
Amelioration of Insulin Resistance by Whey Protein in a High-Fat Diet-Induced Pediatric Obesity Male Mouse Model

 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
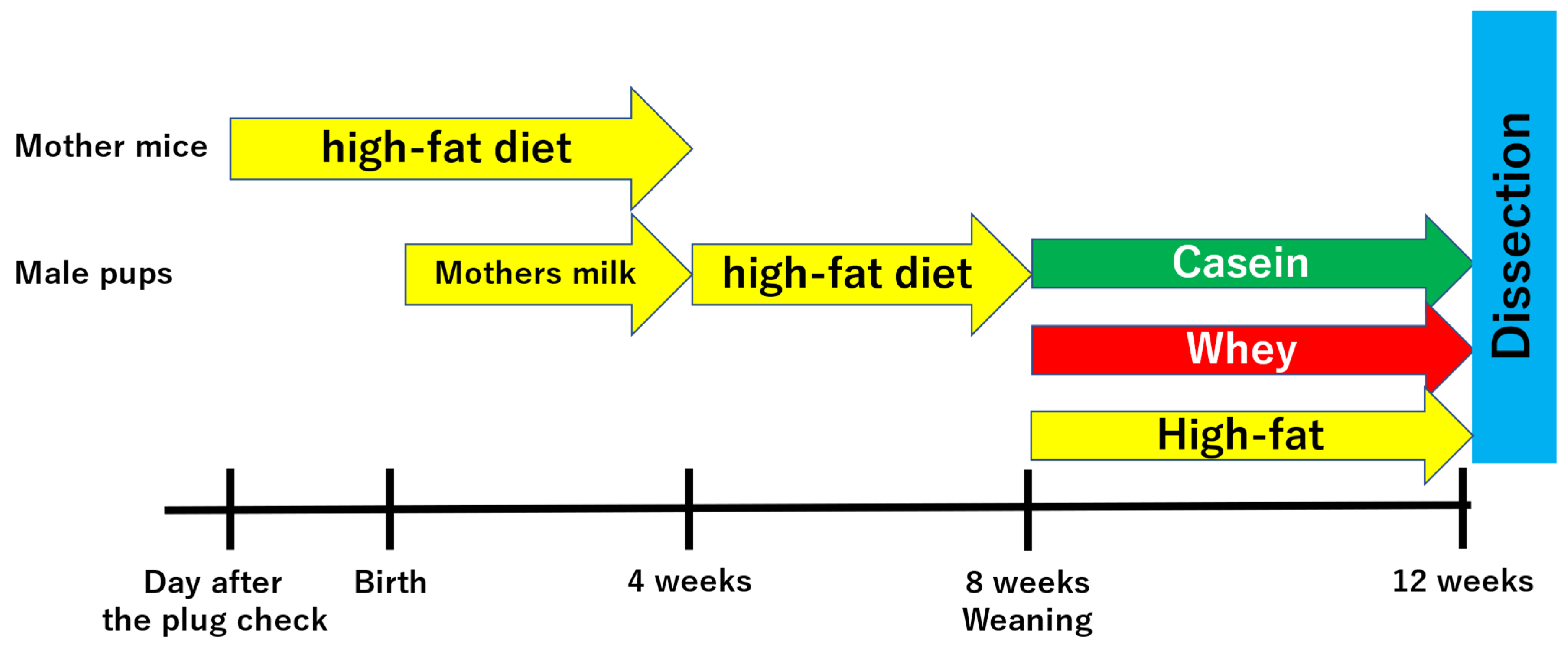
2.1. Experimental Animals
2.2. Rearing Conditions
2.3. Body Weight
2.4. Fasting Blood Glucose Level, Serum Insulin Level, and Insulin Resistance (HOMA-IR)
2.5. Body Composition and Fat Weight
2.6. Serum Lipoprotein Fractionation
2.7. Oxidative Stress Markers
2.8. Gene Expression Analysis of the Liver Tissue
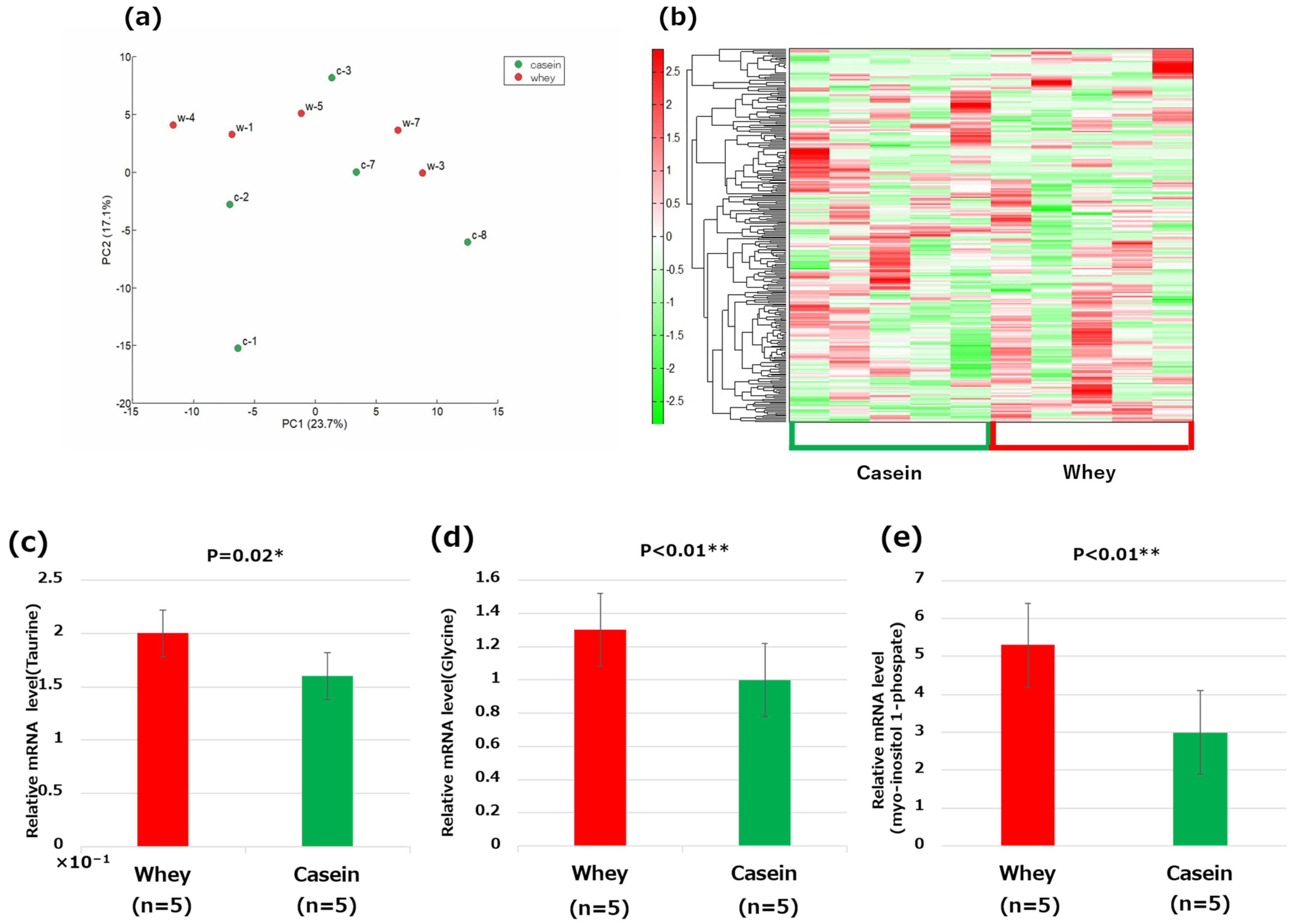
2.9. Metabolome Analysis of the Liver Tissue
2.10. Statistical Analysis
3. Results
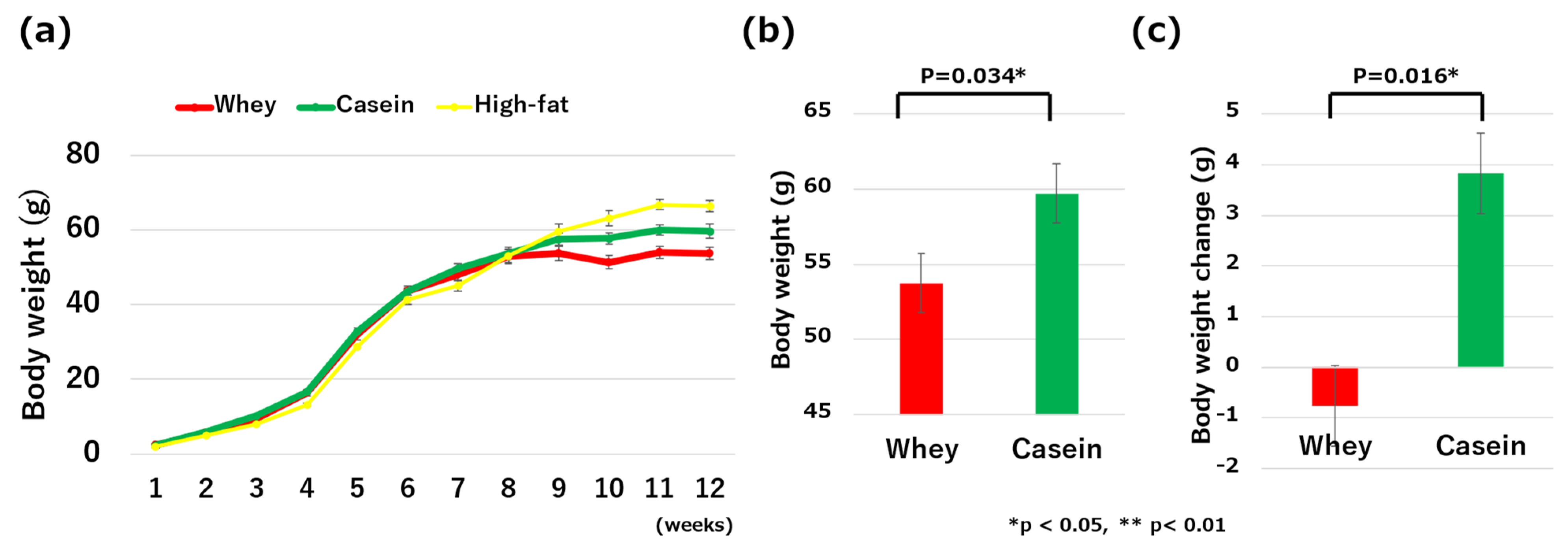
3.1. Weight Change
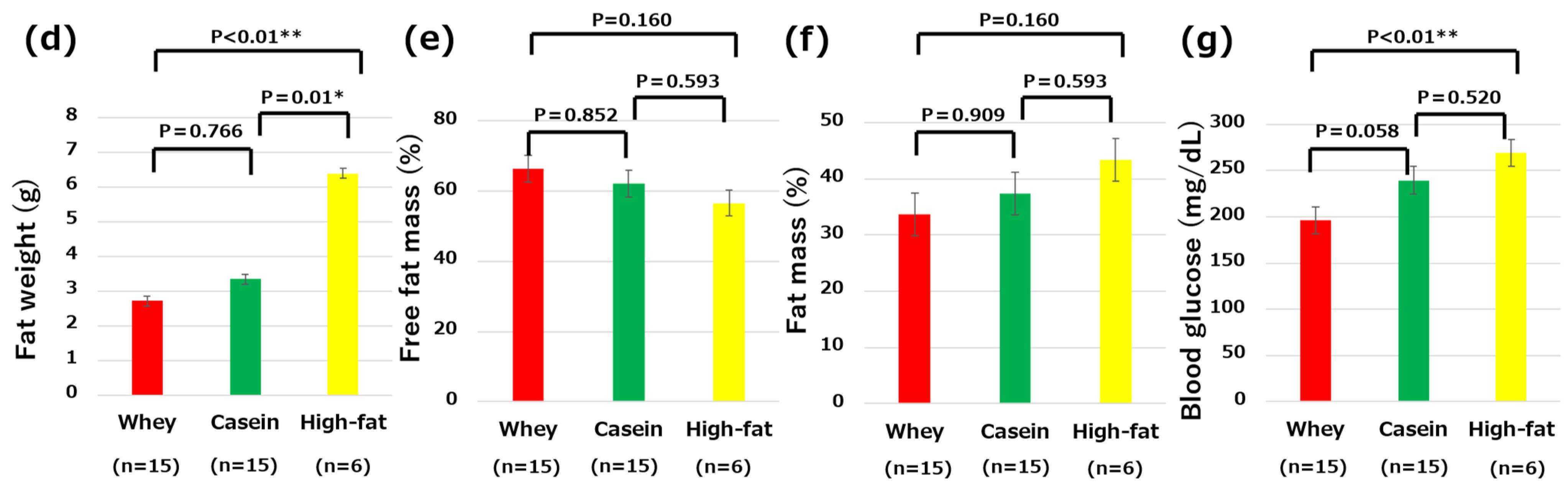
3.2. Fat Weight and Body Composition
3.3. Fasting Blood Glucose
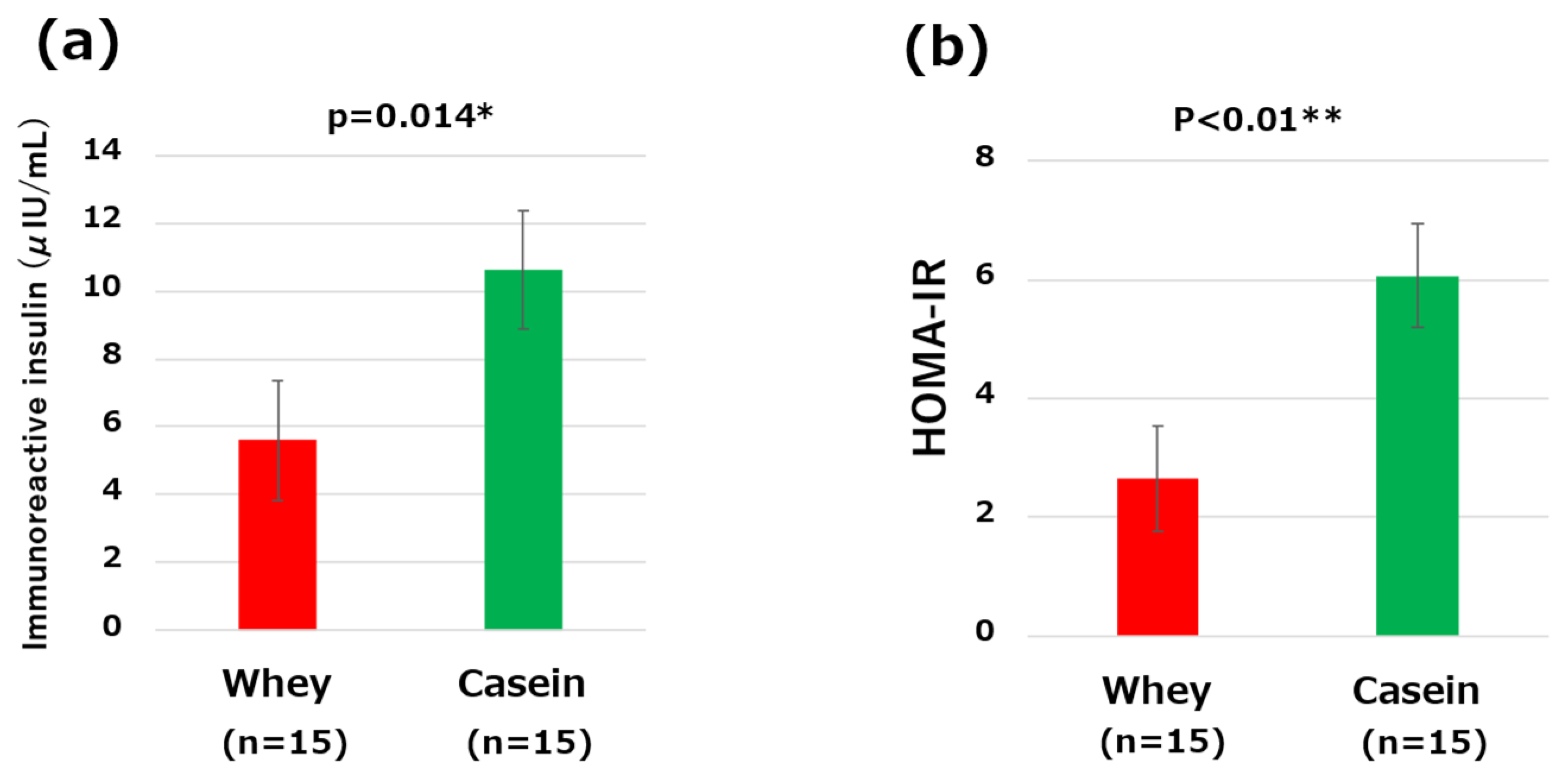
3.4. IRI, and HOMA-IR
3.5. Serum Lipoprotein Fractionation
3.6. Oxidative Stress, Biological Antioxidant Capacity, and Oxidative Stress Index
3.7. Analysis of Genes Related to Lipid Metabolism in the Liver
3.8. Metabolome Analysis of the Liver
4. Discussion
4.1. Lipid Metabolism-Related Genes
4.2. Involvement of Taurine
4.3. Glycine and Myo-Inositol 1-Phosphate
4.4. Limitations
4.5. Future Prospects
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barker, D.J. Obesity and early life. Obes. Rev. 2007, 8 (Suppl. 1), 45–49. [Google Scholar] [CrossRef]
- Gluckman, P.D.; Hanson, M.A. Living with the past: Evolution, development, and patterns of disease. Science 2004, 305, 1733–1736. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Jin, Z.; Yang, Y.; Jiang, F.; Huang, H.; Liu, S.; Jin, X. Association of low birth weight with thinness and severe obesity in children aged 3-12 years: A large-scale population-based cross-sectional study in Shanghai, China. BMJ Open 2019, 9, e028738. [Google Scholar] [CrossRef] [PubMed]
- Kubo, T. Common approach to childhood obesity in Japan. J. Pediatr. Endocrinol. Metab. 2014, 27, 581–592. [Google Scholar] [CrossRef]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef] [PubMed]
- Toffolon, A.; de Rocco-Ponce, M.; Vettore, M.; Iori, E.; Lante, A.; Tessari, P. Effect of Reversal of Whey-Protein to Casein Ratio of Cow Milk, on Insulin, Incretin, and Amino Acid Responses in Humans. Mol. Nutr. Food Res. 2021, 65, e2100069. [Google Scholar] [CrossRef] [PubMed]
- Pennings, B.; Boirie, Y.; Senden, J.M.; Gijsen, A.P.; Kuipers, H.; van Loon, L.J. Whey protein stimulates postprandial muscle protein accretion more effectively than do casein and casein hydrolysate in older men. Am. J. Clin. Nutr. 2011, 93, 997–1005. [Google Scholar] [CrossRef]
- Marshall, K. Therapeutic applications of whey protein. Altern. Med. Rev. 2004, 9, 136–156. [Google Scholar]
- Nonogaki, K.; Kaji, T. Whey protein isolate inhibits hepatic FGF21 production, which precedes weight gain, hyperinsulinemia and hyperglycemia in mice fed a high-fat diet. Sci. Rep. 2020, 10, 15784. [Google Scholar] [CrossRef]
- Tulipano, G.; Sibilia, V.; Caroli, A.M.; Cocchi, D. Whey proteins as a source of dipeptidyl dipeptidase IV (dipeptidyl peptidase-4) inhibitors. Peptides 2011, 32, 835–838. [Google Scholar] [CrossRef]
- Kawase, M.; Hashimoto, H.; Hosoda, M.; Morita, H.; Hosono, A. Effect of administration of fermented milk containing whey protein concentrate to rats on serum lipids and blood pressure. J. Dairy Sci. 2000, 83, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Prokopidis, K.; Mazidi, M.; Sankaranarayanan, R.; Tajik, B.; McArdle, A.; Isanejad, M. Effects of whey and soy protein supplementation on inflammatory cytokines in older adults: A systematic review and meta-analysis. Br. J. Nutr. 2023, 129, 759–770. [Google Scholar] [CrossRef] [PubMed]
- Veskoukis, A.S.; Kerasioti, E.; Skaperda, Z.; Papapostolou, P.A.; Nepka, C.; Spandidos, D.A.; Asprodini, E.; Taitzoglou, I.; Kouretas, D. Whey protein boosts the antioxidant profile of rats by enhancing the activities of crucial antioxidant enzymes in a tissue-specific manner. Food Chem. Toxicol. 2020, 142, 111508. [Google Scholar] [CrossRef]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2004, 114, 1752–1761. [Google Scholar] [CrossRef]
- Maddux, B.A.; See, W.; Lawrence, J.C., Jr.; Goldfine, A.L.; Goldfine, I.D.; Evans, J.L. Protection against oxidative stress-induced insulin resistance in rat L6 muscle cells by mircomolar concentrations of alpha-lipoic acid. Diabetes 2001, 50, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, T.; Kajimoto, Y.; Watada, H.; Kaneto, H.; Kishimoto, M.; Umayahara, Y.; Fujitani, Y.; Kamada, T.; Kawamori, R.; Yamasaki, Y. Glycation-dependent, reactive oxygen species-mediated suppression of the insulin gene promoter activity in HIT cells. J. Clin. Investig. 1997, 99, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Nomiyama, T.; Perez-Tilve, D.; Ogawa, D.; Gizard, F.; Zhao, Y.; Heywood, E.B.; Jones, K.L.; Kawamori, R.; Cassis, L.A.; Tschöp, M.H.; et al. Osteopontin mediates obesity-induced adipose tissue macrophage infiltration and insulin resistance in mice. J. Clin. Investig. 2007, 117, 2877–2888. [Google Scholar] [CrossRef] [PubMed]
- Nakazaki, K.; Nagano, N.; Katayama, D.; Shimizu, S.; Matsuda, K.; Tokunaga, W.; Aoki, R.; Fuwa, K.; Morioka, I. Body Fat-Reducing Effects of Whey. Nutrients 2023, 15, 2263. [Google Scholar] [CrossRef] [PubMed]
- Nemes, A.; Homoki, J.R.; Kiss, R.; Hegedűs, C.; Kovács, D.; Peitl, B.; Gál, F.; Stündl, L.; Szilvássy, Z.; Remenyik, J. Effect of Anthocyanin-Rich Tart Cherry Extract on Inflammatory Mediators and Adipokines Involved in Type 2 Diabetes in a High Fat Diet Induced Obesity Mouse Model. Nutrients 2019, 11, 1966. [Google Scholar] [CrossRef] [PubMed]
- Nagy, C.; Einwallner, E. Study of In Vivo Glucose Metabolism in High-fat Diet-fed Mice Using Oral Glucose Tolerance Test (OGTT) and Insulin Tolerance Test (ITT). J. Vis. Exp. 2018, 131, 56672. [Google Scholar] [CrossRef]
- Dorfman, M.D.; Krull, J.E.; Douglass, J.D.; Fasnacht, R.; Lara-Lince, F.; Meek, T.H.; Shi, X.; Damian, V.; Nguyen, H.T.; Matsen, M.E.; et al. Sex differences in microglial CX3CR1 signalling determine obesity susceptibility in mice. Nat. Commun. 2017, 8, 14556. [Google Scholar] [CrossRef] [PubMed]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Lukaski, H.C.; Johnson, P.E.; Bolonchuk, W.W.; Lykken, G.I. Assessment of fat-free mass using bioelectrical impedance measurements of the human. Am. J. Clin. Nutr. 1985, 41, 810–817. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, M.; Yamashita, S. Recent Advances in Analytical Methods on Lipoprotein Subclasses: Calculation of Particle Numbers from Lipid Levels by Gel Permeation HPLC Using “Spherical Particle Model”. J. Oleo Sci. 2016, 65, 265–282. [Google Scholar] [CrossRef] [PubMed]
- Usui, S.; Hara, Y.; Hosaki, S.; Okazaki, M. A new on-line dual enzymatic method for simultaneous quantification of cholesterol and triglycerides in lipoproteins by HPLC. J. Lipid Res. 2002, 43, 805–814. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Rodríguez, M.A.; Mendoza-Núñez, V.M. Oxidative Stress Indexes for Diagnosis of Health or Disease in Humans. Oxid. Med. Cell. Longev. 2019, 2019, 4128152. [Google Scholar] [CrossRef]
- Sasaki, H. Whey Protein Intake Modulates Lipid Metabolism by Transcriptionally Affecting PPARs and SREBP1c and Their Downstream Enzymes in Mice. Food Nutr. Sci. 2019, 10, 1045–1055. [Google Scholar] [CrossRef]
- Sugimoto, M.; Wong, D.T.; Hirayama, A.; Soga, T.; Tomita, M. Capillary electrophoresis mass spectrometry-based saliva metabolomics identified oral, breast and pancreatic cancer-specific profiles. Metabolomics 2010, 6, 78–95. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Fujimori, T.; Sato, H.; Ishikawa, G.; Kami, K.; Ohashi, Y. Statistical hypothesis testing of factor loading in principal component analysis and its application to metabolite set enrichment analysis. BMC Bioinform. 2014, 15, 51. [Google Scholar] [CrossRef]
- Berger, J.; Leibowitz, M.D.; Doebber, T.W.; Elbrecht, A.; Zhang, B.; Zhou, G.; Biswas, C.; Cullinan, C.A.; Hayes, N.S.; Li, Y.; et al. Novel peroxisome proliferator-activated receptor (PPAR) gamma and PPARdelta ligands produce distinct biological effects. J. Biol. Chem. 1999, 274, 6718–6725. [Google Scholar] [CrossRef]
- Dubois, V.; Eeckhoute, J.; Lefebvre, P.; Staels, B. Distinct but complementary contributions of PPAR isotypes to energy homeostasis. J. Clin. Investig. 2017, 127, 1202–1214. [Google Scholar] [CrossRef]
- McArdle, M.A.; Finucane, O.M.; Connaughton, R.M.; McMorrow, A.M.; Roche, H.M. Mechanisms of obesity-induced inflammation and insulin resistance. Front. Endocrinol. 2013, 4, 52. [Google Scholar] [CrossRef]
- Ota, T. Chemokine systems link obesity to insulin resistance. Diabetes Metab. J. 2013, 37, 165–172. [Google Scholar] [CrossRef]
- Althaher, A.R. An Overview of Hormone-Sensitive Lipase (HSL). ScientificWorldJournal 2022, 2022, 1964684. [Google Scholar] [CrossRef]
- Reid, B.N.; Ables, G.P.; Otlivanchik, O.A.; Schoiswohl, G.; Zechner, R.; Blaner, W.S.; Goldberg, I.J.; Schwabe, R.F.; Chua, S.C., Jr.; Huang, L.S. Hepatic overexpression of hormone-sensitive lipase and adipose triglyceride lipase promotes fatty acid oxidation, stimulates direct release of free fatty acids, and ameliorates steatosis. J. Biol. Chem. 2008, 283, 13087–13099. [Google Scholar] [CrossRef]
- Yamaguchi, K.; Hosokawa, Y. Cysteine dioxygenase. Methods Enzymol. 1987, 143, 395–403. [Google Scholar] [CrossRef]
- Erbersdobler, H.F.; Trautwein, E.; Greulich, H.G. Determinations of taurine in milk and infant formula diets. Eur. J. Pediatr. 1984, 142, 133–134. [Google Scholar] [CrossRef]
- Chitapanarux, T.; Tienboon, P.; Pojchamarnwiputh, S.; Leelarungrayub, D. Open-labeled pilot study of cysteine-rich whey protein isolate supplementation for nonalcoholic steatohepatitis patients. J. Gastroenterol. Hepatol. 2009, 24, 1045–1050. [Google Scholar] [CrossRef]
- Tsuboyama-Kasaoka, N.; Shozawa, C.; Sano, K.; Kamei, Y.; Kasaoka, S.; Hosokawa, Y.; Ezaki, O. Taurine (2-aminoethanesulfonic acid) deficiency. Endocrinology 2006, 147, 3276–3284. [Google Scholar] [CrossRef]
- Murakami, S.; Fujita, M.; Nakamura, M.; Sakono, M.; Nishizono, S.; Sato, M.; Imaizumi, K.; Mori, M.; Fukuda, N. Taurine ameliorates cholesterol metabolism by stimulating bile acid production in high-cholesterol-fed rats. Clin. Exp. Pharmacol. Physiol. 2016, 43, 372–378. [Google Scholar] [CrossRef]
- Fukuda, N.; Yoshitama, A.; Sugita, S.; Fujita, M.; Murakami, S. Dietary taurine reduces hepatic secretion of cholesteryl ester and enhances fatty acid oxidation in rats fed a high-cholesterol diet. J. Nutr. Sci. Vitaminol. 2011, 57, 144–149. [Google Scholar] [CrossRef]
- De Carvalho, F.G.; Brandao, C.F.C.; Batitucci, G.; Souza, A.O.; Ferrari, G.D.; Alberici, L.C.; Muñoz, V.R.; Pauli, J.R.; De Moura, L.P.; Ropelle, E.R.; et al. Taurine supplementation associated with exercise increases mitochondrial activity and fatty acid oxidation gene expression in subcutaneous white adipose tissue of obese women. Clin. Nutr. 2021, 40, 2180–2187. [Google Scholar] [CrossRef]
- Ruiz-Ramírez, A.; Ortiz-Balderas, E.; Cardozo-Saldaña, G.; Diaz-Diaz, E.; El-Hafidi, M. Glycine restores glutathione and protects against oxidative stress in vascular tissue from sucrose-fed rats. Clin. Sci. 2014, 126, 19–29. [Google Scholar] [CrossRef]
- El-Hafidi, M.; Franco, M.; Ramírez, A.R.; Sosa, J.S.; Flores, J.A.P.; Acosta, O.L.; Salgado, M.C.; Cardoso-Saldaña, G. Glycine Increases Insulin Sensitivity and Glutathione Biosynthesis and Protects against Oxidative Stress in a Model of Sucrose-Induced Insulin Resistance. Oxidative Med. Cell. Longev. 2018, 2018, 2101562. [Google Scholar] [CrossRef]
- Croze, M.L.; Soulage, C.O. Potential role and therapeutic interests of myo-inositol in metabolic diseases. Biochimie 2013, 95, 1811–1827. [Google Scholar] [CrossRef]
- Tu-Sekine, B.; Kim, S.F. The Inositol Phosphate System-A Coordinator of Metabolic Adaptability. Int. J. Mol. Sci. 2022, 23, 6747. [Google Scholar] [CrossRef]
- Tang, X.; Xu, G.; Pan, Y.-X.; Chen, H. Epigenetic Regulations of Genes Related to Lipid Metabolism by MicroRNA in Mice Fed High Fat Diet. FASEB J. 2018, 32, 648.20. [Google Scholar] [CrossRef]
- Kuwabara, R.; Urakami, T.; Yoshida, K.; Morioka, I. Case of type 2 diabetes possibly caused by excessive accumulation of visceral fat in a child born small-for-gestational age. J. Diabetes Investig. 2020, 11, 1366–1369. [Google Scholar] [CrossRef]
- Matsubasa, T.; Uchino, T.; Karashima, S.; Kondo, Y.; Maruyama, K.; Tanimura, M.; Endo, F. Oxidative stress in very low birth weight infants as measured by urinary 8-OHdG. Free Radic. Res. 2002, 36, 189–193. [Google Scholar] [CrossRef]
- Kunz, C.; Lönnerdal, B. Re-evaluation of the whey protein/casein ratio of human milk. Acta Paediatr. 1992, 81, 107–112. [Google Scholar] [CrossRef]
- Sluik, D.; Brouwer-Brolsma, E.M.; Berendsen, A.A.M.; Mikkilä, V.; Poppitt, S.D.; Silvestre, M.P.; Tremblay, A.; Pérusse, L.; Bouchard, C.; Raben, R.A.; et al. Protein intake and the incidence of pre-diabetes and diabetes in four population-based studies: The PREVIEW project. Am. J. Clin. Nutr. 2019, 109, 1310–1318. [Google Scholar] [CrossRef] [PubMed]
- Pietrobelli, A.; Agosti, M. Nutrition in the First 1000 Days: Ten Practices to Minimize Obesity Emerging from Published Science. Int. J. Environ. Res. Public Health 2017, 14, 1491. [Google Scholar] [CrossRef]
- Katayama, D.; Nagano, N.; Shimizu, S.; Nakazaki, K.; Matsuda, K.; Tokunaga, W.; Fuwa, K.; Aoki, R.; Morioka, I. A Non-Obese Hyperglycemic Mouse Model that Develops after Birth with Low Birthweight. Biomedicines 2022, 10, 1642. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Direction | Sequence | Start Position | Size |
|---|---|---|---|---|
| PPARα | Forward | CTCAGGGTACCACTACGGAGTTCAC | 1579 | 153 |
| Reverse | TGAATCTTGCAGCTCCGATCAC | 1731 | ||
| PPARγ | Forward | GGAGCCTAAGTTTGAGTTTGCTGTG | 1579 | 153 |
| Reverse | TGCAGCAGGTTGTCTTGGATG | 1731 | ||
| HSL | Forward | TCCTGGAACTAAGTGGACGCAAG | 2093 | 93 |
| Reverse | CAGACACACTCCTGCGCATAGAC | 2185 | ||
| LPL | Forward | AGAGGCTATAGCTGGGAGCAGAAAC | 3440 | 137 |
| Reverse | GCAAGGGCTAACATTCCAGCA | 3616 |
| Casein (n = 10) | Whey (n = 10) | p-Value | |
|---|---|---|---|
| Total-Cho | 159.29 (117.69–212.91) | 139.2 (91.83–182.78) | 0.12 |
| CM-Cho | 0.61 (0.31–0.85) | 0.69 (0.3–1.23) | 0.68 |
| VLDL-Cho | 11.94 (5.71–19.12) | 13.04 (6.03–19.56) | 0.52 |
| LDL-Cho | 26.18 (14.66–33.65) | 19.12 (8.96–27.14) | 0.16 |
| HDL-Cho | 120.55 (71.51–173.39) | 106.33 (70.46–127.63) | 0.21 |
| Total-TG | 95.58 (42.82–162.86) | 91.37 (42.76–146.48) | 0.79 |
| CM-TG | 4.99 (1.09–9.45) | 4.69 (1–11.66) | 0.85 |
| VLDL-TG | 69.21 (26.22–131.14) | 68.64 (25.09–115.98) | 0.91 |
| LDL-TG | 17.17 (11.72–22.78) | 14.29 (9.64–24.01) | 0.09 |
| HDL-TG | 4.21 (2.03–6.39) | 3.749 (2.36–4.49) | 0.24 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsuda, K.; Nagano, N.; Nakazaki, K.; Katayama, D.; Tokunaga, W.; Okuda, K.; Shimizu, S.; Aoki, R.; Fuwa, K.; Shirai, K.; et al. Amelioration of Insulin Resistance by Whey Protein in a High-Fat Diet-Induced Pediatric Obesity Male Mouse Model. Nutrients 2024, 16, 1622. https://doi.org/10.3390/nu16111622
Matsuda K, Nagano N, Nakazaki K, Katayama D, Tokunaga W, Okuda K, Shimizu S, Aoki R, Fuwa K, Shirai K, et al. Amelioration of Insulin Resistance by Whey Protein in a High-Fat Diet-Induced Pediatric Obesity Male Mouse Model. Nutrients. 2024; 16(11):1622. https://doi.org/10.3390/nu16111622
Chicago/Turabian StyleMatsuda, Kengo, Nobuhiko Nagano, Kimitaka Nakazaki, Daichi Katayama, Wataru Tokunaga, Koh Okuda, Shoichi Shimizu, Ryoji Aoki, Kazumasa Fuwa, Keisuke Shirai, and et al. 2024. "Amelioration of Insulin Resistance by Whey Protein in a High-Fat Diet-Induced Pediatric Obesity Male Mouse Model" Nutrients 16, no. 11: 1622. https://doi.org/10.3390/nu16111622
APA StyleMatsuda, K., Nagano, N., Nakazaki, K., Katayama, D., Tokunaga, W., Okuda, K., Shimizu, S., Aoki, R., Fuwa, K., Shirai, K., Fujioka, K., & Morioka, I. (2024). Amelioration of Insulin Resistance by Whey Protein in a High-Fat Diet-Induced Pediatric Obesity Male Mouse Model. Nutrients, 16(11), 1622. https://doi.org/10.3390/nu16111622