
Consumption of Nutritionally Enriched Hen Eggs Enhances Endothelium-Dependent Vasodilation via Cyclooxygenase Metabolites in Healthy Young People—A Randomized Study

 , , ,
, , ,  ,
,  ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
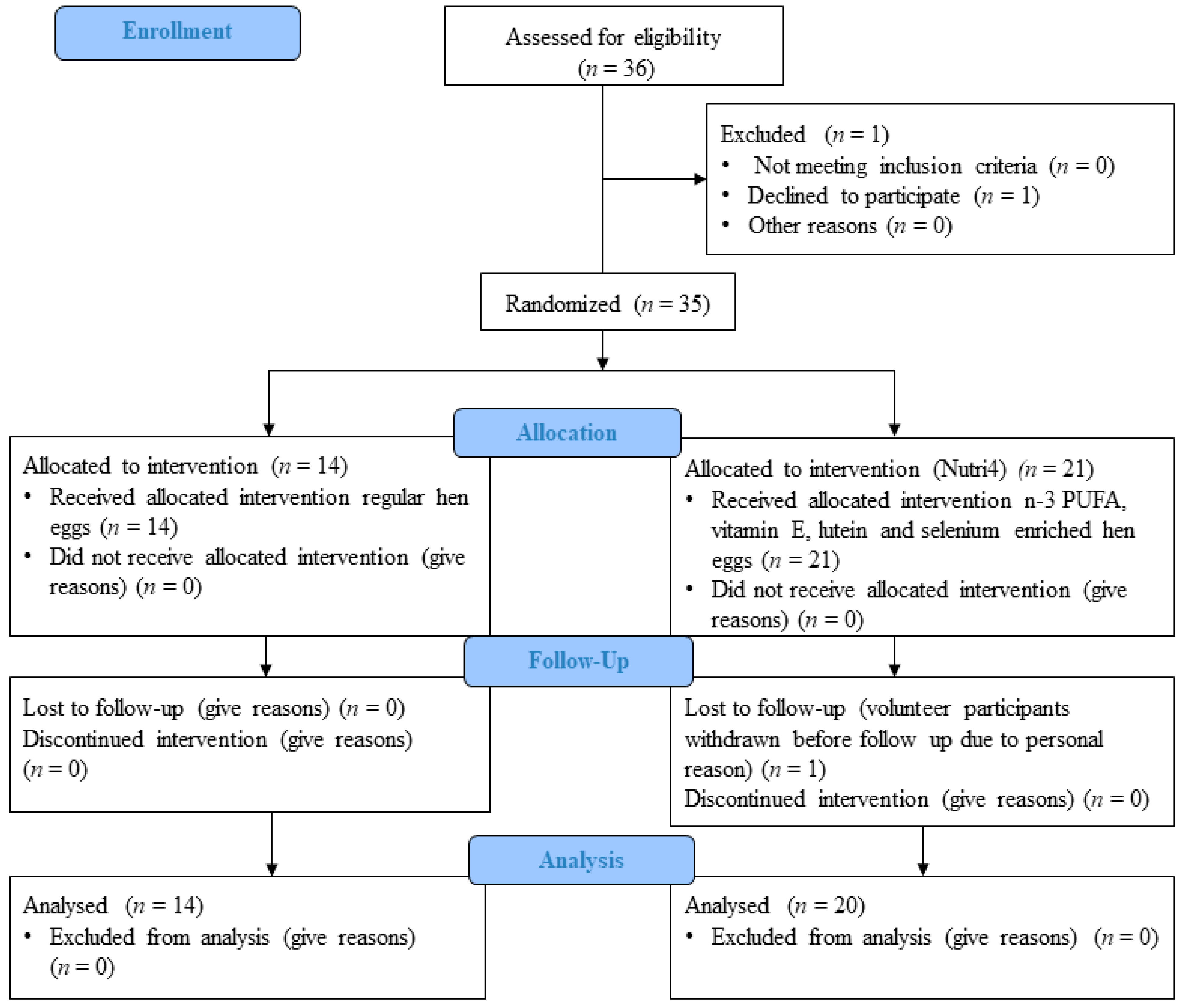
2.1. The Study Population
2.2. Study Protocol
2.3. Anthropometric and Arterial Blood Pressure Measurements
2.4. Assessment of Microvascular Endothelium-Dependent and Endothelium-Independent Vasodilation
2.5. Isolation Peripheral Blood Mononuclear Cell (PBMC)
2.6. Protein Expression of Enzymes Important in the Mechanisms of Microvascular Reactivity
2.7. Venous Blood Sample Analysis
2.8. Measurements of Serum Fatty Acids Profile
2.9. Measurements of Serum Vitamin E Concentration
2.10. Serum Selenium Concentration Measurements
2.11. Measurements of Serum Lutein Concentration
2.12. Statistical Analysis
3. Results
3.1. Effects of Diets on Anthropometric and Biochemical Measurements in Study Population
3.2. Free Fatty Acids Profile, Vitamin E, Lutein, and Selenium in Serum
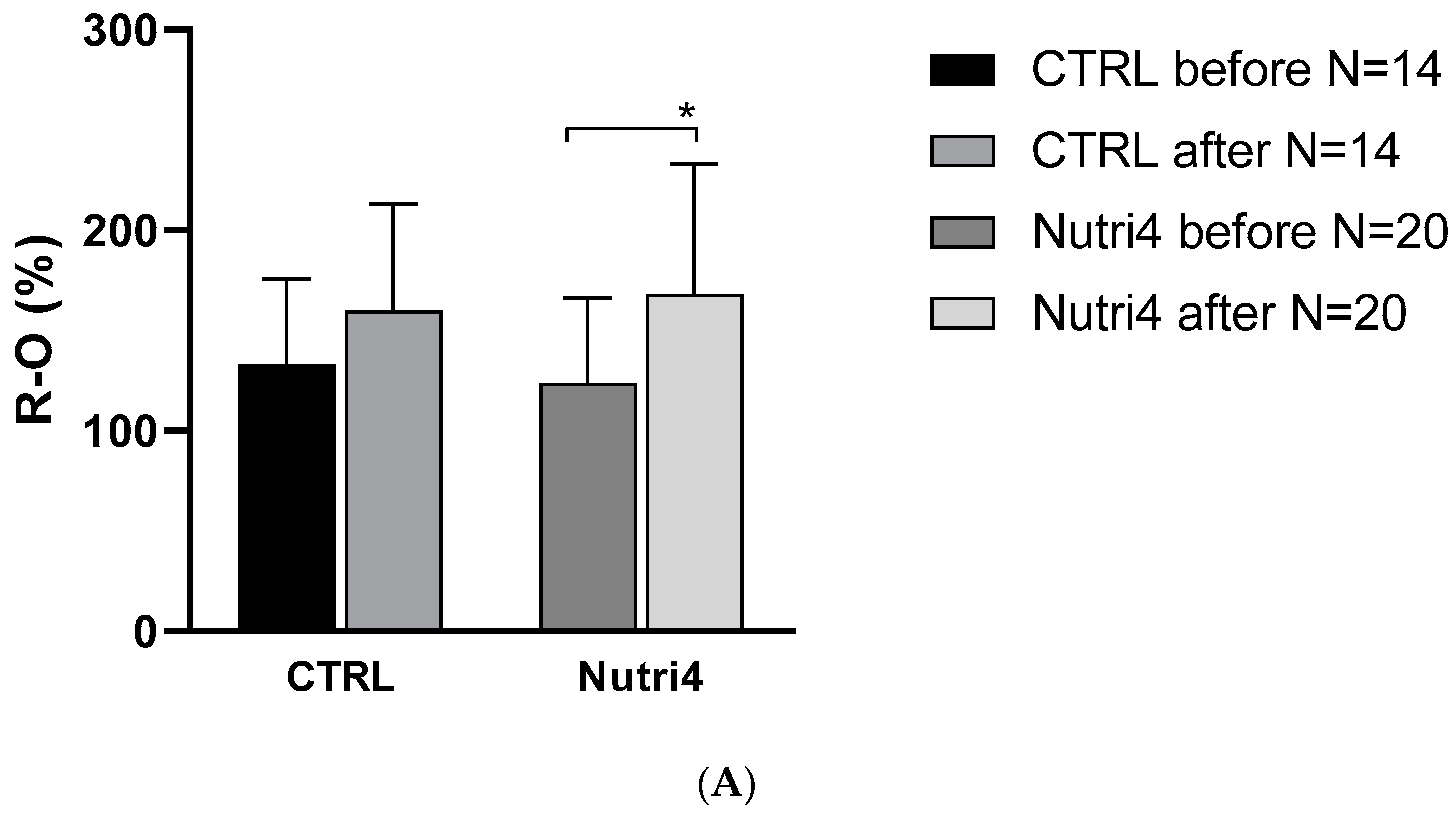
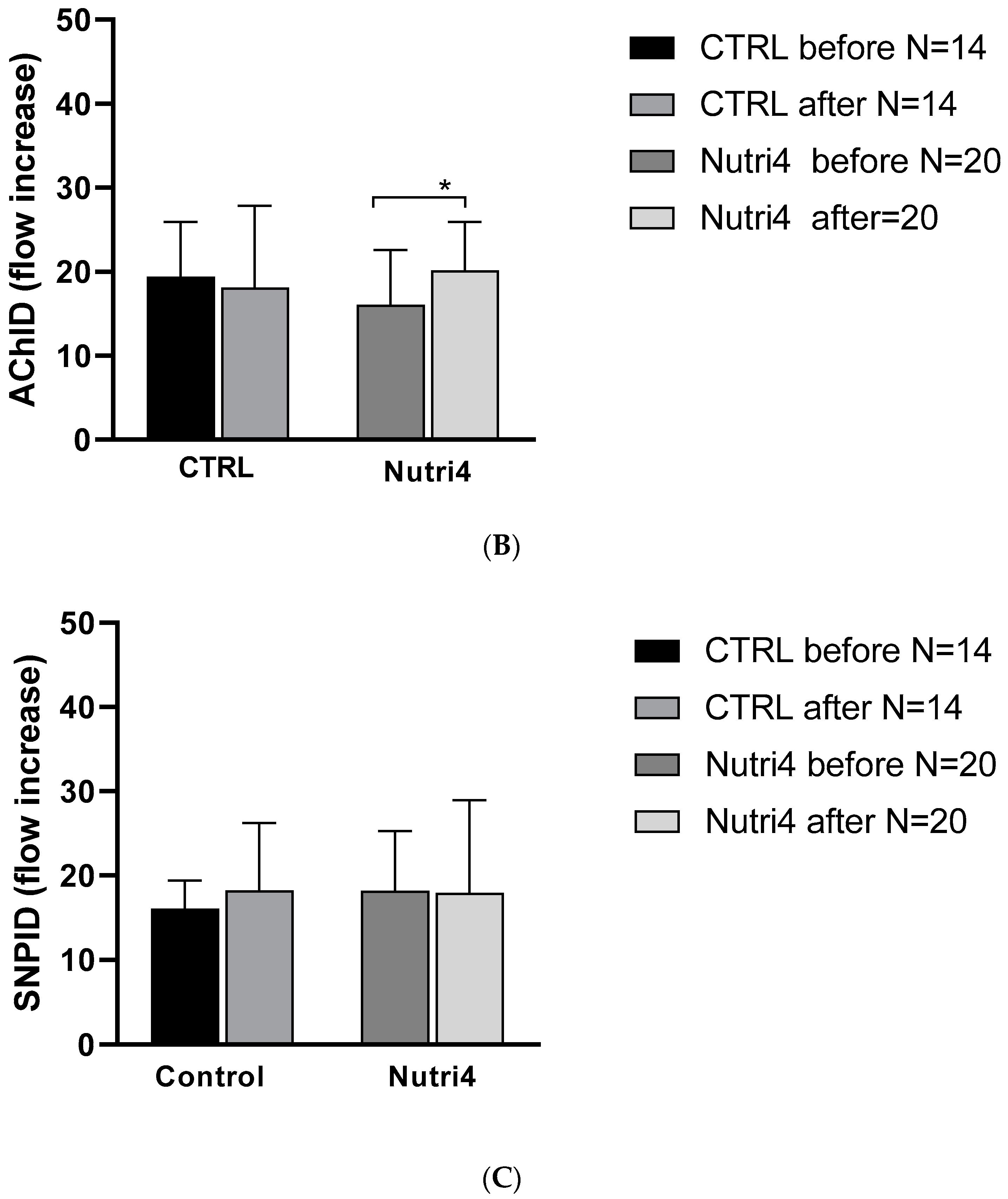
3.3. Skin Microvascular Reactivity to PORH, Acetylcholine, and Sodium Nitroprusside
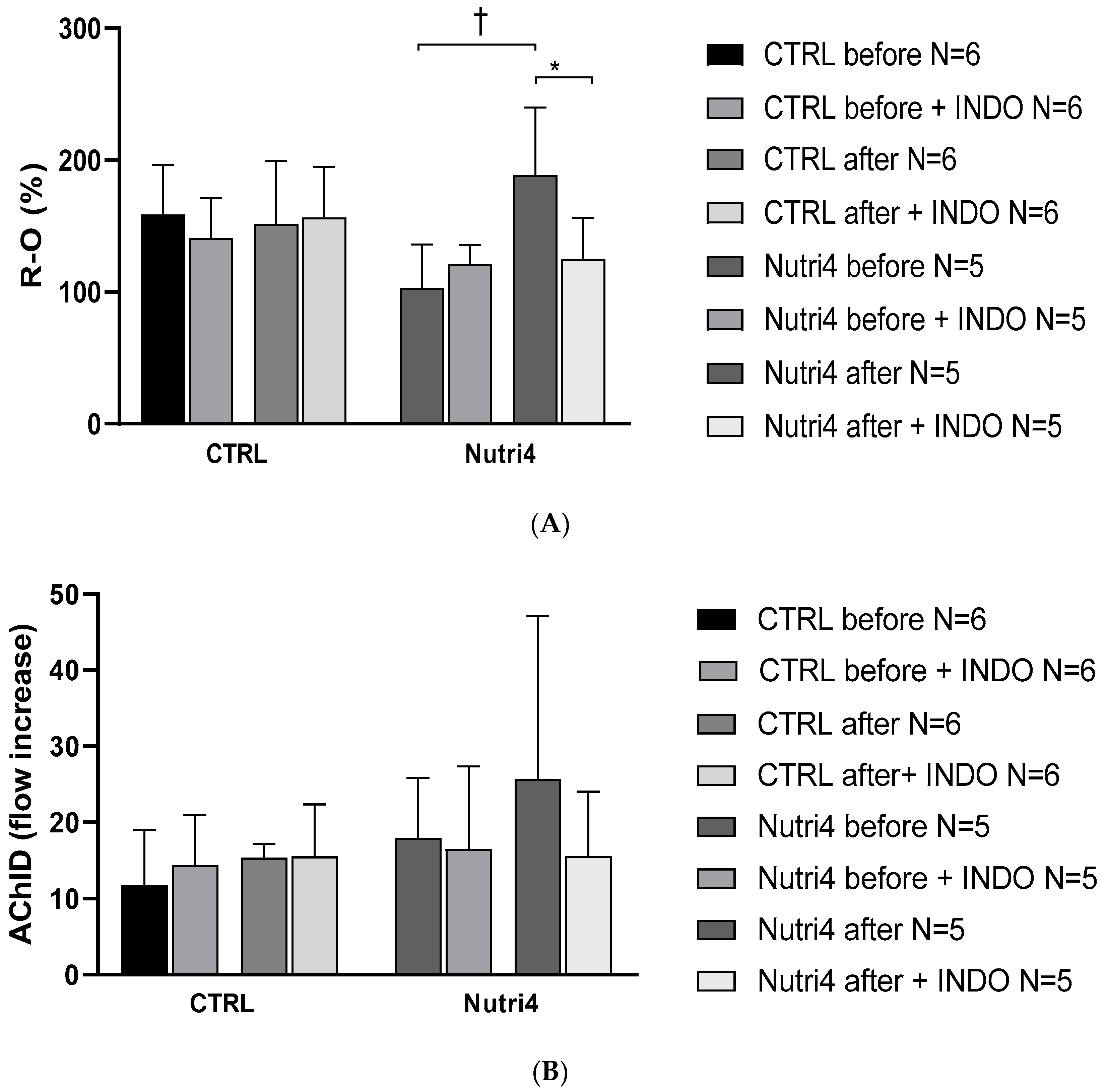
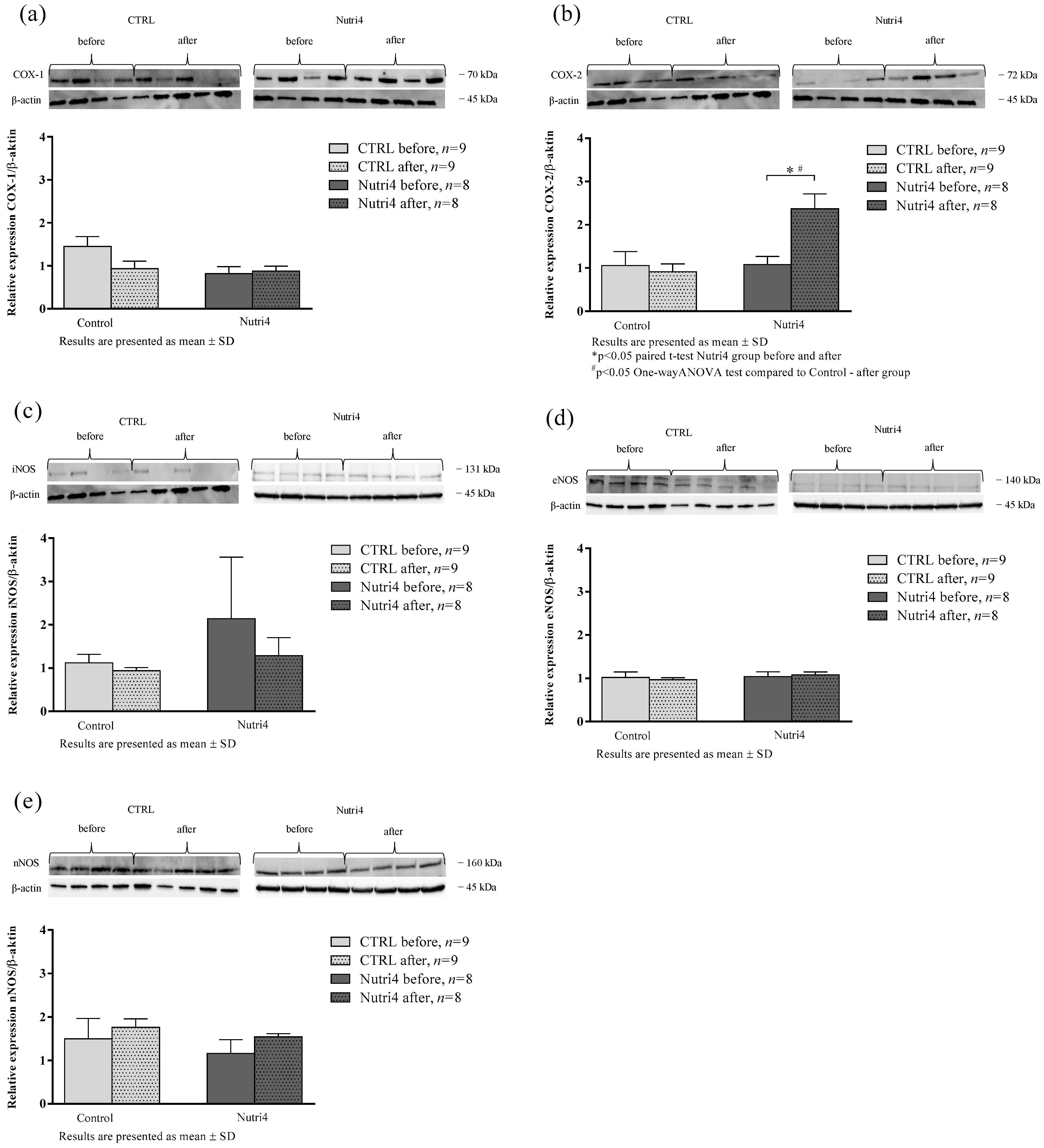
3.4. Protein Expression of Enzymes Important in the Mechanisms of Microvascular Reactivity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Auger, C.; Said, A.; Nguyen, P.N.; Chabert, P.; Idris-Khodja, N.; Schini-Kerth, V.B. Potential of food and natural products to promote endothelial and vascular health. J. Cardiovasc. Pharmacol. 2016, 68, 11–18. [Google Scholar] [CrossRef]
- Ulu, A.; Lee, K.S.S.; Miyabe, C.B.; Yang, J.; Hammock, B.G.; Dong, H. An omega-3 epoxide of docosahexaenoic acid lowers blood pressure in angiotensin-II—Dependent hypertension. J. Cardiovasc. Pharmacol. 2014, 64, 87–99. [Google Scholar] [CrossRef]
- Renier, G.; Skamene, E.; DeSanctis, J.; Radzioch, D. Dietary n-3 polyunsaturated fatty acids prevent the development of atherosclerotic lesions in mice. Modulation of macrophage secretory activities. Arter. Thromb. A J. Vasc. Biol. 1993, 13, 1515–1524. [Google Scholar] [CrossRef]
- Kris-Etherton, P.M.; Harris, W.S.; Appel, L.J. Fish consumption, fish oil, omega-3 fatty acids, and cardiovascular disease. Circulation 2002, 106, 2747–2757. [Google Scholar] [CrossRef]
- Schachinger, V.; Britten, M.B.; Zeiher, A.M. Prognostic impact of coronary vasodilator dysfunction on adverse long-term outcome of coronary heart disease. Circulation 2000, 101, 1899–1906. [Google Scholar] [CrossRef]
- Stupin, A.; Mihalj, M.; Kolobarić, N.; Šušnjara, P.; Kolar, L.; Mihaljević, Z.; Matić, A.; Stupin, M.; Jukić, I.; Kralik, Z.; et al. Anti-inflammatory potential of n-3 polyunsaturated fatty acids enriched hen eggs consumption in improving microvascular endothelial function of healthy individuals—Clinical trial. Int. J. Mol. Sci. 2020, 21, 4149. [Google Scholar] [CrossRef]
- Stupin, A.; Rasic, L.; Matic, A.; Stupin, M.; Kralik, Z.; Kralik, G.; Grcevic, M.; Drenjancevic, I. Omega-3 polyunsaturated fatty acids-enriched hen eggs consumption enhances microvascular reactivity in young healthy individuals. Appl. Physiol. Nutr. Metab. 2018, 43, 988–995. [Google Scholar] [CrossRef]
- Barić, L.; Drenjančević, I.; Mihalj, M. Enhanced antioxidative defense by vitamins C and E consumption prevents 7-day high-salt diet-induced microvascular endothelial function impairment in young healthy individuals. J. Clin. Med. 2020, 9, 843. [Google Scholar] [CrossRef]
- Ćuric, Ž.B.; Masle, A.M.; Kibel, A. Effects of n-3 polyunsaturated fatty acid-enriched hen egg consumption on the inflammatory biomarkers and microvascular function in patients with acute and chronic coronary syndrome—A randomized study. Biology 2021, 10, 774. [Google Scholar] [CrossRef]
- Armah, C.K.; Jackson, K.G.; Doman, I.; James, L.; Cheghani, F.; Minihane, A.M. Fish oil fatty acids improve postprandial vascular reactivity in healthy men. Clin. Sci. 2008, 114, 679–686. [Google Scholar] [CrossRef]
- Jiang, Q. Natural forms of vitamin E: Metabolism, antioxidant and anti-inflammatory activities and the role in disease pre-vention and therapy. Free Radic Biol. Med. 2014, 72, 76–90. [Google Scholar] [CrossRef]
- Yan, J.-H.; Guan, B.-J.; Gao, H.-Y.; Peng, X.-E. Omega-3 polyunsaturated fatty acid supplementation and non-alcoholic fatty liver disease. Medicine 2018, 97, e12271. [Google Scholar] [CrossRef]
- Traber, M.G.; Stevens, J.F. Vitamins C and E: Beneficial effects from a mechanistic perspective. Free. Radic. Biol. Med. 2011, 51, 1000–1013. [Google Scholar] [CrossRef] [PubMed]
- Leme Goto, P.; Cinato, M.; Merachli, F. In vitro and in vivo cardioprotective and metabolic efficacy of vitamin E TPGS/Apelin. J. Mol. Cell. Cardiol. 2020, 138, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Mehrdad, R.; Reza, J.N.M.; Tourandokht, B. Endothelium-dependent effect of sesame seed feeding on vascular reactivity of streptozotocin-diabetic rats: Underlying mechanisms. Iran. J. Pharm. Res. 2013, 12, 377–385. [Google Scholar]
- Kipp, A.P.; Strohm, D.; Brigelius-Flohé, R. Revised reference values for selenium intake. J. Trace Elem. Med. Biol. 2015, 32, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Xing, H.; Liu, C.; Zhang, Z.; Xu, S. Effect of selenium deficiency on nitric oxide and heat shock proteins in chicken erythrocytes. Biol. Trace Element Res. 2015, 171, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Kuruppu, D.; Hendrie, H.C.; Yang, L.; Gao, S. Selenium levels and hypertension: A systematic review of the literature. Public Health Nutr. 2013, 17, 1342–1352. [Google Scholar] [CrossRef]
- Rees, K.; Hartley, L.; Day, C. Selenium supplementation for the primary prevention of cardiovascular disease. Cochrane Database Syst. Rev. 2013, 2013, CD009671. [Google Scholar] [CrossRef]
- Hajizadeh-Sharafabad, F.; Ghoreishi, Z.; Maleki, V.; Tarighat-Esfanjani, A. Mechanistic insights into the effect of lutein on atherosclerosis, vascular dysfunction, and related risk factors: A systematic review of in vivo, ex vivo and in vitro studies. Pharmacol. Res. 2019, 149, 104477. [Google Scholar] [CrossRef]
- Ahn, Y.J.; Kim, H. Lutein as a modulator of oxidative stress-mediated inflammatory diseases. Antioxidants 2021, 10, 1448. [Google Scholar] [CrossRef]
- Drenjancevic, I. Effect of Enriched QUARTET® Hen Eggs on Cardiovascular Function in Cardiovascular Patients and Healthy Individuals. Clinical Trial Registration study/NCT04564690, clinicaltrials.gov. Available online: https://clinicaltrials.gov/ct2/show/study/NCT04564690 (accessed on 17 October 2022).
- Suresh, K. An overview of randomization techniques: An unbiased assessment of outcome in clinical research. J. Hum. Reprod. Sci. 2011, 4, 8–11. [Google Scholar] [CrossRef]
- Kralik, Z.; Kralik, G.; Grčević, M.; Kralik, I.; Gantner, V. Physical-chemical characteristics of designer and conventional eggs. Braz J. Poult. Sci. 2018, 20, 119–126. [Google Scholar] [CrossRef]
- Kralik, Z.; Kralik, G.; Galović, O. Nutricines content in table eggs of Croatian producers. Book of Abstracts of the 13th International Scientific and Professional Conference with Food to Health = Knjiga Sažetaka s 13 Međunarodnog Znanstveno-Stručnog skupa Hranom do Zdravlja. 2011. Available online: https://www.bib.irb.hr/1147732 (accessed on 15 April 2022).
- Cavka, A.; Cosic, A.; Jukic, I.; Jelakovic, B.; Lombard, J.H.; Phillips, S.A.; Seric, V.; Mihaljevic, I.; Drenjancevic, I. The role of cyclo-oxygenase-1 in high-salt diet-induced microvascular dysfunction in humans. J. Physiol. 2015, 593, 5313–5324. [Google Scholar] [CrossRef]
- Stupin, M.; Stupin, A.; Rasic, L.; Cosic, A.; Kolar, L.; Seric, V.; Lenasi, H.; Izakovic, K.; Drenjancevic, I. Acute exhaustive rowing exercise reduces skin microvascular dilator function in young adult rowing athletes. Eur. J. Appl. Physiol. 2017, 118, 461–474. [Google Scholar] [CrossRef]
- Walser, B.; Giordano, R.M.; Stebbins, C.L. Supplementation with omega-3 polyunsaturated fatty acids augments brachial artery dilation and blood flow during forearm contraction. Eur. J. Appl. Physiol. 2006, 97, 347–354. [Google Scholar] [CrossRef]
- Kolobarić, N.; Drenjančević, I.; Matić, A. Dietary intake of n-3 PUFA-enriched hen eggs changes inflammatory markers’ concentration and Treg/Th17 cells distribution in blood of young healthy adults—A randomised study. Nutrients 2021, 13, 1851. [Google Scholar] [CrossRef]
- Mihaljević, Z.; Matić, A.; Stupin, A.; Frkanec, R.; Tavčar, B.; Kelava, V.; Bujak, I.T.; Kolobarić, N.; Kibel, A.; Drenjančević, I. Arachidonic acid metabolites of CYP450 enzymes and HIF-1α modulate endothelium-dependent vasorelaxation in sprague-dawley rats under acute and intermittent hyperbaric oxygenation. Int. J. Mol. Sci. 2020, 21, 6353. [Google Scholar] [CrossRef]
- Matic, A.; Jukic, I.; Stupin, A.; Baric, L.; Mihaljevic, Z.; Unfirer, S.; Bujak, I.T.; Mihaljevic, B.; Lombard, J.H.; Drenjancevic, I. High salt intake shifts the mechanisms of flow-induced dilation in the middle cerebral arteries of Sprague-Dawley rats. Am. J. Physiol. Circ. Physiol. 2018, 315, H718–H730. [Google Scholar] [CrossRef]
- Mihaljević, Z.; Matić, A.; Stupin, A.; Rašić, L.; Jukić, I.; Drenjančević, I. Acute hyperbaric oxygenation, contrary to intermittent hyperbaric oxygenation, adversely affects vasorelaxation in healthy sprague-dawley rats due to increased oxidative stress. Oxidative Med. Cell Longev. 2018, 2018, e7406027. [Google Scholar] [CrossRef]
- Wang, L.Y.; Summerhill, K.; Rodriguez-Canas, C.; Mather, I.; Patel, P.; Eiden, M.; Young, S.; Forouhi, N.G.; Koulman, A. Development and validation of a robust automated analysis of plasma phospholipid fatty acids for metabolic phenotyping of large epidemiological studies. Genome Med. 2013, 5, 39. [Google Scholar] [CrossRef] [PubMed]
- Jargar, J.G.; Dhundasi, S.A.; Punekar, M.D. Status of α-tocopherol concentration and oxidative stress in infertile females of Vija-yapur District, northern Karnataka. Natl. J. Physiol. Pharm. Pharmacol. 2017, 7, 5. [Google Scholar] [CrossRef]
- Tzeng, M.-S.; Yang, F.-L.; Wang-Hsu, G.-S.; Chen, B.-H. Determination of major carotenoids in human serum by liquid chromatography. J. Food Drug Anal. 2004, 12, 1709–1712. [Google Scholar] [CrossRef]
- Leeson, S.; Caston, L. Enrichment of eggs with lutein. Poult. Sci. 2004, 83, 1709–1712. [Google Scholar] [CrossRef] [PubMed]
- Das, U.N. Arachidonic acid in health and disease with focus on hypertension and diabetes mellitus: A review. J. Adv. Res. 2018, 11, 43–55. [Google Scholar] [CrossRef]
- Schmitz, G.; Ecker, J. The opposing effects of n-3 and n-6 fatty acids. Prog. Lipid Res. 2008, 47, 147–155. [Google Scholar] [CrossRef]
- Arnold, C.; Konkel, A.; Fischer, R. Cytochrome P450-dependent metabolism of w-6 and w-3 long-chain polyunsaturated fatty acids. Pharmacol. Rep. 2010, 12, 536–547. [Google Scholar] [CrossRef]
- Kolar, L.; Stupin, M.; Stupin, A. Does the endothelium of competitive athletes benefit from consumption of n-3 poly-unsaturated fatty acid-enriched hen eggs? Prev. Nutr. Food Sci. 2021, 26, 388–399. [Google Scholar] [CrossRef]
- Stupin, A.; Cosic, A.; Novak, S.; Vesel, M.; Jukic, I.; Popovic, B.; Karalic, K.; Loncaric, Z.; Drenjancevic, I. Reduced dietary selenium impairs vascular function by increasing oxidative stress in Sprague-Dawley rat aortas. Int. J. Environ. Res. Public Health 2017, 14, 591. [Google Scholar] [CrossRef]
- Minieri, M.; Di Nardo, P. Nutrients: The environmental regulation of cardiovascular gene expression. Genes Nutr. 2007, 2, 163–168. [Google Scholar] [CrossRef]
- Iacoviello, L.; Santimone, I.; Latella, M.C.; de Gaetano, G.; Donati, M.B. Nutrigenomics: A case for the common soil between cardiovascular disease and cancer. Genes Nutr. 2008, 3, 19–24. [Google Scholar] [CrossRef]
- Marchioli, R.; Barzi, F.; Bomba, E. Early protection against sudden death by n-3 polyunsaturated fatty acids after myocardial infarction. Circulation 2002, 105, 1897–1903. [Google Scholar] [CrossRef]
- Streppel, M.T.; Ocké, M.C.; Boshuizen, H.C.; Kok, F.J.; Kromhout, D. Long-term fish consumption and n-3 fatty acid intake in relation to (sudden) coronary heart disease death: The Zutphen study. Eur. Heart J. 2008, 29, 2024–2030. [Google Scholar] [CrossRef]
- Tousoulis, D.; Plastiras, A.; Siasos, G.; Oikonomou, E.; Verveniotis, A.; Kokkou, E.; Maniatis, K.; Gouliopoulos, N.; Miliou, A.; Paraskevopoulos, T.; et al. Omega-3 PUFAs improved endothelial function and arterial stiffness with a parallel antiinflammatory effect in adults with metabolic syndrome. Atherosclerosis 2014, 232, 10–16. [Google Scholar] [CrossRef]
- Tagawa, H.; Shimokawa, H.; Tagawa, T. Long-term treatment with Eicosapentaenoic acid augments both nitric oxide-mediated and non-nitric oxide-mediated endothelium-dependent forearm vasodilatation in patients with coronary artery disease. J. Cardiovasc. Pharmacol. 1999, 33, 633–640. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| An L-Grade Egg with an Average Weight of 68 g Has about 60 g of Edible Portion | ||
|---|---|---|
| Parameters | Regular Eggs | Nutri4 Eggs |
| Vitamin E (mg) | 0.595 ± 0.174 | 1.098 * ± 0.339 |
| Lutein (mg) | 0.11 ± 0.011 | 0.616 * ± 0.085 |
| Selenium (mg) | 0.0183 ± 0.002 | 0.02305 * ± 0.0012 |
| Fatty acids | ||
| ∑SFA | 1566 ± 346 | 1442 ± 185 |
| ∑MUFA | 1976 ± 189 | 2419 ± 139 |
| ∑n-6 PUFA | 1263 ± 148 | 747 ± 46 * |
| LA | 1165 ± 140 | 702 ± 43 |
| AA | 89 ± 9 | 44 ± 4 * |
| ∑n-3 PUFA | 146 ± 20 | 342 ± 25 * |
| ALA | 71 ± 11 | 189 ± 16 * |
| EPA | n.d. | 19 ± 2 * |
| DHA | 75 ± 11 | 135 ± 11 * |
| ∑n-6/∑n-3 PUFA | 8.71 | 2.18 * |
| Parameter | CTRL | Nutri4 | ||
|---|---|---|---|---|
| Before | After | Before | After | |
| N (W/M) | 14 (6/9) | 20 (7/13) | ||
| Age (years) | 22 ± 3 | 22 ± 2 | ||
| BW (kg) | 78 ± 17 | 77 ± 15 | 72 ± 15 | 72 ± 15 |
| BMI (kg/m2) | 24.3 ± 3.6 | 24.0 ± 3.2 | 23.5 ± 3.1 | 23.5 ± 3.1 |
| WHR | 0.84 ± 0.07 | 0.84 ± 0.07 | 0.81 ± 0.10 | 0.81 ± 0.10 |
| SBP (mmHg) | 110 ± 17 | 108 ± 15 | 109 ± 11 | 104 ± 14 |
| DBP (mmHg) | 72 ± 12 | 69 ± 4 | 78 ± 16 | 72 ± 6 |
| MAP (mmHg) | 85 ± 7 | 82 ± 7 | 88 ± 13 | 84 ± 6 |
| HR (beats per minute) | 65 ± 8 | 65 ± 9 | 78 ± 12 † | 78 ± 11 † |
| Erythrocytes (×1012/L) | 4.7 ± 0.4 | 4.8 ± 0.4 | 4.9 ± 0.5 | 4.9 ± 0.5 |
| Hemoglobin (g/L) | 141 ± 14 | 141 ± 16 | 143 ± 12 | 142 ± 13 |
| Hematocrit (%) | 39 ± 3 | 32 ± 16 | 40 ± 3 | 38 ± 10 |
| Leukocytes (×109/L) | 6.1 ± 1.9 | 6.0 ± 1.2 | 6.3 ± 1.7 | 5.8 ± 1.9 |
| Thrombocytes (×109/L) | 201 ± 65 | 216 ± 64 | 233 ± 32 | 221 ± 32 |
| Urea (mmol/L) | 5.0 ± 1.5 | 5.5 ± 1.3 | 5.0 ± 1.0 | 5.7 ± 1.6 * |
| Creatinine (µmol/L) | 86 ± 18 | 87 ± 14 | 78 ± 13 | 75 ± 12 |
| Urate (µmol/L) | 330 ± 67 | 318 ± 88 | 296 ± 77 | 289 ± 58 |
| Aspartate aminotransferase (U/L) | 25 ± 8 | 25 ± 6 | 22 ± 6 | 22 ± 5 |
| Alanine aminotransferase (U/L) | 23 ± 8 | 22 ± 7 | 20 ± 15 | 23 ± 12 |
| Gamma-glutamyltransferase (U/L) | 14 ± 4 | 14 ± 4 | 17 ± 12 | 18 ± 11 |
| Sodium (mmol/L) | 139 ± 2 | 139 ± 2 | 139± 2 | 139 ± 2 |
| Potassium (mmol/L) | 4.2 ± 0.3 | 4.3 ± 0.3 | 4.1 ± 0.2 | 4.2 ± 0.3 |
| Iron (µmol/L) | 16.4 ± 6.7 | 13.8 ± 7.1 | 21.3 ± 6.3 * | 15.5 ± 6.3 |
| Transferrin (g/L) | 2.73 ± 0.5 | 2.78 ± 0.71 | 2.62 ± 0.34 | 2.66 ± 0.32 |
| Glucose (mmol/L) | 4.6 ± 0.3 | 4.6 ± 0.5 | 4.4 ± 0.4 | 4.9 ± 0.8 * |
| hsCRP (mg/L) | 0.6 ± 0.5 | 1.1 ± 1.5 | 1.1 ± 0.9 | 2.3 ± 0.9 |
| cholesterol (mmol/L) | 4.3 ± 0.8 | 4.6 ± 1.5 | 4.5 ± 0.5 | 4.7 ± 0.7 |
| triglycerides (mmol/L) | 0.8 ± 0.3 | 0.9 ± 0.3 | 1.1 ± 0.5 | 0.9 ± 0.3 |
| HDL cholesterol (mmol/L) | 1.5 ± 0.4 | 1.4 ± 0.4 | 1.3 ± 0.2 | 1.3 ± 0.2 |
| LDL cholesterol (mmol/L) | 2.5 ± 0.7 | 2.9 ± 0.8 * | 2.8 ± 0.4 | 2.9 ± 0.7 |
| Parameter | CTRL | Nutri4 | |||
|---|---|---|---|---|---|
| Before | After | Before | After | ||
| SFA (μmol/L) | |||||
| C4:0 Butyric acid | N/F | N/F | N/F | N/F | |
| C6:0 Caproic acid | N/F | N/F | N/F | N/F | |
| C8:0 Caprylic acid | N/F | N/F | N/F | N/F | |
| C10:0 Capric acid | <LOQ | <LOQ | <LOQ | <LOQ | |
| C11:0 Undecylic acid | N/F | N/F | N/F | N/F | |
| C12:0 Lauric acid | <LOQ | 20.80 | <LOQ | <LOQ | |
| C13:0 Tridecylic acid | <LOQ | <LOQ | <LOQ | <LOQ | |
| C14:0 Myristic acid | 40.4 ± 9.1 | 36.3 ± 6.6 | 50.5 ± 32.2 | 40.3 ± 11.1 | |
| C15:0 Pentadecylic acid | 12.2 ± 1.6 | 12.7 ± 1.3 | 13.7 ± 2.4 | 27.9 ± 39.6 | |
| C16:0 Palmitic Acid | 1258 ± 350 | 1304 ± 332 | 1295 ± 431 | 1242 ± 314 | |
| C17:0 Margaric acid | 13.5 ± 1.7 | 13.6 ± 2.6 | 15.8 ± 3.8 | 14.7 ± 4.0 | |
| C18:0 Stearic acid | 404 ± 111 | 416 ± 137 | 515 ± 140 | 491 ± 132 | |
| C20:0 Arachidic acid | <LOQ | <LOQ | <LOQ | <LOQ | |
| C21:0 Heneicosanoic acid | N/F | N/F | N/F | N/F | |
| C22:0 Behenic acid | <LOQ | 10.2 | <LOQ | <LOQ | |
| C23:0 Tricosanoic acid | <LOQ | <LOQ | <LOQ | <LOQ | |
| C24:0 Lignoceric acid | <LOQ | <LOQ | <LOQ | <LOQ | |
| PUFA (μmol/L) | |||||
| n-5 | C14:1[cis-9] Myristoleic acid | <LOQ | <LOQ | 11 | <LOQ |
| C15:1[cis-10] Cis-10-pentadecenoic acid | N/F | N/F | N/F | N/F | |
| n-7 | C16:1[cis-9] Palmitoleic acid | 58.3 ± 14.9 | 54.1 ± 17.7 | 95.4 ± 59.3 † | 73.6 ± 34.8 |
| C17:1[cis-10] cis-10-Heptadecenoic acid | N/F | N/F | 11.7 ± 3.6 | 10.7 ± 1.3 | |
| n-9 | C18:1[trans-9] Elaidic acid | N/F | N/F | N/F | N/F |
| C18:1[cis-9] Oleic acid | 691 ± 218 | 656 ± 135 | 872 ± 393 | 758 ± 279 | |
| C20:1[cis-11] 11-Eicosenoic acid | 7.8 ± 2.0 | 6.2 ± 0.6 | 8.8 ± 4.4 | 9.2 ± 2.2 | |
| C22:1[cis-13] Erucic acid | 9.2 | 9.1 ± 0.7 | 7.4 ± 1.0 | 7.9 | |
| C24:1[cis-15] Nervonic acid | <LOQ | 8.7 ± 0.6 | 7.2 ± 0.7 | 7.8 ± 0.7 | |
| n-6 | C18:2[trans-9,12] Linoelaidic acid | N/F | N/F | 12,5 | N/F |
| C18:2[cis-9,12] Linoleic acid | 1148 ± 300 | 1170 ± 298 | 1223 ± 290 | 1277 ± 461 | |
| C18:3[cis-6,9,12] gamma-Linolenic acid | 19.9 ± 6.7 | 17.2 ± 3.7 | 27.3 ± 13.8 | 23.2 ± 7.4 | |
| C21:2[cis-11,14] Eicosadienoic acid | 8.6 ± 1.9 | 9.5 ± 2.6 | 12.3 ± 5.8 | 11.1 ± 3.6 | |
| C20:3[cis-8,11,14] Dihomo-gamma-linolenic acid | 54.1 ± 16.2 | 54.1 ± 12.9 | 78.7 ± 42.6 | 69.3 ± 42.0 | |
| C20:4[cis-5,8,11,14] Arachidonic acid | 298 ± 52 | 354 ± 53 | 390 ± 127 | 369 ± 106 | |
| C22:2[cis-13,16] 13,16-Docosadienoic acid | N/F | N/F | N/F | N/F | |
| n-3 | C18:3[cis-9,12,15] alpha-Linolenic acid | 17.2 ± 7.7 | 16.0 ± 5.1 | 16.4 ± 6.3 | 21.1 ± 8.0 |
| C20:3[cis-11,14,17] 11,14,17-Eicosatrienoic acid | N/F | N/F | N/F | N/F | |
| C20:4[cis-5,8,11,14] Eicosa-5,8,11,14,17-pentaenoic acid | 15.1 ± 6.9 | 14.1 ± 4.6 | 13.7 ± 2.9 | 18.6 ± 5.6 *† | |
| C22:6[cis-4,7,10,13,16,19] cis-4,7,10,13,16,19-Docosahexaenoic acid | 54.0 ± 19.9 | 89.9 ± 37.7 * | 65.6 ± 25.1 | 132.1 ± 64.7 * | |
| n-6/n-3 ratio | 11.1 | 8.3 | 10.5 | 6.3 * | |
| Vitamin E μg/mL | 10.27 ± 3.67 | 10.30 ± 3.687 | 6.63 ± 3.22 | 11.26 ± 2.87 * | |
| Selenium μg/L | 62.44 ± 9.37 | 66.37± 10.09 | 64.88 ± 17.65 | 68.88 ± 3.082 | |
| Lutein μmol/L | 0.199 ± 0.104 | 0.199 ± 0.202 | 0.153 ± 0.073 | 0.232 ± 0.078 * | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šušnjara, P.; Mihaljević, Z.; Stupin, A.; Kolobarić, N.; Matić, A.; Jukić, I.; Kralik, Z.; Kralik, G.; Miloloža, A.; Pavošević, T.; et al. Consumption of Nutritionally Enriched Hen Eggs Enhances Endothelium-Dependent Vasodilation via Cyclooxygenase Metabolites in Healthy Young People—A Randomized Study. Nutrients 2023, 15, 1599. https://doi.org/10.3390/nu15071599
Šušnjara P, Mihaljević Z, Stupin A, Kolobarić N, Matić A, Jukić I, Kralik Z, Kralik G, Miloloža A, Pavošević T, et al. Consumption of Nutritionally Enriched Hen Eggs Enhances Endothelium-Dependent Vasodilation via Cyclooxygenase Metabolites in Healthy Young People—A Randomized Study. Nutrients. 2023; 15(7):1599. https://doi.org/10.3390/nu15071599
Chicago/Turabian StyleŠušnjara, Petar, Zrinka Mihaljević, Ana Stupin, Nikolina Kolobarić, Anita Matić, Ivana Jukić, Zlata Kralik, Gordana Kralik, Anđelina Miloloža, Tihana Pavošević, and et al. 2023. "Consumption of Nutritionally Enriched Hen Eggs Enhances Endothelium-Dependent Vasodilation via Cyclooxygenase Metabolites in Healthy Young People—A Randomized Study" Nutrients 15, no. 7: 1599. https://doi.org/10.3390/nu15071599
APA StyleŠušnjara, P., Mihaljević, Z., Stupin, A., Kolobarić, N., Matić, A., Jukić, I., Kralik, Z., Kralik, G., Miloloža, A., Pavošević, T., Šerić, V., Lončarić, Z., Kerovec, D., Galović, O., & Drenjančević, I. (2023). Consumption of Nutritionally Enriched Hen Eggs Enhances Endothelium-Dependent Vasodilation via Cyclooxygenase Metabolites in Healthy Young People—A Randomized Study. Nutrients, 15(7), 1599. https://doi.org/10.3390/nu15071599