
The Association between Dietary Iron Intake, SNP of the MTNR1B rs10830963, and Glucose Metabolism in Chinese Population

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
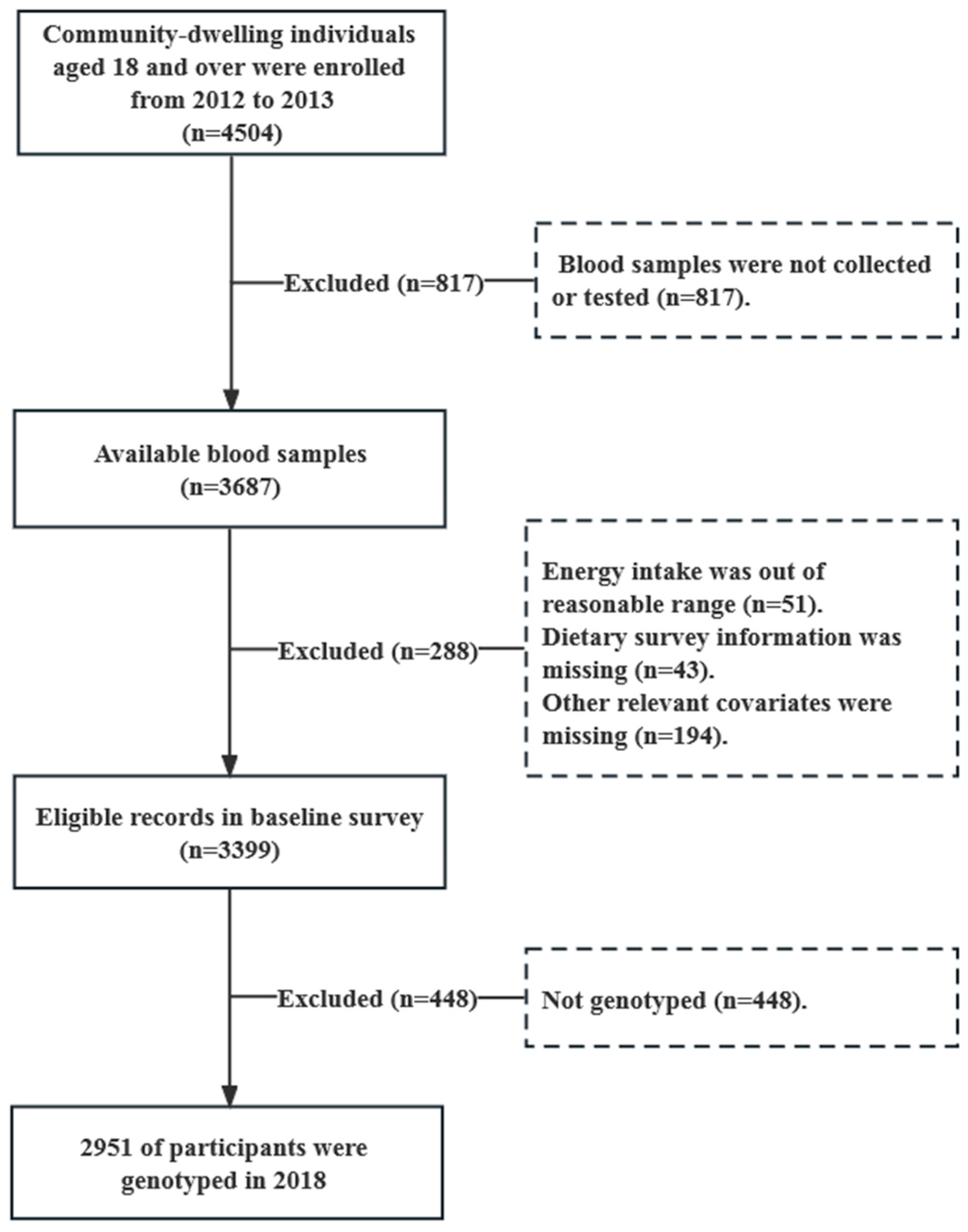
2.1. Study Participants
2.2. Dietary Assessment
2.3. Laboratory Measurements
2.4. Genotyping
2.5. Identification of Elevated Fasting Glucose and Calculation of HOMA2-IR
2.6. Potential Confounders
2.7. Statistical Analysis
3. Results
3.1. Characteristics of the Participants
3.2. Genotypes of the MTNR1B rs10830963
3.3. Associations between Dietary Iron and Risk on the Glucose Metabolism When Stratified by G Allele on the rs10830963 Site of MTNR1B Gene
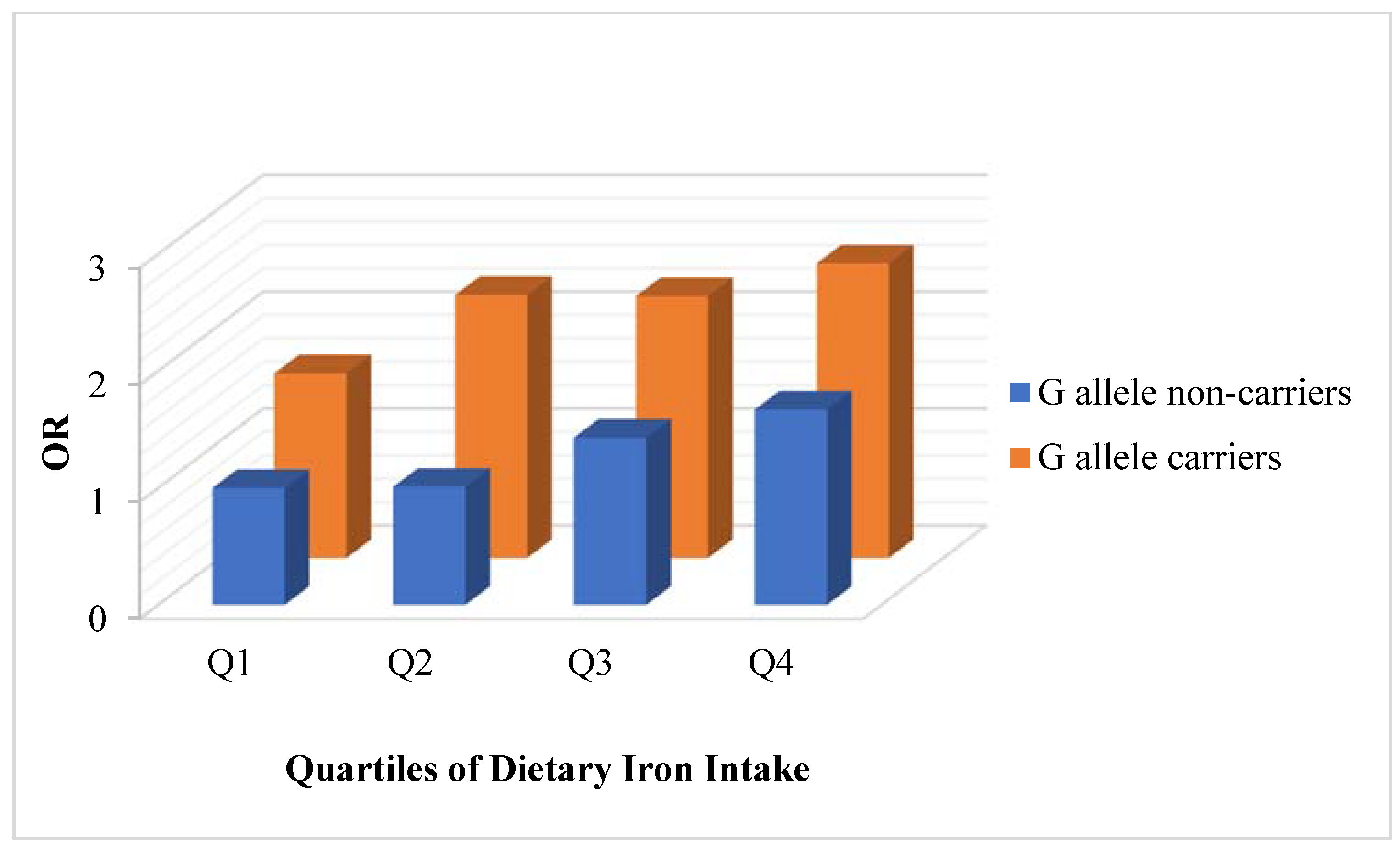
3.3.1. Associations between Dietary Iron and Risk on Elevated Fasting Glucose Stratified by the rs10830963 Risk Allele in the MTNR1B Gene
3.3.2. Associations between Dietary Iron and Risk on Fasting Glucose Stratified by the rs10830963 Risk Allele in the MTNR1B Gene
3.3.3. Associations between Dietary Iron and Risk on HbA1c Stratified by the rs10830963 Risk Allele in the MTNR1B Gene
3.3.4. Associations between Dietary Iron and Risk on HOMA2-IR Stratified by the rs10830963 Risk Allele in the MTNR1B Gene
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9(th) edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.H.; Dou, K.F.; Song, W.J. Prevalence of Diabetes among Men and Women in China. N. Engl. J. Med. 2010, 362, 1090–1101. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Teng, D.; Shi, X.; Qin, G.; Qin, Y.; Quan, H.; Shi, B.; Sun, H.; Ba, J.; Chen, B.; et al. Prevalence of diabetes recorded in mainland China using 2018 diagnostic criteria from the American Diabetes Association: National cross sectional study. BMJ 2020, 369, m997. [Google Scholar] [CrossRef] [PubMed]
- Simcox, J.A.; McClain, D.A. Iron and diabetes risk. Cell Metab. 2013, 17, 329–341. [Google Scholar] [CrossRef]
- Li, S.Y.; Wang, F.; Lu, X.T.; Zhong, R.H.; Long, J.A.; Fang, A.P.; Zhu, H.L. Dietary iron intake and the risk of type 2 diabetes mellitus in middle-aged and older adults in urban China: A prospective cohort study. Br. J. Nutr. 2021, 126, 1091–1099. [Google Scholar] [CrossRef]
- Jung, D.H.; Hong, K.W.; Park, B.; Lee, Y.J. Dietary iron to total energy intake ratio and type 2 diabetes incidence in a longitudinal 12-year analysis of the Korean Genome and Epidemiology Cohort Study. Eur. J. Nutr. 2021, 60, 4453–4461. [Google Scholar] [CrossRef]
- Rajpathak, S.N.; Crandall, J.P.; Wylie-Rosett, J.; Kabat, G.C.; Rohan, T.E.; Hu, F.B. The role of iron in type 2 diabetes in humans. Biochim. Biophys. Acta 2009, 1790, 671–681. [Google Scholar] [CrossRef]
- Jaworek, J.; Nawrot-Porabka, K.; Leja-Szpak, A.; Bonior, J.; Szklarczyk, J.; Kot, M.; Konturek, S.J.; Pawlik, W.W. Melatonin as modulator of pancreatic enzyme secretion and pancreatoprotector. J. Physiol. Pharmacol. 2007, 58 (Suppl. 6), 65–80. [Google Scholar]
- Ramracheya, R.D.; Muller, D.S.; Squires, P.E.; Brereton, H.; Sugden, D.; Huang, G.C.; Amiel, S.A.; Jones, P.M.; Persaud, S.J. Function and expression of melatonin receptors on human pancreatic islets. J. Pineal. Res. 2008, 44, 273–279. [Google Scholar] [CrossRef]
- Ronn, T.; Wen, J.; Yang, Z.; Lu, B.; Du, Y.; Groop, L.; Hu, R.; Ling, C. A common variant in MTNR1B, encoding melatonin receptor 1B, is associated with type 2 diabetes and fasting plasma glucose in Han Chinese individuals. Diabetologia 2009, 52, 830–833. [Google Scholar] [CrossRef]
- Liu, C.; Wu, Y.; Li, H.; Qi, Q.; Langenberg, C.; Loos, R.J.; Lin, X. MTNR1B rs10830963 is associated with fasting plasma glucose, HbA1C and impaired beta-cell function in Chinese Hans from Shanghai. BMC Med. Genet. 2010, 11, 59. [Google Scholar] [CrossRef] [PubMed]
- Sparso, T.; Bonnefond, A.; Andersson, E.; Bouatia-Naji, N.; Holmkvist, J.; Wegner, L.; Grarup, N.; Gjesing, A.P.; Banasik, K.; Cavalcanti-Proenca, C.; et al. G-allele of intronic rs10830963 in MTNR1B confers increased risk of impaired fasting glycemia and type 2 diabetes through an impaired glucose-stimulated insulin release: Studies involving 19,605 Europeans. Diabetes 2009, 58, 1450–1456. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Shi, Y.; You, L.; Wang, L.; Chen, Z.J. Association of rs10830963 and rs10830962 SNPs in the melatonin receptor (MTNR1B) gene among Han Chinese women with polycystic ovary syndrome. Mol. Hum. Reprod. 2011, 17, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Prokopenko, I.; Langenberg, C.; Florez, J.C.; Saxena, R.; Soranzo, N.; Thorleifsson, G.; Loos, R.J.; Manning, A.K.; Jackson, A.U.; Aulchenko, Y.; et al. Variants in MTNR1B influence fasting glucose levels. Nat. Genet. 2009, 41, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Gaulton, K.J.; Ferreira, T.; Lee, Y.; Raimondo, A.; Magi, R.; Reschen, M.E.; Mahajan, A.; Locke, A.; Rayner, N.W.; Robertson, N.; et al. Genetic fine mapping and genomic annotation defines causal mechanisms at type 2 diabetes susceptibility loci. Nat. Genet. 2015, 47, 1415–1425. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.R.; Shah, Y.M. Iron homeostasis in the liver. Compr. Physiol. 2013, 3, 315–330. [Google Scholar] [CrossRef]
- Zhu, Z.; Wang, Z.; Zang, J.; Lu, Y.; Xiao, Z.; Zheng, G.; Wu, F. The SNP rs516946 Interacted in the Association of MetS with Dietary Iron among Chinese Males but Not Females. Nutrients 2022, 14, 2024. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, G.; Pan, X. Chinese Food Composition Book 1, 2nd ed.; Peking University Medical Press: Beijing, China, 2009. [Google Scholar]
- Yang, Y.; Wang, G.; Pan, X. Chinese Food Composition Book 2, 1st ed.; Peking University Medical Press: Beijing, China, 2005. [Google Scholar]
- Chinese Nutrition Society. Chinese Dietary Reference Intakes (2013 Edition); Science Press: Beijing, China, 2014. [Google Scholar]
- Grundy, S.M.; Cleeman, J.I.; Daniels, S.R.; Donato, K.A.; Eckel, R.H.; Franklin, B.A.; Gordon, D.J.; Krauss, R.M.; Savage, P.J.; Smith, S.C., Jr.; et al. Diagnosis and management of the metabolic syndrome: An American Heart Association/National Heart, Lung, and Blood Institute Scientific Statement. Circulation 2005, 112, 2735–2752. [Google Scholar] [CrossRef]
- Wallace, T.M.; Levy, J.C.; Matthews, D.R. Use and Abuse of HOMA Modeling. Diabetes Care 2004, 27, 1487–1495. [Google Scholar] [CrossRef]
- Zaret, K.S.; Carroll, J.S. Pioneer transcription factors: Establishing competence for gene expression. Genes. Dev. 2011, 25, 2227–2241. [Google Scholar] [CrossRef]
- Gao, N.; LeLay, J.; Vatamaniuk, M.Z.; Rieck, S.; Friedman, J.R.; Kaestner, K.H. Dynamic regulation of Pdx1 enhancers by Foxa1 and Foxa2 is essential for pancreas development. Genes. Dev. 2008, 22, 3435–3448. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.S.; Friedman, J.R.; Fulmer, J.T.; Kaestner, K.H. The initiation of liver development is dependent on Foxa transcription factors. Nature 2005, 435, 944–947. [Google Scholar] [CrossRef]
- Tuomainen, T.P.; Nyyssönen, K.; Salonen, R.; Tervahauta, A.; Korpela, H.; Lakka, T.; Kaplan, G.A.; Salonen, J.T. Body Iron Stores Are Associated with Serum Insulin and Blood Glucose Concentrations: Population study in 1,013 eastern Finnish men. Diabetes Care 1997, 20, 426–428. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Y.; Babitt, J.L. Liver iron sensing and body iron homeostasis. Blood 2019, 133, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Nelson, L.M.; Ward, K.; Ogilvie, J.W. Genetic variants in melatonin synthesis and signaling pathway are not associated with adolescent idiopathic scoliosis. Spine 2011, 36, 37–40. [Google Scholar] [CrossRef] [PubMed]
- The 1000Genomes Project: Allele Frequency of MTNR1B SNP rs10830963. Available online: https://www.ncbi.nlm.nih.gov/snp/rs10830963#seq_hash (accessed on 20 October 2022).
- Tan, X.; Ciuculete, D.M.; Schiöth, H.B.; Benedict, C. Associations between chronotype, MTNR1B genotype and risk of type 2 diabetes in UK Biobank. J. Intern Med. 2020, 287, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Jabbari, F.; Mohseni Bandpei, A.; Daneshpour, M.S.; Shahsavani, A.; Hashemi Nazari, S.S.; Faraji Sabokbar, H.; Azizi, F. Role of Air Pollution and rs10830963 Polymorphism on the Incidence of Type 2 Diabetes: Tehran Cardiometabolic Genetic Study. J. Diabetes Res. 2020, 2020, 2928618. [Google Scholar] [CrossRef]
- Olsson, L.; Pettersen, E.; Ahlbom, A.; Carlsson, S.; Midthjell, K.; Grill, V. No effect by the common gene variant rs10830963 of the melatonin receptor 1B on the association between sleep disturbances and type 2 diabetes: Results from the Nord-Trøndelag Health Study. Diabetologia 2011, 54, 1375–1378. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| All | G allele Non-Carriers | G allele Carriers 1 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Male | Female | All | Male | Female | All | Male | Female | All | |
| n (%) | 1403 (47.5) | 1548 (52.5) | 2951 (100.0) | 452 (47.7) | 495 (52.3) | 947 (32.1) | 951 (47.5) | 1053 (52.5) | 2004 (67.9) |
| Age, % | |||||||||
| 15−44 years | 426 (30.4) | 503 (32.5) | 929 (31.5) | 125 (27.6) | 169 (34.1) | 294 (31.0) | 301 (31.7) | 334 (31.7) | 635 (31.7) |
| 45–59 years | 502 (35.8) | 555 (35.9) | 1057 (35.8) | 168 (37.2) | 185 (37.4) | 353 (37.3) | 334 (35.1) | 370 (35.2) | 704 (35.1) |
| 60− years | 475 (33.8) | 490 (31.6) | 965 (32.7) | 159 (35.2) | 141 (28.5) | 300 (31.7) | 316 (33.2) | 349 (33.1) | 665 (33.2) |
| Annual Household Income, % | |||||||||
| Above average level (RMB > 60,000) | 772 (55.0) | 879 (56.8) | 1651 (56.0) | 229 (50.7) | 287 (58.0) | 516 (54.5) | 543 (57.1) | 592 (56.2) | 1135 (56.6) |
| Average level (RMB 30,000–59,999) | 453 (32.3) | 466 (30.1) | 919 (31.1) | 164 (36.3) | 142 (28.7) | 306 (32.3) | 289 (30.4) | 324 (30.8) | 613 (30.6) |
| Below average level (RMB < 30,000) | 106 (7.6) | 132 (8.5) | 238 (8.0) | 40 (8.8) | 37 (7.4) | 77 (8.1) | 66 (6.9) | 95 (9.0) | 161 (8.0) |
| No answer | 72 (5.1) | 71 (4.6) | 143 (4.9) | 19 (4.2) | 29 (5.9) | 48 (5.1) | 53 (5.6) | 42 (4.0) | 95 (4.7) |
| Years of Education, years (SD) | 10.3 (4.0) | 9.1 (4.8) | 9.7 (4.5) | 10.2 (4.0) | 9.2 (4.7) | 9.7 (4.4) | 10.3 (4.0) | 9.1 (4.9) | 9.7 (4.5) |
| Physical Activity Level, % | |||||||||
| Sedentary | 1119 (79.8) | 1383 (89.4) | 2502 (84.8) | 368 (81.4) | 439 (88.7) | 807 (85.2) | 751 (79.0) | 944 (89.7) | 1695 (84.6) |
| Moderate | 244 (17.4) | 151 (9.8) | 395 (13.4) | 70 (15.5) | 52 (10.5) | 122 (12.9) | 174 (18.3) | 99 (9.4) | 273 (13.6) |
| Vigorous | 40 (2.8) | 13 (0.8) | 53 (1.8) | 14 (3.1) | 4 (0.8) | 18 (1.9) | 26 (2.7) | 9 (0.9) | 35 (1.8) |
| Intentional Physical Exercise, % | 353 (25.3) | 388 (25.1) | 741 (25.2) | 127 (28.0) | 127 (25.7) | 254 (26.8) | 226 (23.9) | 261 (24.8) | 487 (24.4) |
| Smoking Status, % | |||||||||
| Never smoked | 571 (40.7) | 1527 (98.6) | 2098 (71.1) | 185 (41.0) | 487 (98.4) | 672 (71.0) | 386 (40.6) | 1040 (98.7) | 1426 (71.2) |
| Former smoker | 147 (10.5) | 6 (0.4) | 153 (5.2) | 61 (13.5) | 3 (0.6) | 64 (6.8) | 86 (9.1) | 3 (0.3) | 89 (4.4) |
| Current smoker | 684 (48.8) | 15 (1.0) | 699 (23.7) | 206 (45.5) | 5 (1.0) | 211 (22.2) | 478 (50.3) | 10 (1.0) | 488 (24.4) |
| Alcohol use, % | |||||||||
| Lifetime abstainers | 828 (64.2) | 1408 (94.9) | 2236 (80.6) | 274 (66.8) | 449 (95.6) | 723 (82.1) | 554 (63.1) | 959 (94.6) | 1513 (80.0) |
| Nonheavy drinkers | 353 (27.4) | 68 (4.5) | 421 (15.2) | 111 (26.9) | 18 (3.8) | 129 (14.6) | 242 (27.6) | 50 (4.9) | 292 (15.4) |
| Infrequent heavy drinkers | 29 (2.3) | 4 (0.3) | 33 (1.2) | 5 (1.2) | 2 (0.4) | 7 (0.8) | 24 (2.7) | 2 (0.2) | 26 (1.4) |
| Frequent heavy drinkers | 79 (6.1) | 4 (0.3) | 83 (3.0) | 21 (5.1) | 1 (0.2) | 22 (2.5) | 58 (6.6) | 3 (0.3) | 61 (3.2) |
| Dietary Intake | |||||||||
| Energy, kcal/day (SD) | 1945.8 (918.4) | 1608.9 (776.4) | 1769.1 (863.3) | 1882.5 (746.8) | 1580.9 (649.4) | 1724.9 (713.3) | 1975.8 (988.6) | 1622.1 (829.4) | 1790.1 (925.2) |
| Total iron, mg/day (SD) | 22.5 (21.5) | 17.8 (11.4) | 20.0 (17.1) | 21.9 (19.2) | 17.8 (10.2) | 19.8 (15.3) | 22.7 (22.5) | 17.8 (11.9) | 20.0 (17.9) |
| Glucose Metabolism Index | |||||||||
| Elevated fasting glucose, % | 351 (25.0) | 326 (21.1) | 677 (22.9) | 93 (20.5) | 96 (19.4) | 189 (19.9) | 258 (27.0) | 230 (21.8) | 488 (24.3) |
| Fasting Glucose, mmol/L | 5.2 (1.2) | 5.1 (1.1) | 5.2 (1.1) | 5.1 (1.1) | 5.1 (1.0) | 5.1 (1.1) | 5.2 (1.1) | 5.2 (1.1) | 5.2 (1.1) |
| HbA1c, % | 5.7 (1.0) | 5.7 (0.9) | 5.7 (1.0) | 5.6 (1.0) | 5.6 (0.8) | 5.6 (0.9) | 5.8 (1.0) | 5.8 (0.9) | 5.8 (1.0) |
| HOMA2-IR | 0.6 (0.5) | 0.7 (0.6) | 0.7 (0.6) | 0.6 (0.5) | 0.8 (0.7) | 0.7 (0.6) | 0.7 (0.5) | 0.7 (0.6) | 0.7 (0.5) |
| Frequency (%) | |||
|---|---|---|---|
| All (n = 2951) | Male (n = 1403) | Female (n = 1548) | |
| Genotype | |||
| GG | 17.6 | 18.4 | 16.9 |
| GC | 50.3 | 49.4 | 51.1 |
| CC | 32.1 | 32.2 | 32.0 |
| MAF | |||
| G | 42.8 | 43.1 | 42.5 |
| Quartiles of Dietary Iron Intake (mg/day), ORs (95% CI) 2 | |||||||
|---|---|---|---|---|---|---|---|
| Q1 | Q2 | Q3 | Q4 | p-Value for Trend 3 | |||
| Total Iron Intake (mg/day) | <12.82 | 12.82–16.59 | 16.59–22.07 | ≥22.07 | |||
| n | 736 | 739 | 740 | 736 | |||
| Elevated fasting glucose | |||||||
| All | Model 1 | G allele non-carriers | Reference | 1.03 (0.62, 1.70) | 1.53 (0.95, 2.45) | 1.76 (1.10, 2.82) | 0.005 |
| G allele carriers | 1.44 (0.95, 2.18) | 1.88 (1.39, 2.54) | 1.75 (1.29, 2.38) | 1.71 (1.26, 2.32) | 0.375 | ||
| Model 2 | G allele non-carriers | Reference | 1.01 (0.58, 1.75) | 1.43 (0.82, 2.49) | 1.67 (0.87, 3.18) | 0.066 | |
| G allele carriers | 1.58 (1.01, 2.47) | 2.25 (1.63, 3.10) | 2.24 (1.59, 3.15) | 2.52 (1.69, 3.76) | 0·033 | ||
| Male | Model 1 | G allele non-carriers | Reference | 0.78 (0.35, 1.72) | 1.46 (0.71, 3.00) | 1.61 (0.79, 3.25) | 0·046 |
| G allele carriers | 1.75 (0.89, 3.46) | 2.01 (1.25, 3.23) | 1.68 (1.07, 2.65) | 2.04 (1.32, 3.16) | 0.667 | ||
| Model 2 | G allele non-carriers | Reference | 0.91 (0.37, 2.23) | 1.35 (0.55, 3.30) | 2.48 (0.90, 6.85) | 0.034 | |
| G allele carriers | 2.03 (0.93, 4.44) | 2.67 (1.59, 4.47) | 2.34 (1.39, 3.95) | 3.22 (1.82, 5.70) | 0.180 | ||
| Female | Model 1 | G allele non-carriers | Reference | 1.27 (0.66, 2.43) | 1.53 (0.81, 2.88) | 1.84 (0.97, 3.51) | 0.051 |
| G allele carriers | 1.27 (0.75, 2.14) | 1.80 (1.22, 2.67) | 1.96 (1.30, 2.95) | 1.37 (0.88, 2.15) | 0.410 | ||
| Model 2 | G allele non-carriers | Reference | 1.12 (0.53, 2.37) | 1.53 (0.72, 3.25) | 1.19 (0.48, 2.97) | 0.512 | |
| G allele carriers | 1.44 (0.82, 2.53) | 2.12 (1.39, 3.23) | 2.55 (1.61, 4.04) | 1.99 (1.10, 3.61) | 0.095 | ||
| Quartiles of Dietary Iron Intake (mg/day), βs (95% CI) 2 | |||||||
|---|---|---|---|---|---|---|---|
| Q1 | Q2 | Q3 | Q4 | p-Value for Trend 3 | |||
| Total Iron Intake (mg/day) | <12.82 | 12.82–16.59 | 16.59–22.07 | ≥22.07 | |||
| n 4 | 691 | 707 | 691 | 692 | |||
| Fasting glucose | |||||||
| All | Model 1 | G allele non-carriers | Reference | 0.08 (−0.12, 0.29) | 0.13 (−0.07, 0.34) | 0.28 (0.08, 0.49) | 0.007 |
| G allele carriers | 0.16 (0.00, 0.33) | 0.25 (0.11, 0.40) | 0.24 (0.09, 0.38) | 0.27 (0.12, 0.42) | 0.185 | ||
| Model 2 | G allele non-carriers | Reference | 0.02 (−0.20, 0.25) | 0.02 (−0.21, 0.26) | 0.17 (−0.11, 0.45) | 0.288 | |
| G allele carriers | 0.18 (0.01, 0.35) | 0.32 (0.16, 0.48) | 0.31 (0.14, 0.47) | 0.40 (0.21, 0.60) | 0.040 | ||
| Male | Model 1 | G allele non-carriers | Reference | −0.08 (−0.44, 0.27) | 0.07 (−0.27, 0.41) | 0.26 (−0.07, 0.60) | 0.043 |
| G allele carriers | 0.20 (−0.13, 0.52) | 0.20 (−0.05, 0.45) | 0.19 (−0.05, 0.43) | 0.24 (0.00, 0.48) | 0.708 | ||
| Model 2 | G allele non-carriers | Reference | −0.07 (−0.46, 0.31) | −0.01 (−0.42, 0.39) | 0.34 (−0.13, 0.81) | 0.121 | |
| G allele carriers | 0.24 (−0.12, 0.60) | 0.31 (0.03, 0.58) | 0.29 (0.02, 0.56) | 0.37 (0.07, 0.67) | 0.442 | ||
| Female | Model 1 | G allele non-carriers | Reference | 0.18 (−0.07, 0.42) | 0.16 (−0.09, 0.40) | 0.25 (−0.01, 0.50) | 0.070 |
| G allele carriers | 0.15 (−0.03, 0.33) | 0.30 (0.11, 0.46) | 0.27 (0.08, 0.47) | 0.30 (0.10, 0.50) | 0.138 | ||
| Model 2 | G allele non-carriers | Reference | 0.07 (−0.20, 0.33) | 0.01 (−0.28, 0.30) | −0.01 (−0.36, 0.34) | 0.908 | |
| G allele carriers | 0.12 (−0.05, 0.29) | 0.32 (0.13, 0.51) | 0.32 (0.10, 0.53) | 0.43 (0.17, 0.70) | 0.021 | ||
| Quartiles of Dietary Iron Intake (mg/day), βs (95% CI) 2 | |||||||
|---|---|---|---|---|---|---|---|
| Q1 | Q2 | Q3 | Q4 | p-Value for Trend 3 | |||
| Total Iron Intake (mg/day) | <12.82 | 12.82–16.59 | 16.59–22.07 | ≥22.07 | |||
| n 4 | 691 | 707 | 691 | 692 | |||
| HbA1c | |||||||
| All | Model 1 | G allele non-carriers | Reference | 0.09 (−0.09, 0.27) | 0.04 (−0.13, 0.22) | 0.13 (−0.05, 0.30) | 0.259 |
| G allele carriers | 0.07 (−0.06, 0.20) | 0.24 (0.12, 0.37) | 0.27 (0.14, 0.39) | 0.22 (0.10, 0.34) | 0.019 | ||
| Model 2 | G allele non-carriers | Reference | 0.06 (−0.13, 0.24) | −0.05 (−0.24, 0.15) | 0.03 (−0.21, 0.26) | 0.890 | |
| G allele carriers | 0.10 (−0.03, 0.23) | 0.31 (0.18, 0.44) | 0.34 (0.20, 0.47) | 0.34 (0.17, 0.50) | 0.003 | ||
| Male | Model 1 | G allele non-carriers | Reference | 0.09 (−0.21, 0.40) | 0.06 (−0.24, 0.37) | 0.07 (−0.23, 0.36) | 0.802 |
| G allele carriers | 0.13 (−0.13, 0.39) | 0.18 (−0.03, 0.40) | 0.23 (0.02, 0.44) | 0.22 (0.02, 0.43) | 0.336 | ||
| Model 2 | G allele non-carriers | Reference | 0.09 (−0.25, 0.40) | −0.10 (−0.44, 0.24) | 0.00 (−0.40, 0.40) | 0.693 | |
| G allele carriers | 0.19 (−0.08, 0.46) | 0.28 (0.04, 0.51) | 0.34 (0.11, 0.57) | 0.39 (0.14, 0.65) | 0.101 | ||
| Female | Model 1 | G allele non-carriers | Reference | 0.07 (−0.14, 0.27) | 0.01 (−0.19, 0.22) | 0.19 (−0.02, 0.41) | 0.157 |
| G allele carriers | 0.04 (−0.10, 0.18) | 0.28 (0.14, 0.43) | 0.30 (0.14, 0.46) | 0.20 (0.04, 0.36) | 0.018 | ||
| Model 2 | G allele non-carriers | Reference | 0.05 (−0.17, 0.27) | 0.00 (−0.24, 0.24) | 0.07 (−0.22, 0.36) | 0.764 | |
| G allele carriers | 0.07 (−0.07, 0.21) | 0.16 (−0.08, 0.39) | 0.22 (−0.01, 0.45) | 0.27 (0.02, 0.53) | 0.014 | ||
| Quartiles of Dietary Iron Intake (mg/day), βs (95% CI) 2 | |||||||
|---|---|---|---|---|---|---|---|
| Q1 | Q2 | Q3 | Q4 | p-Value for Trend 3 | |||
| Total Iron Intake (mg/day) | <12.82 | (12.82–16.59) | (16.59–22.07) | ≥22.07 | |||
| n 4 | 659 | 675 | 676 | 670 | |||
| HOMA2-IR | |||||||
| All | Model 1 | G allele non-carriers | Reference | −0.08 (−0.20, 0.04) | −0.03 (−0.15, 0.09) | −0.10 (−0.22, 0.02) | 0.208 |
| G allele carriers | −0.09 (−0.19, 0.01) | −0.06 (−0.13, 0.01) | −0.04 (−0.11, 0.03) | −0.02 (−0.10, 0.05) | 0.057 | ||
| Model 2 | G allele non-carriers | Reference | −0.12 (−0.25, 0.01) | −0.09 (−0.23, 0.04) | −0.17 (−0.33, 0.00) | 0.090 | |
| G allele carriers | −0.12 (−0.22, −0.01) | −0.10 (−0.17, −0.02) | −0.08 (−0.16, 0.00) | −0.07 (−0.17, 0.02) | 0.268 | ||
| Male | Model 1 | G allele non-carriers | Reference | −0.14 (−0.28, 0.01) | −0.04 (−0.18, 0.10) | −0.08 (−0.22, 0.06) | 0.655 |
| G allele carriers | −0.08 (−0.23, 0.06) | −0.07 (−0.18, 0.04) | −0.03 (−0.14, 0.07) | −0.02 (−0.12, 0.08) | 0.168 | ||
| Model 2 | G allele non-carriers | Reference | −0.17 (−0.32, −0.01) | −0.07 (−0.24, 0.09) | −0.10 (−0.30, 0.09) | 0.656 | |
| G allele carriers | −0.12 (−0.28, 0.03) | −0.09 (−0.20, 0.03) | −0.07 (−0.19, 0.04) | −0.06 (−0.19, 0.06) | 0.404 | ||
| Female | Model 1 | G allele non-carriers | Reference | −0.04 (−0.22, 0.15) | −0.03 (−0.21, 0.15) | −0.13 (−0.32, 0.06) | 0.233 |
| G allele carriers | −0.10 (−0.23, 0.03) | −0.06 (−0.16, 0.03) | −0.05 (−0.15, 0.05) | −0.04 (−0.14, 0.07) | 0.192 | ||
| Model 2 | G allele non-carriers | Reference | −0.10 (−0.30, 0.10) | −0.12 (−0.34, 0.10) | −0.23 (−0.49, 0.04) | 0.105 | |
| G allele carriers | −0.13 (−0.27, 0.01) | −0.12 (−0.22, −0.02) | −0.09 (−0.20, 0.02) | −0.09 (−0.22, 0.05) | 0.434 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, L.; Wang, Z.; Zang, J.; Liu, H.; Lu, Y.; He, X.; Wu, C.; Su, J.; Zhu, Z. The Association between Dietary Iron Intake, SNP of the MTNR1B rs10830963, and Glucose Metabolism in Chinese Population. Nutrients 2023, 15, 1986. https://doi.org/10.3390/nu15081986
Shen L, Wang Z, Zang J, Liu H, Lu Y, He X, Wu C, Su J, Zhu Z. The Association between Dietary Iron Intake, SNP of the MTNR1B rs10830963, and Glucose Metabolism in Chinese Population. Nutrients. 2023; 15(8):1986. https://doi.org/10.3390/nu15081986
Chicago/Turabian StyleShen, Liping, Zhengyuan Wang, Jiajie Zang, Hong Liu, Ye Lu, Xin He, Chunfeng Wu, Jin Su, and Zhenni Zhu. 2023. "The Association between Dietary Iron Intake, SNP of the MTNR1B rs10830963, and Glucose Metabolism in Chinese Population" Nutrients 15, no. 8: 1986. https://doi.org/10.3390/nu15081986
APA StyleShen, L., Wang, Z., Zang, J., Liu, H., Lu, Y., He, X., Wu, C., Su, J., & Zhu, Z. (2023). The Association between Dietary Iron Intake, SNP of the MTNR1B rs10830963, and Glucose Metabolism in Chinese Population. Nutrients, 15(8), 1986. https://doi.org/10.3390/nu15081986