, M.P.; Wylie, J.; Plumb, J. Influence of biofilms on iron and manganese deposition in drinking water distribution systems. Biofouling 2011, 27, 151–163. [Google Scholar] [CrossRef]
- ATSDR. Toxicological Profile for Manganese; U.S. Department of Health and Human Services. Public Health Service. Agency for Toxic Substances and Disease Registry: Atlanta, GA, USA, 2012; 556p.
- Barceloux, D.G.; Barceloux, D. Manganese. J. Toxicol. Clin. Toxicol. 1999, 37, 293–307. [Google Scholar] [CrossRef]
- Leahy, P.; Thompson, T. Overview of the National Water-Quality Assessment Program; Open-File Report; U.S. Geological Survey: Reston, VA, USA, 1994. [CrossRef]
- USGS. US Geological Survey National Water Quality Assessment Data Warehouse. 2001. Available online: https://www.usgs.gov/mission-areas/water-resources/science/national-water-quality-assessment-nawqa (accessed on 22 August 2024).
- Gherardi, M.J.; Rengel, Z. Deep placement of manganese fertiliser improves sustainability of Lucerne growing on bauxite residue: A glasshouse study. Plant Soil 2003, 257, 85–95. [Google Scholar] [CrossRef]
- Mills, R.F.; Doherty, M.L.; López-Marqués, R.L.; Weimar, T.; Dupree, P.; Palmgren, M.G.; Pittman, J.K.; Williams, L.E. ECA3, a golgi-localized P2A-type ATPase, plays a crucial role in manganese nutrition in arabidopsis. Plant Physiol. 2007, 146, 116–128. [Google Scholar] [CrossRef]
- Humphries, J.; Stangoulis, J.; Graham, R. Manganese. In Handbook of Plant Nutrition; Barker, A., Pilbeam, D., Eds.; Taylor and Francis: New York, NY, USA, 2007; pp. 351–366. [Google Scholar]
- Clarkson, D.T. The uptake and translocation of manganese by plant roots. In Manganese in Soils and Plants; Springer: Dordrecht, The Netherlands, 1988; pp. 101–111. [Google Scholar] [CrossRef]
- Maas, E.V.; Moore, D.P.; Mason, B.J. Manganese absorption by excised Barley roots. Plant Physiol. 1968, 43, 527–530. [Google Scholar] [CrossRef]
- Takahashi, M.; Sugiura, M. Strategies for uptake of a soil micronutrient, manganese, by plant roots. Riken Rev. 2001, 35, 76–77. [Google Scholar]
- Zhang, H.; Lombi, E.; Smolders, E.; McGrath, S. Kinetics of Zn release in soils and prediction of Zn concentration in plants using diffusive gradients in thin films. Environ. Sci. Technol. 2004, 38, 3608–3613. [Google Scholar] [CrossRef] [PubMed]
- Rengel, Z.; Marschner, P. Nutrient availability and management in the rhizosphere: Exploiting genotypic differences. New Phytol. 2005, 168, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, M.J.; Sharma, A. Manganese in cell metabolism of higher plants. Bot. Rev. 1991, 57, 117–149. [Google Scholar] [CrossRef]
- Jackson, C.; Dench, J.; Moore, A.L.; Halliwell, B.; Foyer, C.H.; Hall, D.O. Subcellular localisation and identification of superoxide Dismutase in the leaves of higher plants. Eur. J. Biochem. 1978, 91, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Uehara, K.; Fujimoto, S.; Taniguchi, T.; Nakai, K. Studies on Violet-colored acid Phosphatase of sweet potato. J. Biochem. 1974, 75, 639–649. [Google Scholar] [CrossRef]
- Burnell, J.N. The biochemistry of manganese in plants. In Manganese in Soils and Plants; Springer: Dordrecht, The Netherlands, 1988; pp. 125–137. [Google Scholar] [CrossRef]
- Anderson, J.M.; Pyliotis, N. Studies with manganese-deficient spinach chloroplasts. Biochim. Biophys. Acta (BBA)-Bioenerg. 1969, 189, 280–293. [Google Scholar] [CrossRef]
- Ness, P.J.; Woolhouse, H.W. RNA synthesis in PhaseolusChloroplasts. J. Exp. Bot. 1980, 31, 235–245. [Google Scholar] [CrossRef]
- Mousavi, S.R.; Shahsavari, M.; Rezaei, M. A General Overview on Manganese (Mn) Importance for Crops Production. Aust. J. Basic Appl. Sci. 2011, 5, 1799–1803. [Google Scholar]
- Ndakidemi, P.A.; Bambara, S.J.; Makoi, H.J.R. Micronutrient uptake in common bean (Phaseolus vulgaris L.) as affected by Rhizobium inoculation, and the supply of molybdenum and lime. Plant Omics J. Plant Biol. Omics 2011, 4, 40–52. [Google Scholar]
- Ahangar, A.G.; Karimian, N.; Abtahi, A.; Assad, M.T.; Emam, Y. Growth and manganese uptake by soybean in highly calcareous soils as affected by native and applied manganese and predicted by nine different extractants. Commun. Soil Sci. Plant Anal. 1995, 26, 1441–1454. [Google Scholar] [CrossRef]
- Polle, A.; Chakrabarti, K.; Chakrabarti, S.; Seifert, F.; Schramel, P.; Rennenberg, H. Antioxidants and manganese deficiency in needles of Norway spruce (Picea abies L.) trees. Plant Physiol. 1992, 99, 1084–1089. [Google Scholar] [CrossRef] [PubMed]
- Ohki, K.; Wilson, D.O.; Anderson, O.E. Manganese deficiency and toxicity sensitivities of soybean cultivars. Agron. J. 1980, 72, 713–716. [Google Scholar] [CrossRef]
- Messant, M.; Hani, U.; Hennebelle, T.; Guérard, F.; Gakière, B.; Gall, A.; Thomine, S.; Krieger-Liszkay, A. Manganese concentration affects chloroplast structure and the photosynthetic apparatus in Marchantia polymorpha. Plant Physiol. 2023, 192, 356–369. [Google Scholar] [CrossRef]
- Alejandro, S.; Höller, S.; Meier, B.; Peiter, E. Manganese in plants: From acquisition to subcellular allocation. Front. Plant Sci. 2020, 11, 517877. [Google Scholar] [CrossRef]
- Zewide, I.; Sherefu, A. Review paper on effect of micronutrients for crop production. Nutr. Food Process. 2021, 4, 01–08. [Google Scholar] [CrossRef] [PubMed]
- TEAGASC. Manganese: Manganese Deficiency in CROPS. 2017. Available online: https://www.teagasc.ie/crops/soil--soil-fertility/nutrient-deficiencies/manganese/ (accessed on 6 September 2024).
- Jordan, D. Manganese Deficiency in Peanuts. Peanut Notes No. 140. 2024. Available online: https://peanut.ces.ncsu.edu/2024/07/manganese-deficiency-in-peanuts-peanut-notes-no-140-2024/ (accessed on 6 September 2024).
- Longnecker, N.E.; Graham, R.D.; Card, G. Effects of manganese deficiency on the pattern of tillering and development of Barley (Hordeum vulgare c.v. Galleon). Field Crops Res. 1991, 28, 85–102. [Google Scholar] [CrossRef]
- Sharma, C.P.; Sharma, P.N.; Chatterjee, C.; Agarwala, S.C. Manganese deficiency in maize affects pollen viability. Plant Soil 1991, 138, 139–142. [Google Scholar] [CrossRef]
- Smith, M.W.; Cheary, B.S.; Landgraf, B.S. Manganese deficiency in pecan. HortScience 2001, 36, 1075–1076. [Google Scholar] [CrossRef]
- Batey, T. Manganese and boron deficiency in trace elements in soil and crops. Tech. Bull. 1971, 21, 137–148. [Google Scholar]
- Wissemeier, A.H.; Horst, W.J. Effect of light intensity on manganese toxicity symptoms and callose formation in cowpea (Vigna unguiculata (L.) Walp.). Plant Soil 1992, 143, 299–309. [Google Scholar] [CrossRef]
- Reichman, S.M. The Responses of Plants to Metal Toxicity: A Review Focusing on Copper, Manganese, and Zinc; Australian Minerals and Energy Environment Foundation: Melbourne, Australia, 2002; ISBN 1-876205-13-X.
- Bachman, G.R.; Miller, W.B. Iron chelate inducible iron/manganese toxicity in zonal geranium. J. Plant Nutr. 1995, 18, 1917–1929. [Google Scholar] [CrossRef]
- Wu, J.D. Effect of manganese excess on the soybean plant cultivated under various growth conditions. J. Plant Nutr. 1994, 17, 991–1003. [Google Scholar] [CrossRef]
- Brown, J.C.; Ambler, J.E.; Chaney, R.L.; Foy, C.D. Differential responses of plant genotypes to micronutrients. In Micronutrients in Agriculture; Soil Science Society of America: Madison, WI, USA, 1972; pp. 389–418. [Google Scholar]
- Edwards, D.G.; Asher, C.J. Tolerance of crop and pasture species to manganese toxicity. In Proceedings of the Ninth Plant Nutrition Colloquium, Warwick, UK, 22–27 August 1982; pp. 145–150. [Google Scholar]
- Rezai, K.; Farboodnia, T. The response of pea plant to manganese toxicity in solution culture. J. Agric. Sci. 2008, 3, 248–251. [Google Scholar]
- Terry, N.; Evans, P.S.; Thomas, D.E. Manganese toxicity effects on leaf cell multiplication and expansion and on dry matter yield of sugar Beets1. Crop. Sci. 1975, 15, 205–208. [Google Scholar] [CrossRef]
- Guan, W.; Egel, D. Manganese Toxicity on Cantaloupes. Purdue University, 2017. Available online: https://vegcropshotline.org/article/manganese-toxicity-on-cantaloupes/ (accessed on 7 August 2024).
- Rabêlo, V.M.; Magalhães, P.C.; Bressanin, L.A.; Carvalho, D.T.; Reis, C.O.; Karam, D.; Doriguetto, A.C.; Santos, M.H.; Santos Filho, P.R.; Souza, T.C. The foliar application of a mixture of semisynthetic chitosan derivatives induces tolerance to water deficit in maize, improving the antioxidant system and increasing photosynthesis and grain yield. Sci. Rep. 2019, 9, 8164. [Google Scholar] [CrossRef]
- Mousavi, S.R.; Galavi, M.; Ahmadvand, G. Effect of zinc and manganese foliar application on yield, quality, and enrichment on potato (Solanum tuberosum L.). Asian J. Plant Sci. 2007, 6, 1256–1260. [Google Scholar] [CrossRef]
- Hiller, L.K. Foliar Fertilization Bumps Potato Yields in Northwest: Rate and Timing of Application, Plus Host of Other Considerations, Are Critical in Applying Foliars to Potatoes. Fluid J. 1995, 10, 28–30. [Google Scholar]
- Brown, S.; Walworth, J.L. Crop production and soil management series. In Field Crop Fertilizer Recommendations for Alaska Potatoes; University of Alaska Fairbanks, Cooperative Extension Service: Fairbanks, AK, USA, 1998. [Google Scholar]
- Bansal, R.L.; Nayyar, V.K. Differential tolerance of soybean (Glycine max) to manganese in Mn-deficient soil. Indian J. Agric. Sci. 1994, 64. Available online: https://epubs.icar.org.in/index.php/IJAgS/article/view/29447 (accessed on 22 August 2024).
- Mahler, R.L.; Li, G.C.; Wattenbarger, D.W. Manganese relationships in spring wheat and spring Barley production in northern Idaho. Commun. Soil Sci. Plant Anal. 1992, 23, 1671–1692. [Google Scholar] [CrossRef]
- Aref, F. Influence of zinc and boron nutrition on copper, manganese, and iron concentration in the maize leaf. Aust. J. Basic Appl. Sci. 2011, 5, 52–62. [Google Scholar]
- Spiers, J.M. Nitrogen, calcium, and magnesium fertilization affects growth and leaf elemental content of ‘dormanred’ raspberry. J. Plant Nutr. 1993, 16, 2333–2339. [Google Scholar] [CrossRef]
- Hewitt, E.J. Relation of manganese and some other metals to the iron status of plants. Nature 1948, 161, 489–490. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Smith, F.A.; Smith, S.E. Phosphorus efficiencies and their effects on Zn, Cu, and Mn nutrition of different Barley (Hordeum vulgare) cultivars grown in sand culture. Aust. J. Agric. Res. 2002, 53, 211. [Google Scholar] [CrossRef]
- Ponnamperuma, F. The chemistry of submerged soils. Adv. Agron. 1972, 29–96. [Google Scholar] [CrossRef]
- Dhaliwal, S.S.; Walia, S.S. Integrated nutrient management for sustaining maximum productivity of rice-wheat system under Punjab conditions. J. Res. Punjab Agric. Univ. 2008, 45, 12–16. [Google Scholar]
- Walia, S.S.; Kler, D.S. Effect of organic and inorganic sources of nutrition on growth, macro and micro nutrient uptake in maize under maize-wheat sequence. Indian J. Ecol. 2010, 37, 27–29. [Google Scholar]
- Hansen, S.L.; Trakooljul, N.; Liu, H.-C.; Moeser, A.J.; Spears, J.W. Iron transporters are differentially regulated by dietary iron, and modifications are associated with changes in manganese metabolism in young pigs. J. Nutr. 2009, 139, 1474–1479. [Google Scholar] [CrossRef]
- Lönnerdal, B.; Keen, C.; Hurley, L. Manganese binding proteins in human and cow’s milk. Am. J. Clin. Nutr. 1985, 41, 550–559. [Google Scholar] [CrossRef]
- Davidson, L.A.; Lonnerdal, B. FE-saturation and proteolysis of human lactoferrin: Effect on brush-border receptor-mediated uptake of FE and Mn. Am. J. Physiol.-Gastrointest. Liver Physiol. 1989, 257, G930–G934. [Google Scholar] [CrossRef]
- Soares, A.T.; Silva, A.D.; Tinkov, A.A.; Khan, H.; Santamaría, A.; Skalnaya, M.G.; Skalny, A.V.; Tsatsakis, A.; Bowman, A.B.; Aschner, M.; et al. The impact of manganese on neurotransmitter systems. J. Trace Elem. Med. Biol. 2020, 61, 126554. [Google Scholar] [CrossRef]
- Davidson, L.; Cederblad, A.; Lönnerdal, B.; Sandström, B. The effect of individual dietary components on manganese absorption in humans. Am. J. Clin. Nutr. 1991, 54, 1065–1070. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.D.; Zech, L.; Greger, J.L. Manganese metabolism in rats: An improved methodology for assessing gut endogenous losses. Exp. Biol. Med. 1993, 202, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Raghib, M.H.; Wai-Yee, C.; Rennert, M.O. Comparative biological availability of manganese from extrinsically labelled milk diets using sucking rats as a model. Br. J. Nutr. 1986, 55, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Keen, C.L.; Bell, J.G.; Lonnerdal, B. The effect of age on manganese uptake and retention from milk and infant formulas in rats. J. Nutr. 1986, 116, 395–402. [Google Scholar] [CrossRef]
- Guan, H.; Wang, M.; Li, X.; Piao, F.; Li, Q.; Xu, L.; Kitamura, F.; Yokoyama, K. Manganese concentrations in maternal and umbilical cord blood: Related to birth size and environmental factors. Eur. J. Public Health 2013, 24, 150–157. [Google Scholar] [CrossRef]
- Zota, A.R.; Ettinger, A.S.; Bouchard, M.; Amarasiriwardena, C.J.; Schwartz, J.; Hu, H.; Wright, R.O. Maternal blood manganese levels and infant birth weight. Epidemiology 2009, 20, 367–373. [Google Scholar] [CrossRef]
- Yu, E.; Zhao, R.; Cai, Y.; Huang, J.; Li, C.; Li, C.; Mei, L.; Bao, L.; Chen, J.; Zhu, S. Determination of manganese content in cottonseed meal using near-infrared spectrometry and multivariate calibration. J. Cotton Res. 2019, 2, 12. [Google Scholar] [CrossRef]
- Chen, L.; Ding, G.; Gao, Y.; Wang, P.; Shi, R.; Huang, H.; Tian, Y. Manganese concentrations in maternal–infant blood and birth weight. Environ. Sci. Pollut. Res. 2014, 21, 6170–6175. [Google Scholar] [CrossRef]
- Couper, J. On the effects of black oxide of manganese when inhaled into the lungs. Br. Ann. Med. Pharm. Vital Stat. Gen. Sci. 1837, 1, 41–42. [Google Scholar]
- Huang, C.C.; Chu, N.S.; Lu, C.S.; Wang, J.D.; Tsai, J.L.; Tzeng, J.L.; Wolters, E.C.; Calne, D.B. Chronic manganese intoxication. Arch. Neurol. 1989, 46, 1104–1106. [Google Scholar] [CrossRef]
- Dorman, D.C.; Struve, M.F.; James, R.A.; McManus, B.E.; Marshall, M.W.; Wong, B.A. Influence of dietary manganese on the pharmacokinetics of inhaled manganese sulfate in male CD rats. Toxicol. Sci. 2001, 60, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Russell, R.; Beard, J.L.; Cousins, R.J.; Dunn, J.T.; Ferland, G.; Hambidge, K.M.; Lynch, S.; Penland, J.G.; Ross, A.C.; Stoecker, B.J.; et al. Dietary Reference Intakes for Vitamin A, Vitamin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium, and Zinc; Food and Nutrition Board, Institute of Medicine. National Academy Press: Washington, DC, USA, 2001. [Google Scholar] [CrossRef]
- US EPA. Health Effects Support Document for Manganese; US Environmental Protection Agency, Office of Water: Washington, DC, USA, 2002.
- Gordon, N.P.; Schaffer, D.M. Use of dietary supplements by female seniors in a large Northern California health plan. BMC Geriatr. 2005, 5, 4. [Google Scholar] [CrossRef] [PubMed]
- Freeland-Graves, J.H.; Bales, C.W.; Behmardi, F. Manganese requirements of humans. In Nutritional Bioavailability of Manganese; Kies, C., Ed.; American Chemical Society: Washington, DC, USA, 1987; pp. 90–104. [Google Scholar]
- Greger, J.L. Nutrition versus toxicology of manganese in humans: Evaluation of potential biomarkers. Neurotoxicology 1999, 20, 205–212. [Google Scholar] [PubMed]
- Chen, P.; Bornhorst, J.; Aschner, M. Manganese metabolism in humans. Front. Biosci. (Landmark Ed). 2018, 23, 1655–1679. [Google Scholar] [CrossRef] [PubMed]
- Aschner, J.L.; Aschner, M. Nutritional aspects of manganese homeostasis. Mol. Asp. Med. 2005, 26, 353–362. [Google Scholar] [CrossRef]
- Finley, J.; Johnson, P.; Johnson, L. Sex affects manganese absorption and retention by humans from a diet adequate in manganese. Am. J. Clin. Nutr. 1994, 60, 949–955. [Google Scholar] [CrossRef]
- O’Neal, S.L.; Zheng, W. Manganese toxicity upon overexposure: A decade in review. Curr. Environ. Health Rep. 2015, 2, 315–328. [Google Scholar] [CrossRef]
- Greger, J. Dietary standards for manganese: Overlap between nutritional and toxicological studies. J. Nutr. 1998, 128, 368S–371S. [Google Scholar] [CrossRef]
- NIH. Manganese. National Institutes of Health, Office of Dietary Supplements. 2015. Available online: https://ods.od.nih.gov/factsheets/Manganese-Consumer/ (accessed on 12 September 2024).
- Khan, K.; Wasserman, G.A.; Liu, X.; Ahmed, E.; Parvez, F.; Slavkovich, V.; Levy, D.; Mey, J.; van Geen, A.; Graziano, J.H. Manganese exposure from drinking water and children’s academic achievement. Neurotoxicology 2012, 33, 91–97. [Google Scholar] [CrossRef]
- DeSimone, L.A.; Hamilton, P.A.; Gilliom, R.J. Quality of Water from Domestic Wells in Principal Aquifers of the United States, 1991–2004; US Department of the Interior, US Geological Survey: Reston, VA, USA, 2009.
- Khan, K.; Factor-Litvak, P.; Wasserman, G.A.; Liu, X.; Ahmed, E.; Parvez, F.; Slavkovich, V.; Levy, D.; Mey, J.; van Geen, A. Manganese exposure from drinking water and children’s classroom behavior in Bangladesh. Environ. Health Perspect. 2011, 119, 1501. [Google Scholar] [CrossRef]
- Collipp, P.; Chen, S.; Maitinsky, S. Manganese in infant formulas and learning disability. Ann. Nutr. Metab. 1983, 27, 488–494. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.T.; Chowanadisai, W.; Crinella, F.M.; Chicz-DeMet, A.; Lönnerdal, B. Effect of high dietary manganese intake of neonatal rats on tissue mineral accumulation, striatal dopamine levels, and neurodevelopmental status. Neurotoxicology 2002, 23, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Leblondel, G.; Allain, P. Manganese transport by caco-2 cells. Biol. Trace Elem. Res. 1999, 67, 13–28. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Aranda, J.A.; Wapnir, R.A.; Lifshitz, F. In vivo intestinal absorption of manganese in the rat. J. Nutr. 1983, 113, 2601–2607. [Google Scholar] [CrossRef] [PubMed]
- Mena, I.; Horiuchi, K.; Burke, K.; Cotzias, G.C. Chronic manganese poisoning: Individual susceptibility and absorption of iron. Neurology 1969, 19, 1000. [Google Scholar] [CrossRef]
- Garcia, S.J.; Gellein, K.; Syversen, T.; Aschner, M. Iron deficient and manganese supplemented diets alter metals and transporters in the developing rat brain. Toxicol. Sci. 2007, 95, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Lönnerdal, B. Lactoferrin receptors in intestinal brush border membranes. In Lactoferrin Structure and Function; Springer: Boston, MA, USA, 1994; pp. 171–175. [Google Scholar] [CrossRef]
- Keen, C.; Ensunsa, J.; Lönnerdal, B.; Zidenberg-Cherr, S. Manganese. In Encyclopedia of Human Nutrition; Elsevier: Amsterdam, The Netherlands, 2005; pp. 217–225. [Google Scholar] [CrossRef]
- Dorman, D.C.; Foster, M.L. Olfactory transport of manganese: Implications for neurotoxicity. In Manganese in Health and Disease; The Royal Society of Chemistry: Cambridge, UK, 2014; pp. 119–132. [Google Scholar] [CrossRef]
- Ávila, D.S.; Rocha, J.B.; Tizabi, Y.; Dos Santos, A.P.; Santamaría, A.; Bowman, A.B.; Aschner, M. Manganese neurotoxicity. In Handbook of Neurotoxicity; Springer International Publishing: Cham, Switzerland, 2021; pp. 1–26. [Google Scholar] [CrossRef]
- Bowler, R.M.; Gocheva, V.; Harris, M.; Ngo, L.; Abdelouahab, N.; Wilkinson, J.; Doty, R.L.; Park, R.; Roles, H.A. Prospective study on neurotoxic effects in manganese-exposed bridge construction welders. Neurotoxicology 2011, 32, 596–605. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Chakraborty, S.; Mukhopadhyay, S.; Lee, E.; Paoliello, M.M.; Bowman, A.B.; Aschner, M. Manganese homeostasis in the nervous system. J. Neurochem. 2015, 134, 601–610. [Google Scholar] [CrossRef] [PubMed]
- de Bie, R.M.; Gladstone, R.M.; Strafella, A.P.; Ko, J.-H.; Lang, A.E. Manganese-induced Parkinsonism associated with methcathinone (Ephedrone) abuse. Arch. Neurol. 2007, 64, 886–889. [Google Scholar] [CrossRef]
- Sikk, K.; Taba, P.; Haldre, S.; Bergquist, J.; Nyholm, D.; Askmark, H.; Danfors, T.; Sörensen, J.; Thurfjell, L.; Raininko, R. Clinical, neuroimaging and neurophysiological features in addicts with manganese-ephedrone exposure. Acta Neurol. Scand. 2010, 121, 237–243. [Google Scholar] [CrossRef]
- Sikk, K.; Haldre, S.; Aquilonius, S.M.; Asser, A.; Paris, M.; Roose, Ä.; Petterson, J.; Eriksson, S.L.; Bergquist, J.; Taba, P. Manganese-induced parkinsonism in methcathinone abusers: Biomarkers of exposure and follow-up. Eur. J. Neurol. 2013, 20, 915–920. [Google Scholar] [CrossRef] [PubMed]
- Sikk, K.; Haldre, S.; Aquilonius, S.; Taba, P. Manganese-induced parkinsonism due to Ephedrone abuse. Park. Dis. 2011, 2011, 865319. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.-L.; Lu, L.; Pan, Y.-J.; Ding, C.-G.; Xu, D.-Y.; Huang, C.-F.; Pan, X.-F.; Zheng, W. Baseline blood levels of manganese, lead, cadmium, copper, and zinc in residents of Beijing suburb. Environ. Res. 2015, 140, 10–17. [Google Scholar] [CrossRef]
- Leggett, R.W. A biokinetic model for manganese. Sci. Total Environ. 2011, 409, 4179–4186. [Google Scholar] [CrossRef] [PubMed]
- Crossgrove, J.S.; Yokel, R.A. Manganese distribution across the blood–brain barrier III. The divalent metal transporter-1 is not the major mechanism mediating brain manganese uptake. Neurotoxicology 2004, 25, 451–460. [Google Scholar] [CrossRef]
- Jiang, Y.; Zheng, W.; Long, L.; Zhao, W.; Li, X.; Mo, X.; Lu, J.; Fu, X.; Li, W.; Liu, S. Brain magnetic resonance imaging and manganese concentrations in red blood cells of smelting workers: Search for biomarkers of manganese exposure. Neurotoxicology 2007, 28, 126–135. [Google Scholar] [CrossRef]
- Reaney, S.H.; Kwik-Uribe, C.L.; Smith, D.R. Manganese oxidation state and its implications for toxicity. Chem. Res. Toxicol. 2002, 15, 1119–1126. [Google Scholar] [CrossRef]
- Harris, W.R.; Chen, Y. Electron paramagnetic resonance and difference ultraviolet studies of Mn2+ binding to serum transferrin. J. Inorg. Biochem. 1994, 54, 1–19. [Google Scholar] [CrossRef]
- Nischwitz, V.; Berthele, A.; Michalke, B. Speciation analysis of selected metals and determination of their total contents in paired serum and cerebrospinal fluid samples: An approach to investigate the permeability of the human blood-cerebrospinal fluid-barrier. Anal. Chim. Acta 2008, 627, 258–269. [Google Scholar] [CrossRef]
- Jursa, T.; Smith, D.R. Ceruloplasmin alters the tissue disposition and neurotoxicity of manganese, but not its loading onto transferrin. Toxicol. Sci. 2009, 107, 182–193. [Google Scholar] [CrossRef]
- Gunter, T.E.; Gerstner, B.; Gunter, K.K.; Malecki, J.; Gelein, R.; Valentine, W.M.; Aschner, M.; Yule, D.I. Manganese transport via the transferrin mechanism. Neurotoxicology 2013, 34, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Nam, H.; Wang, C.-Y.; Zhang, L.; Zhang, W.; Hojyo, S.; Fukada, T.; Knutson, M.D. ZIP14 and DMT1 in the liver, pancreas, and heart are differentially regulated by iron deficiency and overload: Implications for tissue iron uptake in iron-related disorders. Haematologica 2013, 98, 1049–1057. [Google Scholar] [CrossRef] [PubMed]
- Aisen, P.; Leibman, A.; Zweier, J. Stoichiometric and site characteristics of the binding of iron to human transferrin. J. Biol. Chem. 1978, 253, 1930–1937. [Google Scholar] [CrossRef] [PubMed]
- Moos, T.; Morgan, E. Transferrin and transferrin receptor function in brain barrier systems. Cell. Mol. Neurobiol. 2000, 20, 77–95. [Google Scholar] [CrossRef] [PubMed]
- Jenkitkasemwong, S.; Wang, C.-Y.; Mackenzie, B.; Knutson, M. Physiologic implications of metal-ion transport by ZIP14 and ZIP8. BioMetals 2012, 25, 643–655. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Zhuang, L.; Maddox, D.M.; Smith, S.B.; Ganapathy, V. Structure, function, and expression pattern of a novel sodium-coupled citrate transporter (NaCT) cloned from mammalian brain. J. Biol. Chem. 2002, 277, 39469–39476. [Google Scholar] [CrossRef]
- Quadri, M.; Federico, A.; Zhao, T.; Breedveld, G.J.; Battisti, C.; Delnooz, C.; Severijnen, L.-A.; Di Toro Mammarella, L.; Mignarri, A.; Monti, L.; et al. Mutations in SLC30A10 cause parkinsonism and dystonia with hypermanganesemia, polycythemia, and chronic liver disease. Am. J. Hum. Genet. 2012, 90, 467–477. [Google Scholar] [CrossRef]
- Donovan, A.; Lima, C.A.; Pinkus, J.L.; Pinkus, G.S.; Zon, L.I.; Robine, S.; Andrews, N.C. The iron exporter ferroportin/Slc40a1 is essential for iron homeostasis. Cell Metab. 2005, 1, 191–200. [Google Scholar] [CrossRef]
- Sharon, L.; Feng, M.; Muend, S.; Braiterman, L.T.; Hubbard, A.L.; Rao, R. Vesicular Distribution of Secretory Pathway Ca2+-ATPase Isoform 1 and a Role in Manganese Detoxification in Liver-Derived Polarized Cells. Biometals 2011, 24, 159–170. [Google Scholar]
- Schroeder, H.A.; Balassa, J.J.; Tipton, I.H. Essential trace metals in man: Manganese. J. Chronic Dis. 1966, 19, 545–571. [Google Scholar] [CrossRef]
- Rahil-Khazen, R.; Bolann, B.J.; Myking, A.; Ulvik, R.J. Multi-element analysis of trace element levels in human autopsy tissues by using inductively coupled atomic emission spectrometry technique (ICP-AES). J. Trace Elem. Med. Biol. 2002, 16, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Pejović-Milić, A.; Chettle, D.R.; Oudyk, J.; Pysklywec, M.W.; Haines, T. Bone manganese as a biomarker of manganese exposure: A feasibility study. Am. J. Ind. Med. 2009, 52, 742–750. [Google Scholar] [CrossRef] [PubMed]
- O’Neal, S.L.; Hong, L.; Fu, S.; Jiang, W.; Jones, A.; Nie, L.H.; Zheng, W. Manganese accumulation in bone following chronic exposure in rats: Steady-state concentration and half-life in bone. Toxicol. Lett. 2014, 229, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Kodama, H.; Shimojo, N.; Suzuki, K.T. Distribution of manganese in rat pancreas and identification of its primary binding protein as pro-carboxypeptidase B. Biochem. J. 1991, 278, 857–862. [Google Scholar] [CrossRef]
- Rorsman, P.; Berggren, P.-O.; Hellman, B. Manganese accumulation in pancreatic β-cells and its stimulation by glucose. Biochem. J. 1982, 202, 435–444. [Google Scholar] [CrossRef]
- Koh, E.S.; Kim, S.J.; Yoon, H.E.; Chung, J.H.; Chung, S.; Park, C.W.; Chang, Y.S.; Shin, S.J. Association of blood manganese level with diabetes and renal dysfunction: A cross-sectional study of the Korean general population. BMC Endocr. Disord. 2014, 14, 24. [Google Scholar] [CrossRef]
- Ponnapakkam, T.; Iszard, M.; Henry-Sam, G. Effects of oral administration of manganese on the kidneys and urinary bladder of Sprague-Dawley rats. Int. J. Toxicol. 2003, 22, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Henriksson, J.; Tjalve, H. Manganese taken up into the CNS via the olfactory pathway in rats affects astrocytes. Toxicol. Sci. 2000, 55, 392–398. [Google Scholar] [CrossRef]
- Zogzas, C.E.; Aschner, M.; Mukhopadhyay, S. Structural elements in the transmembrane and cytoplasmic domains of the metal transporter SLC30A10 are required for its manganese efflux activity. J. Biol. Chem. 2016, 291, 15940–15957. [Google Scholar] [CrossRef]
- Ng, E.; Lind, P.M.; Lindgren, C.; Ingelsson, E.; Mahajan, A.; Morris, A.; Lind, L. Genome-wide association study of toxic metals and trace elements reveals novel associations. Hum. Mol. Genet. 2015, 24, 4739–4745. [Google Scholar] [CrossRef]
- Fujishiro, H.; Yano, Y.; Takada, Y.; Tanihara, M.; Himeno, S. Roles of ZIP8, ZIP14, and DMT1 in transport of cadmium and manganese in mouse kidney proximal tubule cells. Metallomics 2012, 4, 700–708. [Google Scholar] [CrossRef] [PubMed]
- Leyva-Illades, D.; Chen, P.; Zogzas, C.E.; Hutchens, S.; Mercado, J.M.; Swaim, C.D.; Morrisett, R.A.; Bowman, A.B.; Aschner, M.; Mukhopadhyay, S. SLC30A10 is a cell surface-localized manganese efflux transporter, and parkinsonism-causing mutations block its intracellular trafficking and efflux activity. J. Neurosci. 2014, 34, 14079–14095. [Google Scholar] [CrossRef] [PubMed]
- Avila, D.S.; Puntel, R.L.; Aschner, M. Manganese in health and disease. Met. Ions Life Sci. 2013, 13, 199–227. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Food | Range of Mean Concentrations (mg kg−1) |
|---|---|
| Nuts and nut products | 18.2–46.8 |
| Grains and grain products | 0.42–40.7 |
| Legumes | 2.24–6.73 |
| Vegetables and vegetable products | 0.42–6.64 |
| Fruits | 0.20–10.4 |
| Infant foods | 0.17–4.83 |
| Fruit juices and drinks | 0.05–11.5 |
| Desserts | 0.04–7.98 |
| Meat, poultry, fish and eggs | 0.10–3.99 |
| Mixed dishes | 0.69–2.98 |
| Condiments, fats and sweeteners | 0.04–1.45 |
| Beverages (including tea) | 0.00–2.09 |
| Soups | 0.19–0.65 |
| Milk and milk products | 0.02–0.49 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Obeng, S.K.; Kulhánek, M.; Balík, J.; Černý, J.; Sedlář, O. Manganese: From Soil to Human Health—A Comprehensive Overview of Its Biological and Environmental Significance. Nutrients 2024, 16, 3455. https://doi.org/10.3390/nu16203455
Obeng SK, Kulhánek M, Balík J, Černý J, Sedlář O. Manganese: From Soil to Human Health—A Comprehensive Overview of Its Biological and Environmental Significance. Nutrients. 2024; 16(20):3455. https://doi.org/10.3390/nu16203455
Chicago/Turabian StyleObeng, Sarfo Kwaku, Martin Kulhánek, Jiří Balík, Jindřich Černý, and Ondřej Sedlář. 2024. "Manganese: From Soil to Human Health—A Comprehensive Overview of Its Biological and Environmental Significance" Nutrients 16, no. 20: 3455. https://doi.org/10.3390/nu16203455
APA StyleObeng, S. K., Kulhánek, M., Balík, J., Černý, J., & Sedlář, O. (2024). Manganese: From Soil to Human Health—A Comprehensive Overview of Its Biological and Environmental Significance. Nutrients, 16(20), 3455. https://doi.org/10.3390/nu16203455