
Use of a Silkworm (Bombyx mori) Larvae By-Product for the Treatment of Atopic Dermatitis: Inhibition of NF-κB Nuclear Translocation and MAPK Signaling

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Powdered by-Products of Mature Silkworm Larvae
2.3. Treatment of HaCaT Cells with Powdered Silkworm Larval by-Products
2.4. Reverse Transcription–Quantitative Polymerase Chain Reaction
2.5. Cell Viability
2.6. Immunofluorescence Analysis
2.7. Western Blotting
2.8. Stimulation of AD-like Skin Lesion and an Experimental Animal Model
2.9. RT-qPCR of RNA Extracted from DNCB-Induced AD Mice
2.10. Histological Observation of DNCB-Treated AD Mice
2.11. Serum Immunoglobulin Analysis of DNCB-Induced AD Mice
2.12. Amino Acid Analysis
2.13. Statistical Analysis
3. Results
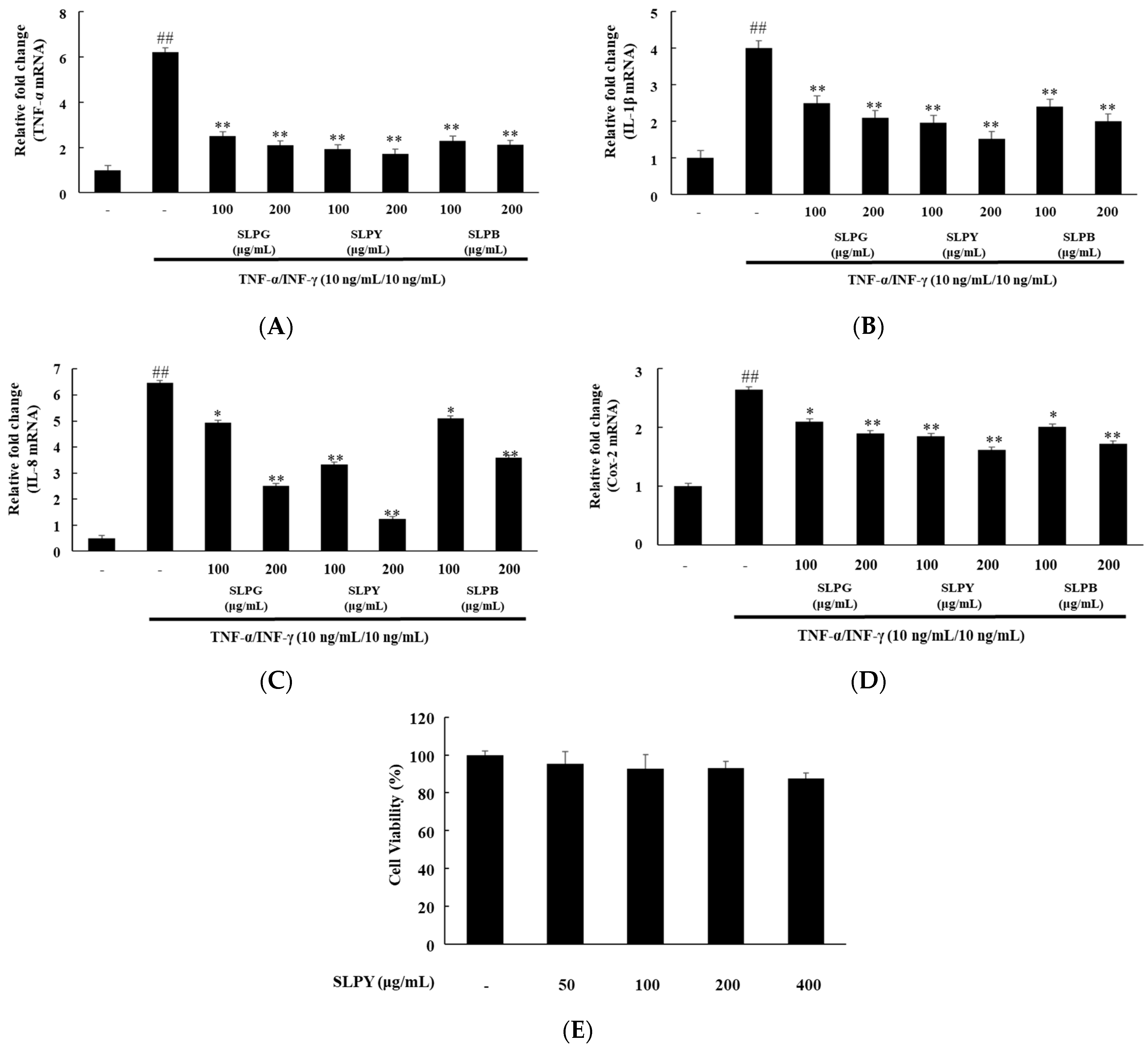
3.1. Powdered by-Products of Silkworm Larvae Exhibit Anti-Inflammatory Effects on TNF-α/IFN-γ-Treated HaCaT Cells
3.2. Percentage of Cell Viability after SLPY Treatment
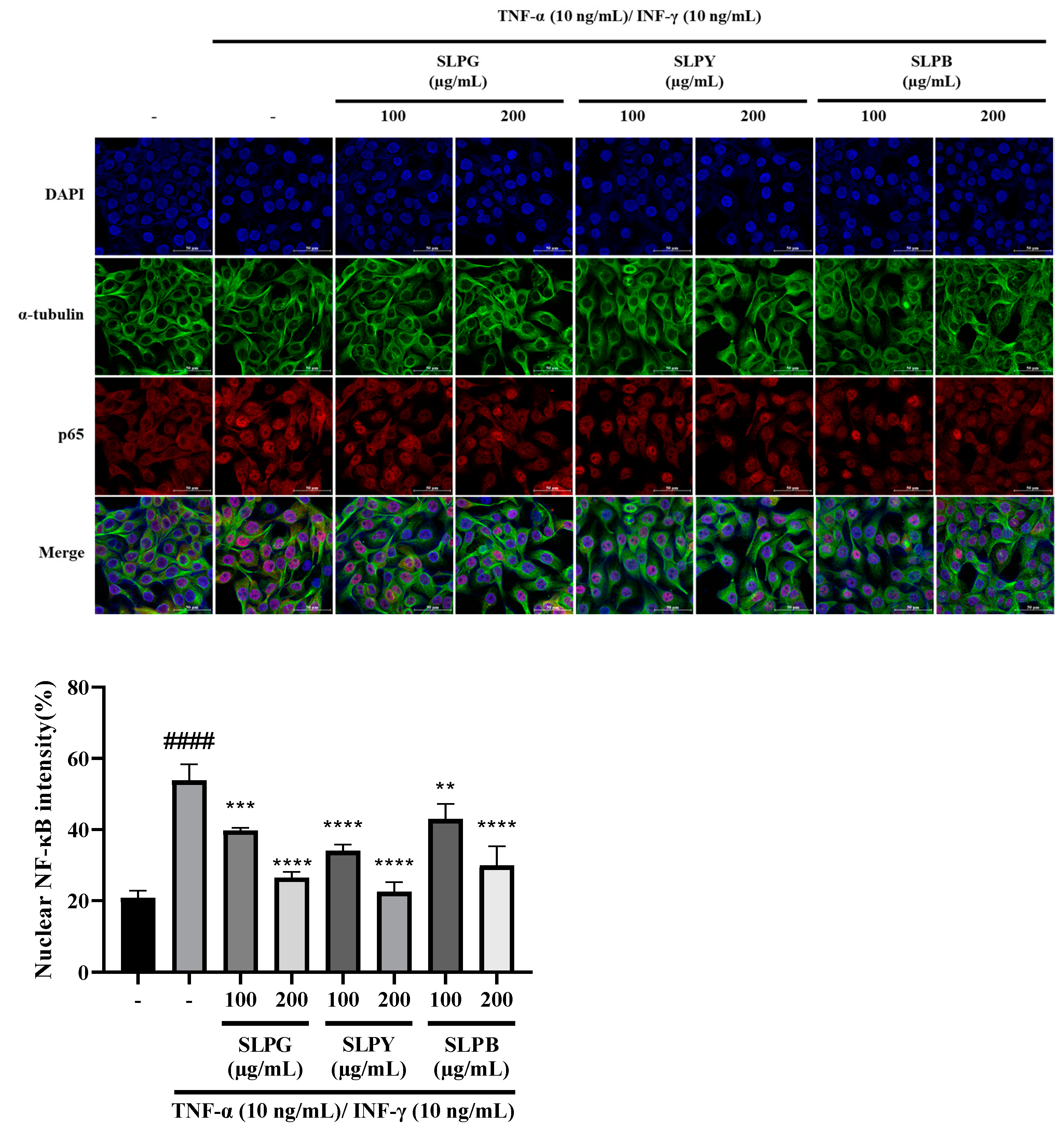
3.3. Powdered by-Products of Silkworm Larvae Attenuate NF-kB p65 Nuclear Translocation in Stimulated HaCaT Cells
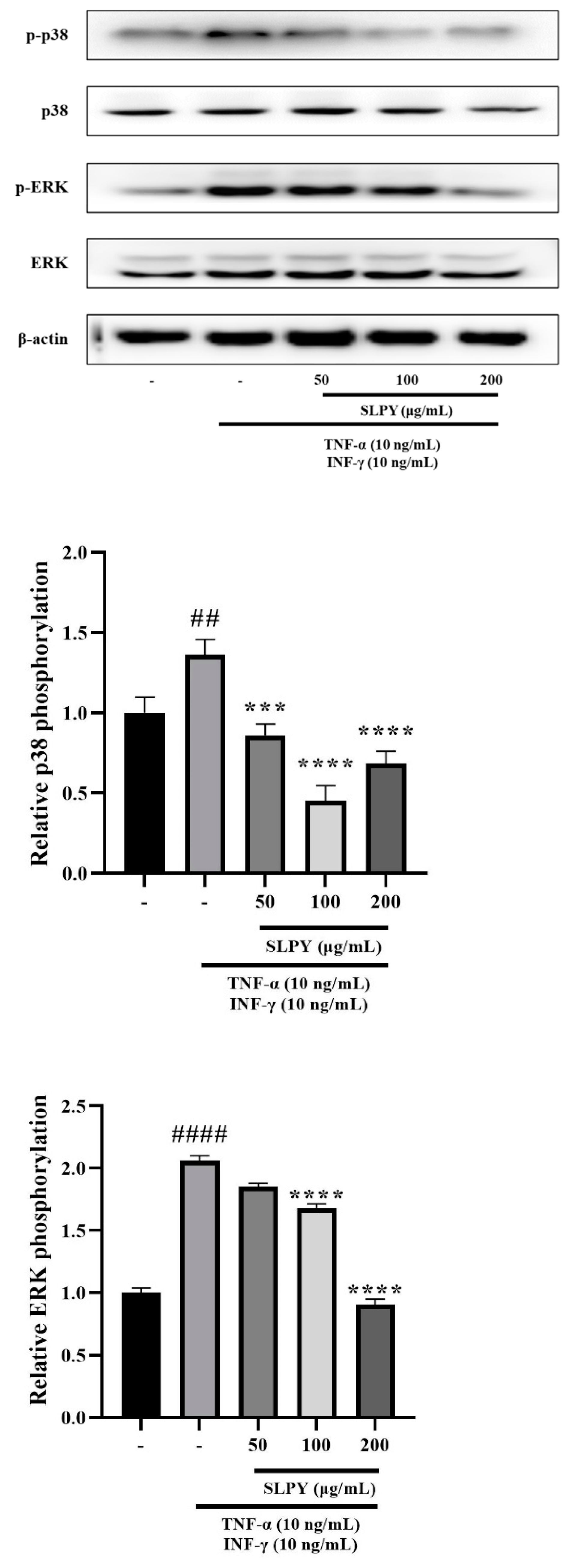
3.4. SLPY Downregulates Expression of Inflammation-Related Proteins
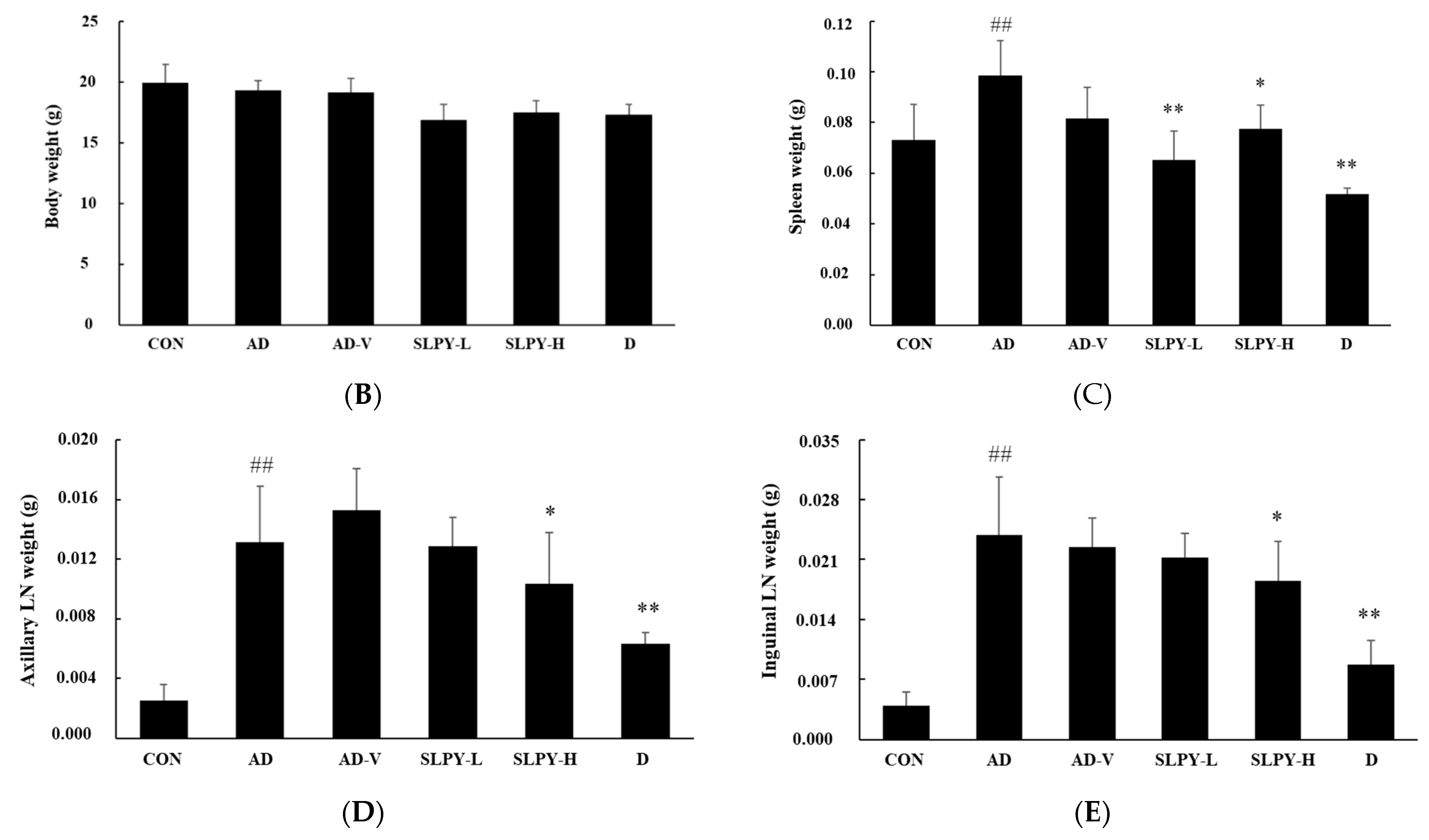
3.5. SLPY Ameliorates AD-like Skin Lesions in DNCB-Treated BALB/c Mice
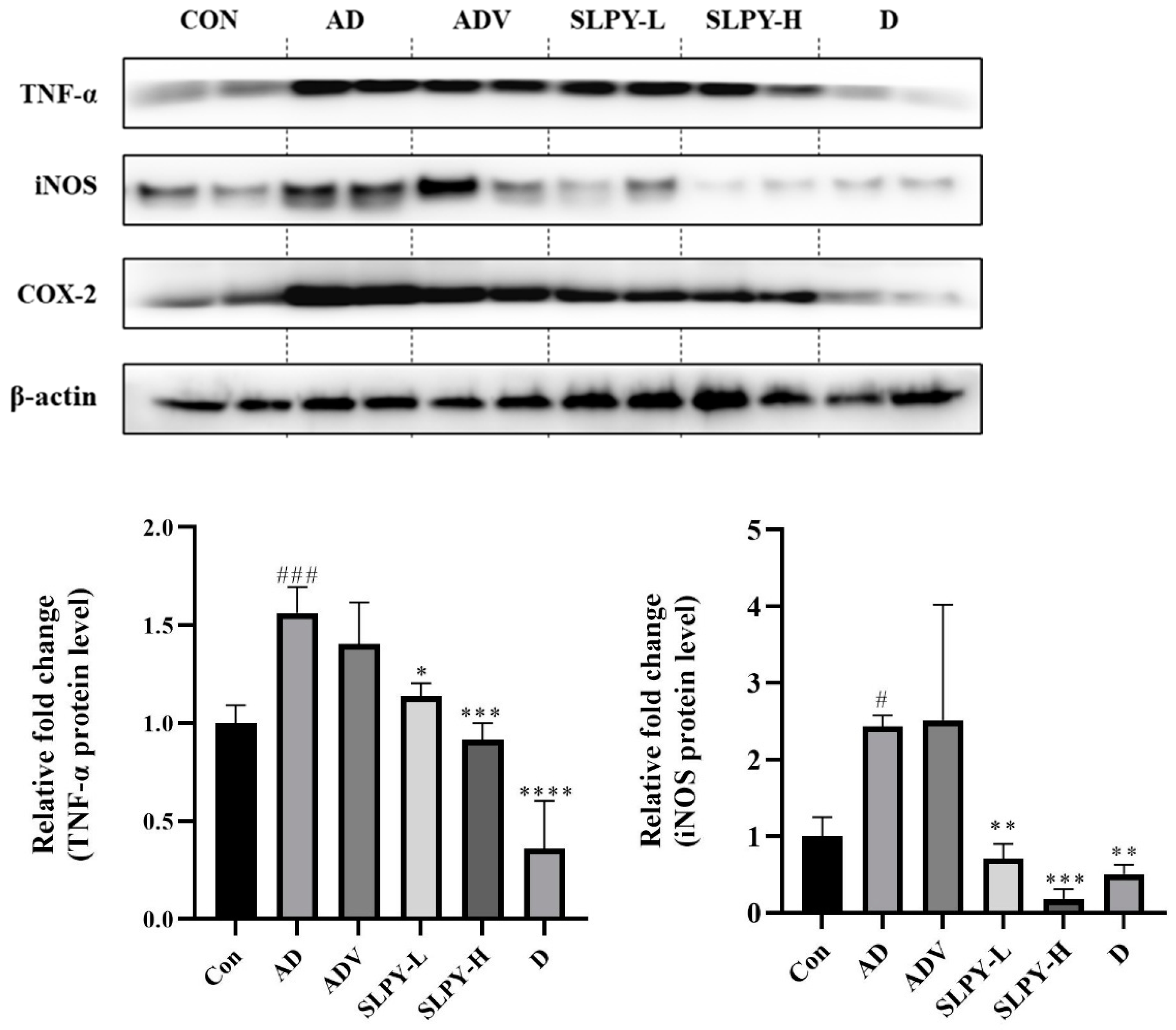
3.6. Effect of SLPY on Inflammatory-Related Protein Expression in Skin Tissues of DNCB-Treated Mice
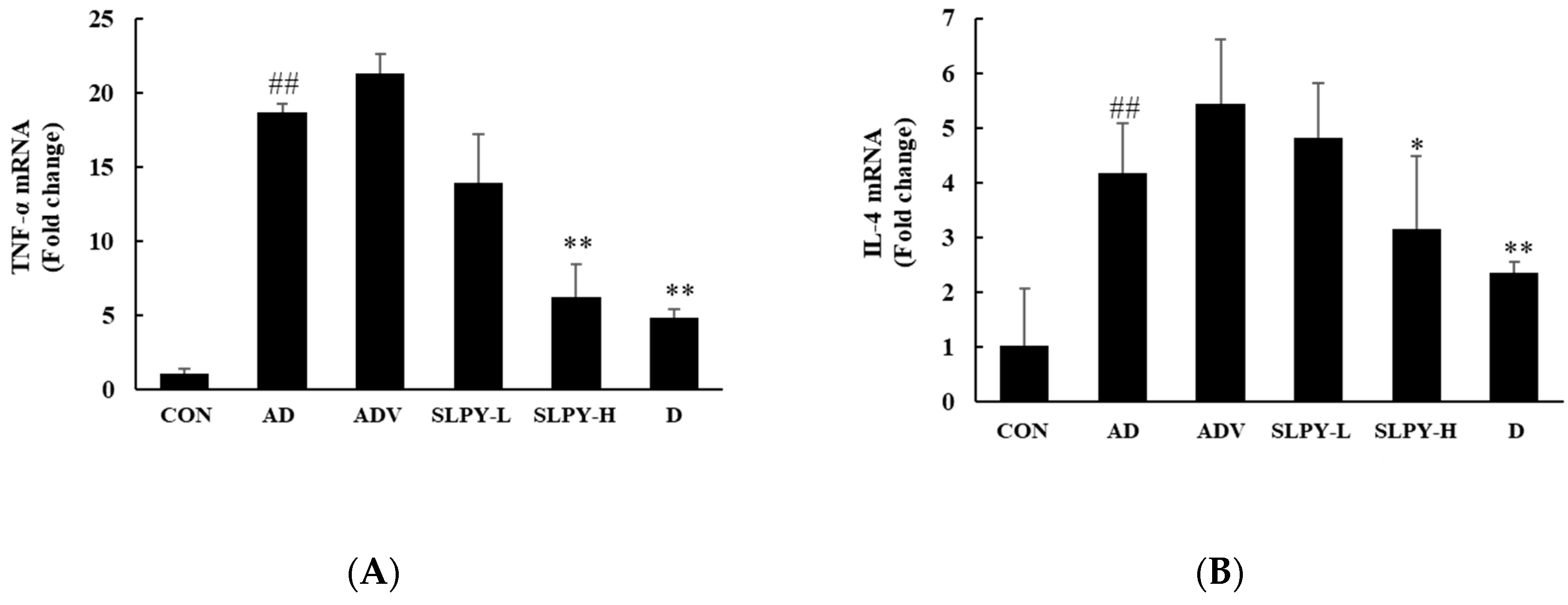
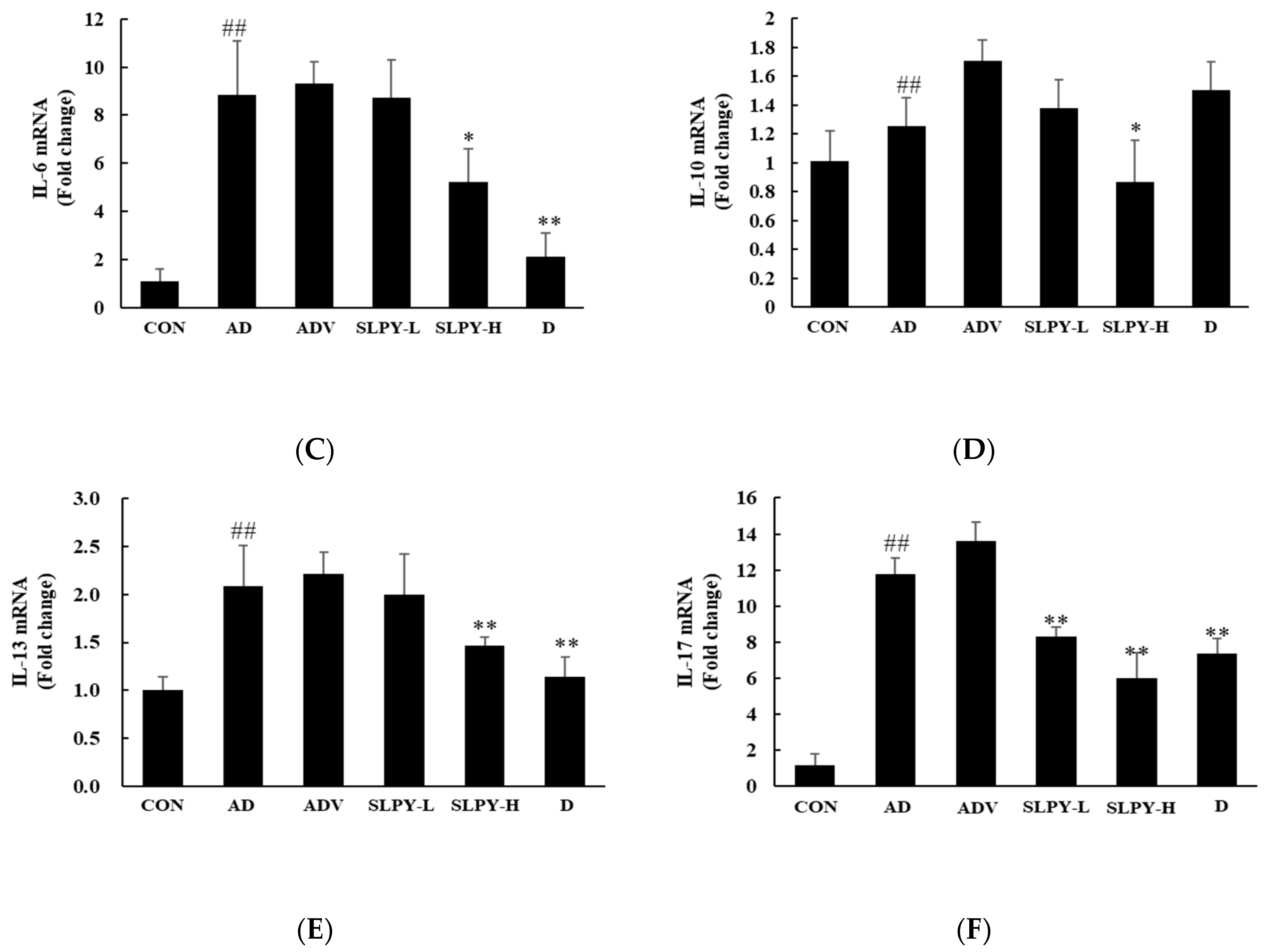
3.7. SLPY Inhibits the Expression Levels of Inflammatory Cytokines in DNCB-Induced Mice
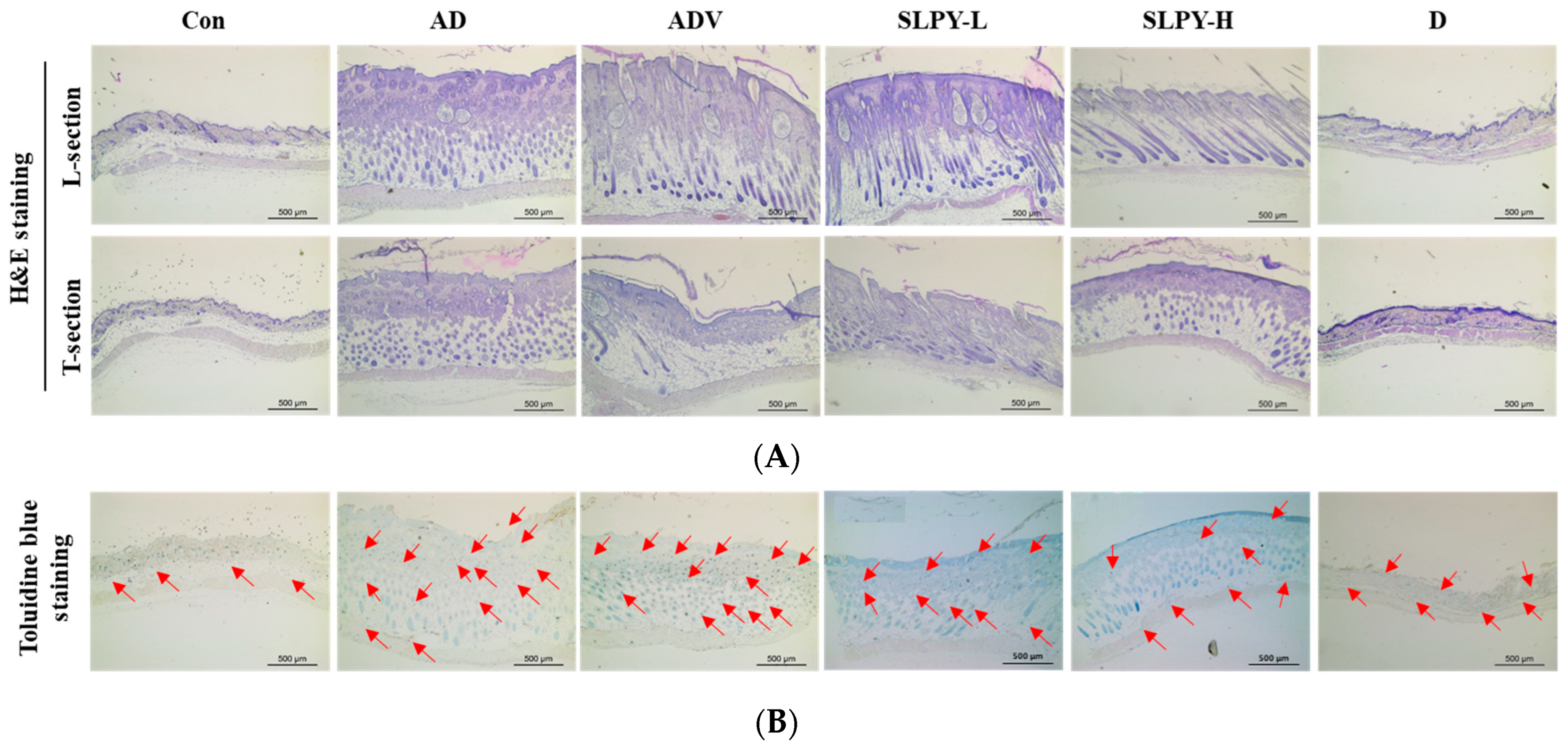
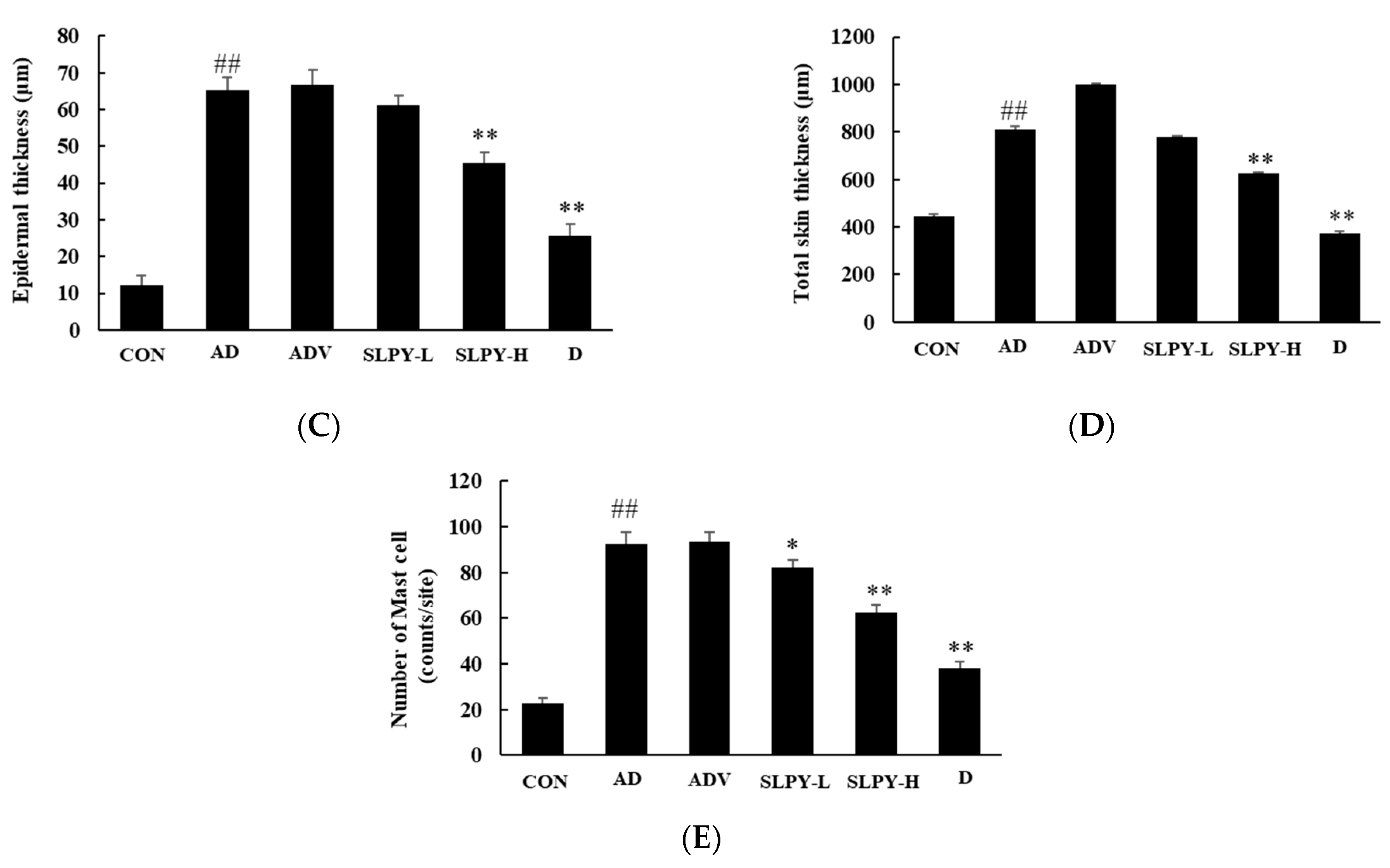
3.8. SLPY Reduces Pathological Damage in Skin Tissues of DNCB-Treated Mice
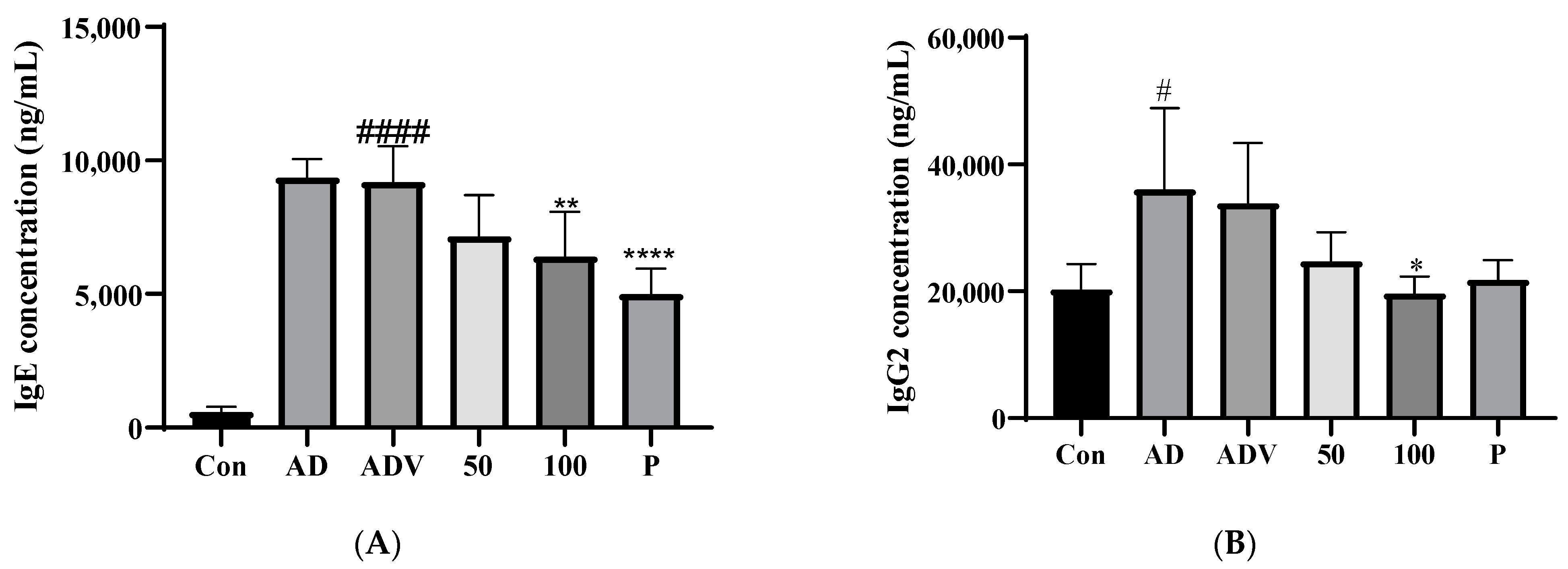
3.9. SLPY Application Reduced IgE and IgG2a Levels in DNCB-Treated AD Mice
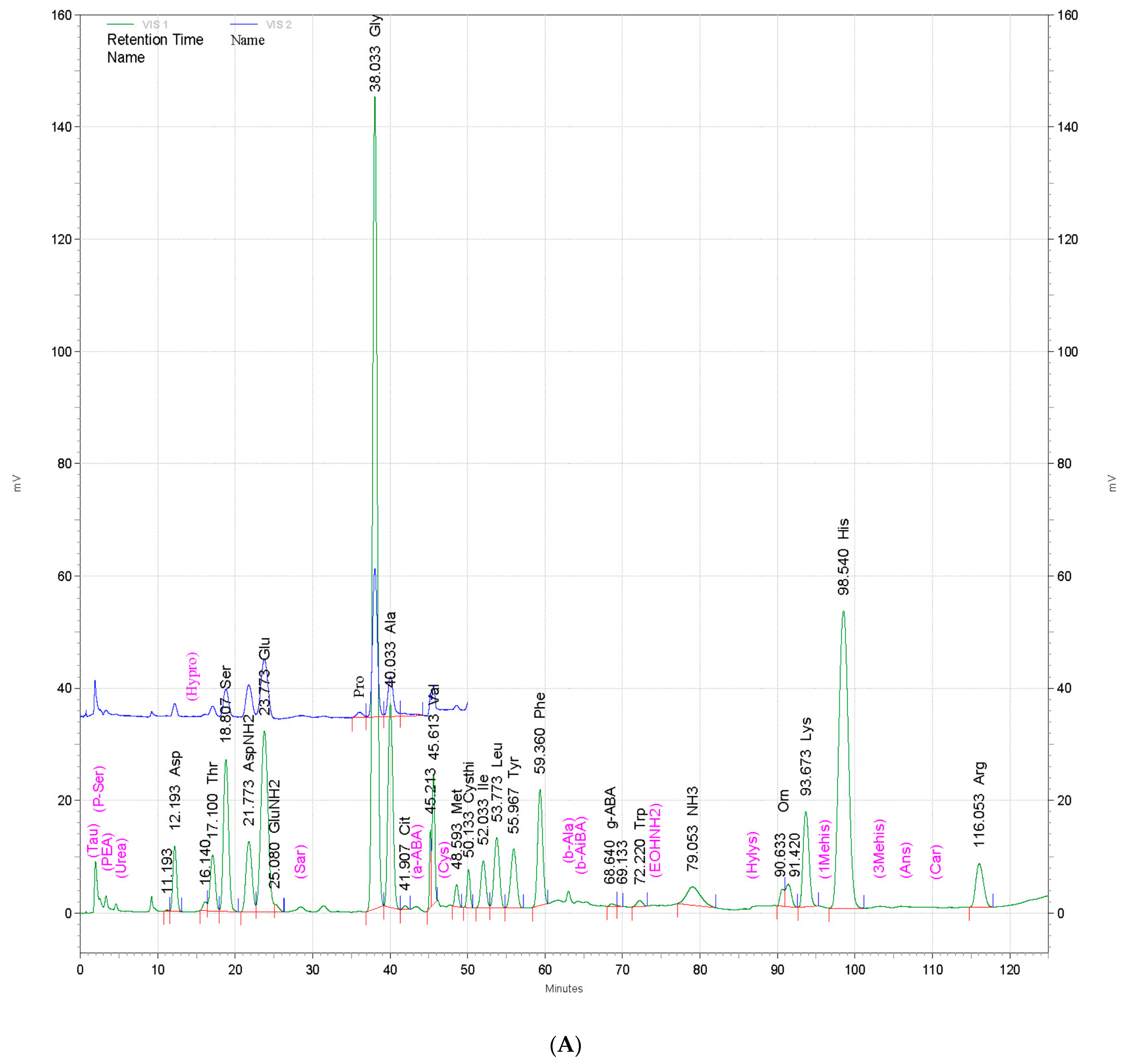
3.10. Amino Acid Composition of Powdered By-Products of Silkworm Larvae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oykhman, P.; Dookie, J.; Al-Rammahy, H.; de Benedetto, A.; Asiniwasis, R.N.; LeBovidge, J.; Wang, J.; Ong, P.Y.; Lio, P.; Gutierrez, A.; et al. Dietary Elimination for the Treatment of Atopic Dermatitis: A Systematic Review and Meta-Analysis. J. Allergy Clin. Immunol. Pract. 2022, 10, 2657–2666.e8. [Google Scholar] [CrossRef]
- Kim, Y.J.; Choi, M.J.; Bak, D.H.; Lee, B.C.; Ko, E.J.; Ahn, G.R.; Ahn, S.W.; Kim, M.J.; Na, J.; Kim, B.J. Topical Administration of EGF Suppresses Immune Response and Protects Skin Barrier in DNCB-Induced Atopic Dermatitis in NC/Nga Mice. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef]
- Hadi, H.A.; Tarmizi, A.I.; Khalid, K.A.; Gajdács, M.; Aslam, A.; Jamshed, S. The Epidemiology and Global Burden of Atopic Dermatitis: A Narrative Review. Life 2021, 11, 936. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Gao, X.; Wang, X.; Li, X.; Jiang, X.; Xie, Z.; Ma, K.; Ma, J.; Umezawa, K.; Zhang, Y. Comparison of Anti-Atopic Dermatitis Activities between DHMEQ and Tacrolimus Ointments in Mouse Model without Stratum Corneum. Int. Immunopharmacol. 2019, 71, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Nakajima-Adachi, H.; Wang, Y.; Zhou, Y.; Gu, W.; Hiraide, E.; Morinaga, M.; Nakagawa, R.; Nakamura, S.; Takano, T.; et al. Regulation of Th2 Responses by Lactococcus Lactis Subsp. Cremoris YRC3780 Alleviates DNCB-Induced Atopic Dermatitis in the Mouse Model. J. Funct. Foods 2022, 94, 105091. [Google Scholar] [CrossRef]
- Fujikawa, M.; Sugimoto, H.; Tamura, R.; Fujikawa, K.; Yamagishi, A.; Ueda, Y. Effects of Mucopolysaccharide Polysulphate on Tight Junction Barrier in Human Epidermal Keratinocytes. Exp. Dermatol. 2022, 31, 1676–1684. [Google Scholar] [CrossRef] [PubMed]
- Möckel, M.; De La Cruz, N.C.; Rübsam, M.; Wirtz, L.; Tantcheva-Poor, I.; Malter, W.; Zinser, M.; Bieber, T.; Knebel-Mörsdorf, D. Herpes Simplex Virus 1 Can Bypass Impaired Epidermal Barriers upon Ex Vivo Infection of Skin from Atopic Dermatitis Patients. J. Virol. 2022, 96, e00864-22. [Google Scholar] [CrossRef] [PubMed]
- Belmesk, L.; Muntyanu, A.; Cantin, E.; AlHalees, Z.; Jack, C.S.; Le, M.; Sasseville, D.; Iannattone, L.; Ben-Shoshan, M.; Litvinov, I.V.; et al. Prominent Role of Type 2 Immunity in Skin Diseases: Beyond Atopic Dermatitis. J. Cutan. Med. Surg. 2022, 26, 33–49. [Google Scholar] [CrossRef]
- Kader, H.A.; Azeem, M.; Jwayed, S.A.; Al-Shehhi, A.; Tabassum, A.; Ayoub, M.A.; Hetta, H.F.; Waheed, Y.; Iratni, R.; Al-Dhaheri, A.; et al. Current Insights into Immunology and Novel Therapeutics of Atopic Dermatitis. Cells 2021, 10, 1392. [Google Scholar] [CrossRef]
- Pan, J.; Xu, H.; Cheng, Y.; Mintah, B.K.; Dabbour, M.; Yang, F.; Chen, W.; Zhang, Z.; Dai, C.; He, R.; et al. Recent Insight on Edible Insect Protein: Extraction, Functional Properties, Allergenicity, Bioactivity, and Applications. Foods 2022, 11, 2931. [Google Scholar] [CrossRef]
- Ji, S.D.; Nguyen, P.; Yoon, S.M.; Kim, K.Y.; Son, J.G.; Kweon, H.Y.; Koh, Y.H. Comparison of Nutrient Compositions and Pharmacological Effects of Steamed and Freeze-Dried Mature Silkworm Powders Generated by Four Silkworm Varieties. J. Asia. Pac. Entomol. 2017, 20, 1410–1418. [Google Scholar] [CrossRef]
- Ji, S.D.; Kim, N.S.; Kweon, H.Y.; Choi, B.H.; Yoon, S.M.; Kim, K.Y.; Koh, Y.H. Nutrient Compositions of Bombyx Mori Mature Silkworm Larval Powders Suggest Their Possible Health Improvement Effects in Humans. J. Asia. Pac. Entomol. 2016, 19, 1027–1033. [Google Scholar] [CrossRef]
- Lee, D.-Y.; Cho, J.-M.; Yun, S.-M.; Hong, K.-S.; Ji, S.-D.; Son, J.-G.; Kim, E.-H. Comparative Effect of Silkworm Powder from 3 Bombyx Mori Varieties on Ethanol-Induced Gastric Injury in Rat Model. Int. J. Ind. Entomol. 2017, 35, 14–21. [Google Scholar]
- Ji, S.D.; Kim, S.B.; Kim, K.Y.; Kim, N.S.; Kim, S.W.; Jo, Y.Y.; Kim, J.G.; Kim, Y.K.; Seok, Y.S.; Lim, J.R.; et al. Contents of Nutrients in Ultra-Fine Powders of Steamed and Lyophilized Mature Silkworms Generated by Four Silkworm Varieties. J. Asia. Pac. Entomol. 2019, 22, 969–974. [Google Scholar] [CrossRef]
- Hong, K.S.; Yun, S.M.; Cho, J.M.; Lee, D.Y.; Ji, S.D.; Son, J.G.; Kim, E.H. Silkworm (Bombyx Mori) Powder Supplementation Alleviates Alcoholic Fatty Liver Disease in Rats. J. Funct. Foods 2018, 43, 29–36. [Google Scholar] [CrossRef]
- Lee, D.Y.; Hong, K.S.; Song, M.Y.; Yun, S.M.; Ji, S.D.; Son, J.G.; Kim, E.H. Hepatoprotective Effects of Steamed and Freeze-Dried Mature Silkworm Larval Powder against Ethanol-Induced Fatty Liver Disease in Rats. Foods 2020, 9, 285. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Yun, S.M.; Song, M.Y.; Ji, S.D.; Son, J.G.; Kim, E.H. Administration of Steamed and Freeze-Dried Mature Silkworm Larval Powder Prevents Hepatic Fibrosis and Hepatocellular Carcinogenesis by Blocking TGF-β/STAT3 Signaling Cascades in Rats. Cells 2020, 9, 568. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Kim, K.Y.; Ji, S.D.; Lee, H.T. Anti-Melanogenic Activity of Steamed and Freeze-Dried Mature Silkworm Powder. J. Asia. Pac. Entomol. 2017, 20, 1001–1006. [Google Scholar] [CrossRef]
- Nguyen, P.; Kim, K.Y.; Kim, A.Y.; Choi, B.H.; Osabutey, A.F.; Park, Y.H.; Lee, H.T.; Ji, S.D.; Koh, Y.H. Mature Silkworm Powders Ameliorated Scopolamine-Induced Amnesia by Enhancing Mitochondrial Functions in the Brains of Mice. J. Funct. Foods 2020, 67, 103886. [Google Scholar] [CrossRef]
- Kang, P.-D.; Sung, G.-B.; Kim, K.-Y.; Lee, H.-S.; Kim, M.-J.; Ji, S.-D.; Hong, I.-P.; Kweon, H.-Y.; Park, K.-Y.; Lee, K.-G.; et al. Breeding of New Silkworm Strain Yeonnokjam. Int. J. Ind. Entomol. 2011, 23, 175–178. [Google Scholar] [CrossRef]
- Lee, Y.; Choi, H.K.; N’deh, K.P.U.; Choi, Y.J.; Fan, M.; Kim, E.K.; Chung, K.H.; An, J.H. Inhibitory Effect of Centella Asiatica Extract on DNCB-Induced Atopic Dermatitis in HaCaT Cells and BALB/c Mice. Nutrients 2020, 12, 411. [Google Scholar] [CrossRef]
- Meiqi, F.; Erandi, W.N.; Young-Jin, C.; Qun, Z.; Mun, B.S.; Eun-Kyung, K. Tenebrio Molitor Larva Trypsin Hydrolysate Ameliorates Atopic. Nutrients 2023, 15, 93. [Google Scholar] [CrossRef]
- Guttman-Yassky, E.; Krueger, J.G.; Lebwohl, M.G. Systemic Immune Mechanisms in Atopic Dermatitis and Psoriasis with Implications for Treatment. Exp. Dermatol. 2018, 27, 409–417. [Google Scholar] [CrossRef]
- Feng, S.; Liu, W.; Deng, S.; Song, G.; Zhou, J.; Zheng, Z.; Song, Z. An Atopic Dermatitis-Like Mouse Model by Alternate Epicutaneous Application of Dinitrofluorobenzene and an Extract of Dermatophagoides Farinae. Front. Med. 2022, 9, 843230. [Google Scholar] [CrossRef]
- Yang, C.C.; Hung, Y.L.; Ko, W.C.; Tsai, Y.J.; Chang, J.F.; Liang, C.W.; Chang, D.C.; Hung, C.F. Effect of Neferine on Dncb-Induced Atopic Dermatitis in Hacat Cells and Balb/c Mice. Int. J. Mol. Sci. 2021, 22, 8237. [Google Scholar] [CrossRef]
- Min, G.Y.; Kim, E.Y.; Hong, S.; Kim, J.H.; Kim, M.; Kim, E.J.; Park, J.H.; Sohn, Y.; Jung, H.S. Lycopus Lucidus Turcz Ameliorates DNCB-Induced Atopic Dermatitis in BALB/c Mice. Mol. Med. Rep. 2021, 24, 827. [Google Scholar] [CrossRef]
- Wu, X.; He, K.; Velickovic, T.C.; Liu, Z. Nutritional, Functional, and Allergenic Properties of Silkworm Pupae. Food Sci. Nutr. 2021, 9, 4655–4665. [Google Scholar] [CrossRef] [PubMed]
- Zarate, M.A.; Nguyen, L.M.; De Dios, R.K.; Zheng, L.; Wright, C.J. Maturation of the Acute Hepatic TLR4/NF-ΚB Mediated Innate Immune Response Is P65 Dependent in Mice. Front. Immunol. 2020, 11, 1892. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Jo, E.H.; Lee, B.; Noh, H.M.; Park, S.; Lee, Y.M.; Kim, D.K.; Park, M.C. Soshiho-Tang, a Traditional Herbal Medicine, Alleviates Atopic Dermatitis Symptoms via Regulation of Inflammatory Mediators. Front. Pharmacol. 2019, 10, 742. [Google Scholar] [CrossRef]
- Auteri, M.; Zizzo, M.G.; Serio, R. GABA and GABA Receptors in the Gastrointestinal Tract: From Motility to Inflammation. Pharmacol. Res. 2015, 93, 11–21. [Google Scholar] [CrossRef]
- Gibbs, N.K. L-Histidine Supplementation in Adults and Young Children with Atopic Dermatitis (Eczema). J. Nutr. 2020, 150, 2576S–2579S. [Google Scholar] [CrossRef] [PubMed]
- Aranda, C.S.; Pereira, K.C.; Velasco, H.F.; Mallozi, M.C.; Andrade, R.S.; Rodrigues, W.D.; Pesquero, J.B.; Martins, A.M.; Solé, D. Isoleucine and atopic dermatitis. Pediatr. Allergy Immunol. 2017, 28, 495–497. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence |
|---|---|---|
| GAPDH | Forward | 5’-CCCCTGGCCAAGGTCATCCATGACAACTTT-3’ |
| Reverse | 5’-GGCCATGAGGTCCACCACCCTGTTGCTGTA-3’ | |
| TNF-α | Forward | 5’-CCCTCCAGTTCTAGTTCTATC-3’ |
| Reverse | 5’-GGGGAAAGAATCATTCAACCAG-3’ | |
| IL-1β | Forward | 5’-ACGATGCACCTGTACGATCA-3’ |
| Reverse | 5’-TCTTTCAACACGCAGGACAG-3’ | |
| IL-8 | Forward | 5’-ACAGCAGAGCACACAAGCTT-3’ |
| Reverse | 5’-CTGGCAACCCTACAACAGAC-3’ | |
| Cox-2 | Forward | 5’-AACAGGAGCATCCTGAATGG-3’ |
| Reverse | 5’-GGTCAATGGAAGCCTGTGAT-3’ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, M.; Choi, Y.-J.; Wedamulla, N.E.; Zhang, Q.; Kim, S.W.; Bae, S.M.; Seok, Y.-S.; Kim, E.-K. Use of a Silkworm (Bombyx mori) Larvae By-Product for the Treatment of Atopic Dermatitis: Inhibition of NF-κB Nuclear Translocation and MAPK Signaling. Nutrients 2023, 15, 1775. https://doi.org/10.3390/nu15071775
Fan M, Choi Y-J, Wedamulla NE, Zhang Q, Kim SW, Bae SM, Seok Y-S, Kim E-K. Use of a Silkworm (Bombyx mori) Larvae By-Product for the Treatment of Atopic Dermatitis: Inhibition of NF-κB Nuclear Translocation and MAPK Signaling. Nutrients. 2023; 15(7):1775. https://doi.org/10.3390/nu15071775
Chicago/Turabian StyleFan, Meiqi, Young-Jin Choi, Nishala Erandi Wedamulla, Qun Zhang, Seong Wan Kim, Sung Moon Bae, Young-Seek Seok, and Eun-Kyung Kim. 2023. "Use of a Silkworm (Bombyx mori) Larvae By-Product for the Treatment of Atopic Dermatitis: Inhibition of NF-κB Nuclear Translocation and MAPK Signaling" Nutrients 15, no. 7: 1775. https://doi.org/10.3390/nu15071775
APA StyleFan, M., Choi, Y.-J., Wedamulla, N. E., Zhang, Q., Kim, S. W., Bae, S. M., Seok, Y.-S., & Kim, E.-K. (2023). Use of a Silkworm (Bombyx mori) Larvae By-Product for the Treatment of Atopic Dermatitis: Inhibition of NF-κB Nuclear Translocation and MAPK Signaling. Nutrients, 15(7), 1775. https://doi.org/10.3390/nu15071775