
Alleviation of Porphyromonas gingivalis or Its Extracellular Vesicles Provoked Periodontitis and Cognitive Impairment by Lactobacillus pentosus NK357 and Bifidobacterium bifidum NK391

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Culture of PG
2.3. Preparation of pEVs
2.4. Culture of NK357 and NK391
2.5. Culture of SH-SY5Y Cells and BV2 Cells
2.6. Animals
2.7. Determination of Biomarkers
2.8. Immunofluorescence Assay
2.9. Microbiota 16S rRNA Sequencing
2.10. Quantitative Real-Time Polymerase Chain Reaction (qPCR)
2.11. Statistical Analysis
3. Results
3.1. NK357 and NK391 Decreased PG- or pEVs-Induced TNF-α Expression in BV2 Cells and Increased PG- or pEVs-Suppressed BDNF Expression in SH-SY5Y Cells
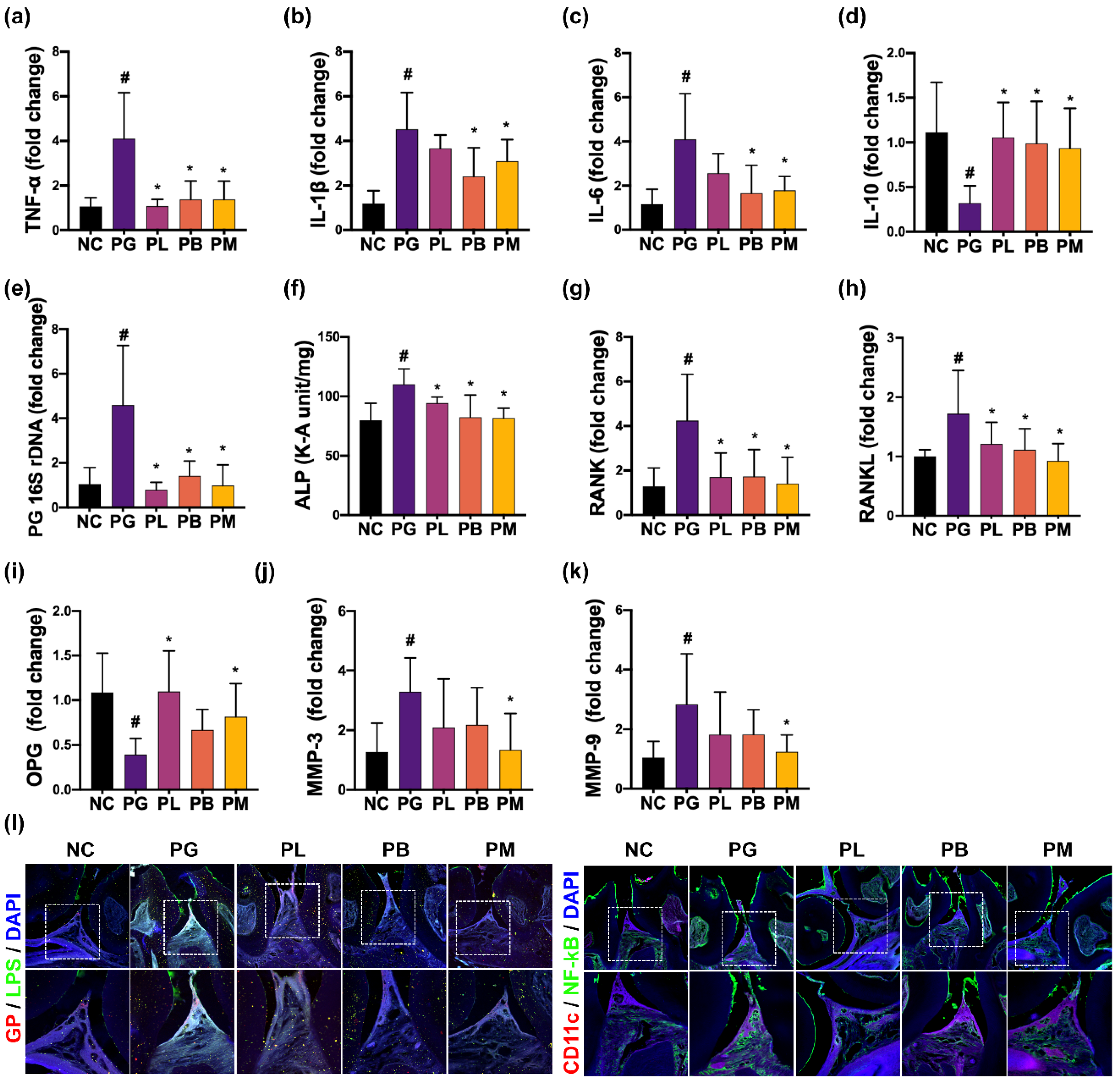
3.2. NK357 and NK391 Alleviated PG-Induced Periodontitis in Mice
3.3. NK357 and NK391 Alleviated PG-Induced CI in Mice
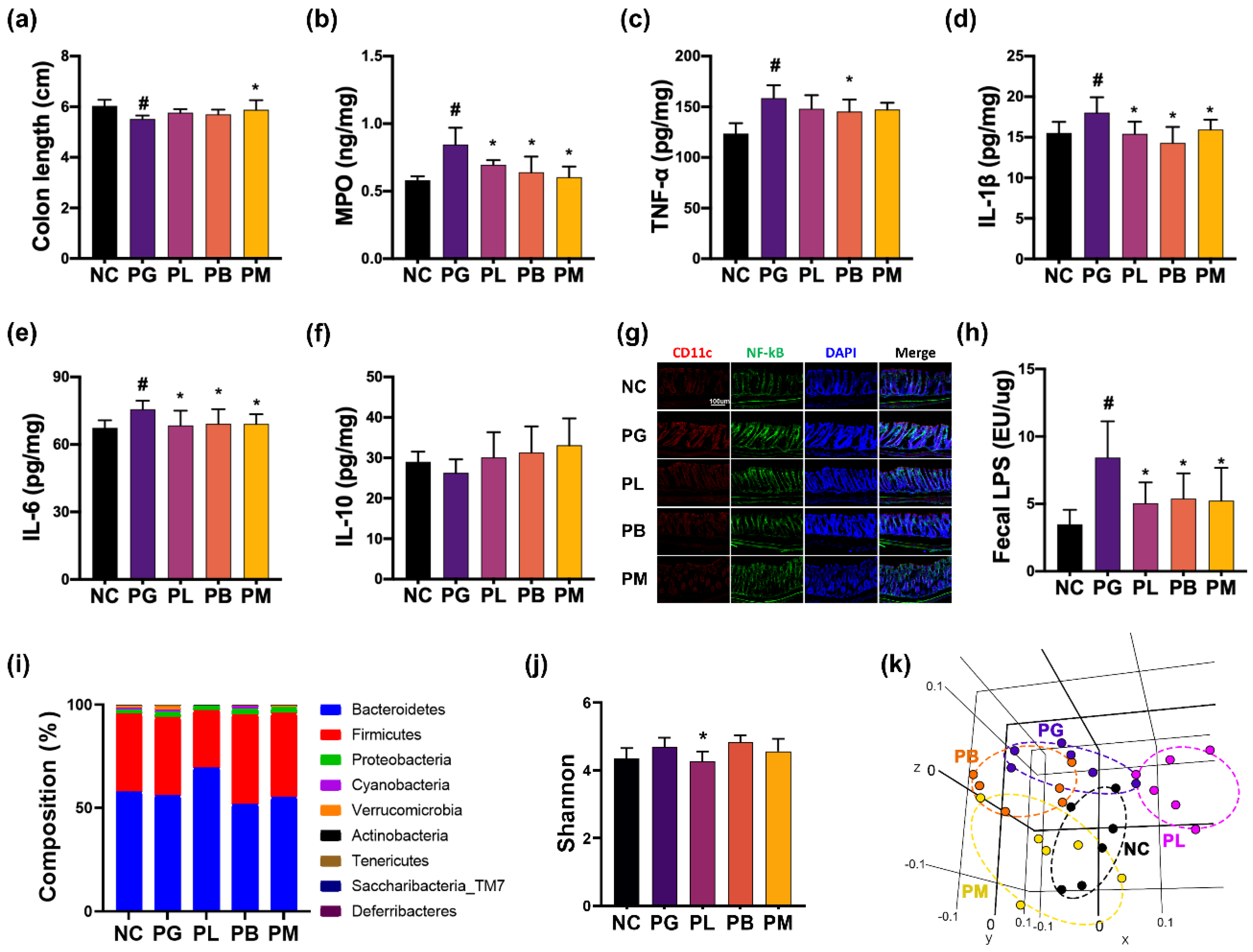
3.4. NK357 and NK391 Alleviated PG-Induced Colitis and Gut Dysbiosis in Mice
3.5. NKc Alleviated pEVs-Induced Periodontitis, CI, and Colitis in Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- How, K.Y.; Song, K.P.; Chan, K.G. Porphyromonas gingivalis: An overview of periodontopathic pathogen below the gum line. Front Microbiol. 2016, 7, 53. [Google Scholar] [CrossRef] [PubMed]
- Elwishahy, A.; Antia, K.; Bhusari, S.; Ilechukwu, N.C.; Horstick, O.; Winkler, V. Porphyromonas gingivalis as a risk factor to Alzheimer’s disease: A systematic review. J. Alzheimers Dis. Rep. 2021, 5, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, K.; Yamada-Furukawa, M.; Kurosawa, M.; Shikama, Y. Periodontal disease and eriodontal disease-related bacteria involved in the pathogenesis of Alzheimer’s disease. J. Inflamm. Res. 2020, 13, 275–283. [Google Scholar] [CrossRef]
- Guo, H.; Chang, S.; Pi, X.; Hua, F.; Jiang, H.; Liu, C.; Du, M. The effect of periodontitis on dementia and cognitive impairment: A meta-analysis. Int. J. Environ Res. Public Health 2021, 18, 6823. [Google Scholar] [CrossRef] [PubMed]
- Imamura, T. The role of gingipains in the pathogenesis of periodontal disease. J. Periodontol. 2003, 74, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Bahar, B.; Singhrao, S.K. An evaluation of the molecular mode of action of trans-resveratrol in the Porphyromonas gingivalis lipopolysaccharide challenged neuronal cell model. Mol. Biol. Rep. 2021, 48, 147–156. [Google Scholar] [CrossRef]
- Ding, Y.; Ren, J.; Yu, H.; Yu, W.; Zhou, Y. Porphyromonas gingivalis, a periodontitis causing bacterium, induces memory impairment and age-dependent neuroinflammation in mice. Immun. Ageing 2018, 15, 6. [Google Scholar] [CrossRef]
- Zhang, J.; Yu, C.; Zhang, X.; Chen, H.; Dong, J.; Lu, W.; Song, Z.; Zhou, W. Porphyromonas gingivalis lipopolysaccharide induces cognitive dysfunction, mediated by neuronal inflammation via activation of the TLR4 signaling pathway in C57BL/6 mice. J. Neuroinflammation 2018, 15, 37. [Google Scholar] [CrossRef]
- Dominy, S.S.; Lynch, C.; Ermini, F.; Benedyk, M.; Marczyk, A.; Konradi, A.; Nguyen, M.; Haditsch, U.; Raha, D.; Griffin, C.; et al. Porphyromonas gingivalis in Alzheimer’s disease brains: Evidence for disease causation and treatment with small-molecule inhibitors. Sci. Adv. 2019, 5, eaau3333. [Google Scholar] [CrossRef]
- Ryder, M.I. Porphyromonas gingivalis and Alzheimer disease: Recent findings and potential therapies. J. Periodontol. 2020, 91 (Suppl. S1), S45–S49. [Google Scholar] [CrossRef]
- Abraham, B.P.; Quigley, E.M.M. Probiotics in inflammatory bowel disease. Gastroenterol. Clin. N. Am. 2017, 46, 769–782. [Google Scholar] [CrossRef]
- Guo, L.; Xu, J.; Du, Y.; Wu, W.; Nie, W.; Zhang, D.; Luo, Y.; Lu, H.; Lei, M.; Xiao, S.; et al. Effects of gut microbiota and probiotics on Alzheimer’s disease. Transl. Neurosci. 2021, 12, 573–580. [Google Scholar] [CrossRef]
- Borchers, A.T.; Selmi, C.; Meyers, F.J.; Keen, C.L.; Gershwin, M.E. Probiotics and immunity. J. Gastroenterol. 2009, 44, 26–46. [Google Scholar] [CrossRef]
- Sheikhi, A.; Giti, H.; Heibor, M.R.; Jafarzadeh, A.; Shakerian, M.; Baharifar, N.; Niruzad, F.; Moghaddam, A.S.; Kokhaei, P.; Baghaeifar, M. Lactobacilus delbrueckii subsp. bulgaricus modulates the secretion of Th1/Th2 and Treg cell-related cytokines by PBMCs from patients with atopic dermatitis. Drug Res. (Stuttg.) 2017, 67, 724–729. [Google Scholar] [CrossRef] [PubMed]
- Mantegazza, C.; Molinari, P.; D’Auria, E.; Sonnino, M.; Morelli, L.; Zuccotti, G.V. Probiotics and antibiotic-associated diarrhea in children: A review and new evidence on Lactobacillus rhamnosus GG during and after antibiotic treatment. Pharmacol. Res. 2018, 128, 63–72. [Google Scholar] [CrossRef]
- Kim, J.K.; Lee, K.E.; Lee, S.A.; Jang, H.M.; Kim, D.H. Interplay between human gut bacteria Escherichia coli and Lactobacillus mucosae in the occurrence of neuropsychiatric disorders in mice. Front Immunol. 2020, 11, 273. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.Y.; Son, Y.H.; Yoo, J.W.; Joo, M.K.; Kim, D.H. Tight junction protein expression-inducing probiotics alleviate TNBS-induced cognitive impairment with colitis in mice. Nutrients 2022, 14, 2975. [Google Scholar] [CrossRef]
- Lee, H.J.; Lee, K.E.; Kim, J.K.; Kim, D.H. Suppression of gut dysbiosis by Bifidobacterium longum alleviates cognitive decline in 5XFAD transgenic and aged mice. Sci. Rep. 2019, 9, 11814. [Google Scholar] [CrossRef]
- Gatej, S.M.; Marino, V.; Bright, R.; Fitzsimmons, T.R.; Gully, N.; Zilm, P.; Gibson, R.J.; Edwards, S.; Bartold, P.M. Probiotic Lactobacillus rhamnosus GG prevents alveolar bone loss in a mouse model of experimental periodontitis. J. Clin. Periodontol. 2018, 45, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Morales, A.; Carvajal, P.; Silva, N.; Hernandez, M.; Godoy, C.; Rodriguez, G.; Cabello, R.; Garcia-Sesnich, J.; Hoare, A.; Diaz, P.I.; et al. Clinical effects of Lactobacillus rhamnosus in non-surgical treatment of chronic periodontitis: A randomized placebo-controlled trial with 1-year follow-up. J. Periodontol. 2016, 87, 944–952. [Google Scholar] [CrossRef]
- Boutaga, K.; van Winkelhoff, A.J.; Vandenbroucke-Grauls, C.M.; Savelkoul, P.H. Comparison of real-time PCR and culture for detection of Porphyromonas gingivalis in subgingival plaque samples. J. Clin. Microbiol. 2003, 41, 4950–4954. [Google Scholar] [CrossRef] [PubMed]
- Okamura, H.; Hirota, K.; Yoshida, K.; Weng, Y.; He, Y.; Shiotsu, N.; Ikegame, M.; Uchida-Fukuhara, Y.; Tanai, A.; Guo, J. Outer membrane vesicles of Porphyromonas gingivalis: Novel communication tool and strategy. Jpn. Dent. Sci. Rev. 2021, 57, 138–146. [Google Scholar] [CrossRef]
- Jang, H.M.; Lee, K.E.; Kim, D.H. The Preventive and curative effects of Lactobacillus reuteri NK33 and Bifidobacterium adolescentis NK98 on immobilization stress-induced anxiety/depression and colitis in mice. Nutrients 2019, 11, 819. [Google Scholar] [CrossRef]
- Lee, K.E.; Kim, J.K.; Han, S.K.; Lee, D.Y.; Lee, H.J.; Yim, S.V.; Kim, D.H. The extracellular vesicle of gut microbial Paenalcaligenes hominis is a risk factor for vagus nerve-mediated cognitive impairment. Microbiome 2020, 8, 107. [Google Scholar] [CrossRef]
- Jang, S.E.; Lim, S.M.; Jeong, J.J.; Jang, H.M.; Lee, H.J.; Han, M.J.; Kim, D.H. Gastrointestinal inflammation by gut microbiota disturbance induces memory impairment in mice. Mucosal. Immunol. 2018, 11, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Sarter, M.; Bodewitz, G.; Stephens, D.N. Attenuation of scopolamine-induced impairment of spontaneous alteration behaviour by antagonist but not inverse agonist and agonist beta-carbolines. Psychopharmacology (Berl.) 1988, 94, 491–495. [Google Scholar] [CrossRef] [PubMed]
- Rogers, G.B.; Keating, D.J.; Young, R.L.; Wong, M.L.; Licinio, J.; Wesselingh, S. From gut dysbiosis to altered brain function and mental illness: Mechanisms and pathways. Mol. Psychiatry 2016, 21, 738–748. [Google Scholar] [CrossRef]
- Gomaa, E.Z. Human gut microbiota/microbiome in health and diseases: A review. Antonie Van Leeuwenhoek 2020, 113, 2019–2040. [Google Scholar] [CrossRef] [PubMed]
- Fukui, H. Increased intestinal permeability and decreased barrier function: Does it really influence the risk of inflammation? Inflamm. Intest Dis. 2016, 1, 135–145. [Google Scholar] [CrossRef]
- Schietroma, M.; Pessia, B.; Carlei, F.; Mariani, P.; Sista, F.; Amicucci, G. Intestinal permeability and systemic endotoxemia in patients with acute pancreatitis. Ann. Ital. Chir. 2016, 87, 138–144. [Google Scholar] [PubMed]
- Bischoff, S.C.; Barbara, G.; Buurman, W.; Ockhuizen, T.; Schulzke, J.D.; Serino, M.; Tilg, H.; Watson, A.; Wells, J.M. Intestinal permeability--a new target for disease prevention and therapy. BMC Gastroenterol. 2014, 14, 189. [Google Scholar] [CrossRef]
- Jang, H.M.; Lee, K.E.; Lee, H.J.; Kim, D.H. Immobilization stress-induced Escherichia coli causes anxiety by inducing NF-κB activation through gut microbiota disturbance. Sci. Rep. 2018, 8, 13897. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Shin, Y.J.; Kim, J.K.; Jang, H.M.; Joo, M.K.; Kim, D.H. Alleviation of cognitive impairment by gut microbiota lipopolysaccharide production-suppressing Lactobacillus plantarum and Bifidobacterium longum in mice. Food Funct. 2021, 12, 10750–10763. [Google Scholar] [CrossRef] [PubMed]
- Haditsch, U.; Roth, T.; Rodriguez, L.; Hancock, S.; Cecere, T.; Nguyen, M.; Arastu-Kapur, S.; Broce, S.; Raha, D.; Lynch, C.C.; et al. Alzheimer’s disease-like neurodegeneration in Porphyromonas gingivalis infected neurons with persistent expression of active gingipains. J. Alzheimers Dis. 2020, 75, 1361–1376. [Google Scholar] [CrossRef]
- Afonso, P.; De Luca, P.; Carvalho, R.S.; Cortes, L.; Pinheiro, P.; Oliveiros, B.; Almeida, R.D.; Mele, M.; Duarte, C.B. BDNF increases synaptic NMDA receptor abundance by enhancing the local translation of Pyk2 in cultured hippocampal neurons. Sci. Signal. 2019, 12, eaav3577. [Google Scholar] [CrossRef]
- Maqsood, R.; Stone, T.W. The gut-brain axis, BDNF, NMDA and CNS disorders. Neurochem. Res. 2016, 41, 2819–2835. [Google Scholar] [CrossRef] [PubMed]
- Kairisalo, M.; Korhonen, L.; Sepp, M.; Pruunsild, P.; Kukkonen, J.P.; Kivinen, J.; Timmusk, T.; Blomgren, K.; Lindholm, D. NF-kappaB-dependent regulation of brain-derived neurotrophic factor in hippocampal neurons by X-linked inhibitor of apoptosis protein. Eur. J. Neurosci. 2009, 30, 958–966. [Google Scholar] [CrossRef]
- Marini, A.M.; Jiang, X.; Wu, X.; Tian, F.; Zhu, D.; Okagaki, P.; Lipsky, R.H. Role of brain-derived neurotrophic factor and NF-kappaB in neuronal plasticity and survival: From genes to phenotype. Restor. Neurol. Neurosci. 2004, 22, 121–130. [Google Scholar]
- Kim, J.W.; Jung, B.H.; Lee, J.H.; Yoo, K.Y.; Lee, H.; Kang, M.S.; Lee, J.K. Effect of Weissella cibaria on the reduction of periodontal tissue destruction in mice. J. Periodontol. 2020, 91, 1367–1374. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, X.; Yoo, J.-W.; Shin, Y.-J.; Park, H.-S.; Son, Y.-H.; Kim, D.-H. Alleviation of Porphyromonas gingivalis or Its Extracellular Vesicles Provoked Periodontitis and Cognitive Impairment by Lactobacillus pentosus NK357 and Bifidobacterium bifidum NK391. Nutrients 2023, 15, 1068. https://doi.org/10.3390/nu15051068
Ma X, Yoo J-W, Shin Y-J, Park H-S, Son Y-H, Kim D-H. Alleviation of Porphyromonas gingivalis or Its Extracellular Vesicles Provoked Periodontitis and Cognitive Impairment by Lactobacillus pentosus NK357 and Bifidobacterium bifidum NK391. Nutrients. 2023; 15(5):1068. https://doi.org/10.3390/nu15051068
Chicago/Turabian StyleMa, Xiaoyang, Jong-Wook Yoo, Yoon-Jung Shin, Hee-Seo Park, Young-Hoo Son, and Dong-Hyun Kim. 2023. "Alleviation of Porphyromonas gingivalis or Its Extracellular Vesicles Provoked Periodontitis and Cognitive Impairment by Lactobacillus pentosus NK357 and Bifidobacterium bifidum NK391" Nutrients 15, no. 5: 1068. https://doi.org/10.3390/nu15051068
APA StyleMa, X., Yoo, J.-W., Shin, Y.-J., Park, H.-S., Son, Y.-H., & Kim, D.-H. (2023). Alleviation of Porphyromonas gingivalis or Its Extracellular Vesicles Provoked Periodontitis and Cognitive Impairment by Lactobacillus pentosus NK357 and Bifidobacterium bifidum NK391. Nutrients, 15(5), 1068. https://doi.org/10.3390/nu15051068