






Biomarkers of Micronutrients and Phytonutrients and Their Application in Epidemiological Studies
 ,
,
Abstract
1. Introduction
2. Methods
2.1. Information Sources and Search Strategy
2.2. Criteria and Study Selection
2.3. Synthesis of Results
3. Vitamin Biomarker
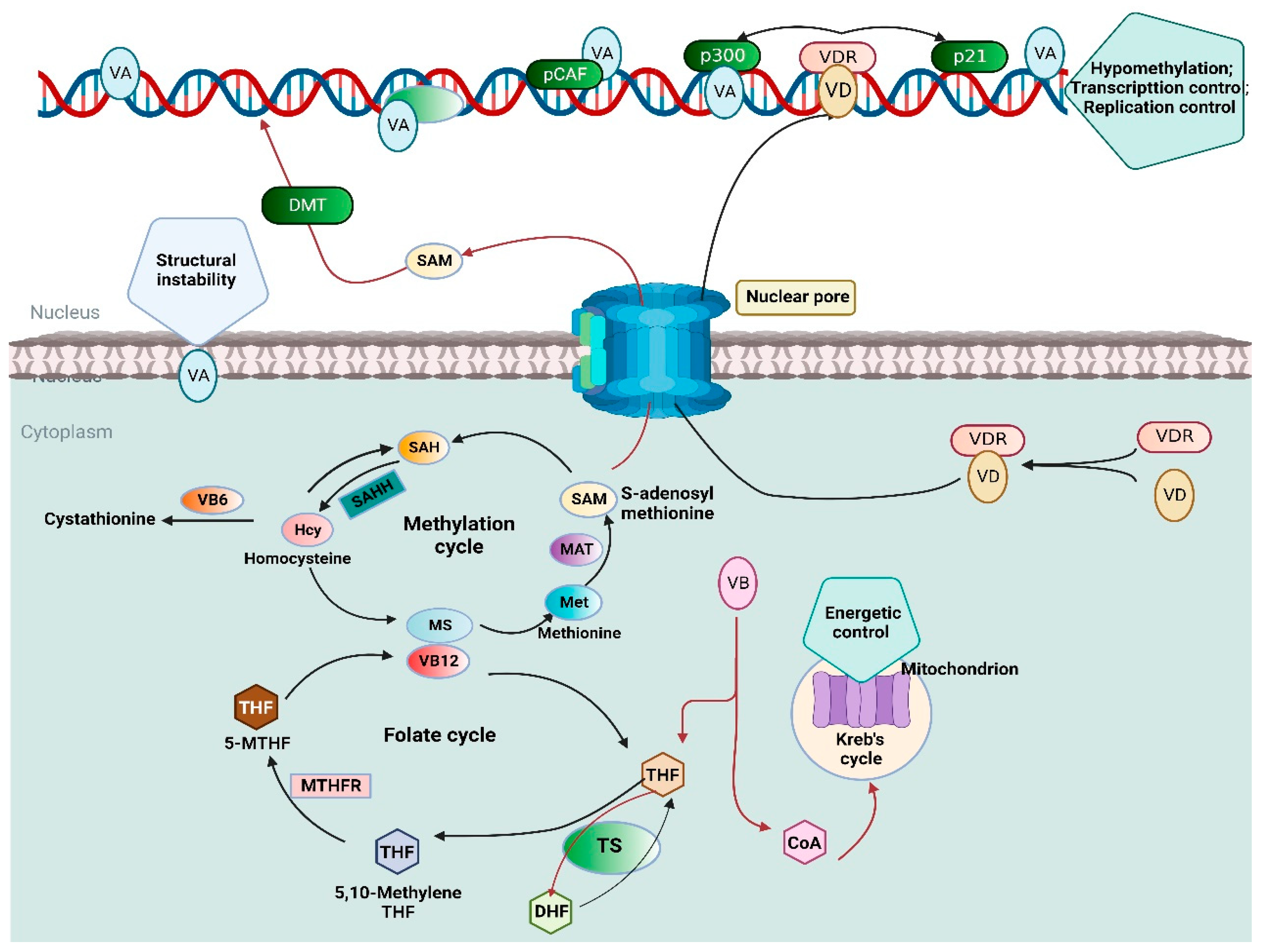
3.1. Vitamin A Biomarker
Application of Vitamin A Biomarkers
3.2. Vitamin D Biomarker
Application of Vitamin D Biomarkers
3.3. Vitamin B9 (Folate) Biomarker
Application of Folate Biomarker
3.4. Vitamin B12 Biomarker
Application of Vitamin B12 Biomarker
4. Mineral Biomarker
4.1. Iodine Biomarker
Application of Iodine Biomarker
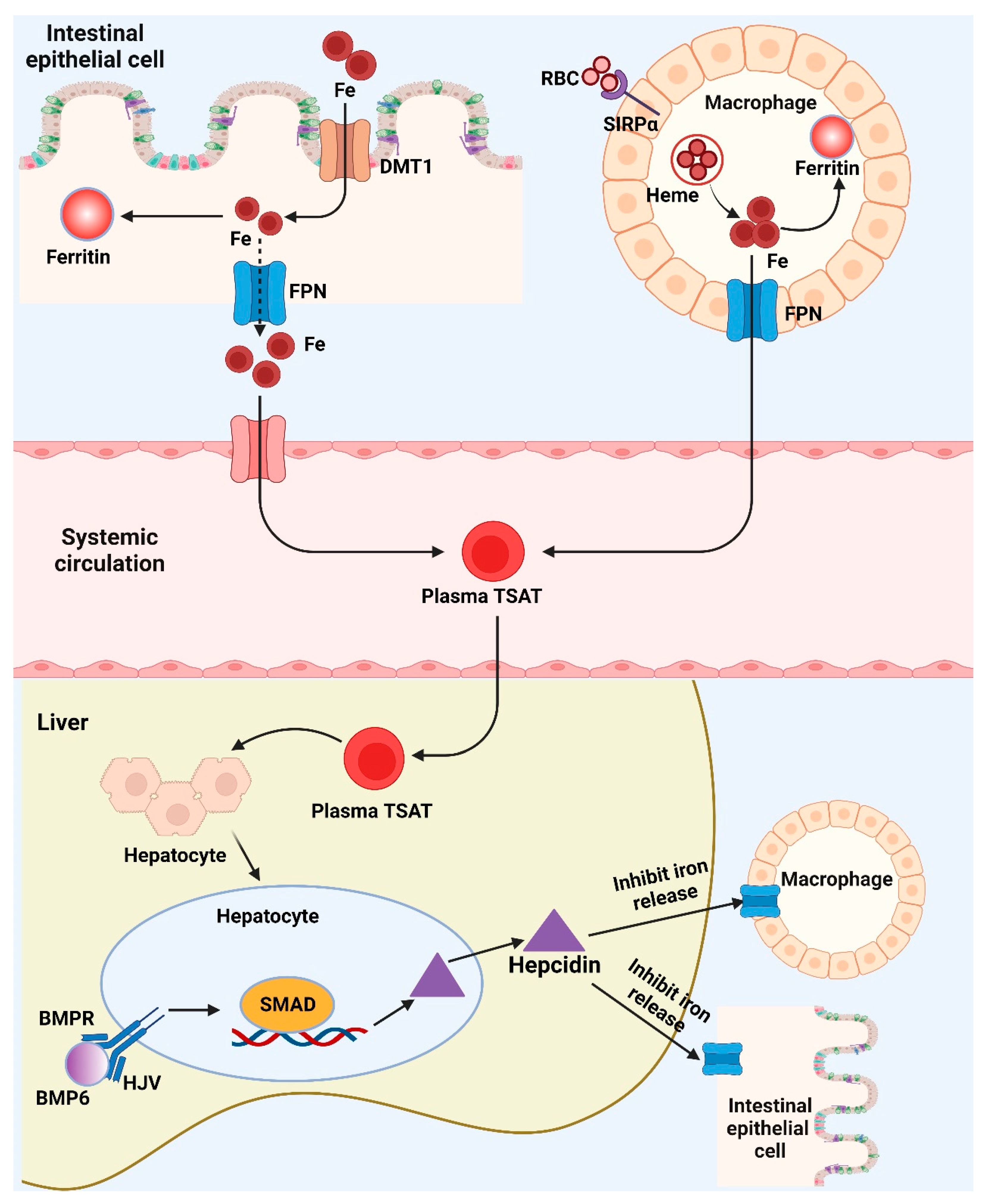
4.2. Iron Biomarker
Application of Iron Biomarker
4.3. Zinc Biomarker
Application of Zinc Biomarker
5. Phytonutrient Biomarker
5.1. Biomarker of Polyphenol and Its Application
5.2. Biomarker of Carotenoid and Its Application
6. Limitations of Current Nutritional Research
7. Conclusions and Prospective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mao, X.Y.; Yin, X.X.; Guan, Q.W.; Xia, Q.X.; Yang, N.; Zhou, H.H.; Liu, Z.Q.; Jin, W.L. Dietary nutrition for neurological disease therapy: Current status and future directions. Pharmacol. Ther. 2021, 226, 107861. [Google Scholar] [CrossRef]
- Lichtenstein, A.H.; Appel, L.J.; Vadiveloo, M.; Hu, F.B.; Kris-Etherton, P.M.; Rebholz, C.M.; Rebholz, C.M.; Sacks, F.M.; Thorndike, A.N.; Van Horn, L.; et al. 2021 Dietary Guidance to Improve Cardiovascular Health: A Scientific Statement from the American Heart Association. Circulation 2021, 144, e472–e487. [Google Scholar] [CrossRef]
- Jun, S.; Cowan, A.E.; Dodd, K.W.; Tooze, J.A.; Gahche, J.J.; Eicher-Miller, H.A.; Guenther, P.M.; Dwyer, J.T.; Potischman, N.; Bhadra, A.; et al. Association of food insecurity with dietary intakes and nutritional biomarkers among US children, National Health and Nutrition Examination Survey (NHANES) 2011-2016. Am. J. Clin. Nutr. 2021, 114, 1059–1069. [Google Scholar] [CrossRef]
- Autier, P. Vitamin D status as a synthetic biomarker of health status. Endocrine 2016, 51, 201–202. [Google Scholar] [CrossRef] [PubMed]
- Nestel, P.J.; Mori, T.A. Dietary patterns, dietary nutrients and cardiovascular disease. Rev. Cardiovasc. Med. 2022, 23, 17. [Google Scholar] [CrossRef]
- Wu, Q.; Gao, Z.J.; Yu, X.; Wang, P. Dietary regulation in health and disease. Signal Transduct. Target. Ther. 2022, 7, 252. [Google Scholar] [CrossRef]
- Schulz, C.A.; Oluwagbemigun, K.; Nothlings, U. Advances in dietary pattern analysis in nutritional epidemiology. Eur. J. Nutr. 2021, 60, 4115–4130. [Google Scholar] [CrossRef]
- Lee, E.C.; Fragala, M.S.; Kavouras, S.A.; Queen, R.M.; Pryor, J.L.; Casa, D.J. Biomarkers in Sports and Exercise: Tracking Health, Performance, and Recovery in Athletes. J. Strength Cond. Res. 2017, 31, 2920–2937. [Google Scholar] [CrossRef]
- Odriozola, L.; Corrales, F.J. Discovery of nutritional biomarkers: Future directions based on omics technologies. Int. J. Food Sci. Nutr. 2015, 66 (Suppl. S1), S31–S40. [Google Scholar] [CrossRef]
- Rubio-Aliaga, I.; Kochhar, S.; Silva-Zolezzi, I. Biomarkers of nutrient bioactivity and efficacy: A route toward personalized nutrition. J. Clin. Gastroenterol. 2012, 46, 545–554. [Google Scholar] [CrossRef]
- Combs, G.F., Jr.; Trumbo, P.R.; McKinley, M.C.; Milner, J.; Studenski, S.; Kimura, T.; Watkins, S.M.; Raiten, D.J. Biomarkers in nutrition: New frontiers in research and application. Ann. N. Y. Acad. Sci. 2013, 1278, 1–10. [Google Scholar] [CrossRef]
- Zebrowska, A.; Sadowska-Krepa, E.; Stanula, A.; Waskiewicz, Z.; Lakomy, O.; Bezuglov, E.; Nikolaidis, P.T.; Rosemann, T.; Knechtle, B. The effect of vitamin D supplementation on serum total 25(OH) levels and biochemical markers of skeletal muscles in runners. J. Int. Soc. Sport. Nutr. 2020, 17, 18. [Google Scholar] [CrossRef]
- Crispin, P. Comment on Ferritin as a functional biomarker of iron status in children and young adults. Br. J. Haematol. 2019, 187, 261–263. [Google Scholar] [CrossRef]
- Raghavan, R.; Ashour, F.S.; Bailey, R. A Review of Cutoffs for Nutritional Biomarkers. Adv. Nutr. 2016, 7, 112–120. [Google Scholar] [CrossRef]
- Allen, L.H.; Miller, J.W.; de Groot, L.; Rosenberg, I.H.; Smith, A.D.; Refsum, H.; Raiten, D.J. Biomarkers of Nutrition for Development (BOND): Vitamin B-12 Review. J. Nutr. 2018, 148 (Suppl. S4), 1995S–2027S. [Google Scholar] [CrossRef]
- Bailey, L.B.; Stover, P.J.; McNulty, H.; Fenech, M.F.; Gregory, J.F., 3rd; Mills, J.L.; Pfeiffer, C.M.; Fazili, Z.; Zhang, M.; Ueland, P.M.; et al. Biomarkers of Nutrition for Development-Folate Review. J. Nutr. 2015, 145, 1636S–1680S. [Google Scholar] [CrossRef] [PubMed]
- King, J.C.; Brown, K.H.; Gibson, R.S.; Krebs, N.F.; Lowe, N.M.; Siekmann, J.H.; Raiten, D.J. Biomarkers of Nutrition for Development (BOND)-Zinc Review. J. Nutr. 2015, 146, 858S–885S. [Google Scholar] [CrossRef]
- Lynch, S.; Pfeiffer, C.M.; Georgieff, M.K.; Brittenham, G.; Fairweather-Tait, S.; Hurrell, R.F.; McArdle, H.J.; Raiten, D.J. Biomarkers of Nutrition for Development (BOND)-Iron Review. J. Nutr. 2018, 148 (Suppl. S1), 1001S–1067S. [Google Scholar] [CrossRef] [PubMed]
- Rohner, F.; Zimmermann, M.; Jooste, P.; Pandav, C.; Caldwell, K.; Raghavan, R.; Raiten, D.J. Biomarkers of nutrition for development--iodine review. J. Nutr. 2014, 144, 1322S–1342S. [Google Scholar] [CrossRef] [PubMed]
- Tanumihardjo, S.A.; Russell, R.M.; Stephensen, C.B.; Gannon, B.M.; Craft, N.E.; Haskell, M.J.; Lietz, G.; Schulze, K.; Raiten, D.J. Biomarkers of Nutrition for Development (BOND)-Vitamin A Review. J. Nutr. 2016, 146, 1816S–1848S. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yang, K.; Chen, L.; Liao, X.; Deng, L.; Chen, S.; Ji, Y. Vitamin A deficiency in critically ill children with sepsis. Crit. Care 2019, 23, 267. [Google Scholar] [CrossRef]
- Colt, S.; Gannon, B.M.; Finkelstein, J.L.; Zambrano, M.P.; Andrade, J.K.; Centeno-Tablante, E.; August, A.; Erickson, D.; Cárdenas, W.B.; Mehta, S. Vitamin A status, inflammation adjustment, and immunologic response in the context of acute febrile illness: A pilot cohort study among pediatric patients. Clin. Nutr. 2021, 40, 2837–2844. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.Q.; Yin, H.; Wu, J.Z.; Chen, R.Z.; Xia, Y.; Wang, L.; Zhu, H.Y.; Fan, L.; Li, J.Y.; Liang, J.H.; et al. 25-Hydroxy vitamin D deficiency predicts inferior prognosis in Hodgkin lymphoma. Leuk. Res. 2021, 105, 106580. [Google Scholar] [CrossRef]
- Xu, D.M.; Liang, J.H.; Wang, L.; Zhu, H.Y.; Xia, Y.; Fan, L.; Li, J.Y.; Xu, W. 25-Hydroxy vitamin D deficiency predicts inferior prognosis in mantle cell lymphoma. J. Cancer Res. Clin. Oncol. 2020, 146, 1003–1009. [Google Scholar] [CrossRef]
- Shi, H.; Chen, H.; Zhang, Y.; Li, J.; Fu, K.; Xue, W.; Teng, W.; Tian, L. 25-Hydroxyvitamin D level, vitamin D intake, and risk of stroke: A dose-response meta-analysis. Clin. Nutr. 2020, 39, 2025–2034. [Google Scholar] [CrossRef] [PubMed]
- Rotstein, A.; Kodesh, A.; Goldberg, Y.; Reichenberg, A.; Levine, S.Z. Serum folate deficiency and the risks of dementia and all-cause mortality: A national study of old age. Evid. Based Ment. Health 2022, 25, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Zhang, H.L.; Xin, M.; Wang, D.; Zheng, N.; Wang, S.; Xu, J.; Wang, Y.; Zhu, J.; Feng, J. Serum Folate Correlates with Severity of Guillain-Barre Syndrome and Predicts Disease Progression. Biomed. Res. Int. 2018, 2018, 5703279. [Google Scholar] [CrossRef]
- Awwad, H.M.; Ohlmann, C.H.; Stoeckle, M.; Geisel, J.; Obeid, R. Serum concentrations of folate vitamers in patients with a newly diagnosed prostate cancer or hyperplasia. Clin. Biochem. 2018, 56, 41–46. [Google Scholar] [CrossRef]
- Kornerup, L.S.; Hvas, C.L.; Abild, C.B.; Richelsen, B.; Nexo, E. Early changes in vitamin B12 uptake and biomarker status following Roux-en-Y gastric bypass and sleeve gastrectomy. Clin. Nutr. 2019, 38, 906–911. [Google Scholar] [CrossRef]
- Khosravi, M.; Sotoudeh, G.; Amini, M.; Raisi, F.; Mansoori, A.; Hosseinzadeh, M. The relationship between dietary patterns and depression mediated by serum levels of Folate and vitamin B12. BMC Psychiatry 2020, 20, 63. [Google Scholar] [CrossRef]
- Masini, G.; Graham, F.J.; Pellicori, P.; Cleland, J.G.F.; Cuthbert, J.J.; Kazmi, S.; Inciardi, R.M.; Clark, A.L. Criteria for Iron Deficiency in Patients With Heart Failure. J. Am. Coll. Cardiol. 2022, 79, 341–351. [Google Scholar] [CrossRef]
- Ueda, T.; Kawakami, R.; Nogi, K.; Nogi, M.; Ishihara, S.; Nakada, Y.; Nakano, T.; Hashimoto, Y.; Nakagawa, H.; Nishida, T.; et al. Serum iron: A new predictor of adverse outcomes independently from serum hemoglobin levels in patients with acute decompensated heart failure. Sci. Rep. 2021, 11, 2395. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Xu, J.; Yang, C.; Hou, X.; Yang, P. Urinary ferritin creatinine ratio, a potential biomarker for lupus nephritis activity. Clin. Rheumatol. 2021, 40, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Neven, K.Y.; Cox, B.; Cosemans, C.; Gyselaers, W.; Penders, J.; Plusquin, M.; Roels, H.A.; Vrijens, K.; Ruttens, A.; Nawrot, T.S. Lower iodine storage in the placenta is associated with gestational diabetes mellitus. BMC Med. 2021, 19, 47. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Sharp, A.; Villanueva, E.; Ma, Z.F. Breast Milk Iodine Concentration (BMIC) as a Biomarker of Iodine Status in Lactating Women and Children <2 Years of Age: A Systematic Review. Nutrients 2022, 14, 1691. [Google Scholar] [PubMed]
- Vogel-Gonzalez, M.; Tallo-Parra, M.; Herrera-Fernandez, V.; Perez-Vilaro, G.; Chillon, M.; Nogues, X.; Gómez-Zorrilla, S.; López-Montesinos, I.; Arnau-Barrés, I.; Sorli-Redó, M.L.; et al. Low Zinc Levels at Admission Associates with Poor Clinical Outcomes in SARS-CoV-2 Infection. Nutrients 2021, 13, 562. [Google Scholar] [CrossRef]
- Kim, M.C.; Lee, J.I.; Kim, J.H.; Kim, H.J.; Cho, Y.K.; Jeon, W.K.; Kim, B.I.; Sohn, W. Serum zinc level and hepatic fibrosis in patients with nonalcoholic fatty liver disease. PLoS ONE 2020, 15, e0240195. [Google Scholar] [CrossRef] [PubMed]
- Rabassa, M.; Cherubini, A.; Zamora-Ros, R.; Urpi-Sarda, M.; Bandinelli, S.; Ferrucci, L.; Andres-Lacueva, C. Low Levels of a Urinary Biomarker of Dietary Polyphenol Are Associated with Substantial Cognitive Decline over a 3-Year Period in Older Adults: The Invecchiare in Chianti Study. J. Am. Geriatr. Soc. 2015, 63, 938–946. [Google Scholar] [CrossRef] [PubMed]
- Murphy, N.; Achaintre, D.; Zamora-Ros, R.; Jenab, M.; Boutron-Ruault, M.C.; Carbonnel, F.; Savoye, I.; Kaaks, R.; Kühn, T.; Boeing, H.; et al. A prospective evaluation of plasma polyphenol levels and colon cancer risk. Int. J. Cancer 2018, 143, 1620–1631. [Google Scholar] [CrossRef]
- Toh, D.W.K.; Loh, W.W.; Sutanto, C.N.; Yao, Y.; Kim, J.E. Skin carotenoid status and plasma carotenoids: Biomarkers of dietary carotenoids, fruits and vegetables for middle-aged and older Singaporean adults. Br. J. Nutr. 2021, 126, 1398–1407. [Google Scholar] [CrossRef]
- Sanjeevi, N.; Lipsky, L.M.; Nansel, T.R. Hyperglycemia and Carotenoid Intake Are Associated with Serum Carotenoids in Youth with Type 1 Diabetes. J. Acad. Nutr. Diet. 2019, 119, 1340–1348. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. J. Clin. Epidemiol. 2021, 134, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Schardt, C.; Adams, M.B.; Owens, T.; Keitz, S.; Fontelo, P. Utilization of the PICO framework to improve searching PubMed for clinical questions. BMC Med. Inform. Decis. Mak. 2007, 7, 16. [Google Scholar] [CrossRef]
- Martini, L.; Pecoraro, L.; Salvottini, C.; Piacentini, G.; Atkinson, R.; Pietrobelli, A. Appropriate and inappropriate vitamin supplementation in children. J. Nutr. Sci. 2020, 9, e20. [Google Scholar] [CrossRef] [PubMed]
- Uusi-Rasi, K.; Patil, R.; Karinkanta, S.; Tokola, K.; Kannus, P.; Lamberg-Allardt, C.; Sievänen, H. Serum 25-hydroxyvitamin D levels and incident falls in older women. Osteoporos. Int. 2019, 30, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Murdaca, G.; Banchero, S.; Tonacci, A.; Nencioni, A.; Monacelli, F.; Gangemi, S. Vitamin D and Folate as Predictors of MMSE in Alzheimer’s Disease: A Machine Learning Analysis. Diagnostics 2021, 11, 940. [Google Scholar] [CrossRef] [PubMed]
- Selhub, J.; Jacques, P.F.; Dallal, G.; Choumenkovitch, S.; Rogers, G. The use of blood concentrations of vitamins and their respective functional indicators to define folate and vitamin B12 status. Food Nutr. Bull. 2008, 29, S67–S73. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.W.; Lee, H.C. Vitamin D and health—The missing vitamin in humans. Pediatr. Neonatol. 2019, 60, 237–244. [Google Scholar] [CrossRef]
- McLean, E.; Klemm, R.; Subramaniam, H.; Greig, A. Refocusing vitamin A supplementation programmes to reach the most vulnerable. BMJ Glob. Health 2020, 5, e001997. [Google Scholar] [CrossRef] [PubMed]
- Li, C.C.; Liu, C.; Fu, M.; Hu, K.Q.; Aizawa, K.; Takahashi, S.; Hiroyuki, S.; Cheng, J.; von Lintig, J.; Wang, X.D. Tomato Powder Inhibits Hepatic Steatosis and Inflammation Potentially Through Restoring SIRT1 Activity and Adiponectin Function Independent of Carotenoid Cleavage Enzymes in Mice. Mol. Nutr. Food Res. 2018, 62, e1700738. [Google Scholar] [CrossRef]
- Cheng, J.; Eroglu, A. The Promising Effects of Astaxanthin on Lung Diseases. Adv. Nutr. 2021, 12, 850–864. [Google Scholar] [CrossRef]
- Chen, W.; Chen, G. The Roles of Vitamin A in the Regulation of Carbohydrate, Lipid, and Protein Metabolism. J. Clin. Med. 2014, 3, 453–479. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Balbuena, E.; Miller, B.; Eroglu, A. The Role of beta-Carotene in Colonic Inflammation and Intestinal Barrier Integrity. Front. Nutr. 2021, 8, 723480. [Google Scholar] [CrossRef] [PubMed]
- Debelo, H.; Novotny, J.A.; Ferruzzi, M.G. Vitamin A. Adv. Nutr. 2017, 8, 992–994. [Google Scholar] [CrossRef]
- Marie, A.; Darricau, M.; Touyarot, K.; Parr-Brownlie, L.C.; Bosch-Bouju, C. Role and Mechanism of Vitamin A Metabolism in the Pathophysiology of Parkinson’s Disease. J. Park. Dis. 2021, 11, 949–970. [Google Scholar] [CrossRef] [PubMed]
- Tanumihardjo, S.A. The Dawn of a New Era in Vitamin A Assessment. J. Nutr. 2020, 150, 185–187. [Google Scholar] [CrossRef] [PubMed]
- Tepasse, P.R.; Vollenberg, R.; Fobker, M.; Kabar, I.; Schmidt, H.; Meier, J.A.; Nowacki, T.; Hüsing-Kabar, A. Vitamin A Plasma Levels in COVID-19 Patients: A Prospective Multicenter Study and Hypothesis. Nutrients 2021, 13, 2173. [Google Scholar] [CrossRef] [PubMed]
- Tuckey, R.C.; Cheng, C.Y.S.; Slominski, A.T. The serum vitamin D metabolome: What we know and what is still to discover. J. Steroid Biochem. Mol. Biol. 2019, 186, 4–21. [Google Scholar] [CrossRef]
- Jeenduang, N.; Plyduang, T.; Horpet, D. Association of 25-hydroxyvitamin D levels and metabolic syndrome in Thai postmenopausal women. Diabetes Metab. Syndr. 2020, 14, 1585–1590. [Google Scholar] [CrossRef]
- Bikle, D.; Christakos, S. New aspects of vitamin D metabolism and action—Addressing the skin as source and target. Nat. Rev. Endocrinol. 2020, 16, 234–252. [Google Scholar] [CrossRef]
- Hanel, A.; Carlberg, C. Vitamin D and evolution: Pharmacologic implications. Biochem. Pharmacol. 2020, 173, 113595. [Google Scholar] [CrossRef] [PubMed]
- Onwuneme, C.; Martin, F.; McCarthy, R.; Carroll, A.; Segurado, R.; Murphy, J.; Twomey, A.; Murphy, N.; Kilbane, M.; McKenna, M.; et al. The Association of Vitamin D Status with Acute Respiratory Morbidity in Preterm Infants. J. Pediatr. 2015, 166, 1175–1180.e1. [Google Scholar] [CrossRef]
- Shab-Bidar, S.; Bours, S.P.; Geusens, P.P.; van der Velde, R.Y.; Janssen, M.J.; van den Bergh, J.P. Suboptimal effect of different vitamin D3 supplementations and doses adapted to baseline serum 25(OH)D on achieved 25(OH)D levels in patients with a recent fracture: A prospective observational study. Eur. J. Endocrinol. 2013, 169, 597–604. [Google Scholar] [CrossRef][Green Version]
- Cashman, K.D.; van den Heuvel, E.G.; Schoemaker, R.J.; Preveraud, D.P.; Macdonald, H.M.; Arcot, J. 25-Hydroxyvitamin D as a Biomarker of Vitamin D Status and Its Modeling to Inform Strategies for Prevention of Vitamin D Deficiency within the Population. Adv. Nutr. 2017, 8, 947–957. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Cheng, C.; Wang, Y.; Xue, Y.; Liu, Y.; Li, W.; Li, X. Serum 25-Hydroxyvitamin D Concentrations and Cardiometabolic Biomarkers in Chinese Rural Population. Horm. Metab. Res. 2021, 53, 105–111. [Google Scholar] [CrossRef]
- Kwon, B.S.; Lee, K.; Kim, E.S.; Jun, S.H.; Lim, S.Y.; Song, M.J.; Kim, Y.W.; Lee, Y.J.; Park, J.S.; Cho, Y.J.; et al. A Prospective Cohort Study of Bioavailable 25-Hydroxyvitamin D Levels as a Marker of Vitamin D Status in Nontuberculous Mycobacterial Pulmonary Disease. Nutrients 2021, 13, 2524. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.R.; Dudenkov, D.V.; Mara, K.C.; Fischer, P.R.; Maxson, J.A.; Thacher, T.D. Serum 25-Hydroxyvitamin D and Subsequent Cancer Incidence and Mortality: A Population-Based Retrospective Cohort Study. Mayo Clin. Proc. 2021, 96, 2157–2167. [Google Scholar] [CrossRef]
- Ebara, S. Nutritional role of folate. Congenit. Anom. 2017, 57, 138–141. [Google Scholar] [CrossRef]
- Jankovic-Karasoulos, T.; Furness, D.L.; Leemaqz, S.Y.; Dekker, G.A.; Grzeskowiak, L.E.; Grieger, J.A.; Andraweera, P.H.; McCullough, D.; McAninch, D.; McCowan, L.M.; et al. Maternal folate, one-carbon metabolism and pregnancy outcomes. Matern. Child. Nutr. 2021, 17, e13064. [Google Scholar] [CrossRef]
- Lu, J.; Trabert, B.; Liao, L.M.; Pfeiffer, R.M.; Michels, K.A. Dietary intake of nutrients involved in folate-mediated one-carbon metabolism and risk for endometrial cancer. Int. J. Epidemiol. 2019, 48, 474–488. [Google Scholar] [CrossRef]
- Naderi, N.; House, J.D. Recent Developments in Folate Nutrition. Adv. Food Nutr. Res. 2018, 83, 195–213. [Google Scholar]
- Van Gool, J.D.; Hirche, H.; Lax, H.; De Schaepdrijver, L. Folic acid and primary prevention of neural tube defects: A review. Reprod. Toxicol. 2018, 80, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Long, P.; Liu, X.; Li, J.; He, S.; Chen, H.; Yuan, Y.; Qiu, G.; Yu, K.; Liu, K.; Jiang, J.; et al. Circulating folate concentrations and risk of coronary artery disease: A prospective cohort study in Chinese adults and a Mendelian randomization analysis. Am. J. Clin. Nutr. 2020, 111, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Ohrvik, V.; Lemming, E.W.; Nalsen, C.; Becker, W.; Ridefelt, P.; Lindroos, A.K. Dietary intake and biomarker status of folate in Swedish adults. Eur. J. Nutr. 2018, 57, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Obeid, R.; Schon, C.; Wilhelm, M.; Pietrzik, K.; Pilz, S. Dietary and lifestyle predictors of folate insufficiency in non-supplemented German women. Int. J. Food Sci. Nutr. 2019, 70, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Solvik, B.S.; Strand, T.A.; Kvestad, I.; Markhus, M.W.; Ueland, P.M.; McCann, A.; Øyen, J. Dietary Intake and Biomarkers of Folate and Cobalamin Status in Norwegian Preschool Children: The FINS-KIDS Study. J. Nutr. 2020, 150, 1852–1858. [Google Scholar] [CrossRef]
- Watanabe, F.; Bito, T. Vitamin B12 sources and microbial interaction. Exp. Biol. Med. 2018, 243, 148–158. [Google Scholar] [CrossRef]
- Soh, Y.; Lee, D.H.; Won, C.W. Association between Vitamin B12 levels and cognitive function in the elderly Korean population. Medicine 2020, 99, e21371. [Google Scholar] [CrossRef]
- Obeid, R.; Heil, S.G.; Verhoeven, M.M.A.; van den Heuvel, E.; de Groot, L.; Eussen, S. Vitamin B12 Intake From Animal Foods, Biomarkers, and Health Aspects. Front. Nutr. 2019, 6, 93. [Google Scholar] [CrossRef]
- Van Wijngaarden, J.P.; Dhonukshe-Rutten, R.A.M.; Brouwer-Brolsma, E.M.; Enneman, A.W.; Swart, K.M.A.; van Dijk, S.C.; In ’t Veld, P.H.; van Schoor, N.M.; van der Velde, N.; de Jonge, R.; et al. Vitamin B12 Intake and Related Biomarkers: Associations in a Dutch Elderly Population. J. Nutr. Health Aging 2017, 21, 1268–1276. [Google Scholar] [CrossRef]
- Lederer, A.K.; Hannibal, L.; Hettich, M.; Behringer, S.; Spiekerkoetter, U.; Steinborn, C.; Gründemann, C.; Zimmermann-Klemd, A.M.; Müller, A.; Simmet, T.; et al. Vitamin B12 Status Upon Short-Term Intervention with a Vegan Diet-A Randomized Controlled Trial in Healthy Participants. Nutrients 2019, 11, 2815. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, S.; Zhang, X.; Xu, Y.; Cheng, J.; Yang, X. The Role of Diet, Eating Behavior, and Nutrition Intervention in Seasonal Affective Disorder: A Systematic Review. Front. Psychol. 2020, 11, 1451. [Google Scholar] [CrossRef]
- Song, J.; Zhou, B.; Kan, J.; Liu, G.; Zhang, S.; Si, L.; Zhang, X.; Yang, X.; Ma, J.; Cheng, J.; et al. Gut microbiota: Linking nutrition and perinatal depression. Front. Cell. Infect. Microbiol. 2022, 12, 1260. [Google Scholar]
- Kocylowski, R.; Lewicka, I.; Grzesiak, M.; Gaj, Z.; Sobanska, A.; Poznaniak, J.; von Kaisenberg, C.; Suliburska, J. Assessment of dietary intake and mineral status in pregnant women. Arch. Gynecol. Obstet. 2018, 297, 1433–1440. [Google Scholar] [CrossRef]
- Naito, Y.; Masuyama, T.; Ishihara, M. Iron and cardiovascular diseases. J. Cardiol. 2021, 77, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Mohammadifard, N.; Humphries, K.H.; Gotay, C.; Mena-Sanchez, G.; Salas-Salvado, J.; Esmaillzadeh, A.; Ignaszewski, A.; Sarrafzadegan, N. Trace minerals intake: Risks and benefits for cardiovascular health. Crit. Rev. Food Sci. Nutr. 2019, 59, 1334–1346. [Google Scholar] [CrossRef] [PubMed]
- Pasricha, S.-R.; Tye-Din, J.; Muckenthaler, M.U.; Swinkels, D.W. Iron deficiency. Lancet 2021, 397, 233–248. [Google Scholar] [CrossRef] [PubMed]
- Bilek, R.; Dvorakova, M.; Grimmichova, T.; Jiskra, J. Iodine, thyroglobulin and thyroid gland. Physiol. Res. 2020, 69, S225–S236. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Diaz, E.; Pearce, E.N. Iodine status and supplementation before, during, and after pregnancy. Best Pr. Res. Clin. Endocrinol. Metab. 2020, 34, 101430. [Google Scholar] [CrossRef]
- Monaghan, A.M.; Mulhern, M.S.; McSorley, E.M.; Strain, J.J.; Dyer, M.; van Wijngaarden, E.; Yeates, A.J. Associations between maternal urinary iodine assessment, dietary iodine intakes and neurodevelopmental outcomes in the child: A systematic review. Thyroid Res. 2021, 14, 14. [Google Scholar] [CrossRef]
- Madar, A.A.; Heen, E.; Hopstock, L.A.; Carlsen, M.H.; Meyer, H.E. Iodine Intake in Norwegian Women and Men: The Population-Based Tromso Study 2015–2016. Nutrients 2020, 12, 3246. [Google Scholar] [CrossRef] [PubMed]
- Bertinato, J. Iodine nutrition: Disorders, monitoring and policies. Adv. Food Nutr. Res. 2021, 96, 365–415. [Google Scholar] [PubMed]
- Cui, T.; Wang, W.; Chen, W.; Pan, Z.; Gao, S.; Tan, L.; Pearce, E.N.; Zimmermann, M.B.; Shen, J.; Zhang, W. Serum Iodine Is Correlated with Iodine Intake and Thyroid Function in School-Age Children from a Sufficient-to-Excessive Iodine Intake Area. J. Nutr. 2019, 149, 1012–1018. [Google Scholar] [CrossRef] [PubMed]
- Hlucny, K.; Alexander, B.M.; Gerow, K.; Larson-Meyer, D.E. Reflection of Dietary Iodine in the 24 h Urinary Iodine Concentration, Serum Iodine and Thyroglobulin as Biomarkers of Iodine Status: A Pilot Study. Nutrients 2021, 13, 2520. [Google Scholar] [CrossRef]
- Pinheiro, C.; Xavier Moreira, N.; Ferreira, P.; Matta Coelho, C.; Guimaraes, J.; Pereira, G.; Cortez, A.; Bracchi, I.; Pestana, D.; Barreiros Mota, I.; et al. Iodine knowledge is associated with iodine status in Portuguese pregnant women: Results from the IoMum cohort study. Br. J. Nutr. 2021, 126, 1331–1339. [Google Scholar] [CrossRef]
- Kanike, N.; Groh-Wargo, S.; Thomas, M.; Chien, E.K.; Mhanna, M.; Kumar, D.; Worley, S.; Singh, R.J.; Shekhawat, P.S. Risk of Iodine Deficiency in Extremely Low Gestational Age Newborns on Parenteral Nutrition. Nutrients 2020, 12, 1636. [Google Scholar] [CrossRef]
- Gattermann, N.; Muckenthaler, M.U.; Kulozik, A.E.; Metzgeroth, G.; Hastka, J. The Evaluation of Iron Deficiency and Iron Overload. Dtsch. Arztebl. Int. 2021, 118, 847–856. [Google Scholar] [CrossRef]
- Georgieff, M.K. Iron deficiency in pregnancy. Am. J. Obs. Gynecol. 2020, 223, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Mei, Z.; Addo, O.Y.; Jefferds, M.E.; Sharma, A.J.; Flores-Ayala, R.C.; Brittenham, G.M. Physiologically based serum ferritin thresholds for iron deficiency in children and non-pregnant women: A US National Health and Nutrition Examination Surveys (NHANES) serial cross-sectional study. Lancet Haematol. 2021, 8, e572–e582. [Google Scholar] [CrossRef]
- Zhang, Q.; Zheng, J.; Wu, X.; Chen, S.; Qiu, J.; Sun, M. Multi-index evaluation system and relevant critical values for monitoring iron status in Chinese athletes. Med. Sport 2022, 75, 59–68. [Google Scholar] [CrossRef]
- Joo, Y.S.; Kim, H.W.; Lee, S.; Nam, K.H.; Yun, H.R.; Jhee, J.H.; Han, S.H.; Yoo, T.H.; Kang, S.W.; Park, J.T. Dietary zinc intake and incident chronic kidney disease. Clin. Nutr. 2021, 40, 1039–1045. [Google Scholar] [CrossRef]
- Baltaci, A.K.; Yuce, K.; Mogulkoc, R. Zinc Metabolism and Metallothioneins. Biol. Trace Elem. Res. 2018, 183, 22–31. [Google Scholar] [CrossRef]
- Ackland, M.L.; Michalczyk, A.A. Zinc and infant nutrition. Arch. Biochem. Biophys. 2016, 611, 51–57. [Google Scholar] [CrossRef]
- Brown, K.H.; Engle-Stone, R.; Krebs, N.F.; Peerson, J.M. Dietary intervention strategies to enhance zinc nutrition: Promotion and support of breastfeeding for infants and young children. Food Nutr. Bull. 2009, 30 (Suppl. S1), S144–S171. [Google Scholar] [CrossRef] [PubMed]
- Lowe, N.M.; Medina, M.W.; Stammers, A.L.; Patel, S.; Souverein, O.W.; Dullemeijer, C.; Serra-Majem, L.; Nissensohn, M.; Hall Moran, V. The relationship between zinc intake and serum/plasma zinc concentration in adults: A systematic review and dose-response meta-analysis by the EURRECA Network. Br. J. Nutr. 2012, 108, 1962–1971. [Google Scholar] [CrossRef] [PubMed]
- Atasoy, H.I.; Bugdayci, G. Zinc deficiency and its predictive capacity for anemia: Unique model in school children. Pediatr. Int. 2018, 60, 703–709. [Google Scholar] [CrossRef] [PubMed]
- Maares, M.; Hackler, J.; Haupt, A.; Heller, R.A.; Bachmann, M.; Diegmann, J.; Moghaddam, A.; Schomburg, L.; Haase, H. Free Zinc as a Predictive Marker for COVID-19 Mortality Risk. Nutrients 2022, 14, 1407. [Google Scholar] [CrossRef] [PubMed]
- Kan, J.; Wu, F.; Wang, F.; Zheng, J.; Cheng, J.; Li, Y.; Yang, Y.; Du, J. Phytonutrients: Sources, bioavailability, interaction with gut microbiota, and their impacts on human health. Front. Nutr. 2022, 9, 960309. [Google Scholar] [CrossRef]
- Fraga, C.G.; Croft, K.D.; Kennedy, D.O.; Tomás-Barberán, F.A. The effects of polyphenols and other bioactives on human health. Food Funct. 2019, 20, 514–528. [Google Scholar] [CrossRef]
- Bacanli, M.; Dilsiz, S.A.; Basaran, N.; Basaran, A.A. Effects of phytochemicals against diabetes. Adv. Food Nutr. Res. 2019, 89, 209–238. [Google Scholar]
- Bulotta, S.; Capriglione, F.; Celano, M.; Pecce, V.; Russo, D.; Maggisano, V. Phytochemicals in thyroid cancer: Analysis of the preclinical studies. Endocrine 2021, 73, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Tresserra-Rimbau, A.; Estruch, R.; Martinez-Gonzalez, M.A.; Medina-Remon, A.; Castaner, O.; Corella, D.; Salas-Salvadó, J.; Lamuela-Raventós, R.M. Effects of Polyphenol, Measured by a Biomarker of Total Polyphenols in Urine, on Cardiovascular Risk Factors After a Long-Term Follow-Up in the PREDIMED Study. Oxidative Med. Cell. Longev. 2016, 2016, 2572606. [Google Scholar] [CrossRef]
- Granado-Lorencio, F.; Blanco-Navarro, I.; Perez-Sacristan, B.; Hernandez-Alvarez, E. Biomarkers of carotenoid bioavailability. Food Res. Int. 2017, 99, 902–916. [Google Scholar] [CrossRef]
- Rana, A.; Samtiya, M.; Dhewa, T.; Mishra, V.; Aluko, R.E. Health benefits of polyphenols: A concise review. J. Food Biochem. 2022, 10, e14264. [Google Scholar] [CrossRef] [PubMed]
- Perez-Jimenez, J.; Fezeu, L.; Touvier, M.; Arnault, N.; Manach, C.; Hercberg, S.; Galan, P.; Scalbert, A. Dietary intake of 337 polyphenols in French adults. Am. J. Clin. Nutr. 2011, 93, 1220–1228. [Google Scholar] [CrossRef] [PubMed]
- Zamora-Ros, R.; Rabassa, M.; Cherubini, A.; Urpi-Sarda, M.S.; Ferrucci, L.; Andres-Lacueva, C. High concentrations of a urinary biomarker of polyphenol intake are associated with decreased mortality in older adults. J. Nutr. 2013, 143, 1445–1450. [Google Scholar] [CrossRef] [PubMed]
- Kan, J.; Wang, M.; Liu, Y.; Liu, H.; Chen, L.; Zhang, X.; Huang, C.; Liu, B.Y.; Gu, Z.; Du, J. A novel botanical formula improves eye fatigue and dry eye: A randomized, double-blind, placebo-controlled study. Am. J. Clin. Nutr. 2020, 112, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Prentice, R.L.; Pettinger, M.; Neuhouser, M.L.; Tinker, L.F.; Huang, Y.; Zheng, C.; Manson, J.E.; Mossavar-Rahmani, Y.; Anderson, G.L.; Lampe, J.W. Application of blood concentration biomarkers in nutritional epidemiology: Example of carotenoid and tocopherol intake in relation to chronic disease risk. Am. J. Clin. Nutr. 2019, 1, 1189–1196. [Google Scholar] [CrossRef]
- Allore, T.; Lemieux, S.; Vohl, M.C.; Couture, P.; Lamarche, B.; Couillard, C. Correlates of the difference in plasma carotenoid concentrations between men and women. Br. J. Nutr. 2019, 121, 172–181. [Google Scholar] [CrossRef]
- Jenab, M.; Slimani, N.; Bictash, M.; Jenab, M.; Slimani, N.; Bictash, M.; Ferrari, P.; Bingham, S.A. Biomarkers in nutritional epidemiology: Applications, needs and new horizons. Hum. Genet. 2009, 125, 507–525. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Lopez, O.; Milagro, F.I.; Allayee, H.; Chmurzynska, A.; Choi, M.S.; Curi, R.; De Caterina, R.; Ferguson, L.R.; Goni, L.; Kang, J.X.; et al. Guide for Current Nutrigenetic, Nutrigenomic, and Nutriepigenetic Approaches for Precision Nutrition Involving the Prevention and Management of Chronic Diseases Associated with Obesity. J. Nutrigenet. Nutrigenomics. 2017, 10, 43–62. [Google Scholar] [CrossRef] [PubMed]
- Fito, M.; Melander, O.; Martinez, J.A.; Toledo, E.; Carpene, C.; Corella, D. Advances in Integrating Traditional and Omic Biomarkers When Analyzing the Effects of the Mediterranean Diet Intervention in Cardiovascular Prevention. Int. J. Mol. Sci. 2016, 17, 1469. [Google Scholar] [CrossRef] [PubMed]
- Zeevi, D.; Korem, T.; Zmora, N.; Israeli, D.; Rothschild, D.; Weinberger, A.; Ben-Yacov, O.; Lador, D.; Avnit-Sagi, T.; Lotan-Pompan, M.; et al. Personalized Nutrition by Prediction of Glycemic Responses. Cell 2015, 163, 1079–1094. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutritional Biomarker | Sample Type | Potential Mechanism | Related Epidemiology Studies | Ref. |
|---|---|---|---|---|
| Vitamin A | Serum | Pro-inflammatory and anti-inflammatory factor imbalances and reduced immunological function in sepsis may result from VA insufficiency. | Sepsis | [21] |
| Retinol binding protein | Blood | Acute phase response, induced by acute infection, are responsible for reductions in circulating vitamin A. | Acute febrile illness | [22] |
| Vitamin D [25-(OH)D] | Serum | Stimulated the expression of PD-L1 and PD-L2, suppressing T-cell mediated anti-tumor immunity | Mantle cell lymphoma/Hodgkin lymphoma | [23,24] |
| Vitamin D [25-(OH)D] | Serum | Activated vitamin D is an inhibitor of the renin angiotensin system (RAS), an important pathway in the regulation of the cardiovascular system. | Stroke | [25] |
| Folate | Serum | Folate deficiency affects homocysteine level and affects impaired DNA repair in neurons, sensitizing them to oxidative damage, | Dementia and all-cause mortality | [26] |
| Folate | Serum | Diminish human immune functions by affecting T and B cell differentials and the proliferation response of lymphocytes/Affects the synthesis of methyl groups and DNA in growing cells | Guillain-Barre syndrome, Prostate cancer | [27,28] |
| Vitamin B12 | Plasma/Serum | Rearrangement compromises vitamin B12 absorption | Bariatric surgery | [29] |
| Vitamin B12 and Folate | Serum | Prevent impaired immune function and inflammation | Depression | [30] |
| Iron | Serum | Increasing hepcidin activity by inflammation in Heart Failure may lead to the development of functional Iron deficiency | Heart failure | [31,32] |
| Ferritin/Creatinine | Urine | Inflammation can stimulate the proliferation and activation of monocyte phagocytic system, which leads to the increase of iron absorption and retention in the reticuloendothelial system. | Lupus nephritis | [33] |
| Iodine | Urine/serum | Altered thyroid hormone homeostasis and dysregulation of glucose metabolism | Gestational diabetes mellitus | [34] |
| Iodine | Urine/breast milk | Maintaining thyroid function and body metabolism | Neurodevelopment/Iodine deficiency disorders | [35] |
| Zinc | Serum/Plasma | Effect on the immune system, as both the adaptive and the innate immunity, are affected by zinc levels. | COVID-19 | [36] |
| Zinc | Serum | Inhibitor of NADPH oxidase, Zn acts as a potent promoter of autophagy-mediated lipophagy in the liver, reducing lipid accumulation and stimulating lipolysis | Hepatic fibrosis | [37] |
| Polyphenols | Urine/serum | Polyphenols reduce neuronal damage and death from oxidative reactions by inhibiting the generation of reactive oxygen species, lipid peroxidation, apoptosis, protein oxidation, metal chelation, and damage to cellular signaling. | Cognition disorder | [38] |
| Polyphenols | Plasma | Antioxidants, anti-inflammatory and estrogenic effects | Colon cancer | [39] |
| Carotenoids | Skin tissue/Plasma | Less susceptible to fluctuations in response to recent dietary intake and potential carotenoid degradation by heat, light and oxygen. | Metabolism disorder/Obesity | [40] |
| Carotenoids | Serum | Oxidative stress can deplete serum carotenoids, and thus attenuate the association of serum carotenoids with intake. | Type 1 diabetes | [41] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, J.; Wu, F.; Wang, F.; Cheng, J.; Zou, H.; Li, Y.; Du, J.; Kan, J. Biomarkers of Micronutrients and Phytonutrients and Their Application in Epidemiological Studies. Nutrients 2023, 15, 970. https://doi.org/10.3390/nu15040970
Zheng J, Wu F, Wang F, Cheng J, Zou H, Li Y, Du J, Kan J. Biomarkers of Micronutrients and Phytonutrients and Their Application in Epidemiological Studies. Nutrients. 2023; 15(4):970. https://doi.org/10.3390/nu15040970
Chicago/Turabian StyleZheng, Jianheng, Feng Wu, Feijie Wang, Junrui Cheng, Hong Zou, Yuan Li, Jun Du, and Juntao Kan. 2023. "Biomarkers of Micronutrients and Phytonutrients and Their Application in Epidemiological Studies" Nutrients 15, no. 4: 970. https://doi.org/10.3390/nu15040970
APA StyleZheng, J., Wu, F., Wang, F., Cheng, J., Zou, H., Li, Y., Du, J., & Kan, J. (2023). Biomarkers of Micronutrients and Phytonutrients and Their Application in Epidemiological Studies. Nutrients, 15(4), 970. https://doi.org/10.3390/nu15040970