
Impact of Peptide Transport and Memory Function in the Brain


Abstract
1. Introduction
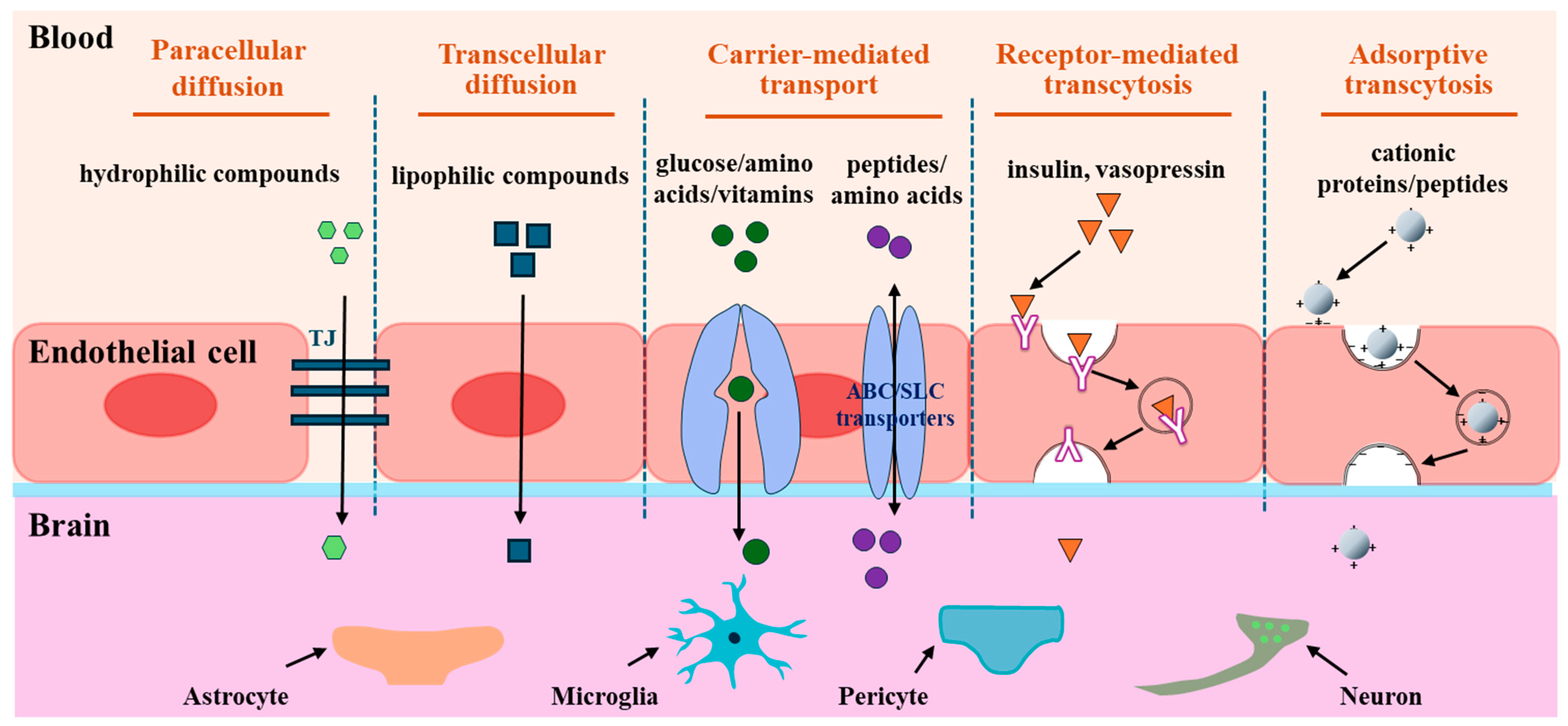
2. Transport of Peptides across the BBB to the Brain
2.1. Passive Diffusion
2.2. Carrier-Mediated Transport
2.3. Receptor-Mediated Transcytosis
2.4. Adsorptive Transcytosis
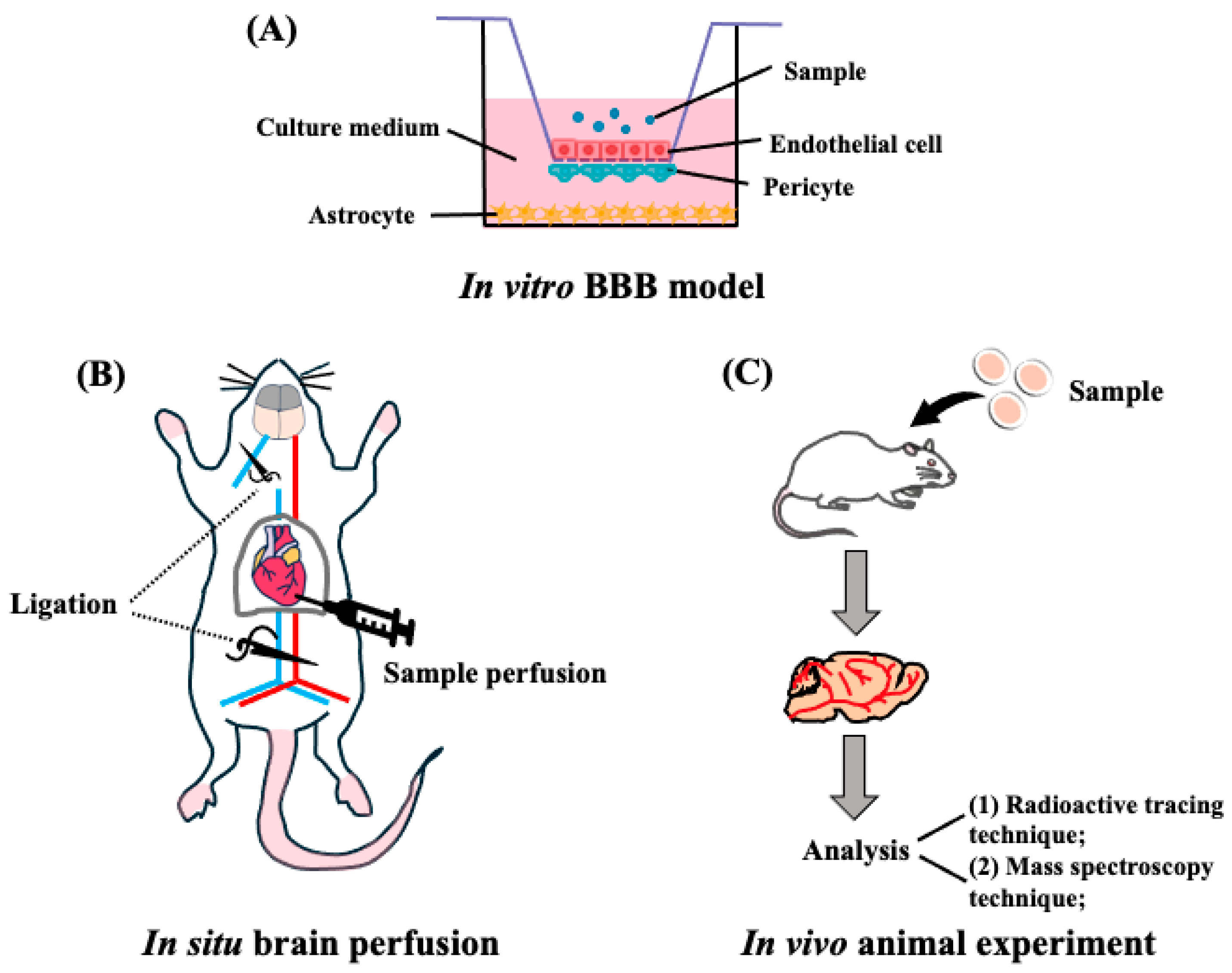
3. Evaluation of Peptide Transportability into the Brain
3.1. In Vitro BBB Reconstituted Models for Peptide Transport
3.2. In Vivo BBB Transport Models for Peptide Transport
3.3. In Vivo Imaging Techniques for Peptide BBB Transport
4. The Effects of Peptides on Alzheimer’s Disease
4.1. The Pathogenesis of the Alzheimer’s Disease
4.2. Alzheimer’s Disease Prevention by Food Peptides
5. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Kesidou, E.; Theotokis, P.; Damianidou, O.; Boziki, M.; Konstantinidou, N.; Taloumtzis, C.; Sintila, S.-A.; Grigoriadis, P.; Evangelopoulos, M.E.; Bakirtzis, C.; et al. CNS Ageing in Health and Neurodegenerative Disorders. J. Clin. Med. 2023, 12, 2255. [Google Scholar] [CrossRef]
- Steinmetz, J.D.; Seeher, K.M.; Schiess, N.; Nichols, E.; Cao, B.; Servili, C.; Cavallera, V.; Cousin, E.; Hagins, H.; Moberg, M.E.; et al. Global, Regional, and National Burden of Disorders Affecting the Nervous System, 1990–2021: A Systematic Analysis for the Global Burden of Disease Study 2021. Lancet Neurol. 2024, 23, 344–381. [Google Scholar] [CrossRef]
- Bhandari, D.; Rafiq, S.; Gat, Y.; Gat, P.; Waghmare, R.; Kumar, V. A Review on Bioactive Peptides: Physiological Functions, Bioavailability and Safety. Int. J. Pept. Res. Ther. 2020, 26, 139–150. [Google Scholar] [CrossRef]
- Jia, L.; Wang, L.; Liu, C.; Liang, Y.; Lin, Q. Bioactive Peptides from Foods: Production, Function, and Application. Food Funct. 2021, 12, 7108–7125. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Sun-Waterhouse, D.; Neil Waterhouse, G.I.; Zheng, L.; Su, G.; Zhao, M. Effects of Food-Derived Bioactive Peptides on Cognitive Deficits and Memory Decline in Neurodegenerative Diseases: A Review. Trends Food Sci. Technol. 2021, 116, 712–732. [Google Scholar] [CrossRef]
- Singh, K.; Gupta, J.K.; Kumar, S.; Soni, U. A Review of the Common Neurodegenerative Disorders: Current Therapeutic Approaches and the Potential Role of Bioactive Peptides. Curr. Protein Pept. Sci. 2024, 25, 507–526. [Google Scholar] [CrossRef] [PubMed]
- Kita, M.; Obara, K.; Kondo, S.; Umeda, S.; Ano, Y. Effect of Supplementation of a Whey Peptide Rich in Tryptophan-Tyrosine-Related Peptides on Cognitive Performance in Healthy Adults: A Randomized, Double-Blind, Placebo-Controlled Study. Nutrients 2018, 10, 899. [Google Scholar] [CrossRef] [PubMed]
- Kita, M.; Kobayashi, K.; Obara, K.; Koikeda, T.; Umeda, S.; Ano, Y. Supplementation with Whey Peptide Rich in β-Lactolin Improves Cognitive Performance in Healthy Older Adults: A Randomized, Double-Blind, Placebo-Controlled Study. Front. Neurosci. 2019, 13, 399. [Google Scholar] [CrossRef]
- Markus, C.R.; Olivier, B.; Panhuysen, G.E.M.; Van der Gugten, J.; Alles, M.S.; Tuiten, A.; Westenberg, H.G.M.; Fekkes, D.; Koppeschaar, H.F.; de Haan, E.E. The Bovine Protein α-Lactalbumin Increases the Plasma Ratio of Tryptophan to the Other Large Neutral Amino Acids, and in Vulnerable Subjects Raises Brain Serotonin Activity, Reduces Cortisol Concentration, and Improves Mood under Stress. Am. J. Clin. Nutr. 2000, 71, 1536–1544. [Google Scholar] [CrossRef]
- Loveday, S.M. Protein Digestion and Absorption: The Influence of Food Processing. Nutr. Res. Rev. 2023, 36, 544–559. [Google Scholar] [CrossRef]
- Kadry, H.; Noorani, B.; Cucullo, L. A Blood-Brain Barrier Overview on Structure, Function, Impairment, and Biomarkers of Integrity. Fluids Barriers CNS 2020, 17, 69. [Google Scholar] [CrossRef]
- Ronaldson, P.T.; Davis, T.P. Regulation of Blood-Brain Barrier Integrity by Microglia in Health and Disease: A Therapeutic Opportunity. J. Cereb. Blood Flow Metab. 2020, 40, S6–S24. [Google Scholar] [CrossRef] [PubMed]
- Pardridge, W.M. Why Is the Global CNS Pharmaceutical Market so Under-Penetrated? Drug Discov. Today 2002, 7, 5–7. [Google Scholar] [CrossRef]
- Pardridge, W.M. The Blood-Brain Barrier: Bottleneck in Brain Drug Development. Neurotherapeutics 2005, 2, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Banks, W.A. Characteristics of Compounds That Cross the Blood-Brain Barrier. BMC Neurol. 2009, 9 (Suppl. S1), S3. [Google Scholar] [CrossRef]
- Kastin, A.J.; Nissen, C.; Nikolics, K.; Medzihradszky, K.; Coy, D.H.; Teplan, I.; Schally, A.V. Distribution of 3H-α-MSH in Rat Brain. Brain Res. Bull. 1976, 1, 19–26. [Google Scholar] [CrossRef]
- Matsui, T.; Yoshino, A.; Tanaka, M. A Trip of Peptides to the Brain. Food Prod. Process. Nutr. 2020, 2, 30. [Google Scholar] [CrossRef]
- Abbott, N.J.; Rönnbäck, L.; Hansson, E. Astrocyte-Endothelial Interactions at the Blood-Brain Barrier. Nat. Rev. Neurosci. 2006, 7, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Wolburg, H.; Lippoldt, A. Tight Junctions of the Blood-Brain Barrier: Development, Composition and Regulation. Vascul. Pharmacol. 2002, 38, 323–337. [Google Scholar] [CrossRef]
- Curley, S.M.; Cady, N.C. Biologically-Derived Nanomaterials for Targeted Therapeutic Delivery to the Brain. Sci. Prog. 2018, 101, 273–292. [Google Scholar] [CrossRef]
- Teixidó, M.; Zurita, E.; Malakoutikhah, M.; Tarragó, T.; Giralt, E. Diketopiperazines as a Tool for the Study of Transport across the Blood−Brain Barrier (BBB) and Their Potential Use as BBB-Shuttles. J. Am. Chem. Soc. 2007, 129, 11802–11813. [Google Scholar] [CrossRef]
- Malakoutikhah, M.; Guixer, B.; Arranz-Gibert, P.; Teixidó, M.; Giralt, E. ‘À La Carte’ Peptide Shuttles: Tools to Increase Their Passage across the Blood–Brain Barrier. ChemMedChem 2014, 9, 1594–1601. [Google Scholar] [CrossRef] [PubMed]
- Arranz-Gibert, P.; Guixer, B.; Malakoutikhah, M.; Muttenthaler, M.; Guzmán, F.; Teixidó, M.; Giralt, E. Lipid Bilayer Crossing—The Gate of Symmetry. Water-Soluble Phenylproline-Based Blood-Brain Barrier Shuttles. J. Am. Chem. Soc. 2015, 137, 7357–7364. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Gonzalez, B.; Sanchez-Alegria, K.; Velazquez-Moctezuma, J. From Blood-to-Brain: Regulating the Permeability of the Blood-Brain Barrier. Curr. Psychopharmacol. 2012, 1, 214–227. [Google Scholar] [CrossRef]
- Campos-Bedolla, P.; Walter, F.R.; Veszelka, S.; Deli, M.A. Role of the Blood–Brain Barrier in the Nutrition of the Central Nervous System. Arch. Med. Res. 2014, 45, 610–638. [Google Scholar] [CrossRef]
- Morris, M.E.; Rodriguez-Cruz, V.; Felmlee, M.A. SLC and ABC Transporters: Expression, Localization, and Species Differences at the Blood-Brain and the Blood-Cerebrospinal Fluid Barriers. AAPS J. 2017, 19, 1317–1331. [Google Scholar] [CrossRef] [PubMed]
- Stieger, B.; Gao, B. Drug Transporters in the Central Nervous System. Clin. Pharmacokinet. 2015, 54, 225–242. [Google Scholar] [CrossRef]
- Shawahna, R.; Uchida, Y.; Declèves, X.; Ohtsuki, S.; Yousif, S.; Dauchy, S.; Jacob, A.; Chassoux, F.; Daumas-Duport, C.; Couraud, P.-O.; et al. Transcriptomic and Quantitative Proteomic Analysis of Transporters and Drug Metabolizing Enzymes in Freshly Isolated Human Brain Microvessels. Mol. Pharm. 2011, 8, 1332–1341. [Google Scholar] [CrossRef]
- Ohtsuki, S.; Terasaki, T. Contribution of Carrier-Mediated Transport Systems to the Blood–Brain Barrier as a Supporting and Protecting Interface for the Brain; Importance for CNS Drug Discovery and Development. Pharm. Res. 2007, 24, 1745–1758. [Google Scholar] [CrossRef]
- Puris, E.; Gynther, M.; Auriola, S.; Huttunen, K.M. L-Type Amino Acid Transporter 1 as a Target for Drug Delivery. Pharm. Res. 2020, 37, 88. [Google Scholar] [CrossRef] [PubMed]
- Benz, F.; Liebner, S. Structure and Function of the Blood-Brain Barrier (BBB). Handb. Exp. Pharmacol. 2022, 273, 3–31. [Google Scholar] [PubMed]
- Chafai, A.; Fromm, M.F.; König, J.; Maas, R. The Prognostic Biomarker L-Homoarginine Is a Substrate of the Cationic Amino Acid Transporters CAT1, CAT2A and CAT2B. Sci. Rep. 2017, 7, 4767. [Google Scholar] [CrossRef]
- Smith, D.E.; Clémençon, B.; Hediger, M.A. Proton-Coupled Oligopeptide Transporter Family SLC15: Physiological, Pharmacological and Pathological Implications. Mol. Aspects Med. 2013, 34, 323–336. [Google Scholar] [CrossRef] [PubMed]
- Bauer, K. Carnosine and Homocarnosine, the Forgotten, Enigmatic Peptides of the Brain. Neurochem. Res. 2005, 30, 1339–1345. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Xie, Y.; Keep, R.F.; Smith, D.E. Divergent Developmental Expression and Function of the Proton-Coupled Oligopeptide Transporters PepT2 and PhT1 in Regional Brain Slices of Mouse and Rat. J. Neurochem. 2014, 129, 955–965. [Google Scholar] [CrossRef]
- Hu, C.; Tao, L.; Cao, X.; Chen, L. The Solute Carrier Transporters and the Brain: Physiological and Pharmacological Implications. Asian J. Pharm. Sci. 2020, 15, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.R.; Shusta, E.V. Blood-Brain Barrier Transport of Therapeutics via Receptor-Mediation. Pharm. Res. 2007, 24, 1759–1771. [Google Scholar] [CrossRef]
- Haqqani, A.S.; Stanimirovic, D.B. Brain Delivery of Therapeutics via Transcytosis: Types and Mechanisms of Vesicle-Mediated Transport across the BBB. In Drug Delivery to the Brain; AAPS Advances in the Pharmaceutical Sciences Series; Springer International Publishing: Cham, Switzerland, 2022; pp. 71–91. ISBN 9783030887728. [Google Scholar]
- Bickel, U.; Yoshikawa, T.; Pardridge, W.M. Delivery of Peptides and Proteins through the Blood-Brain Barrier. Adv. Drug Deliv. Rev. 2001, 46, 247–279. [Google Scholar] [CrossRef]
- Guixer, B.; Arroyo, X.; Belda, I.; Sabidó, E.; Teixidó, M.; Giralt, E. Chemically Synthesized Peptide Libraries as a New Source of BBB Shuttles. Use of Mass Spectrometry for Peptide Identification. J. Pept. Sci. 2016, 22, 577–591. [Google Scholar] [CrossRef]
- Sakamoto, K.; Shinohara, T.; Adachi, Y.; Asami, T.; Ohtaki, T. A Novel LRP1-Binding Peptide L57 That Crosses the Blood Brain Barrier. Biochem. Biophys. Rep. 2017, 12, 135–139. [Google Scholar] [CrossRef]
- Hervé, F.; Ghinea, N.; Scherrmann, J.-M. CNS Delivery via Adsorptive Transcytosis. AAPS J. 2008, 10, 455–472. [Google Scholar] [CrossRef] [PubMed]
- Pardridge, W.M.; Boado, R.J. Enhanced Cellular Uptake of Biotinylated Antisense Oligonucleotide or Peptide Mediated by Avidin, a Cationic Protein. FEBS Lett. 1991, 288, 30–32. [Google Scholar] [CrossRef]
- Pardridge, W.M.; Triguero, D.; Buciak, J. Transport of Histone through the Blood-Brain Barrier. J. Pharmacol. Exp. Ther. 1989, 251, 821–826. [Google Scholar] [PubMed]
- Pardridge, W.M.; Buciak, J.L.; Kang, Y.S.; Boado, R.J. Protamine-Mediated Transport of Albumin into Brain and Other Organs of the Rat. Binding and Endocytosis of Protamine-Albumin Complex by Microvascular Endothelium. J. Clin. Investig. 1993, 92, 2224–2229. [Google Scholar] [CrossRef] [PubMed]
- Villegas, J.C.; Broadwell, R.D. Transcytosis of Protein through the Mammalian Cerebral Epithelium and Endothelium. II. Adsorptive Transcytosis of WGA-HRP and the Blood-Brain and Brain-Blood Barriers. J. Neurocytol. 1993, 22, 67–80. [Google Scholar] [CrossRef]
- Azarmi, M.; Maleki, H.; Nikkam, N.; Malekinejad, H. Transcellular Brain Drug Delivery: A Review on Recent Advancements. Int. J. Pharm. 2020, 586, 119582. [Google Scholar] [CrossRef]
- Sharma, G.; Modgil, A.; Zhong, T.; Sun, C.; Singh, J. Influence of Short-Chain Cell-Penetrating Peptides on Transport of Doxorubicin Encapsulating Receptor-Targeted Liposomes across Brain Endothelial Barrier. Pharm. Res. 2014, 31, 1194–1209. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, X.; Gong, M.; Zhang, J. Delivery of a Peptide-Drug Conjugate Targeting the Blood Brain Barrier Improved the Efficacy of Paclitaxel against Glioma. Oncotarget 2016, 7, 79401–79407. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, W.; Ma, L.; Fan, L.; Gao, F.; Ni, J.; Wang, R. The Improved Blood-Brain Barrier Permeability of Endomorphin-1 Using the Cell-Penetrating Peptide SynB3 with Three Different Linkages. Int. J. Pharm. 2014, 476, 1–8. [Google Scholar] [CrossRef]
- Temsamani, J.; Bonnafous, C.; Rousselle, C.; Fraisse, Y.; Clair, P.; Granier, L.-A.; Rees, A.R.; Kaczorek, M.; Scherrmann, J.-M. Improved Brain Uptake and Pharmacological Activity Profile of Morphine-6-Glucuronide Using a Peptide Vector-Mediated Strategy. J. Pharmacol. Exp. Ther. 2005, 313, 712–719. [Google Scholar] [CrossRef]
- Ruczyński, J.; Rusiecka, I.; Turecka, K.; Kozłowska, A.; Alenowicz, M.; Gągało, I.; Kawiak, A.; Rekowski, P.; Waleron, K.; Kocić, I. Transportan 10 Improves the Pharmacokinetics and Pharmacodynamics of Vancomycin. Sci. Rep. 2019, 9, 3247. [Google Scholar] [CrossRef] [PubMed]
- Hanafy, A.S.; Dietrich, D.; Fricker, G.; Lamprecht, A. Blood-Brain Barrier Models: Rationale for Selection. Adv. Drug Deliv. Rev. 2021, 176, 113859. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, S.; Deli, M.A.; Kawaguchi, H.; Shimizudani, T.; Shimono, T.; Kittel, A.; Tanaka, K.; Niwa, M. A New Blood-Brain Barrier Model Using Primary Rat Brain Endothelial Cells, Pericytes and Astrocytes. Neurochem. Int. 2009, 54, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Qi, Y.; Liu, X.; Fang, L.; Gao, Y.; Liu, C.; Wu, D.; Wang, X.; Zhao, F.; Wang, J.; et al. Neuroprotective Effects of Fermented Yak Milk-Derived Peptide LYLKPR on H2O2-Injured HT-22 Cells. Food Funct. 2022, 13, 12021–12038. [Google Scholar] [CrossRef]
- Oller-Salvia, B.; Teixidó, M.; Giralt, E. From Venoms to BBB Shuttles: Synthesis and Blood-Brain Barrier Transport Assessment of Apamin and a Nontoxic Analog. Biopolymers 2013, 100, 675–686. [Google Scholar] [CrossRef]
- Tsuruoka, N.; Beppu, Y.; Koda, H.; Doe, N.; Watanabe, H.; Abe, K. A DKP Cyclo(L-Phe-L-Phe) Found in Chicken Essence Is a Dual Inhibitor of the Serotonin Transporter and Acetylcholinesterase. PLoS ONE 2012, 7, e50824. [Google Scholar] [CrossRef]
- Hayes, M.; Moen, L.F.; Auty, M.A.E.; Lea, T.E. Transport of a Prolyl Endopeptidase Inhibitory Peptide across the Blood-Brain Barrier Demonstrated Using the HCMEC/D3 Cell Line Transcytosis Assay. J. Agric. Food Chem. 2016, 64, 146–150. [Google Scholar] [CrossRef]
- Christensen, B.; Toth, A.E.; Nielsen, S.S.E.; Scavenius, C.; Petersen, S.V.; Enghild, J.J.; Rasmussen, J.T.; Nielsen, M.S.; Sørensen, E.S. Transport of a Peptide from Bovine As1-Casein across Models of the Intestinal and Blood-Brain Barriers. Nutrients 2020, 12, 3157. [Google Scholar] [CrossRef]
- Domenger, D.; Cudennec, B.; Kouach, M.; Touche, V.; Landry, C.; Lesage, J.; Gosselet, F.; Lestavel, S.; Goossens, J.-F.; Dhulster, P.; et al. Food-Derived Hemorphins Cross Intestinal and Blood–Brain Barriers in Vitro. Front. Endocrinol. 2018, 9, 159. [Google Scholar] [CrossRef]
- Wu, D.; Chen, Q.; Chen, X.; Han, F.; Chen, Z.; Wang, Y. The Blood-Brain Barrier: Structure, Regulation, and Drug Delivery. Signal Transduct. Target. Ther. 2023, 8, 217. [Google Scholar] [CrossRef]
- Sun, Z.; Zhang, Z.; Wang, X.; Cade, R.; Elmir, Z.; Fregly, M. Relation of β-Casomorphin to Apnea in Sudden Infant Death Syndrome. Peptides 2003, 24, 937–943. [Google Scholar] [CrossRef] [PubMed]
- Ano, Y.; Ayabe, T.; Kutsukake, T.; Ohya, R.; Takaichi, Y.; Uchida, S.; Yamada, K.; Uchida, K.; Takashima, A.; Nakayama, H. Novel Lactopeptides in Fermented Dairy Products Improve Memory Function and Cognitive Decline. Neurobiol. Aging 2018, 72, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Ano, Y.; Kita, M.; Kitaoka, S.; Furuyashiki, T. Leucine-Histidine Dipeptide Attenuates Microglial Activation and Emotional Disturbances Induced by Brain Inflammation and Repeated Social Defeat Stress. Nutrients 2019, 11, 2161. [Google Scholar] [CrossRef] [PubMed]
- Ano, Y.; Yoshino, Y.; Kutsukake, T.; Ohya, R.; Fukuda, T.; Uchida, K.; Takashima, A.; Nakayama, H. Tryptophan-Related Dipeptides in Fermented Dairy Products Suppress Microglial Activation and Prevent Cognitive Decline. Aging 2019, 11, 2949–2967. [Google Scholar] [CrossRef]
- Min, L.-J.; Kobayashi, Y.; Mogi, M.; Tsukuda, K.; Yamada, A.; Yamauchi, K.; Abe, F.; Iwanami, J.; Xiao, J.-Z.; Horiuchi, M. Administration of Bovine Casein-Derived Peptide Prevents Cognitive Decline in Alzheimer Disease Model Mice. PLoS ONE 2017, 12, e0171515. [Google Scholar] [CrossRef]
- Newcomb, R.; Abbruscato, T.J.; Singh, T.; Nadasdi, L.; Davis, T.P.; Miljanich, G. Bioavailability of Ziconotide in Brain: Influx from Blood, Stability, and Diffusion. Peptides 2000, 21, 491–501. [Google Scholar] [CrossRef]
- Demeule, M.; Beaudet, N.; Régina, A.; Besserer-Offroy, É.; Murza, A.; Tétreault, P.; Belleville, K.; Ché, C.; Larocque, A.; Thiot, C.; et al. Conjugation of a Brain-Penetrant Peptide with Neurotensin Provides Antinociceptive Properties. J. Clin. Investig. 2014, 124, 1199–1213. [Google Scholar] [CrossRef]
- Tanaka, M.; Dohgu, S.; Komabayashi, G.; Kiyohara, H.; Takata, F.; Kataoka, Y.; Nirasawa, T.; Maebuchi, M.; Matsui, T. Brain-Transportable Dipeptides across the Blood-Brain Barrier in Mice. Sci. Rep. 2019, 9, 5769. [Google Scholar] [CrossRef]
- Cheng, L.; Tanaka, M.; Yoshino, A.; Nagasato, Y.; Takata, F.; Dohgu, S.; Matsui, T. A Memory-Improving Dipeptide, Tyr-Pro, Can Reach the Mouse Brain after Oral Administration. Sci. Rep. 2023, 13, 16908. [Google Scholar] [CrossRef]
- Nogimura, D.; Mizushige, T.; Taga, Y.; Nagai, A.; Shoji, S.; Azuma, N.; Kusubata, M.; Adachi, S.-I.; Yoshizawa, F.; Kabuyama, Y. Prolyl-Hydroxyproline, a Collagen-Derived Dipeptide, Enhances Hippocampal Cell Proliferation, Which Leads to Antidepressant-like Effects in Mice. FASEB J. 2020, 34, 5715–5723. [Google Scholar] [CrossRef]
- Tanaka, M.; Kiyohara, H.; Yoshino, A.; Nakano, A.; Takata, F.; Dohgu, S.; Kataoka, Y.; Matsui, T. Brain-Transportable Soy Dipeptide, Tyr-Pro, Attenuates Amyloid β Peptide25-35-Induced Memory Impairment in Mice. Npj Sci. Food 2020, 4, 7. [Google Scholar] [CrossRef]
- Harris, W.J.; Asselin, M.-C.; Hinz, R.; Parkes, L.M.; Allan, S.; Schiessl, I.; Boutin, H.; Dickie, B.R. In Vivo Methods for Imaging Blood-Brain Barrier Function and Dysfunction. Eur. J. Nucl. Med. Mol. Imaging 2023, 50, 1051–1083. [Google Scholar] [CrossRef]
- Raja, R.; Rosenberg, G.A.; Caprihan, A. MRI Measurements of Blood-Brain Barrier Function in Dementia: A Review of Recent Studies. Neuropharmacology 2018, 134, 259–271. [Google Scholar] [CrossRef] [PubMed]
- André, S.; Ansciaux, E.; Saidi, E.; Larbanoix, L.; Stanicki, D.; Nonclercq, D.; Vander Elst, L.; Laurent, S.; Muller, R.N.; Burtea, C. Validation by Magnetic Resonance Imaging of the Diagnostic Potential of a Heptapeptide-Functionalized Imaging Probe Targeted to Amyloid-β and Able to Cross the Blood-Brain Barrier. J. Alzheimers Dis. 2017, 60, 1547–1565. [Google Scholar] [CrossRef] [PubMed]
- Moyaert, P.; Padrela, B.E.; Morgan, C.A.; Petr, J.; Versijpt, J.; Barkhof, F.; Jurkiewicz, M.T.; Shao, X.; Oyeniran, O.; Manson, T.; et al. Imaging Blood-Brain Barrier Dysfunction: A State-of-the-Art Review from a Clinical Perspective. Front. Aging Neurosci. 2023, 15, 1132077. [Google Scholar] [CrossRef] [PubMed]
- Breuer, H.; Meier, M.; Schneefeld, S.; Härtig, W.; Wittneben, A.; Märkel, M.; Ross, T.L.; Bengel, F.M.; Bankstahl, M.; Bankstahl, J.P. Multimodality Imaging of Blood-Brain Barrier Impairment during Epileptogenesis. J. Cereb. Blood Flow Metab. 2017, 37, 2049–2061. [Google Scholar] [CrossRef] [PubMed]
- la Fougère, C.; Rominger, A.; Förster, S.; Geisler, J.; Bartenstein, P. PET and SPECT in Epilepsy: A Critical Review. Epilepsy Behav. 2009, 15, 50–55. [Google Scholar] [CrossRef]
- Bagni, B.; Feggi, L.M.; Carraro, P.L.; Candini, G. Determination of cerebral blood flow using non-diffusible tracers. Radiol. Med. 1983, 69, 788–791. [Google Scholar]
- Wilson, D.M., 3rd; Cookson, M.R.; Van Den Bosch, L.; Zetterberg, H.; Holtzman, D.M.; Dewachter, I. Hallmarks of Neurodegenerative Diseases. Cell 2023, 186, 693–714. [Google Scholar] [CrossRef]
- Tiwari, S.; Atluri, V.; Kaushik, A.; Yndart, A.; Nair, M. Alzheimer’s Disease: Pathogenesis, Diagnostics, and Therapeutics. Int. J. Nanomed. 2019, 14, 5541–5554. [Google Scholar] [CrossRef]
- Kandimalla, R.; Reddy, P.H. Therapeutics of Neurotransmitters in Alzheimer’s Disease. J. Alzheimers Dis. 2017, 57, 1049–1069. [Google Scholar] [CrossRef] [PubMed]
- Querfurth, H.W.; LaFerla, F.M. Alzheimer’s Disease. N. Engl. J. Med. 2010, 362, 329–344. [Google Scholar] [CrossRef]
- Hampel, H.; Hardy, J.; Blennow, K.; Chen, C.; Perry, G.; Kim, S.H.; Villemagne, V.L.; Aisen, P.; Vendruscolo, M.; Iwatsubo, T.; et al. The Amyloid-β Pathway in Alzheimer’s Disease. Mol. Psychiatry 2021, 26, 5481–5503. [Google Scholar] [CrossRef]
- Hu, W.; Wu, F.; Zhang, Y.; Gong, C.-X.; Iqbal, K.; Liu, F. Expression of Tau Pathology-Related Proteins in Different Brain Regions: A Molecular Basis of Tau Pathogenesis. Front. Aging Neurosci. 2017, 9, 311. [Google Scholar] [CrossRef]
- Zhang, H.; Wei, W.; Zhao, M.; Ma, L.; Jiang, X.; Pei, H.; Cao, Y.; Li, H. Interaction between Aβ and Tau in the Pathogenesis of Alzheimer’s Disease. Int. J. Biol. Sci. 2021, 17, 2181–2192. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.-K.; Kuan, Y.-C.; Lin, H.-W.; Hu, C.-J. Clinical Trials of New Drugs for Alzheimer Disease: A 2020–2023 Update. J. Biomed. Sci. 2023, 30, 83. [Google Scholar] [CrossRef]
- Chin, E.; Jaqua, E.; Safaeipour, M.; Ladue, T. Conventional versus New Treatment: Comparing the Effects of Acetylcholinesterase Inhibitors and N-Methyl-D-Aspartate Receptor Antagonist with Aducanumab. Cureus 2022, 14, e31065. [Google Scholar] [CrossRef] [PubMed]
- Praticò, D. Oxidative Stress Hypothesis in Alzheimer’s Disease: A Reappraisal. Trends Pharmacol. Sci. 2008, 29, 609–615. [Google Scholar] [CrossRef]
- Grune, T. Oxidized Protein Aggregates: Formation and Biological Effects. Free Radic. Biol. Med. 2020, 150, 120–124. [Google Scholar] [CrossRef]
- Kinney, J.W.; Bemiller, S.M.; Murtishaw, A.S.; Leisgang, A.M.; Salazar, A.M.; Lamb, B.T. Inflammation as a Central Mechanism in Alzheimer’s Disease. Alzheimer’s Dement. 2018, 4, 575–590. [Google Scholar] [CrossRef]
- Picca, A.; Calvani, R.; Coelho-Junior, H.J.; Landi, F.; Bernabei, R.; Marzetti, E. Mitochondrial Dysfunction, Oxidative Stress, and Neuroinflammation: Intertwined Roads to Neurodegeneration. Antioxidants 2020, 9, 647. [Google Scholar] [CrossRef]
- Li, W.; Zhao, T.; Zhang, J.; Wu, C.; Zhao, M.; Su, G. Comparison of Neuroprotective and Cognition-Enhancing Properties of Hydrolysates from Soybean, Walnut, and Peanut Protein. J. Chem. 2016, 2016, 1–8. [Google Scholar] [CrossRef]
- Ju, D.-T.; Kumar, K.A.; Kuo, W.-W.; Ho, T.-J.; Chang, R.-L.; Lin, W.-T.; Day, C.H.; Viswanadha, V.V.P.; Liao, P.-H.; Huang, C.-Y. Bioactive Peptide VHVV Upregulates the Long-Term Memory-Related Biomarkers in Adult Spontaneously Hypertensive Rats. Int. J. Mol. Sci. 2019, 20, 3069. [Google Scholar] [CrossRef] [PubMed]
- Amakye, W.K.; Hou, C.; Xie, L.; Lin, X.; Gou, N.; Yuan, E.; Ren, J. Bioactive Anti-Aging Agents and the Identification of New Anti-Oxidant Soybean Peptides. Food Biosci. 2021, 42, 101194. [Google Scholar] [CrossRef]
- Cheng, L.; Shi, C.; Nakamura, S.; Esaki, N.; Ichiba, Y.; Tanaka, M.; Sakai, K.; Matsui, T. Adiponectin-Receptor Agonistic Dipeptide Tyr-Pro Stimulates the Acetylcholine Nervous System in NE-4C Cells. J. Agric. Food Chem. 2024, 72, 7121–7129. [Google Scholar] [CrossRef]
- Wang, S.; Su, G.; Zhang, Q.; Zhao, T.; Liu, Y.; Zheng, L.; Zhao, M. Walnut (Juglans regia) Peptides Reverse Sleep Deprivation-Induced Memory Impairment in Rat via Alleviating Oxidative Stress. J. Agric. Food Chem. 2018, 66, 10617–10627. [Google Scholar] [CrossRef]
- Liu, C.; Guo, Y.; Zhao, F.; Qin, H.; Lu, H.; Fang, L.; Wang, J.; Min, W. Potential Mechanisms Mediating the Protective Effects of a Peptide from Walnut (Juglans mandshurica Maxim.) against Hydrogen Peroxide Induced Neurotoxicity in PC12 Cells. Food Funct. 2019, 10, 3491–3501. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Amakye, W.K.; Guo, L.; Gong, C.; Zhao, Y.; Yao, M.; Ren, J. Walnut-Derived Peptide PW5 Ameliorates Cognitive Impairments and Alters Gut Microbiota in APP/PS1 Transgenic Mice. Mol. Nutr. Food Res. 2019, 63, e1900326. [Google Scholar] [CrossRef]
- Wang, S.; Zheng, L.; Zhao, T.; Zhang, Q.; Liu, Y.; Sun, B.; Su, G.; Zhao, M. Inhibitory Effects of Walnut (Juglans regia) Peptides on Neuroinflammation and Oxidative Stress in Lipopolysaccharide-Induced Cognitive Impairment Mice. J. Agric. Food Chem. 2020, 68, 2381–2392. [Google Scholar] [CrossRef]
- Zhao, F.; Wang, J.; Lu, H.; Fang, L.; Qin, H.; Liu, C.; Min, W. Neuroprotection by Walnut-Derived Peptides through Autophagy Promotion via Akt/MTOR Signaling Pathway against Oxidative Stress in PC12 Cells. J. Agric. Food Chem. 2020, 68, 3638–3648. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Su, G.; Zhang, X.; Song, G.; Zhang, L.; Zheng, L.; Zhao, M. Characterization and Exploration of Potential Neuroprotective Peptides in Walnut (Juglans regia) Protein Hydrolysate against Cholinergic System Damage and Oxidative Stress in Scopolamine-Induced Cognitive and Memory Impairment Mice and Zebrafish. J. Agric. Food Chem. 2021, 69, 2773–2783. [Google Scholar] [CrossRef]
- Gao, Y.; Qin, H.; Wu, D.; Liu, C.; Fang, L.; Wang, J.; Liu, X.; Min, W. Walnut Peptide WEKPPVSH in Alleviating Oxidative Stress and Inflammation in Lipopolysaccharide-Activated BV-2 Microglia via the Nrf2/HO-1 and NF-ΚB/P38 MAPK Pathways. J. Biosci. Bioeng. 2021, 132, 496–504. [Google Scholar] [CrossRef]
- Wang, S.; Zheng, L.; Zhao, T.; Zhang, Q.; Su, G.; Zhao, M. The Neuroprotective Effect of Walnut-Derived Peptides against Glutamate-Induced Damage in PC12 Cells: Mechanism and Bioavailability. Food Sci. Hum. Wellness 2022, 11, 933–942. [Google Scholar] [CrossRef]
- Lu, H.; Fang, L.; Wang, J.; Zhao, F.; Liu, C.; Gao, Y.; Liu, J.; Min, W. Pine Nut Antioxidant Peptides Ameliorate the Memory Impairment in a Scopolamine-Induced Mouse Model via SIRT3-Induced Synaptic Plasticity. Food Funct. 2021, 12, 8026–8036. [Google Scholar] [CrossRef]
- Lee, J.K.; Li-Chan, E.C.Y.; Cheung, I.W.Y.; Jeon, Y.-J.; Ko, J.-Y.; Byun, H.-G. Neuroprotective Effect of β-Secretase Inhibitory Peptide from Pacific Hake (Merluccius productus) Fish Protein Hydrolysate. Curr. Alzheimer Res. 2019, 16, 1028–1038. [Google Scholar] [CrossRef] [PubMed]
- Chai, H.-J.; Wu, C.-J.; Yang, S.-H.; Li, T.-L.; Sun Pan, B. Peptides from Hydrolysate of Lantern Fish (Benthosema pterotum) Proved Neuroprotective in Vitro and in Vivo. J. Funct. Foods 2016, 24, 438–449. [Google Scholar] [CrossRef]
- Gao, R.; Shu, W.; Shen, Y.; Sun, Q.; Bai, F.; Wang, J.; Li, D.; Li, Y.; Jin, W.; Yuan, L. Sturgeon Protein-Derived Peptides Exert Anti-Inflammatory Effects in LPS-Stimulated RAW264.7 Macrophages via the MAPK Pathway. J. Funct. Foods 2020, 72, 104044. [Google Scholar] [CrossRef]
- Zhao, Y.; Dong, Y.; Ge, Q.; Cui, P.; Sun, N.; Lin, S. Neuroprotective Effects of NDEELNK from Sea Cucumber Ovum against Scopolamine-Induced PC12 Cell Damage through Enhancing Energy Metabolism and Upregulation of the PKA/BDNF/NGF Signaling Pathway. Food Funct. 2021, 12, 7676–7687. [Google Scholar] [CrossRef]
- Lu, M.; Mishra, A.; Boschetti, C.; Lin, J.; Liu, Y.; Huang, H.; Kaminski, C.F.; Huang, Z.; Tunnacliffe, A.; Kaminski Schierle, G.S. Sea Cucumber-Derived Peptides Alleviate Oxidative Stress in Neuroblastoma Cells and Improve Survival in C. elegans Exposed to Neurotoxic Paraquat. Oxid. Med. Cell. Longev. 2021, 2021, 8842926. [Google Scholar] [CrossRef]
- Wu, D.; Zhang, S.; Sun, N.; Zhu, B.; Lin, S. Neuroprotective Function of a Novel Hexapeptide QMDDQ from Shrimp via Activation of the PKA/CREB/BNDF Signaling Pathway and Its Structure-Activity Relationship. J. Agric. Food Chem. 2020, 68, 6759–6769. [Google Scholar] [CrossRef]
- Zhao, T.; Su, G.; Wang, S.; Zhang, Q.; Zhang, J.; Zheng, L.; Sun, B.; Zhao, M. Neuroprotective Effects of Acetylcholinesterase Inhibitory Peptides from Anchovy (Coilia mystus) against Glutamate-Induced Toxicity in PC12 Cells. J. Agric. Food Chem. 2017, 65, 11192–11201. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Ding, J.; Lu, Z.; Zhu, B.; Lin, S. Digestive and Absorptive Properties of the Antarctic Krill Tripeptide Phe-pro-Phe (FPF) and Its Auxiliary Memory-Enhancing Effect. J. Agric. Food Chem. 2024, 72, 8491–8505. [Google Scholar] [CrossRef] [PubMed]
- Ano, Y.; Kutsukake, T.; Sasaki, T.; Uchida, S.; Yamada, K.; Kondo, K. Identification of a Novel Peptide from β-Casein That Enhances Spatial and Object Recognition Memory in Mice. J. Agric. Food Chem. 2019, 67, 8160–8167. [Google Scholar] [CrossRef] [PubMed]
- Rafique, H.; Hu, X.; Ren, T.; Dong, R.; Aadil, R.M.; Zou, L.; Sharif, M.K.; Li, L. Characterization and Exploration of the Neuroprotective Potential of Oat-Protein-Derived Peptides in PC12 Cells and Scopolamine-Treated Zebrafish. Nutrients 2023, 16, 117. [Google Scholar] [CrossRef]
- Du, Z.; Li, Y. Review and Perspective on Bioactive Peptides: A Roadmap for Research, Development, and Future Opportunities. J. Agric. Food Res. 2022, 9, 100353. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Peptide | Source | Experiment Model | Transportability | Refs |
|---|---|---|---|---|
| fermented yak milk peptide | in vitro BBB model | 6.90 ± 0.73 × 10−7 cm/s | [55] | |
| (LYLKPR) | ||||
| apamin | venom | in vitro BBB model | 1.7 ± 0.1 × 10−6 cm/s | [56] |
| (CNCKAPETALCARRQQH) | ||||
| Cyclo (FF) | chicken essence | in vitro BBB model | ~25 × 10−6 cm/s | [57] |
| PPL | fish skin | in vitro BBB model | na | [58] |
| αS1-casein peptide | milk | in vitro BBB model | 1.09 ± 0.14 × 10−6 cm /s | [59] |
| (PIGSENSEKTTMPLW) | ||||
| H7 | hemoglobin | in vitro BBB model | <1 × 10−6 cm /s | [60] |
| (YPWTQRF) | ||||
| LVV-H4 | hemoglobin | in vitro BBB model | <1 × 10−6 cm /s | [60] |
| (LVVYPWT) | ||||
| VV-H4 | hemoglobin | in vitro BBB model | <1 × 10−6 cm /s | [60] |
| (VVYPWT) | ||||
| VV-H7 | hemoglobin | in vitro BBB model | <1 × 10−6 cm /s | [60] |
| (VVYPWTQRF) | ||||
| casomorphin-5 | milk | mouse | 0.266 nmol/g·min | [62] |
| (YPFPG) | ||||
| casomorphin-7 | milk | mouse | na | [62] |
| (YPFPGPI) | ||||
| GTWY | whey protein | rat oral administration (radioactivity) | tissue/plasma ratio: 0.32 (hippocampus) and 0.39 (cerebral cortex) | [63] |
| LH | - | rat oral administration (radioactivity) | tissue/plasma ratio: 0.2 | [64] |
| WY | fermented dairy products | rat oral administration (radioactivity) | tissue/plasma ratio: 0.23 (hippocampus) and 0.24 (cerebral cortex) | [65] |
| MKP | milk | rat oral administration | autoradiographic image | [66] |
| L57 | - | in situ mouse brain perfusion | radioactivity | [41] |
| (TWPKHFDKHTFYSILKLGKH-OH) | ||||
| ziconotide | sea snails | rat intravenous injection | 0.005%/g brain tissue | [67] |
| (CKGKGAKCSRLMYDCCTGSCRSGKC) | ||||
| Angiopep-2 | - | in vitro BBB model and in situ brain perfusion | radioactivity | [68] |
| (TFFYGGCRGKRNNFKTEEY-COOH) | ||||
| Gly-Sar | soybean | in situ mouse brain perfusion | 7.60 ± 1.29 μL/g·min | [69] |
| GP | soybean | in situ mouse brain perfusion | 3.49 ± 0.66 μL/g·min | [69] |
| YP | soybean | in situ mouse brain perfusion | 3.53 ± 0.74 μL/g·min | [69] |
| YP | soybean | mouse oral administration | AUC0–120 min: 0.34 ± 0.11 pmol·min/mg-dry brain, at 10 mg/kg | [70] |
| PO | collagen | rat oral administration | ~0.5 nmol/mL in cerebrospinal fluid | [71] |
| (Pro-Hyp) |
| Peptide | Source | Experiment Model | Action | Refs |
|---|---|---|---|---|
| VHVV | soybean | rat | activation of CREB-mediated downstream proteins | [94] |
| WPK | soybean | PC12 cell | attenuated H2O2 induced oxidative stress | [95] |
| AYLH | soybean | PC12 cell | [95] | |
| YP | soybean | mouse and NE-4C cell | stimulation of ChAT expression and ACh production | [96] |
| GGW | walnut | PC12 cell | protection against glutamate-induced apoptosis | [97] |
| VYY | walnut | PC12 cell | [97] | |
| LLPF | walnut | PC12 cell | [97] | |
| EVSGPGLSPN | walnut | PC12 cell | protection against H2O2-induced neurotoxicity | [98] |
| PPKNW | walnut | APP/PS1 mouse | inhibition of Aβ42 aggregation | [99] |
| LPF | walnut | mouse | decrease in TNF-α and IL-1β production | [100] |
| GVYY | walnut | mouse | [100] | |
| APTLW | walnut | mouse | [100] | |
| YVLLPSPK | walnut | PC12 cell | Akt/mTOR-mediated autophagy promotion against oxidative stress | [101] |
| TWLPLPR | walnut | PC12 cell | [101] | |
| KVPPLLY | walnut | PC12 cell | [101] | |
| FY | walnut | zebrafish | AChE and Keap1 inhibitors | [102] |
| SGFDAE | walnut | zebrafish | [102] | |
| WEKPPVSH | walnut | BV-2 microglia cell | decreased NO and ROS generation, mitigated secretion of IL-6, TNF-α and IL-1β | [103] |
| WSREEQERE | walnut | PC12 cell | neuroprotective effect against glutamate-induced apoptosis | [104] |
| ADIYTEEAGR | walnut | PC12 cell | [104] | |
| WYPGK | pine nuts | PC12 cell and mouse | SIRT3-induced synaptic plasticity enhancement | [105] |
| LN | pacific hake fish | SH-SY5Y cell | inhibition of Aβ production | [106] |
| FYY | lantern fish | SH-SY5Y cell and mouse | reduced H2O2 induced ROS and apoptotic cell death | [107] |
| DW | lantern fish | [107] | ||
| KIWHHTF | sturgeon | RAW264.7 | anti-inflammatory effect by inhibiting the MAPK pathway | [108] |
| VHYAGTVDY | sturgeon | [108] | ||
| HLDDALRGQE | sturgeon | [108] | ||
| NDEELNK | sea cucumber | PC12 cell | cholinergic system-involved alleviation of cell damage | [109] |
| FETLMPLWGNK | sea cucumber | human neuroblastoma cell and Caenorhabditis elegans | antioxidant activity at both cellular and organism levels | [110] |
| HEPFYGNEGALR | sea cucumber | [110] | ||
| KMYPVPLN | sea cucumber | [110] | ||
| QMDDQ | shrimp | PC12 cell | neuroprotective effect by increasing ACh content and inhibiting AChE activity | [111] |
| PAYCS | anchovy | PC12 cell | AChE inhibition, ROS and Ca2+ influx-mediated cell protection | [112] |
| CVGSY | anchovy | [112] | ||
| FPF | Antarctic krill | mouse | elevation of ACh content, AChE inhibition | [113] |
| KEMPFPKYPVEP | Camembert cheese | mouse | elevation of ACh content, AChE inhibition | [114] |
| LYLKPR | fermented yak milk | HT-22 cell | amelioration of neuronal injury by inhibiting the NLRP3 inflammasome | [55] |
| DFVADHPFLF | oat protein hydrolysate | PC12 cell and zebrafish | neuroprotective activity mediated by upregulation of BDNF, Nrf2, and Erg1 | [115] |
| HGQNFPIL | [115] | |||
| RDFPITWPW | [115] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, L.; Shi, C.; Li, X.; Matsui, T. Impact of Peptide Transport and Memory Function in the Brain. Nutrients 2024, 16, 2947. https://doi.org/10.3390/nu16172947
Cheng L, Shi C, Li X, Matsui T. Impact of Peptide Transport and Memory Function in the Brain. Nutrients. 2024; 16(17):2947. https://doi.org/10.3390/nu16172947
Chicago/Turabian StyleCheng, Lihong, Caiyue Shi, Xixi Li, and Toshiro Matsui. 2024. "Impact of Peptide Transport and Memory Function in the Brain" Nutrients 16, no. 17: 2947. https://doi.org/10.3390/nu16172947
APA StyleCheng, L., Shi, C., Li, X., & Matsui, T. (2024). Impact of Peptide Transport and Memory Function in the Brain. Nutrients, 16(17), 2947. https://doi.org/10.3390/nu16172947