
The Potential Immunomodulatory Effect of Bifidobacterium longum subsp. longum BB536 on Healthy Adults through Plasmacytoid Dendritic Cell Activation in the Peripheral Blood

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. In Vitro Analysis
2.1.1. Preparation of Peripheral-Blood Mononuclear Cells (PBMCs)
2.1.2. Coculture of PBMCs and Heat-Killed B. longum BB536
2.1.3. Fluorescence Activated Cell Sorter (FACS) Analysis
2.1.4. Gene Expression Analysis
2.2. Clinical Study
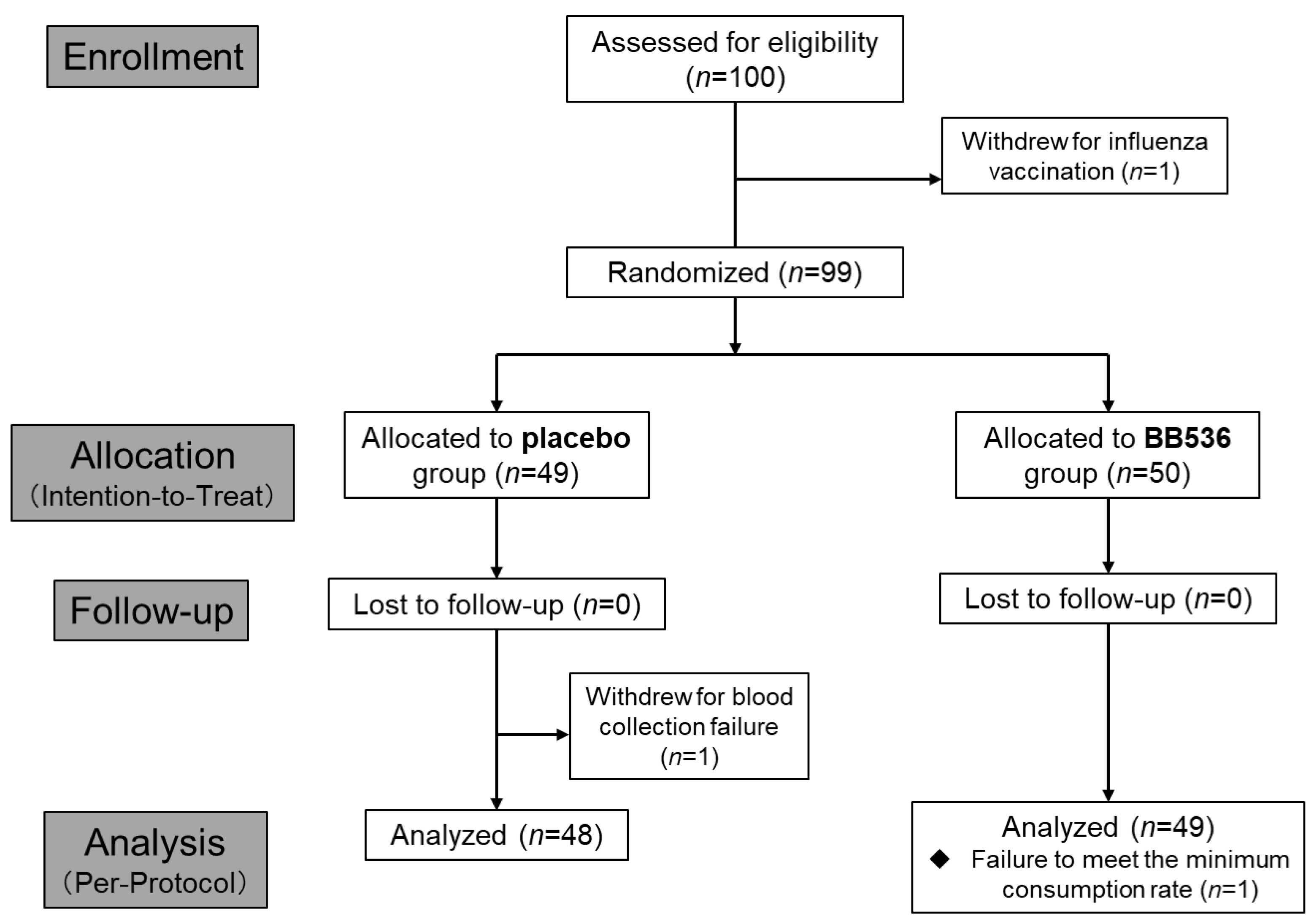
2.2.1. Participants
2.2.2. Study Design and Intervention
2.2.3. FACS Analysis
2.2.4. Measurements of NK Cell Activity, Neutrophilic Phagocytosis, and Bactericidal Activities
2.2.5. Evaluation of Immunological Responses to TLR Ligand of PBMCs Ex Vivo
2.2.6. Measurements of Cytokines in Plasma Isolated from Peripheral Blood
2.3. Statistics
3. Results
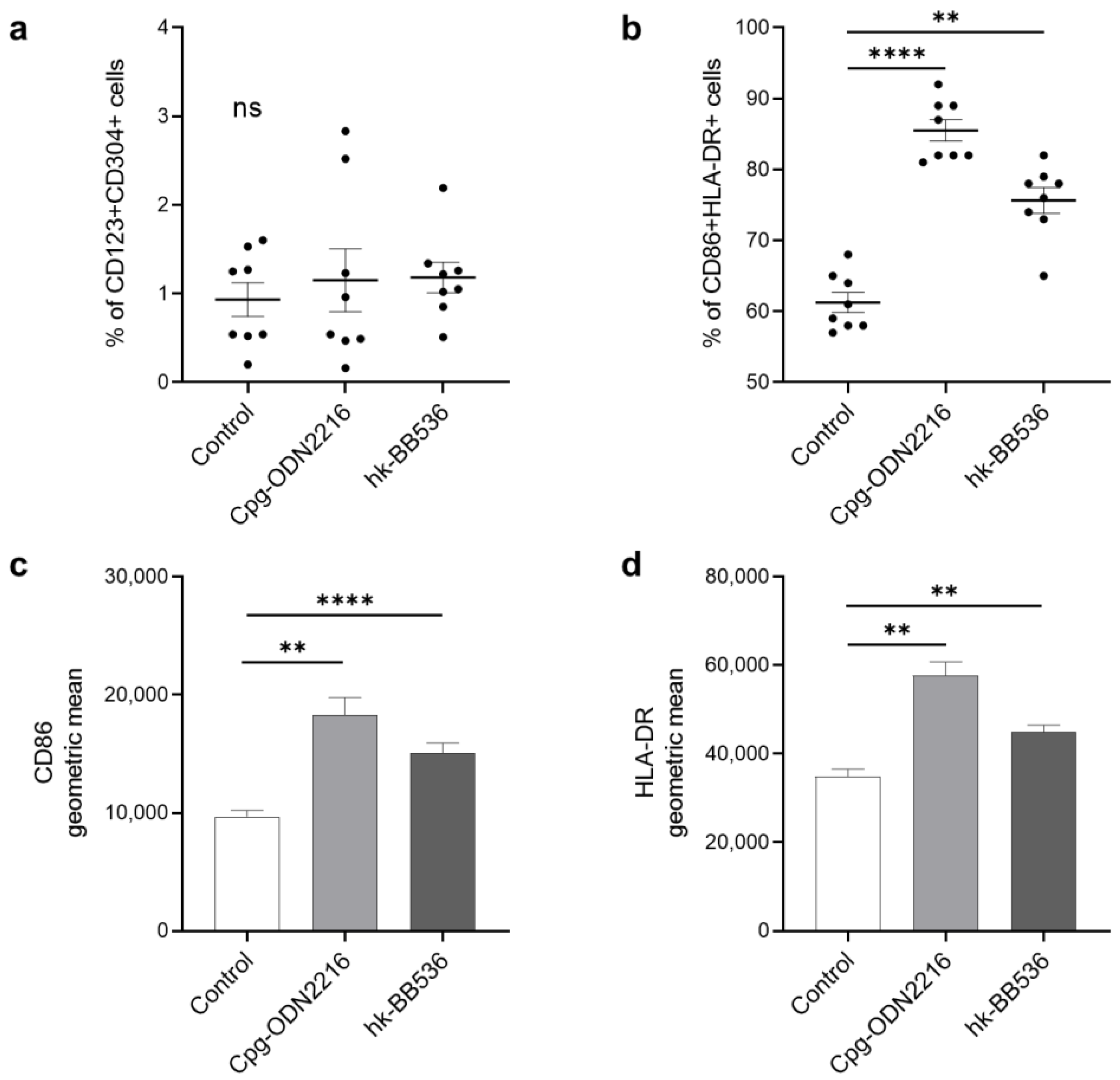
3.1. Heat-Killed BB536 Activated the Surface Markers of pDCs in PBMCs In Vitro
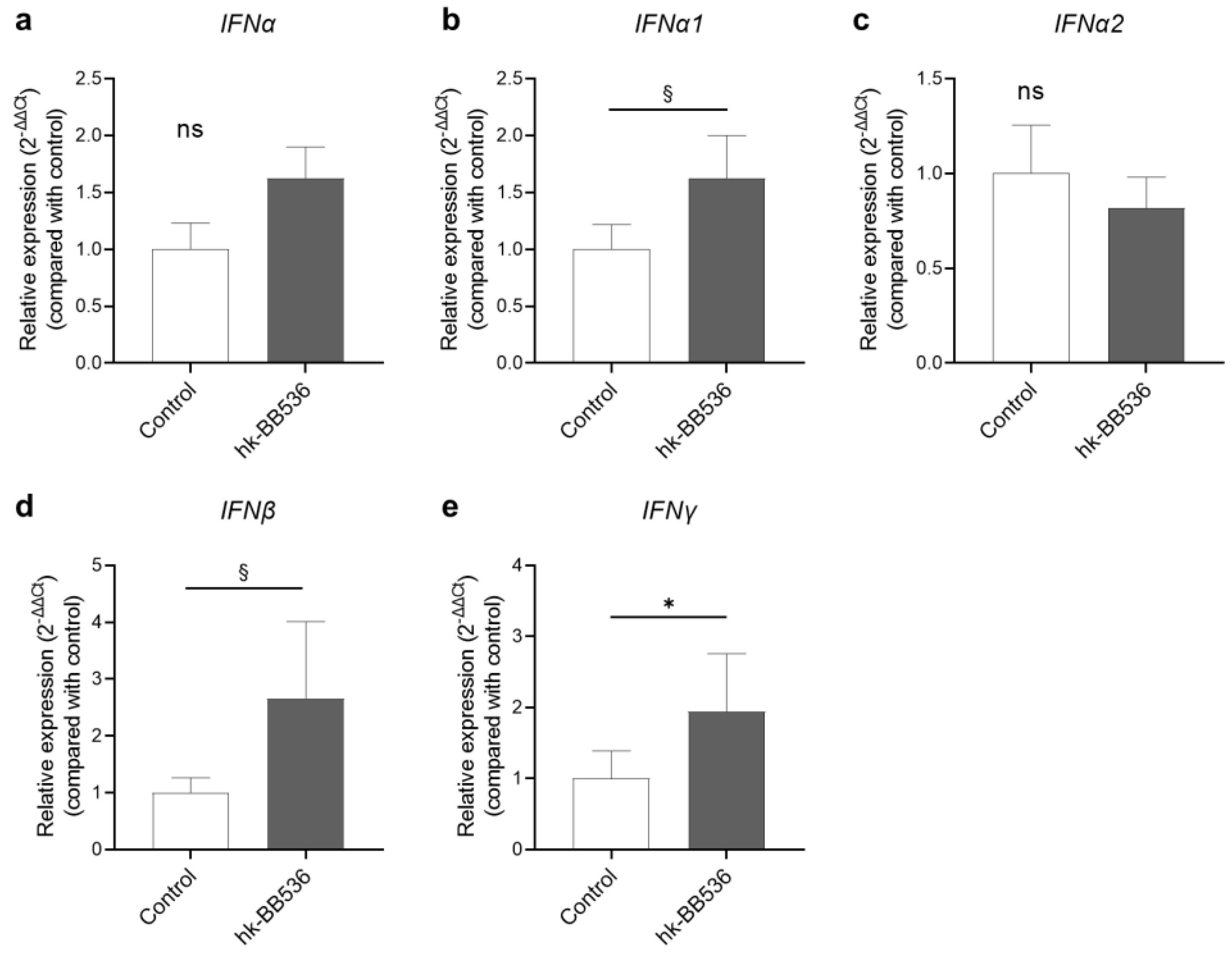
3.2. Effect of Hk-BB536 on Interferon Gene Expression in PBMCs In Vitro
3.3. Initial Participant Characteristics in the Clinical Study
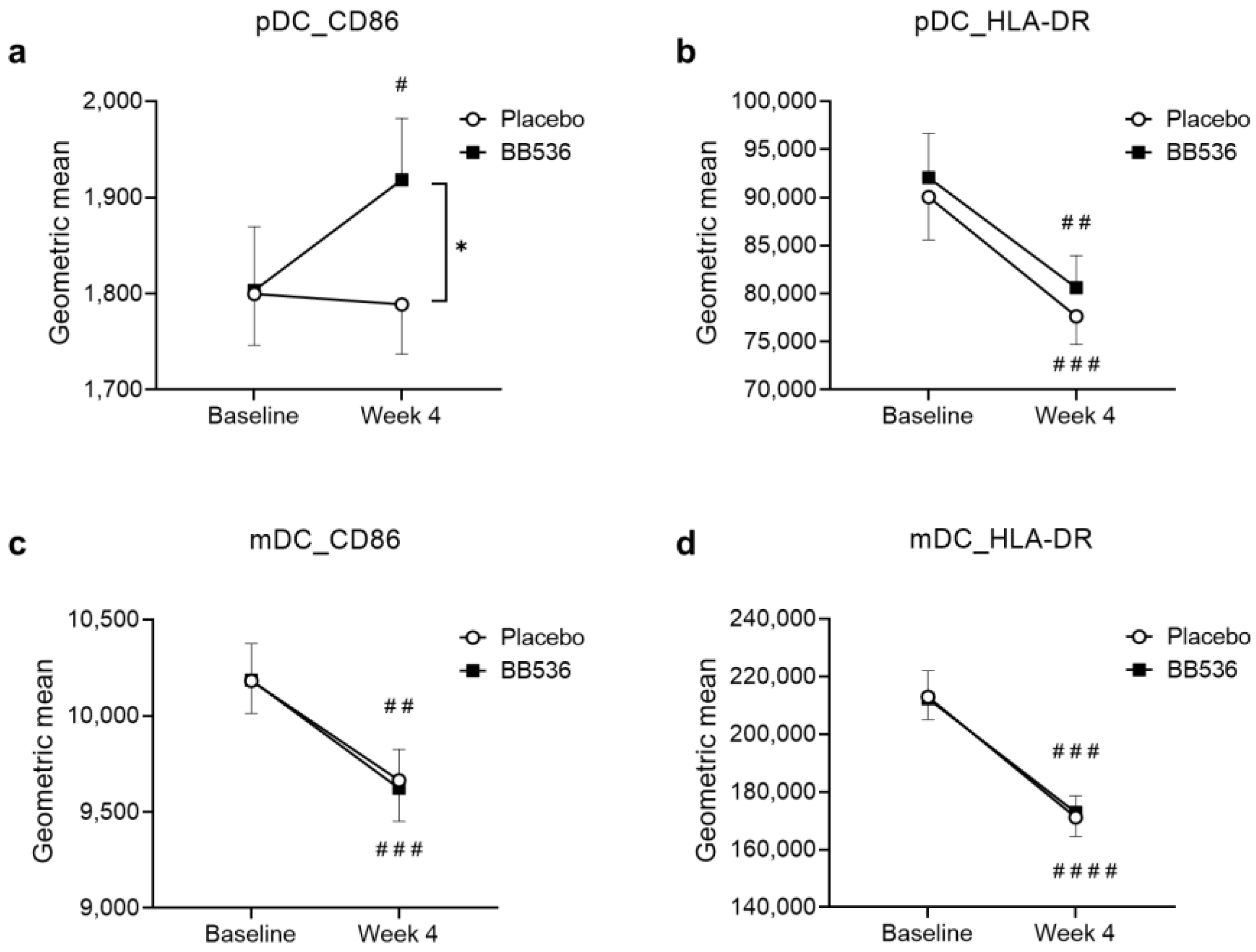
3.4. Effects of BB536 Intake on DC Activity In Vivo
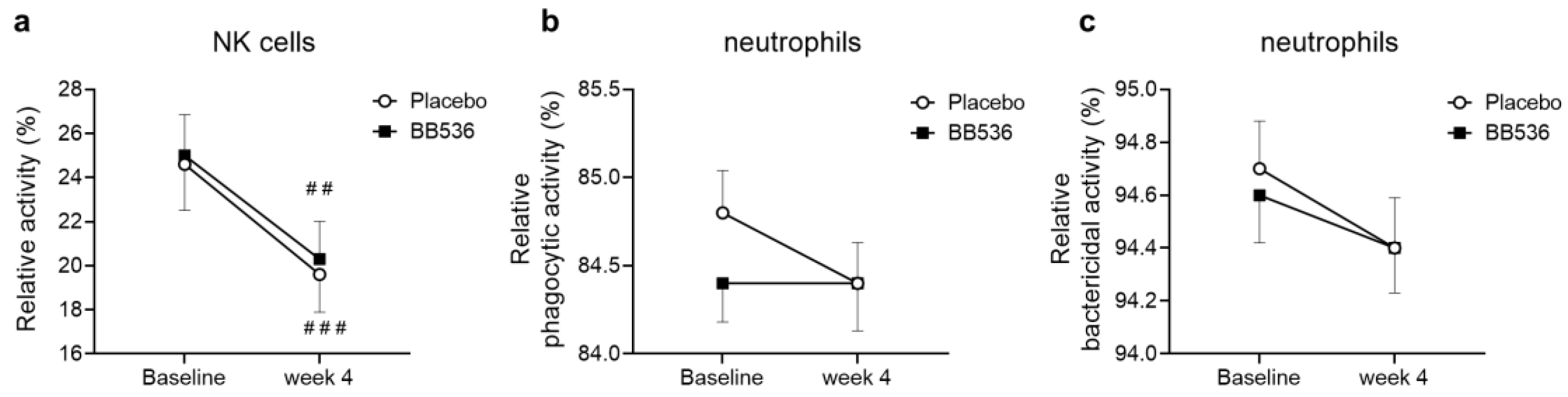
3.5. Effects of BB536 Intake on NK Activity, Neutrophil Phagocytic Activity, and Bactericidal Activity In Vivo
3.6. Effects of BB536 Intake on the IFN Response Ex Vivo
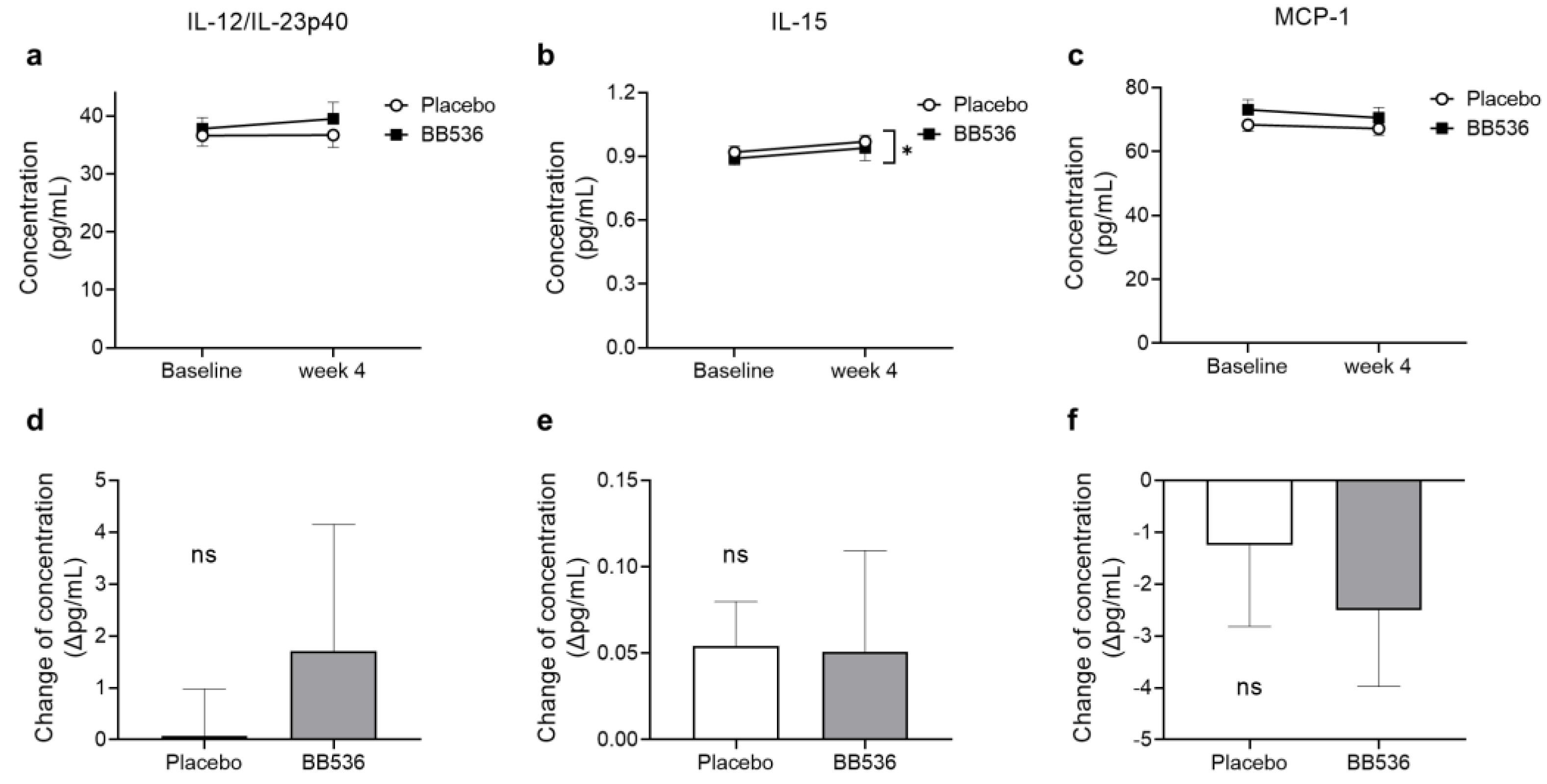
3.7. Effects of BB536 Intake on the IFN Response in Plasma Ex Vivo
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between Microbiota and Immunity in Health and Disease. Cell Res. 2020, 30, 492–506. [Google Scholar] [CrossRef] [PubMed]
- Azad, M.A.K.; Sarker, M.; Wan, D. Immunomodulatory Effects of Probiotics on Cytokine Profiles. BioMed Res. Int. 2019, 2019, 8063647. Available online: https://www.hindawi.com/journals/bmri/2018/8063647/ (accessed on 2 May 2023). [CrossRef] [PubMed]
- Arboleya, S.; Watkins, C.; Stanton, C.; Ross, R.P. Gut Bifidobacteria Populations in Human Health and Aging. Front. Microbiol. 2016, 7, 1204. [Google Scholar] [CrossRef] [PubMed]
- Alessandri, G.; Ossiprandi, M.C.; MacSharry, J.; van Sinderen, D.; Ventura, M. Bifidobacterial Dialogue with Its Human Host and Consequent Modulation of the Immune System. Front. Immunol. 2019, 10, 2348. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, I.; Marchesi, J.R.; Schofield, Z.; Hall, L.J. Exploring the Role of the Microbiota Member Bifidobacterium in Modulating Immune-Linked Diseases. Emerg. Top. Life Sci. 2017, 1, 333–349. [Google Scholar] [CrossRef]
- Wong, C.B.; Odamaki, T.; Xiao, J. Beneficial Effects of Bifidobacterium longum subsp. longum BB536 on Human Health: Modulation of Gut Microbiome as the Principal Action. J. Funct. Foods 2019, 54, 506–519. [Google Scholar] [CrossRef]
- Lau, A.S.-Y.; Yanagisawa, N.; Hor, Y.-Y.; Lew, L.-C.; Ong, J.-S.; Chuah, L.-O.; Lee, Y.-Y.; Choi, S.-B.; Rashid, F.; Wahid, N.; et al. Bifidobacterium longum BB536 Alleviated Upper Respiratory Illnesses and Modulated Gut Microbiota Profiles in Malaysian Pre-School Children. Benef. Microbes 2018, 9, 61–70. [Google Scholar] [CrossRef]
- Namba, K.; Hatano, M.; Yaeshima, T.; Takase, M.; Suzuki, K. Effects of Bifidobacterium longum BB536 Administration on Influenza Infection, Influenza Vaccine Antibody Titer, and Cell-Mediated Immunity in the Elderly. Biosci. Biotechnol. Biochem. 2010, 74, 939–945. [Google Scholar] [CrossRef]
- Wiertsema, S.P.; van Bergenhenegouwen, J.; Garssen, J.; Knippels, L.M.J. The Interplay between the Gut Microbiome and the Immune System in the Context of Infectious Diseases throughout Life and the Role of Nutrition in Optimizing Treatment Strategies. Nutrients 2021, 13, 886. [Google Scholar] [CrossRef]
- O’Keeffe, M.; Mok, W.H.; Radford, K.J. Human Dendritic Cell Subsets and Function in Health and Disease. Cell. Mol. Life Sci. 2015, 72, 4309–4325. [Google Scholar] [CrossRef]
- Alvarez, D.; Vollmann, E.H.; von Andrian, U.H. Mechanisms and Consequences of Dendritic Cell Migration. Immunity 2008, 29, 325–342. [Google Scholar] [CrossRef] [PubMed]
- Randolph, G.J.; Angeli, V.; Swartz, M.A. Dendritic-Cell Trafficking to Lymph Nodes through Lymphatic Vessels. Nat. Rev. Immunol. 2005, 5, 617–628. [Google Scholar] [CrossRef]
- Conrad, C.; Gregorio, J.; Wang, Y.-H.; Ito, T.; Meller, S.; Hanabuchi, S.; Anderson, S.; Atkinson, N.; Ramirez, P.T.; Liu, Y.-J.; et al. Plasmacytoid Dendritic Cells Promote Immunosuppression in Ovarian Cancer via ICOS Costimulation of Foxp3+ T-Regulatory Cells. Cancer Res. 2012, 72, 5240–5249. [Google Scholar] [CrossRef] [PubMed]
- Mittelbrunn, M.; Martínez del Hoyo, G.; López-Bravo, M.; Martín-Cofreces, N.B.; Scholer, A.; Hugues, S.; Fetler, L.; Amigorena, S.; Ardavín, C.; Sánchez-Madrid, F. Imaging of Plasmacytoid Dendritic Cell Interactions with T Cells. Blood 2009, 113, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Fink, K.; Lang, K.S.; Manjarrez-Orduno, N.; Junt, T.; Senn, B.M.; Holdener, M.; Akira, S.; Zinkernagel, R.M.; Hengartner, H. Early Type I Interferon-Mediated Signals on B Cells Specifically Enhance Antiviral Humoral Responses. Eur. J. Immunol. 2006, 36, 2094–2105. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Gregorio, J.D.; Iwahori, T.; Zhang, X.; Choi, O.; Tolentino, L.L.; Prestwood, T.; Carmi, Y.; Engleman, E.G. A Distinct Subset of Plasmacytoid Dendritic Cells Induces Activation and Differentiation of B and T Lymphocytes. Proc. Natl. Acad. Sci. USA 2017, 114, 1988–1993. [Google Scholar] [CrossRef] [PubMed]
- Crother, T.R.; Ma, J.; Jupelli, M.; Chiba, N.; Chen, S.; Slepenkin, A.; Alsabeh, R.; Peterson, E.; Shimada, K.; Arditi, M. Plasmacytoid Dendritic Cells Play a Role for Effective Innate Immune Responses during Chlamydia Pneumoniae Infection in Mice. PLoS ONE 2012, 7, e48655. [Google Scholar] [CrossRef]
- Guillerey, C.; Mouriès, J.; Polo, G.; Doyen, N.; Law, H.K.W.; Chan, S.; Kastner, P.; Leclerc, C.; Dadaglio, G. Pivotal Role of Plasmacytoid Dendritic Cells in Inflammation and NK-Cell Responses after TLR9 Triggering in Mice. Blood 2012, 120, 90–99. [Google Scholar] [CrossRef]
- Nguyen, K.B.; Salazar-Mather, T.P.; Dalod, M.Y.; Deusen, J.B.V.; Wei, X.; Liew, F.Y.; Caligiuri, M.A.; Durbin, J.E.; Biron, C.A. Coordinated and Distinct Roles for IFN-Aβ, IL-12, and IL-15 Regulation of NK Cell Responses to Viral Infection. J. Immunol. 2002, 169, 4279–4287. [Google Scholar] [CrossRef]
- Izumo, T.; Maekawa, T.; Ida, M.; Kishi, A.; Akatani, K.; Kitagawa, Y.; Kiso, Y. Effect of Lactobacillus Pentosus S-PT84 Ingestion on IFN-α Production from Plasmacytoid Dendritic Cells by Virus Stimulation. Biosci. Biotechnol. Biochem. 2011, 75, 370–372. [Google Scholar] [CrossRef]
- Sugimura, T.; Jounai, K.; Ohshio, K.; Tanaka, T.; Suwa, M.; Fujiwara, D. Immunomodulatory Effect of Lactococcus Lactis JCM5805 on Human Plasmacytoid Dendritic Cells. Clin. Immunol. 2013, 149, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Akatsu, H.; Iwabuchi, N.; Xiao, J.-Z.; Matsuyama, Z.; Kurihara, R.; Okuda, K.; Yamamoto, T.; Maruyama, M. Clinical Effects of Probiotic Bifidobacterium longum BB536 on Immune Function and Intestinal Microbiota in Elderly Patients Receiving Enteral Tube Feeding. JPEN J. Parenter. Enter. Nutr. 2013, 37, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Arimura, K.; Takagi, H.; Uto, T.; Fukaya, T.; Nakamura, T.; Choijookhuu, N.; Hishikawa, Y.; Yamashita, Y.; Sato, K. Crucial Role of Plasmacytoid Dendritic Cells in the Development of Acute Colitis through the Regulation of Intestinal Inflammation. Mucosal Immunol. 2017, 10, 957–970. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Gaugler, B.; Mohty, M.; Malard, F. Plasmacytoid Dendritic Cell Biology and Its Role in Immune-Mediated Diseases. Clin. Transl. Immunol. 2020, 9, e1139. [Google Scholar] [CrossRef] [PubMed]
- Greene, J.L.; Leytze, G.M.; Emswiler, J.; Peach, R.; Bajorath, J.; Cosand, W.; Linsley, P.S. Covalent Dimerization of CD28/CTLA-4 and Oligomerization of CD80/CD86 Regulate T Cell Costimulatory Interactions. J. Biol. Chem. 1996, 271, 26762–26771. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, A.; Waters, E.; Rowshanravan, B.; Hinze, C.; Williams, C.; Janman, D.; Fox, T.A.; Booth, C.; Pesenacker, A.M.; Halliday, N.; et al. Differences in CD80 and CD86 Transendocytosis Reveal CD86 as a Key Target for CTLA-4 Immune Regulation. Nat. Immunol. 2022, 23, 1365–1378. [Google Scholar] [CrossRef]
- Ahout, I.M.L.; Jans, J.; Haroutiounian, L.; Simonetti, E.R.; van der Gaast-de Jongh, C.; Diavatopoulos, D.A.; de Jonge, M.I.; de Groot, R.; Ferwerda, G. Reduced Expression of HLA-DR on Monocytes During Severe Respiratory Syncytial Virus Infections. Pediatr. Infect. Dis. J. 2016, 35, e89. [Google Scholar] [CrossRef]
- Gay, D.; Maddon, P.; Sekaly, R.; Talle, M.A.; Godfrey, M.; Long, E.; Goldstein, G.; Chess, L.; Axel, R.; Kappler, J.; et al. Functional Interaction between Human T-Cell Protein CD4 and the Major Histocompatibility Complex HLA-DR Antigen. Nature 1987, 328, 626–629. [Google Scholar] [CrossRef]
- Wittmann, A.; Autenrieth, I.B.; Frick, J.-S. Plasmacytoid Dendritic Cells Are Crucial in Bifidobacterium Adolescentis-Mediated Inhibition of Yersinia Enterocolitica Infection. PLoS ONE 2013, 8, e71338. [Google Scholar] [CrossRef]
- Keppler, S.J.; Rosenits, K.; Koegl, T.; Vucikuja, S.; Aichele, P. Signal 3 Cytokines as Modulators of Primary Immune Responses during Infections: The Interplay of Type I IFN and IL-12 in CD8 T Cell Responses. PLoS ONE 2012, 7, e40865. [Google Scholar] [CrossRef]
- Siegal, F.P.; Kadowaki, N.; Shodell, M.; Fitzgerald-Bocarsly, P.A.; Shah, K.; Ho, S.; Antonenko, S.; Liu, Y.-J. The Nature of the Principal Type 1 Interferon-Producing Cells in Human Blood. Science 1999, 284, 1835–1837. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Kanzler, H.; Duramad, O.; Cao, W.; Liu, Y.-J. Specialization, Kinetics, and Repertoire of Type 1 Interferon Responses by Human Plasmacytoid Predendritic Cells. Blood 2006, 107, 2423–2431. [Google Scholar] [CrossRef] [PubMed]
- Contractor, N.; Louten, J.; Kim, L.; Biron, C.A.; Kelsall, B.L. Cutting Edge: Peyer’s Patch Plasmacytoid Dendritic Cells (PDCs) Produce Low Levels of Type I Interferons: Possible Role for IL-10, TGFβ, and Prostaglandin E2 in Conditioning a Unique Mucosal PDC Phenotype1. J. Immunol. 2007, 179, 2690–2694. [Google Scholar] [CrossRef] [PubMed]
- Medina, M.; Izquierdo, E.; Ennahar, S.; Sanz, Y. Differential Immunomodulatory Properties of Bifidobacterium Logum Strains: Relevance to Probiotic Selection and Clinical Applications. Clin. Exp. Immunol. 2007, 150, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Bradley, L.M.; Dalton, D.K.; Croft, M. A Direct Role for IFN-Gamma in Regulation of Th1 Cell Development. J. Immunol. 1996, 157, 1350–1358. [Google Scholar] [CrossRef] [PubMed]
- Ueki, H.; Furusawa, Y.; Iwatsuki-Horimoto, K.; Imai, M.; Kabata, H.; Nishimura, H.; Kawaoka, Y. Effectiveness of Face Masks in Preventing Airborne Transmission of SARS-CoV-2. mSphere 2020, 5, e00637-20. [Google Scholar] [CrossRef] [PubMed]
- Lévi, F.A.; Canon, C.; Touitou, Y.; Reinberg, A.; Mathé, G. Seasonal Modulation of the Circadian Time Structure of Circulating T and Natural Killer Lymphocyte Subsets from Healthy Subjects. Available online: https://www.jci.org/articles/view/113333/pdf (accessed on 14 August 2023).
- Courtney, J.M.; Bax, A. Hydrating the Respiratory Tract: An Alternative Explanation Why Masks Lower Severity of COVID-19. Biophys. J. 2021, 120, 994–1000. [Google Scholar] [CrossRef]
- Lee, A.; Lee, Y.J.; Yoo, H.J.; Kim, M.; Chang, Y.; Lee, D.S.; Lee, J.H. Consumption of Dairy Yogurt Containing Lactobacillus Paracasei Ssp. Paracasei, Bifidobacterium Animalis Ssp. Lactis and Heat-Treated Lactobacillus Plantarum Improves Immune Function Including Natural Killer Cell Activity. Nutrients 2017, 9, 558. [Google Scholar] [CrossRef]
- Finamore, A.; Roselli, M.; Donini, L.; Brasili, D.E.; Rami, R.; Carnevali, P.; Mistura, L.; Pinto, A.; Giusti, A.; Mengheri, E. Supplementation with Bifidobacterium longum Bar33 and Lactobacillus Helveticus Bar13 Mixture Improves Immunity in Elderly Humans (over 75 Years) and Aged Mice. Nutrition 2019, 63–64, 184–192. [Google Scholar] [CrossRef]
- Bae, W.-Y.; Min, H.; Shin, S.L.; Kim, T.-R.; Lee, H.; Sohn, M.; Park, K.S. Effect of Orally Administered Heat-Treated Lactobacillus Plantarum LM1004 on the Innate Immune System: A Randomized, Placebo-Controlled, Double-Blind Study. J. Funct. Foods 2022, 98, 105293. [Google Scholar] [CrossRef]
- Marcial, G.E.; Ford, A.L.; Haller, M.J.; Gezan, S.A.; Harrison, N.A.; Cai, D.; Meyer, J.L.; Perry, D.J.; Atkinson, M.A.; Wasserfall, C.H.; et al. Lactobacillus Johnsonii N6.2 Modulates the Host Immune Responses: A Double-Blind, Randomized Trial in Healthy Adults. Front. Immunol. 2017, 8, 655. [Google Scholar] [CrossRef] [PubMed]
- Seifert, S.; Bub, A.; Franz, C.M.A.P.; Watzl, B. Probiotic Lactobacillus Casei Shirota Supplementation Does Not Modulate Immunity in Healthy Men with Reduced Natural Killer Cell Activity1–3. J. Nutr. 2011, 141, 978–984. [Google Scholar] [CrossRef] [PubMed]
- Gleeson, M.; Bishop, N.C.; Struszczak, L. Effects of Lactobacillus Casei Shirota Ingestion on Common Cold Infection and Herpes Virus Antibodies in Endurance Athletes: A Placebo-Controlled, Randomized Trial. Eur. J. Appl. Physiol. 2016, 116, 1555–1563. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| β-ACTIN | GAGCGGGAAATCGTGCGTGACATT | TGCCCAGGAAGGAAGGCTGGAAGA |
| IFNα | GACCAGGAGACACGGAATGT | GATGTAATCCTTGCCGTCGT |
| IFNα1 | GCAAGCCCAGAAGTATCTGC | ACTGGTTGCCATCAAACTCC |
| IFNα2 | AAATACAGCCCTTGTGCCTGG | GGTGAGCTGGCATACGAATCA |
| IFNβ | AAGGCCAAGGAGTACAGTC | ATCTTCAGTTTCGGAGGTAA |
| IFNγ | TGACCAGAGCATCCAAAAGA | CTCTTCGACCTCGAAACAGC |
| Characteristic | Placebo (n = 49) | BB536 (n = 50) | p-Value |
|---|---|---|---|
| Female (%) | 28 (58.3%) | 25 (50.0%) | 0.548 |
| Age (years) | 45.9 ± 10.8 | 46.2 ± 10.5 | 0.903 |
| BMI (kg/m2) | 21.78 ± 2.1 | 21.43 ± 2.03 | 0.399 |
| Smoker (%) | 1 (2.08%) | 3 (6.0%) | 0.617 |
| Sleep time (hour) | 6.68 ± 0.75 | 6.66 ± 0.72 | 0.873 |
| White blood cell (/μL) | 5058.4 ± 1048.8 | 5149.2 ± 1198.8 | 0.689 |
| Platelet count (×104/μL) | 27.29 ± 4.97 | 26.45 ± 4.57 | 0.385 |
| Total serum protein (g/dL) | 7.24 ± 0.35 | 7.29 ± 0.36 | 0.457 |
| Albumin (g/dL) | 4.48 ± 0.26 | 4.52 ± 0.27 | 0.425 |
| γ-GTP (U/L) | 21.6 ± 11.7 | 22.8 ± 11.3 | 0.613 |
| HbA1c (%) | 5.18 ± 0.19 | 5.26 ± 0.26 | 0.073 |
| Triglyceride (mg/dL) | 72.4 ± 39.6 | 68.3 ± 32.1 | 0.570 |
| HDL-Cholesterol (mg/dL) | 71.2 ± 14.7 | 68.9 ± 16.6 | 0.482 |
| LDL-Cholesterol (mg/dL) | 115.9 ± 21.9 | 117.0 ± 25.1 | 0.816 |
| Total-Cholesterol (mg/dL) | 204.6 ± 26.1 | 200.2 ± 24.9 | 0.402 |
| Baseline 1 | Week4 | |||||
|---|---|---|---|---|---|---|
| Placebo | BB536 | p-Value | Placebo | BB536 | p-Value | |
| IFNα | 4.671 ± 0.249 | 4.702 ± 0.237 | 0.943 | 4.883 ± 0.248 | 4.623 ± 0.188 | 0.209 |
| IFNα1 | 4.808 ± 0.246 | 4.868 ± 0.236 | 0.937 | 5.128 ± 0.257 | 4.870 ± 0.185 | 0.218 |
| IFNα2 | 5.700 ± 0.300 | 5.862 ± 0.292 | 0.901 | 5.898 ± 0.286 | 5.690 ± 0.228 | 0.421 |
| IFNβ | 4.980 ± 0.230 | 5.019 ± 0.218 | 0.811 | 5.468 ± 0.265 | 5.192 ± 0.207 | 0.268 |
| IFNγ | 4.979 ± 0.331 | 4.973 ± 0.236 | 0.914 | 6.249 ± 0.346 | 6.242 ± 0.191 | 0.736 |
| Baseline 1 | Week4 | |||||
|---|---|---|---|---|---|---|
| Placebo | BB536 | p-Value | Placebo | BB536 | p-Value | |
| IFNα | 4.671 ± 0.249 | 4.702 ± 0.237 | 0.943 | 4.883 ± 0.248 | 4.623 ± 0.188 | 0.209 |
| IFNα1 | 4.808 ± 0.246 | 4.868 ± 0.236 | 0.937 | 5.128 ± 0.257 | 4.870 ± 0.185 | 0.218 |
| IFNα2 | 5.700 ± 0.300 | 5.862 ± 0.292 | 0.901 | 5.898 ± 0.286 | 5.690 ± 0.228 | 0.421 |
| IFNβ | 4.980 ± 0.230 | 5.019 ± 0.218 | 0.811 | 5.468 ± 0.265 | 5.192 ± 0.207 | 0.268 |
| IFNγ | 4.979 ± 0.331 | 4.973 ± 0.236 | 0.914 | 6.249 ± 0.346 | 6.242 ± 0.191 | 0.736 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Arai, S.; Kato, K.; Iwabuchi, S.; Iwabuchi, N.; Muto, N.; Motobayashi, H.; Ebihara, S.; Tanaka, M.; Hashimoto, S. The Potential Immunomodulatory Effect of Bifidobacterium longum subsp. longum BB536 on Healthy Adults through Plasmacytoid Dendritic Cell Activation in the Peripheral Blood. Nutrients 2024, 16, 42. https://doi.org/10.3390/nu16010042
Li Y, Arai S, Kato K, Iwabuchi S, Iwabuchi N, Muto N, Motobayashi H, Ebihara S, Tanaka M, Hashimoto S. The Potential Immunomodulatory Effect of Bifidobacterium longum subsp. longum BB536 on Healthy Adults through Plasmacytoid Dendritic Cell Activation in the Peripheral Blood. Nutrients. 2024; 16(1):42. https://doi.org/10.3390/nu16010042
Chicago/Turabian StyleLi, Yiran, Satoshi Arai, Kumiko Kato, Sadahiro Iwabuchi, Noriyuki Iwabuchi, Natsumi Muto, Hideki Motobayashi, Shukuko Ebihara, Miyuki Tanaka, and Shinichi Hashimoto. 2024. "The Potential Immunomodulatory Effect of Bifidobacterium longum subsp. longum BB536 on Healthy Adults through Plasmacytoid Dendritic Cell Activation in the Peripheral Blood" Nutrients 16, no. 1: 42. https://doi.org/10.3390/nu16010042
APA StyleLi, Y., Arai, S., Kato, K., Iwabuchi, S., Iwabuchi, N., Muto, N., Motobayashi, H., Ebihara, S., Tanaka, M., & Hashimoto, S. (2024). The Potential Immunomodulatory Effect of Bifidobacterium longum subsp. longum BB536 on Healthy Adults through Plasmacytoid Dendritic Cell Activation in the Peripheral Blood. Nutrients, 16(1), 42. https://doi.org/10.3390/nu16010042