
The Effect of Omega-9 on Bone Viscoelasticity and Strength in an Ovariectomized Diet-Fed Murine Model

 , ,
, ,  ,
,  , add
Show full author list
, add
Show full author list
Abstract
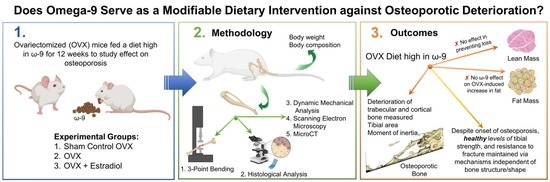
1. Introduction
2. Materials and Methods
2.1. Ovariectomy and Dietary Intervention
2.2. Measurement of Body Weight and Body Composition
2.3. Dynamic Mechanical Analysis
2.4. Three-Point Bending
2.5. Histological Analysis
2.6. Scanning Electron Microscopy
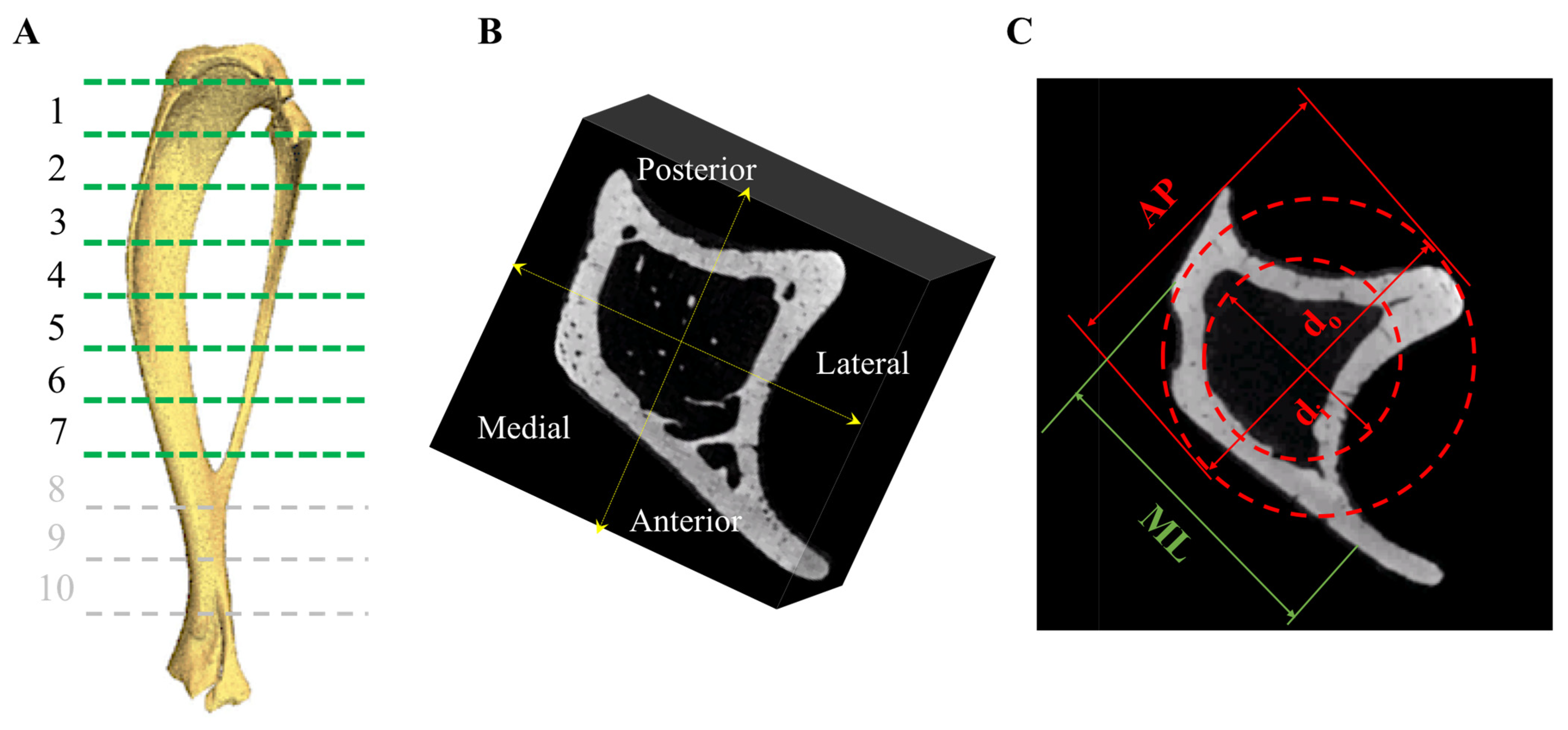
2.7. MicroCT
2.8. Statistical Analysis
3. Results
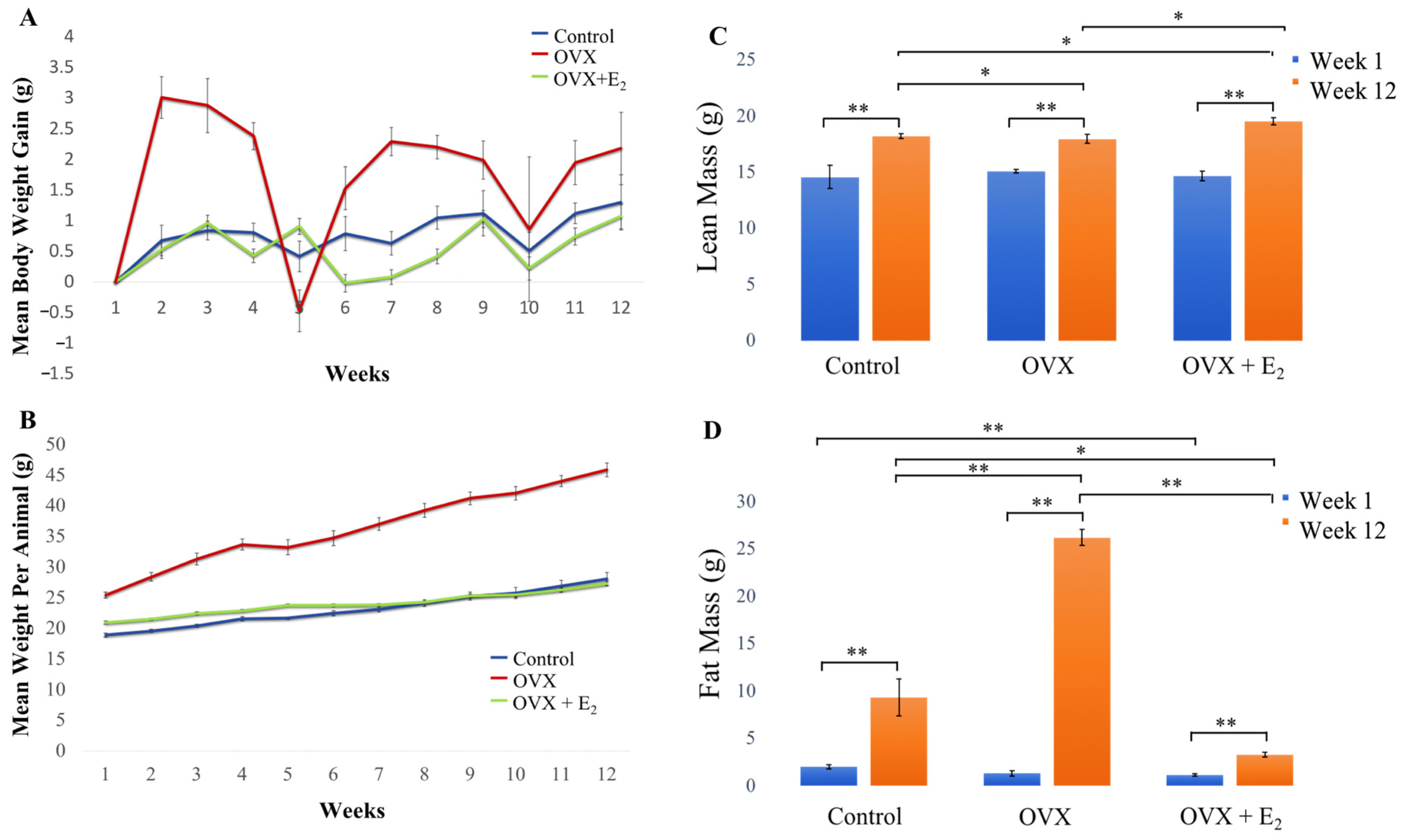
3.1. Body Weight and Body Weight Gain
3.2. Body Composition
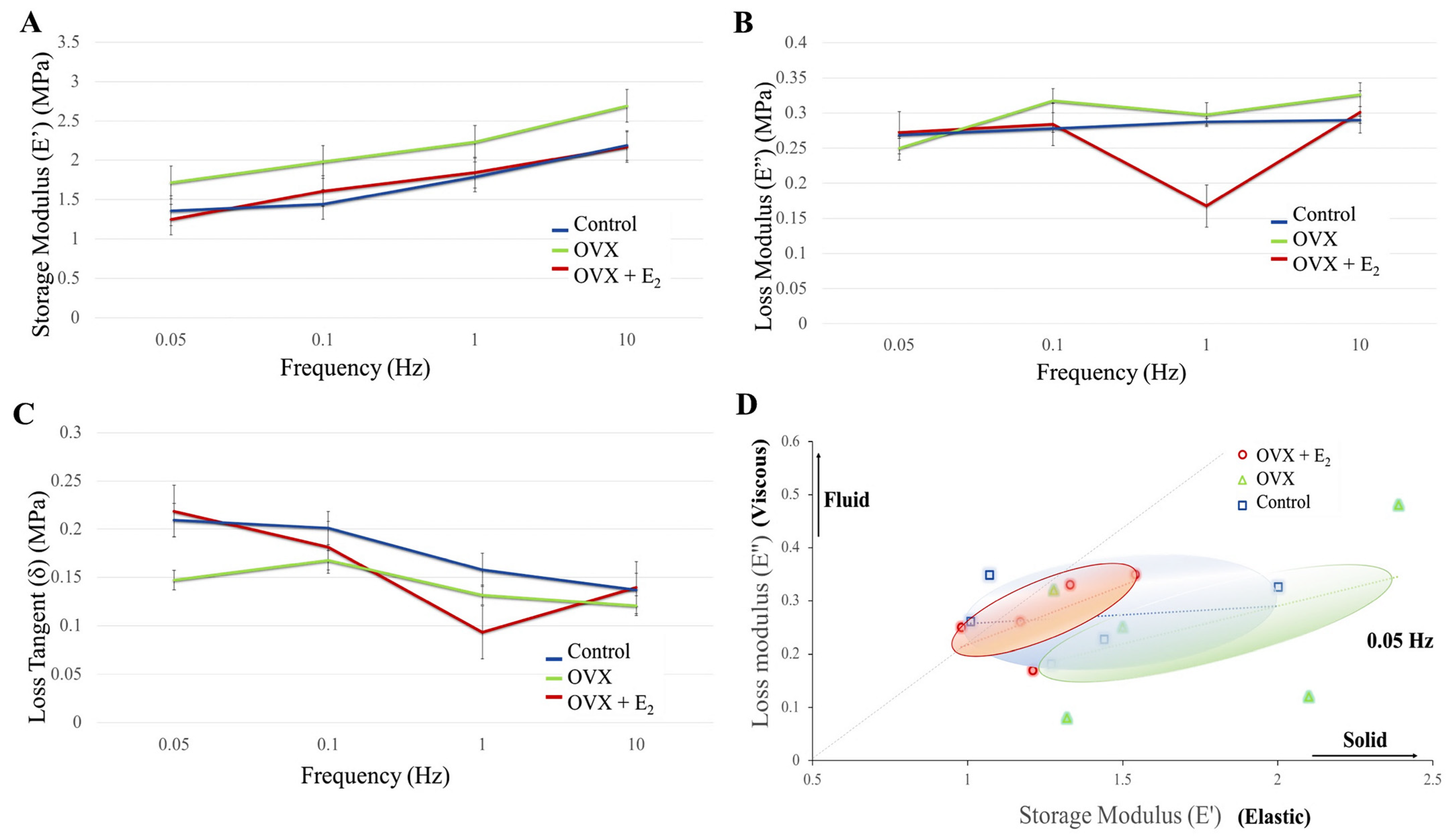
3.3. Dynamic Mechanical Analysis (DMA)
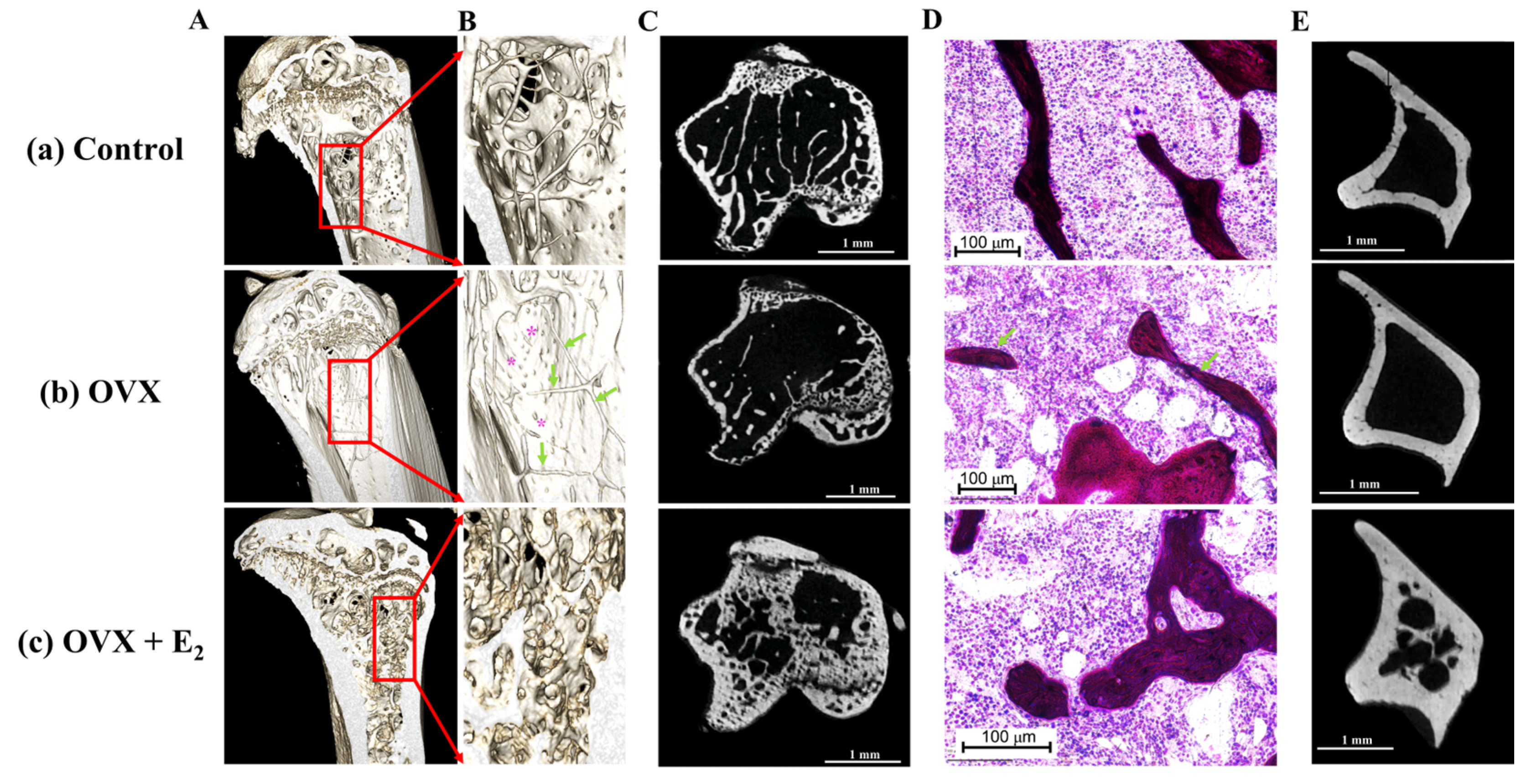
3.4. Tibial Structural Parameters
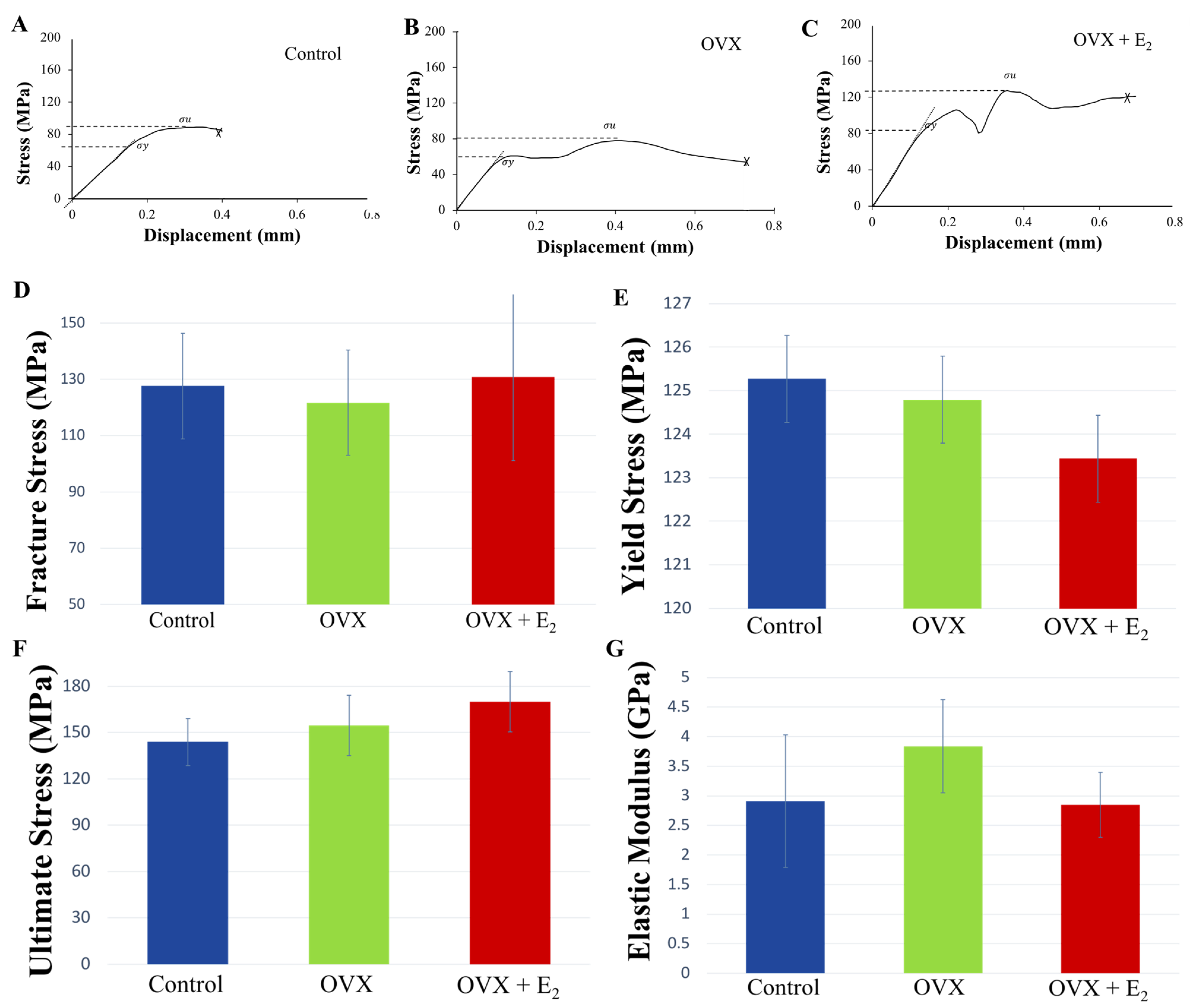
3.5. Three-Point-Bending
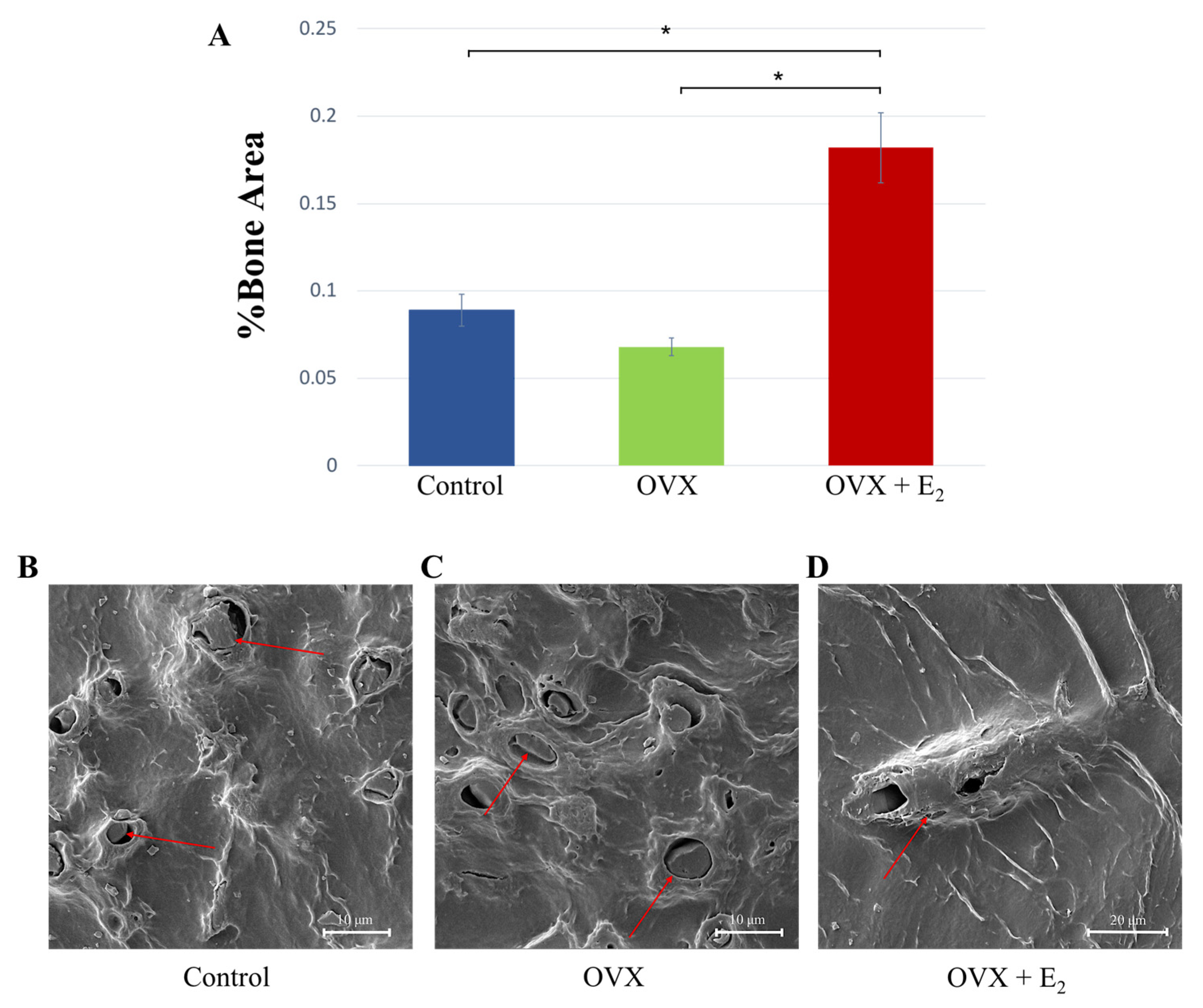
3.6. Histological, Scanning Electron Microscopy, and microCT Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, Q.; Cai, W.; Wang, G.; Shen, X. Prevalence and contributing factors of osteoporosis in the elderly over 70 years old: An epidemiological study of several community health centers in Shanghai. Ann. Palliat. Med. 2020, 9, 231–238. [Google Scholar] [CrossRef]
- Marcucci, G. Rare causes of osteoporosis. Clin. Cases Miner. Bone Metab. 2015, 12, 151–156. [Google Scholar] [CrossRef]
- Peck, W.A. Consensus development conference: Diagnosis, prophylaxis, and treatment of osteoporosis. Am. J. Med. 1993, 94, 646–650. [Google Scholar] [CrossRef]
- Wright, N.C.; Looker, A.C.; Saag, K.G.; Curtis, J.R.; Delzell, E.S.; Randall, S.; Dawson-Hughes, B. The Recent Prevalence of Osteoporosis and Low Bone Mass in the United States Based on Bone Mineral Density at the Femoral Neck or Lumbar Spine. J. Bone Miner. Res. 2014, 29, 2520–2526. [Google Scholar] [CrossRef]
- Salari, N.; Ghasemi, H.; Mohammadi, L.; Behzadi, M.H.; Rabieenia, E.; Shohaimi, S.; Mohammadi, M. The global prevalence of osteoporosis in the world: A comprehensive systematic review and meta-analysis. J. Orthop. Surg. Res. 2021, 16, 609. [Google Scholar] [CrossRef]
- Kemmak, A.R.; Reazpour, A.; Jahangiri, R.; Nikjoo, S.; Farabi, H.; Soleimanpour, S. Economic burden of osteoporosis in the world: A systematic review. Med. J. Islam. Repub. Iran 2020, 34, 154. [Google Scholar] [CrossRef]
- Zhang, X.; Lin, J.; Yang, Y.; Wu, H.; Li, Y.; Yang, X.; Fei, Q. Comparison of three tools for predicting primary osteoporosis in an elderly male population in Beijing: A cross-sectional study. Clin. Interv. Aging 2018, 13, 201–209. [Google Scholar] [CrossRef]
- Conti, V.; Russomanno, G.; Corbi, G.; Toro, G.; Simeon, V.; Filippelli, W.; Ferrara, N.; Grimaldi, M.; D’Argenio, V.; Maffulli, N.; et al. A Polymorphism at the Translation Start Site of the Vitamin D Receptor Gene Is Associated with the Response to Anti-Osteoporotic Therapy in Postmenopausal Women from Southern Italy. Int. J. Mol. Sci. 2015, 16, 5452–5466. [Google Scholar] [CrossRef]
- Tarazona-Santabalbina, F.J.; Belenguer-Varea, Á.; Rovira-Daudi, E.; Salcedo-Mahiques, E.; Cuesta-Peredó, D.; Doménech-Pascual, J.R.; Salvador-Pérez, M.I.; Avellana-Zaragoza, J.A. Early interdisciplinary hospital intervention for elderly patients with hip fractures–functional outcome and mortality. Clinics 2012, 67, 547–555. [Google Scholar] [CrossRef]
- Guzon-Illescas, O.; Fernandez, E.P.; Villarias, N.C.; Donate, F.J.Q.; Peña, M.; Alonso-Blas, C.; Vadillo, A.G.; Mazzucchelli, R. Mortality after osteoporotic hip fracture: Incidence, trends, and associated factors. J. Orthop. Surg. Res. 2019, 14, 203. [Google Scholar] [CrossRef]
- Pugely, A.; Martin, C.T.; Gao, Y.; Klocke, N.F.; Callaghan, J.J.; Marsh, J. A Risk Calculator for Short-Term Morbidity and Mortality After Hip Fracture Surgery. J. Orthop. Trauma 2014, 28, 63–69. [Google Scholar] [CrossRef]
- Clynes, M.A.; Harvey, N.C.; Curtis, E.M.; Fuggle, N.R.; Dennison, E.M.; Cooper, C. The epidemiology of osteoporosis. Br. Med. Bull. 2020, 133, 105–117. [Google Scholar] [CrossRef]
- Brown, J.P.; Adachi, J.D.; Schemitsch, E.; Tarride, J.-E.; Brown, V.; Bell, A.; Reiner, M.; Oliveira, T.; Motsepe-Ditshego, P.; Burke, N.; et al. Mortality in older adults following a fragility fracture: Real-world retrospective matched-cohort study in Ontario. BMC Musculoskelet. Disord. 2021, 22, 105. [Google Scholar] [CrossRef]
- Omsland, T.K.; Emaus, N.; Tell, G.S.; Magnus, J.H.; Ahmed, L.A.; Holvik, K.; Center, J.; Forsmo, S.; Gjesdal, C.G.; Schei, B.; et al. Mortality following the first hip fracture in Norwegian women and men (1999–2008). A NOREPOS study. Bone 2014, 63, 81–86. [Google Scholar] [CrossRef]
- Kanis, J.A.; Johansson, H.; Odén, A.; Harvey, N.C.; Gudnason, V.; Sanders, K.M.; Sigurdsson, G.; Siggeirsdottir, K.; Fitzpatrick, L.A.; Borgström, F.; et al. Characteristics of recurrent fractures. Osteoporos. Int. 2018, 29, 1747–1757. [Google Scholar] [CrossRef]
- Johansson, H.; Siggeirsdóttir, K.; Harvey, N.; Odén, A.; Gudnason, V.; McCloskey, E.; Sigurdsson, G.; Kanis, J.A. Imminent risk of fracture after fracture. Osteoporos. Int. 2016, 28, 775–780. [Google Scholar] [CrossRef]
- Balasubramanian, A.; Zhang, J.; Chen, L.; Wenkert, D.; Daigle, S.G.; Grauer, A.; Curtis, J.R. Risk of subsequent fracture after prior fracture among older women. Osteoporos. Int. 2018, 30, 79–92. [Google Scholar] [CrossRef]
- Dyer, S.M.; Crotty, M.; Fairhall, N.; Magaziner, J.; Beaupre, L.A.; Cameron, I.D.; Sherrington, C. A critical review of the long-term disability outcomes following hip fracture. BMC Geriatr. 2016, 16, 158. [Google Scholar] [CrossRef]
- Cosman, F.; De Beur, S.J.; LeBoff, M.S.; Lewiecki, E.M.; Tanner, B.; Randall, S.; Lindsay, R. Clinician’s Guide to Prevention and Treatment of Osteoporosis. Osteoporos. Int. 2014, 25, 2359–2381. [Google Scholar] [CrossRef]
- Hodsman, A.; Papaioannou, A.; Cranney, A. Clinical practice guidelines for the use of parathyroid hormone in the treatment of osteoporosis. Can. Med. Assoc. J. 2006, 175, 48. [Google Scholar] [CrossRef]
- Ilesanmi-Oyelere, B.L.; Kruger, M.C. Nutrient and Dietary Patterns in Relation to the Pathogenesis of Postmenopausal Osteoporosis—A Literature Review. Life 2020, 10, 220. [Google Scholar] [CrossRef] [PubMed]
- Sahni, S.; Mangano, K.M.; McLean, R.R.; Hannan, M.T.; Kiel, D.P. Dietary Approaches for Bone Health: Lessons from the Framingham Osteoporosis Study. Curr. Osteoporos. Rep. 2015, 13, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Omer, M.; Ali, H.; Orlovskaya, N.; Ballesteros, A.; Cheong, V.S.; Martyniak, K.; Wei, F.; Collins, B.E.; Yarmolenko, S.N.; Asiatico, J.; et al. Omega-9 Modifies Viscoelasticity and Augments Bone Strength and Architecture in a High-Fat Diet-Fed Murine Model. Nutrients 2022, 14, 3165. [Google Scholar] [CrossRef] [PubMed]
- Lau, B.Y.; Fajardo, V.A.; McMeekin, L.; Sacco, S.M.; Ward, W.E.; Roy, B.D.; Peters, S.J.; LeBlanc, P.J. Influence of high-fat diet from differential dietary sources on bone mineral density, bone strength, and bone fatty acid composition in rats. Appl. Physiol. Nutr. Metab. 2010, 35, 598–606. [Google Scholar] [CrossRef] [PubMed]
- Farahnak, Z.; Freundorfer, M.T.; Lavery, P.; Weiler, H.A. Dietary docosahexaenoic acid contributes to increased bone mineral accretion and strength in young female Sprague-Dawley rats. Prostaglandins Leukot. Essent. Fat. Acids 2019, 144, 32–39. [Google Scholar] [CrossRef]
- Maíra, D.C.D.A.; Letícia, R.P.; da Costa, L.R.; Boueri, B.F.D.C.; Carolina, R.P.; Pereira, A.D.; Ribeiro, D.C.; da Silva, E.M.; da Costa, C.A.S.; Gilson, T.B. Flaxseed (Linum usitatissimum) flour contributes to bone health in adult male rats. Nutrition 2018, 49, 48–50. [Google Scholar] [CrossRef]
- Weiler, H.; Kovacs, H.; Nitschmann, E.; Bankovic-Calic, N.; Aukema, H.; Ogborn, M. Feeding flaxseed oil but not secoisolariciresinol diglucoside results in higher bone mass in healthy rats and rats with kidney disease. Prostaglandins Leukot. Essent. Fat. Acids 2007, 76, 269–275. [Google Scholar] [CrossRef]
- Korotkova, M.; Ohlsson, C.; Hanson, L.; Strandvik, B. Dietary n-6: n-3 fatty acid ratio in the perinatal period affects bone parameters in adult female rats. Br. J. Nutr. 2004, 92, 643–648. [Google Scholar] [CrossRef]
- Salari, P.; Rezaie, A.; Larijani, B.; Abdollahi, M. A systematic review of the impact of n-3 fatty acids in bone health and osteoporosis. Med. Sci. Monit. 2008, 14, RA37–RA44. [Google Scholar]
- Farina, E.K.; Kiel, D.P.; Roubenoff, R.; Schaefer, E.J.; Cupples, L.A.; Tucker, K.L. Dietary Intakes of Arachidonic Acid and α-Linolenic Acid Are Associated with Reduced Risk of Hip Fracture in Older Adults. J. Nutr. 2011, 141, 1146–1153. [Google Scholar] [CrossRef]
- Mangano, K.M.; Sahni, S.; Kerstetter, J.E.; Kenny, A.M.; Hannan, M.T. Polyunsaturated Fatty Acids and Their Relation with Bone and Muscle Health in Adults. Curr. Osteoporos. Rep. 2013, 11, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Järvinen, R.; Tuppurainen, M.; Erkkilä, A.T.; Penttinen, P.; Kärkkäinen, M.; Salovaara, K.; Jurvelin, J.S.; Kröger, H. Associations of dietary polyunsaturated fatty acids with bone mineral density in elderly women. Eur. J. Clin. Nutr. 2012, 66, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Farina, E.K.; Kiel, D.P.; Roubenoff, R.; Schaefer, E.J.; Cupples, L.A.; Tucker, K.L. Plasma phosphatidylcholine concentrations of polyunsaturated fatty acids are differentially associated with hip bone mineral density and hip fracture in older adults: The framingham osteoporosis study. J. Bone Miner. Res. 2012, 27, 1222–1230. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Ramírez, M.J.; Palma, S.; A Martínez-González, M.; Delgado-Martínez, A.D.; de la Fuente, C.; Delgado-Rodríguez, M. Dietary fat intake and the risk of osteoporotic fractures in the elderly. Eur. J. Clin. Nutr. 2007, 61, 1114–1120. [Google Scholar] [CrossRef]
- Orchard, T.S.; Ing, S.W.; Lu, B.; Belury, M.A.; Johnson, K.; Wactawski-Wende, J.; Jackson, R.D. The association of red blood cell n-3 and n-6 fatty acids with bone mineral density and hip fracture risk in the women’s health initiative. J. Bone Miner. Res. 2013, 28, 505–515. [Google Scholar] [CrossRef]
- Virtanen, J.K.; Mozaffarian, D.; Cauley, J.A.; Mukamal, K.J.; Robbins, J.; Siscovick, D.S. Fish consumption, bone mineral density, and risk of hip fracture among older adults: The cardiovascular health study. J. Bone Miner. Res. 2010, 25, 1972–1979. [Google Scholar] [CrossRef]
- Dahir, N.S.; Calder, A.N.; McKinley, B.J.; Liu, Y.; Gilbertson, T.A. Sex differences in fat taste responsiveness are modulated by estradiol. Am. J. Physiol. Metab. 2021, 320, E566–E580. [Google Scholar] [CrossRef]
- Jamsa, T.; Jalovaara, P.; Peng, Z.; Väänänen, H.; Tuukkanen, J. Comparison of three-point bending test and peripheral quantitative computed tomography analysis in the evaluation of the strength of mouse femur and tibia. Bone 1998, 23, 155–161. [Google Scholar] [CrossRef]
- Schriefer, J.L.; Robling, A.G.; Warden, S.J.; Fournier, A.J.; Mason, J.J.; Turner, C.H. A comparison of mechanical properties derived from multiple skeletal sites in mice. J. Biomech. 2005, 38, 467–475. [Google Scholar] [CrossRef]
- Deckard, C.; Walker, A.; Hill, B.J.F. Using three-point bending to evaluate tibia bone strength in ovariectomized young mice. J. Biol. Phys. 2017, 43, 139–148. [Google Scholar] [CrossRef]
- Cheong, V.S.; Roberts, B.C.; Kadirkamanathan, V.; Dall’Ara, E. Positive interactions of mechanical loading and PTH treatments on spatio-temporal bone remodelling. Acta Biomater. 2021, 136, 291–305. [Google Scholar] [CrossRef]
- Wolfram, U.; Schwiedrzik, J. Post-yield and failure properties of cortical bone. BoneKEy Rep. 2016, 5, 829. [Google Scholar] [CrossRef]
- He, W.; Goodkind, D.; Kowal, P. An Aging World: 2015; U.S. Government Printing Office: Washington, DC, USA, 2016.
- Colón, C.J.P.; Molina-Vicenty, I.L.; Frontera-Rodríguez, M.; García-Ferré, A.; Rivera, B.P.; Cintrón-Vélez, G.; Frontera-Rodríguez, S. Muscle and Bone Mass Loss in the Elderly Population: Advances in diagnosis and treatment. J. Biomed. 2018, 3, 40–49. [Google Scholar] [CrossRef]
- Föger-Samwald, U.; Dovjak, P.; Azizi-Semrad, U.; Kerschan-Schindl, K.; Pietschmann, P. Osteoporosis: Pathophysiology and therapeutic options. EXCLI J. 2020, 19, 1017–1037. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.; Bradford, C. Omega-3, Omega-6 and Omega-9 Fatty Acids: Implications for Cardiovascular and Other Diseases. J. Glycom. Lipidom. 2014, 4, 1–8. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, X.; Shan, Y. LC-MS-based plasma metabolomics reveals metabolic variations in ovariectomy-induced osteoporosis in female Wistar rats. RSC Adv. 2018, 8, 24932–24941. [Google Scholar] [CrossRef] [PubMed]
- Trichopoulou, A.; Georgiou, E.; Bassiakos, Y.; Lipworth, L.; Lagiou, P.; CharalambosProukakis, M.D.P.; Trichopoulos, D. Energy Intake and Monounsaturated Fat in Relation to Bone Mineral Density among Women and Men in Greece. Prev. Med. 1997, 26, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Dellatore, P.; Douard, V.; Qin, L.; Watford, M.; Ferraris, R.P.; Lin, T.; Shapses, S.A. High fat diet enriched with saturated, but not monounsaturated fatty acids adversely affects femur, and both diets increase calcium absorption in older female mice. Nutr. Res. 2016, 36, 742–750. [Google Scholar] [CrossRef]
- Varela-López, A.; Ochoa, J.J.; Llamas-Elvira, J.M.; López-Frías, M.; Planells, E.; Speranza, L.; Battino, M.; Quiles, J.L. Loss of Bone Mineral Density Associated with Age in Male Rats Fed on Sunflower Oil Is Avoided by Virgin Olive Oil Intake or Coenzyme Q Supplementation. Int. J. Mol. Sci. 2017, 18, 1397. [Google Scholar] [CrossRef]
- Leeners, B.; Geary, N.; Tobler, P.N.; Asarian, L. Ovarian hormones and obesity. Hum. Reprod. Update 2017, 23, 300–321. [Google Scholar] [CrossRef]
- Huang, K.-P.; Raybould, H.E. Estrogen and gut satiety hormones in vagus-hindbrain axis. Peptides 2020, 133, 170389. [Google Scholar] [CrossRef] [PubMed]
- Rogers, N.H.; Perfield, J.W.; Strissel, K.J.; Obin, M.S.; Greenberg, A.S. Reduced Energy Expenditure and Increased Inflammation Are Early Events in the Development of Ovariectomy-Induced Obesity. Endocrinology 2009, 150, 2161–2168. [Google Scholar] [CrossRef] [PubMed]
- Geary, N.; Asarian, L.; Korach, K.S.; Pfaff, N.W.; Ogawa, S. Deficits in E2-Dependent Control of Feeding, Weight Gain, and Cholecystokinin Satiation in ER-α Null Mice. Endocrinology 2001, 142, 4751–4757. [Google Scholar] [CrossRef] [PubMed]
- Roesch, D.M. Effects of selective estrogen receptor agonists on food intake and body weight gain in rats. Physiol. Behav. 2006, 87, 39–44. [Google Scholar] [CrossRef]
- Huang, K.-P.; Ronveaux, C.C.; Knotts, T.A.; Rutkowsky, J.R.; Ramsey, J.J.; Raybould, H.E. Sex differences in response to short-term high fat diet in mice. Physiol. Behav. 2020, 221, 112894. [Google Scholar] [CrossRef]
- Jones, M.E.; Thorburn, A.W.; Britt, K.L.; Hewitt, K.N.; Wreford, N.G.; Proietto, J.; Oz, O.K.; Leury, B.J.; Robertson, K.M.; Yao, S.; et al. Aromatase-deficient (ArKO) mice have a phenotype of increased adiposity. Proc. Natl. Acad. Sci. USA 2000, 97, 12735–12740. [Google Scholar] [CrossRef]
- Burstein, A.H.; Zika, J.M.; Heiple, K.G.; Klein, L. Contribution of collagen and mineral to the elastic-plastic properties of bone. J. Bone Jt. Surg. 1975, 57, 956–961. [Google Scholar] [CrossRef]
- Rochdi, A.; Foucat, L.; Renou, J.P. Effect of thermal denaturation on water-collagen interactions: NMR relaxation and differential scanning calorimetry analysis. Biopolymers 1999, 50, 690–696. [Google Scholar] [CrossRef]
- Linde, F. Elastic and viscoelastic properties of trabecular bone by a compression testing approach. Dan. Med. Bull. 1994, 41, 119–138. [Google Scholar]
- Yamashita, J.; Furman, B.R.; Rawls, H.R.; Wang, X.; Agrawal, C.M. The use of dynamic mechanical analysis to assess the viscoelastic properties of human cortical bone. J. Biomed. Mater. Res. 2001, 58, 47–53. [Google Scholar] [CrossRef]
- Dong, X.N.; Yeni, Y.N.; Les, C.M.; Fyhrie, D.P. Effects of end boundary conditions and specimen geometry on the viscoelastic properties of cancellous bone measured by dynamic mechanical analysis. J. Biomed. Mater. Res. 2004, 68A, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Robinovitch, S.; Hayes, W.C.; McMahon, T.A. Prediction of Femoral Impact Forces in Falls on the Hip. J. Biomech. Eng. 1991, 113, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Turner, C.H. Biomechanics of Bone: Determinants of Skeletal Fragility and Bone Quality. Osteoporos. Int. 2002, 13, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.; Lakes, R.S.; Lal, A. Resonant ultrasound spectroscopy for measurement of mechanical damping: Comparison with broadband viscoelastic spectroscopy. Rev. Sci. Instrum. 2000, 71, 2855–2861. [Google Scholar] [CrossRef]
- Raisz, L.G. Physiology and pathophysiology of bone remodeling. Clin. Chem. 1999, 45, 1353–1358. [Google Scholar]
- Les, C.M.; Vance, J.L.; Christopherson, G.T.; Turner, A.S.; Divine, G.W.; Fyhrie, D.P. Long-term ovariectomy decreases ovine compact bone viscoelasticity. J. Orthop. Res. 2005, 23, 869–876. [Google Scholar] [CrossRef]
- Kim, D.-G.; Huja, S.S.; Lee, H.R.; Tee, B.C.; Hueni, S. Relationships of Viscosity with Contact Hardness and Modulus of Bone Matrix Measured by Nanoindentation. J. Biomech. Eng. 2010, 132, 024502. [Google Scholar] [CrossRef]
- Yang, X.; Muthukumaran, P.; DasDe, S.; Teoh, S.H.; Choi, H.; Lim, S.-K.; Lee, T. Positive alterations of viscoelastic and geometric properties in ovariectomized rat femurs with concurrent administration of ibandronate and PTH. Bone 2013, 52, 308–317. [Google Scholar] [CrossRef]
- Yang, X.; Teoh, S.-H.; DasDe, S.; Lee, T. Administration of PTH and ibandronate increases ovariectomized rat compact bone viscoelasticity. J. Mech. Behav. Biomed. Mater. 2013, 22, 51–58. [Google Scholar] [CrossRef]
- Wen, X.-X.; Wang, F.-Q.; Xu, C.; Wu, Z.-X.; Zhang, Y.; Feng, Y.-F.; Yan, Y.-B.; Lei, W. Time Related Changes of Mineral and Collagen and Their Roles in Cortical Bone Mechanics of Ovariectomized Rabbits. PLoS ONE 2015, 10, e0127973. [Google Scholar] [CrossRef]
- Ozasa, R.; Ishimoto, T.; Miyabe, S.; Hashimoto, J.; Hirao, M.; Yoshikawa, H.; Nakano, T. Osteoporosis Changes Collagen/Apatite Orientation and Young’s Modulus in Vertebral Cortical Bone of Rat. Calcif. Tissue Int. 2019, 104, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Yu, X.; Yao, Q.; Qin, J. Early effects of ovariectomy on bone microstructure, bone turnover markers and mechanical properties in rats. BMC Musculoskelet. Disord. 2022, 23, 316. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Palomar, I.; Shipov, A.; Shahar, R.; Barber, A.H. Mechanical Behavior of Osteoporotic Bone at Sub-Lamellar Length Scales. Front. Mater. 2015, 2, 9. [Google Scholar] [CrossRef]
- Currey, J.D. Bones; Princeton University Press: Princeton, NJ, USA, 2002. [Google Scholar]
- Carter, D.R.; Hayes, W.C. The compressive behavior of bone as a two-phase porous structure. J. Bone Jt. Surg. 1977, 59, 954–962. [Google Scholar] [CrossRef]
- Oliviero, S.; Roberts, M.; Owen, R.; Reilly, G.C.; Bellantuono, I.; Dall’Ara, E. Non-invasive prediction of the mouse tibia mechanical properties from microCT images: Comparison between different finite element models. Biomech. Model. Mechanobiol. 2021, 20, 941–955. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.B.; Burr, D.B.; Sharkey, N.A.; Fyhrie, D.P. Skeletal Tissue Mechanics; Springer: New York, NY, USA, 2015. [Google Scholar]
- Pearson, O.M.; Lieberman, D.E. The aging of Wolff’s “law”: Ontogeny and responses to mechanical loading in cortical bone. Am. J. Phys. Anthr. 2004, 125, 63–99. [Google Scholar] [CrossRef]
- Bayraktar, H.H.; Morgan, E.F.; Niebur, G.L.; Morris, G.E.; Wong, E.K.; Keaveny, T.M. Comparison of the elastic and yield properties of human femoral trabecular and cortical bone tissue. J. Biomech. 2004, 37, 27–35. [Google Scholar] [CrossRef]
- Burstein, A.H.; Currey, J.D.; Frankel, V.H.; Reilly, D.T. The ultimate properties of bone tissue: The effects of yielding. J. Biomech. 1972, 5, 35–44. [Google Scholar] [CrossRef]
- Salim, M.S.; Salleh, A.F.; Daud, R.; Zhang, D.; Zi, B.; Cui, G.; Ding, H. Bone Fracture Modeling and Analysis by Three Point Bending: A Review. MATEC Web Conf. 2016, 77, 11004. [Google Scholar] [CrossRef]
- Wang, Y.; Von Euw, S.; Fernandes, F.; Cassaignon, S.; Selmane, M.; Laurent, G.; Pehau-Arnaudet, G.; Coelho, C.; Bonhomme-Coury, L.; Giraud-Guille, M.-M.; et al. Water-mediated structuring of bone apatite. Nat. Mater. 2013, 12, 1144–1153. [Google Scholar] [CrossRef]
- Tye, C.; Hunter, G.K.; Goldberg, H.A. Identification of the Type I Collagen-binding Domain of Bone Sialoprotein and Characterization of the Mechanism of Interaction. J. Biol. Chem. 2005, 280, 13487–13492. [Google Scholar] [CrossRef]
- Thurner, P.J.; Chen, C.G.; Ionova-Martin, S.; Sun, L.; Harman, A.; Porter, A.; Ager, J.W.; Ritchie, R.O.; Alliston, T. Osteopontin deficiency increases bone fragility but preserves bone mass. Bone 2010, 46, 1564–1573. [Google Scholar] [CrossRef]
- Hang, F.; Gupta, H.S.; Barber, A.H. Nanointerfacial strength between non-collagenous protein and collagen fibrils in antler bone. J. R. Soc. Interface 2014, 11, 20130993. [Google Scholar] [CrossRef]
- Sroga, G.E.; Vashishth, D. Effects of Bone Matrix Proteins on Fracture and Fragility in Osteoporosis. Curr. Osteoporos. Rep. 2012, 10, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Thurner, P.; Lam, S.; Weaver, J.C.; Morse, D.E.; Hansma, P.K. Localization of Phosphorylated Serine, Osteopontin, and Bone Sialoprotein on Bone Fracture Surfaces. J. Adhes. 2009, 85, 526–545. [Google Scholar] [CrossRef]
- Zimmermann, E.A.; Schaible, E.; Gludovatz, B.; Schmidt, F.N.; Riedel, C.; Krause, M.; Vettorazzi, E.; Acevedo, C.; Hahn, M.; Püschel, K.; et al. Intrinsic mechanical behavior of femoral cortical bone in young, osteoporotic and bisphosphonate-treated individuals in low- and high energy fracture conditions. Sci. Rep. 2016, 6, 21072. [Google Scholar] [CrossRef] [PubMed]
- Vashishth, D.; Gibson, G.; Khoury, J.; Schaffler, M.; Kimura, J.; Fyhrie, D. Influence of nonenzymatic glycation on biomechanical properties of cortical bone. Bone 2001, 28, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.; Gong, H.; Cen, H.; Shi, P.; Zhang, R.; Li, Z.; Bi, X. Influence of non-enzymatic glycation on the mechanical properties of cortical bone. J. Mech. Behav. Biomed. Mater. 2021, 119, 104553. [Google Scholar] [CrossRef] [PubMed]
- Role of collagen and other organics in the mechanical properties of bone. Osteoporos. Int. 2003, 14, 29–36. [CrossRef] [PubMed]
- Zappone, B.; Thurner, P.J.; Adams, J.; Fantner, G.E.; Hansma, P.K. Effect of Ca2+ Ions on the Adhesion and Mechanical Properties of Adsorbed Layers of Human Osteopontin. Biophys. J. 2008, 95, 2939–2950. [Google Scholar] [CrossRef] [PubMed]
- Kasonga, A.E.; Kruger, M.C.; Coetzee, M. Free fatty acid receptor 4-β-arrestin 2 pathway mediates the effects of different classes of unsaturated fatty acids in osteoclasts and osteoblasts. Biochim. Biophys. Acta—Mol. Cell Biol. Lipids 2019, 1864, 281–289. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet Type | Diet | Fat Ratio: (Unsaturated: Saturated) | Total Fat (kcal%) | ω-3% | ω-6% | ω-9% |
|---|---|---|---|---|---|---|
| Control Diet * | D07020902 | 1:1 | 10 | 2.8 | 31.6 | 30.7 |
| High Fat Diet | D12492 | 1:1 | 60 | 1.25 | 14.15 | 44.2 |
| * Control Diet | HFD 50:50 | |||
|---|---|---|---|---|
| g | kcal | g | kcal | |
| Protein (% by weight) | 19.2 | 20 | 26.2 | 20 |
| Carbohydrate (% by weight) | 67.3 | 70 | 26.3 | 20 |
| Fat (% by weight) | 4.3 | 10 | 34.9 | 60 |
| Total kcal | 100 | 100 | ||
| kcal/g | 3.85 | 5.24 | ||
| Ingredients | ||||
| Protein Casein, 80 Mesh | 200 | 800 | 200 | 800 |
| L-Cystine | 3 | 12 | 3 | 12 |
| Carbohydrate Corn Starch Maltodextrin 10 Sucrose Cellulose, BW200 | 500 100 100 50 | 2000 400 400 0 | 0 125 68.8 50 | 0 500 275 0 |
| Lipid Soybean Oil Lard CoconutOil, Hydrogenated Safflower Oil Cocoa Butter | 10 5 0 0 30 | 90 45 0 0 270 | 25 245 0 0 0 | 225 2205 0 0 0 |
| Mineral Mix S10026 Dicalcium phosphate Calcium carbonate Potassium citrate, 1 H2O | 13 5.5 16.5 | 0 0 0 | 13 5.5 16.5 | 0 0 0 |
| Vitamin Mix V10001 ** Choline Bitartrate | 10 2 | 40 0 | 10 2 | 40 0 |
| FD&C Yellow Dye #5 FD&C Red Dye #40 FD&C Blue Dye #1 | 0 0 0 | 0 0 0 | 0 0 0.05 | 0 0 0 |
| Total | 1055 | 4057 | 773.85 | 4057 |
| Variables | Control | OVX | OVX + E2 |
|---|---|---|---|
| Starting body weight (g) | 18.92 ± 0.32 | 25.41 ± 0.49 | 20.96 ± 0.23 |
| Final body weight (g) | 28.05 ± 1.10 | 45.09 ± 1.14 | 27.32 ± 0.35 |
| Total cumulative gain in body weight (g) | 9.13 | 20.49 | 6.36 |
| Study Week | Body Weight (BW) | Group Comparison | p-Value * p < 0.05 | Weight Gain (WG) | p-Value * p < 0.05 |
|---|---|---|---|---|---|
| 1 | BW | Control vs. OVX Control vs. OVX + E2 OVX vs. OVX + E2 | 0.004 * 0.002 * 0.002 * | WG | - - - |
| 2 | BW | Control vs. OVX Control vs. OVX + E2 OVX vs. OVX + E2 | 0.004 * 0.003 * 0.002 * | WG | 0.004 * 0.366 0.0028 * |
| 3 | BW | Control vs. OVX Control vs. OVX + E2 OVX vs. OVX + E2 | 0.004 * 0.003 * 0.002 * | WG | 0.004 * 0.605 0.005 * |
| 4 | BW | Control vs. OVX Control vs. OVX + E2 OVX vs. OVX + E2 | 0.004 * 0.010 * 0.002 * | WG | 0.004 * 0.093 0.002 * |
| 5 | BW | Control vs. OVX Control vs. OVX + E2 OVX vs. OVX + E2 | 0.004 * 0.002 * 0.002 * | WG | 0.103 0.121 0.002 * |
| 6 | BW | Control vs. OVX Control vs. OVX + E2 OVX vs. OVX + E2 | 0.004 * 0.028 * 0.002 * | WG | 0.014 * 0.109 0.003 * |
| 7 | BW | Control vs. OVX Control vs. OVX + E2 OVX vs. OVX + E2 | 0.004 * 0.196 0.002 * | WG | 0.004 * 0.039 0.002 * |
| 8 | BW | Control vs. OVX Control vs. OVX + E2 OVX vs. OVX + E2 | 0.004 * 0.796 0.002 * | WG | 0.006 * 0.024 0.002 * |
| 9 | BW | Control vs. OVX Control vs. OVX + E2 OVX vs. OVX + E2 | 0.004 * 0.796 0.002 * | WG | 0.078 0.606 0.020 * |
| 10 | BW | Control vs. OVX Control vs. OVX + E2 OVX vs. OVX + E2 | 0.004 * 1.000 0.002 * | WG | 0.337 0.302 0.302 |
| 11 | BW | Control vs. OVX Control vs. OVX + E2 OVX vs. OVX + E2 | 0.004 * 0.606 0.002 * | WG | 0.037 * 0.121 0.004 * |
| 12 | BW | Control vs. OVX Control vs. OVX + E2 OVX vs. OVX + E2 | 0.004 * 0.518 0.002 * | WG | 0.200 0.897 0.078 |
| Tibial Variables | Control | OVX | OVX + E2 |
|---|---|---|---|
| Total area (mm2) | 1.10 ± 0.04 | 0.84 ± 0.01 | 1.13 ± 0.11 |
| Outer diameter (mm) | 1.46 ± 0.03 | 1.37 ± 0.01 | 1.39 ± 0.08 |
| Inner diameter (mm) | 0.86 ± 0.19 | 0.90 ± 0.009 | 0.72 ± 0.03 |
| Moment of inertia (mm4) | 0.20 ± 0.02 | 0.14 ± 0.006 | 0.18 ± 0.04 |
| Length (mm) | 18.82 ± 0.45 | 21.30 ± 0.35 | 19.64 ± 0.87 |
| Whole Tibial Bone Mineral and Volume Levels (Range) | |||
|---|---|---|---|
| Control | OVX | OVX + E2 | |
| BMD (g.cm3) | 0.461 (0.310–0.673) | 0.422 (0.267–0.623) | 0.585 (0.390–0.844) |
| BMC (g)(sum) | 7.327 | 6.749 | 10.010 |
| BV/TV | 0.547 (0.253–0.732) | 0.490 (0.241–0.646) | 0.689 (0.514–0.853) |
| Whole tibial bone mineral and volume levels (range) | |||
| Control | OVX | OVX + E2 | |
| BMD (g.cm3) | 0.461 (0.310–0.673) | 0.422 (0.267–0.623) | 0.585 (0.390–0.844) |
| BMC (g)(sum) | 7.327 | 6.749 | 10.010 |
| BV/TV | 0.547 (0.253–0.732) | 0.490 (0.241–0.646) | 0.689 (0.514–0.853) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Omer, M.; Ngo, C.; Ali, H.; Orlovskaya, N.; Cheong, V.S.; Ballesteros, A.; Garner, M.T.; Wynn, A.; Martyniak, K.; Wei, F.; et al. The Effect of Omega-9 on Bone Viscoelasticity and Strength in an Ovariectomized Diet-Fed Murine Model. Nutrients 2023, 15, 1209. https://doi.org/10.3390/nu15051209
Omer M, Ngo C, Ali H, Orlovskaya N, Cheong VS, Ballesteros A, Garner MT, Wynn A, Martyniak K, Wei F, et al. The Effect of Omega-9 on Bone Viscoelasticity and Strength in an Ovariectomized Diet-Fed Murine Model. Nutrients. 2023; 15(5):1209. https://doi.org/10.3390/nu15051209
Chicago/Turabian StyleOmer, Mahmoud, Christopher Ngo, Hessein Ali, Nina Orlovskaya, Vee San Cheong, Amelia Ballesteros, Michael Tyrel Garner, Austin Wynn, Kari Martyniak, Fei Wei, and et al. 2023. "The Effect of Omega-9 on Bone Viscoelasticity and Strength in an Ovariectomized Diet-Fed Murine Model" Nutrients 15, no. 5: 1209. https://doi.org/10.3390/nu15051209
APA StyleOmer, M., Ngo, C., Ali, H., Orlovskaya, N., Cheong, V. S., Ballesteros, A., Garner, M. T., Wynn, A., Martyniak, K., Wei, F., Collins, B. E., Yarmolenko, S. N., Asiatico, J., Kinzel, M., Ghosh, R., Meckmongkol, T., Calder, A., Dahir, N., Gilbertson, T. A., ... Coathup, M. (2023). The Effect of Omega-9 on Bone Viscoelasticity and Strength in an Ovariectomized Diet-Fed Murine Model. Nutrients, 15(5), 1209. https://doi.org/10.3390/nu15051209