
The Gut Microbiota–Brain Axis during Aging, Mild Cognitive Impairment and Dementia: Role of Tau Protein, β-Amyloid and LPS in Serum and Curli Protein in Stool

 , ,
, ,  , , , ,
, , , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Anthropometry and Body Composition
2.3. Blood Pressure
2.4. Biochemical Parameters
2.5. Determination of ApoE Polymorphism
2.6. Determination of Serum Tau and β Amyloid
2.7. Determination of LPS
2.8. PCR End Point
2.9. Western Blot of Curli Protein
2.10. Fecal DNA Extraction
2.11. Gut Microbiota Analysis by 16S rRNA Sequencing
2.12. Animals
2.13. Statistical Analysis
3. Results
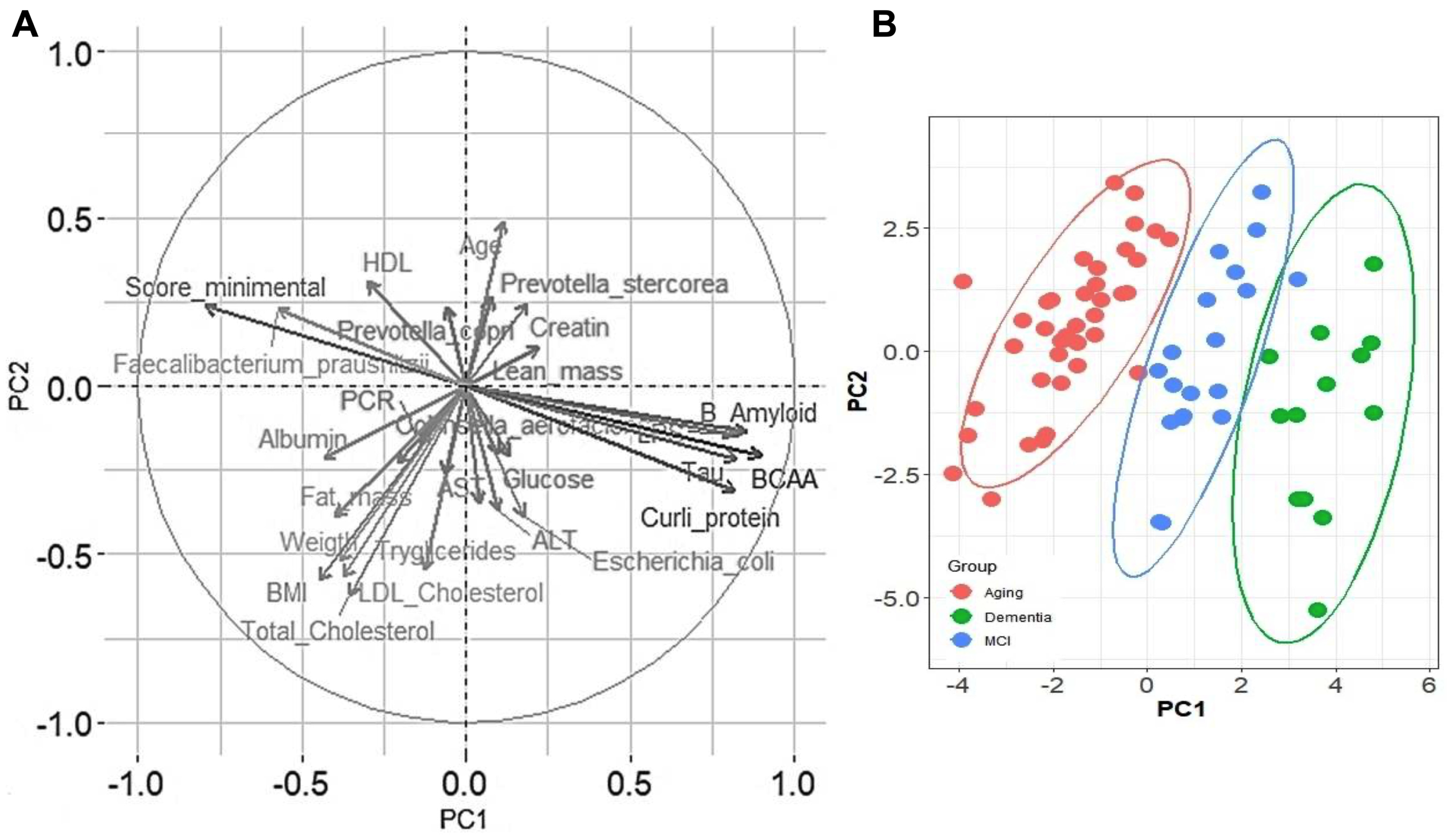
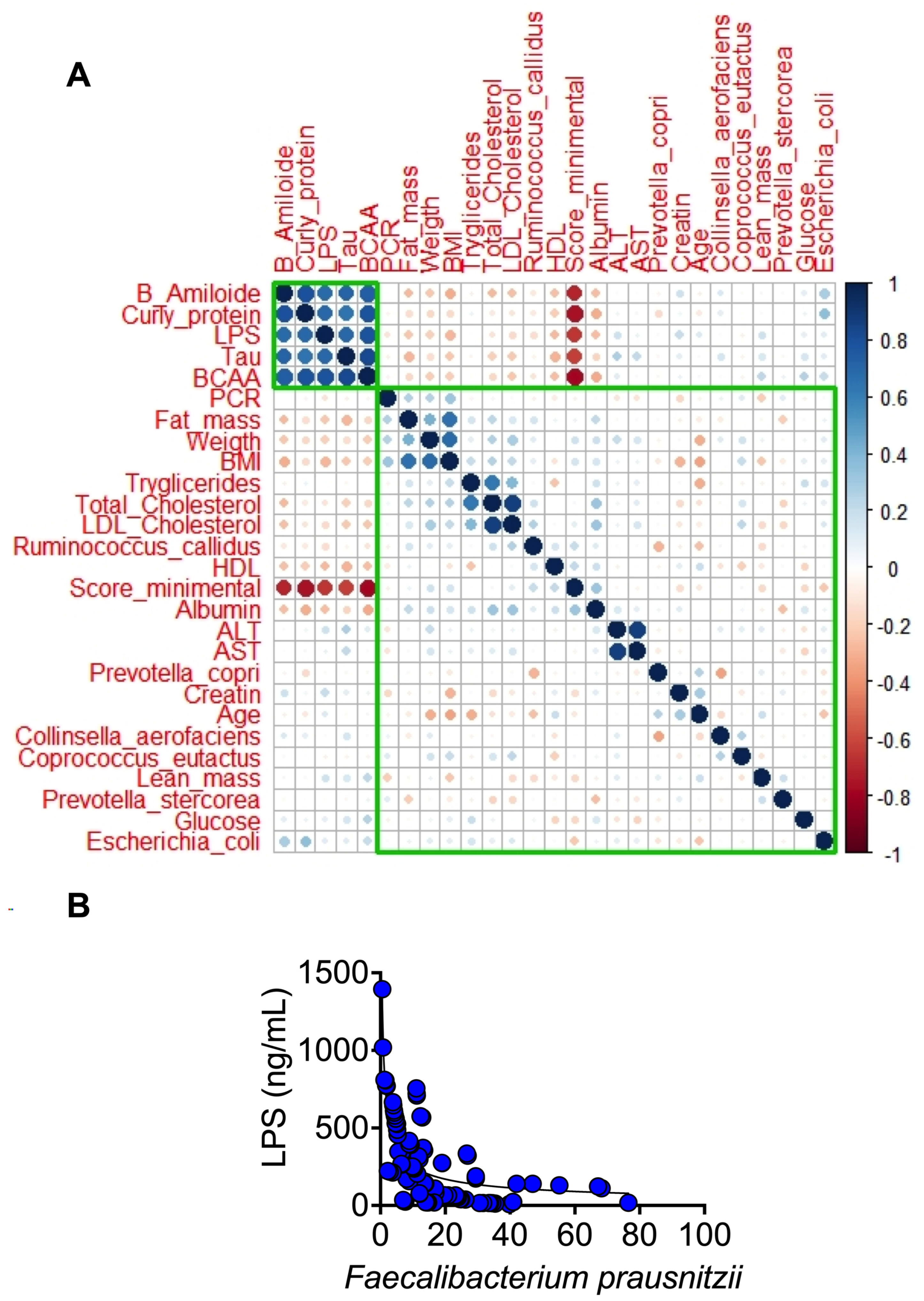
3.1. Anthropometric and Biochemical Parameters
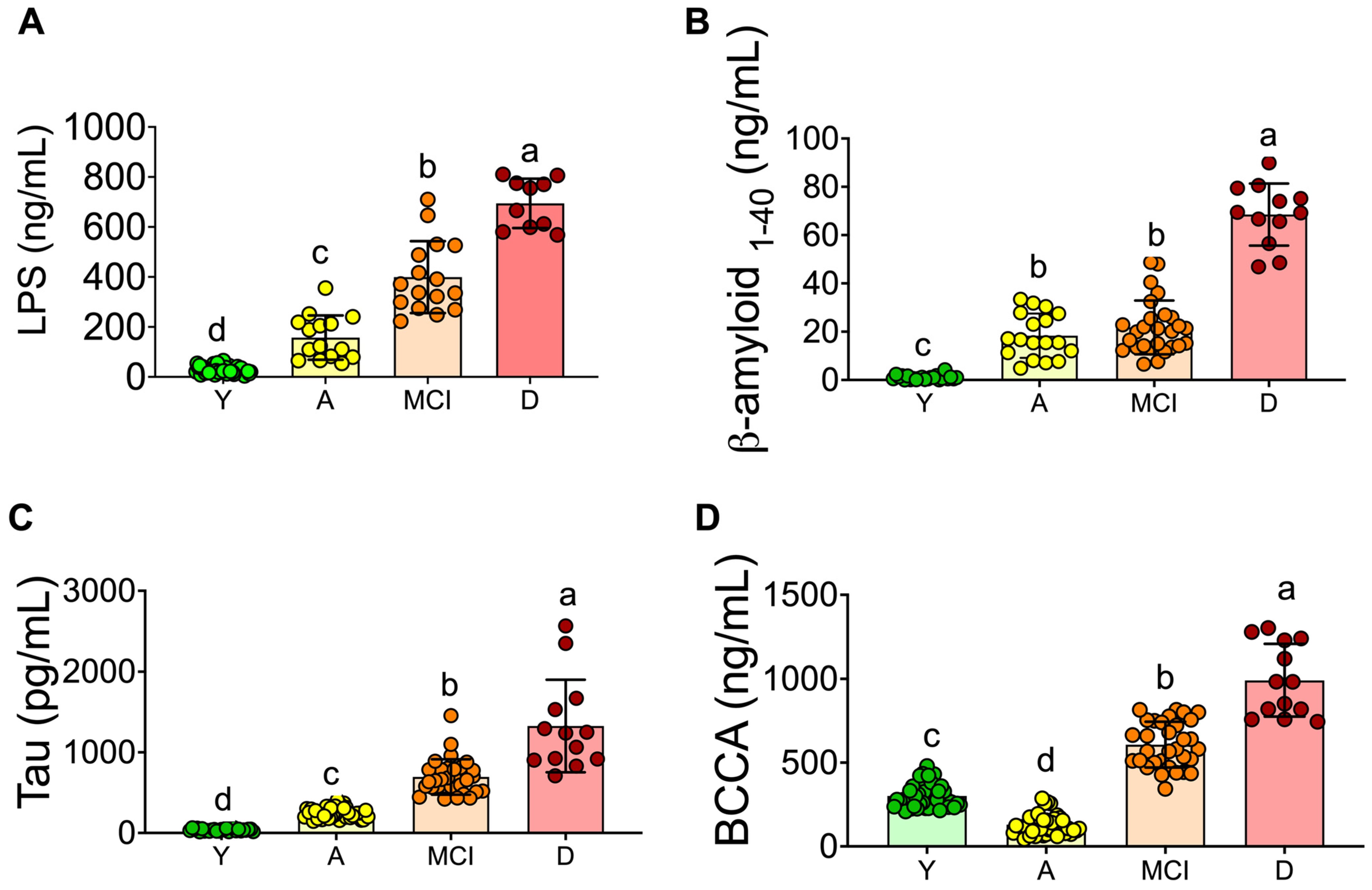
3.2. Inflammation and Neuronal Biomarkers
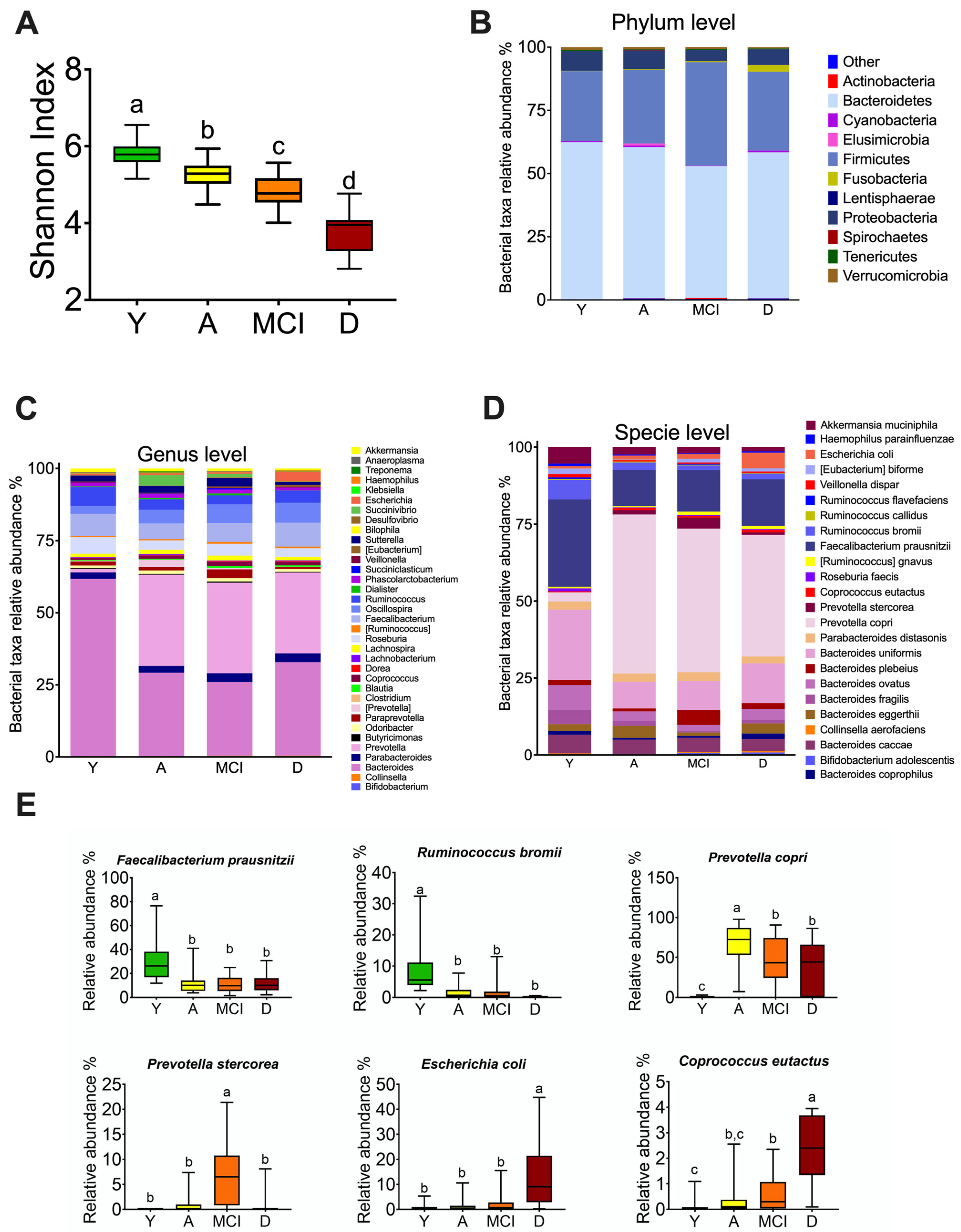
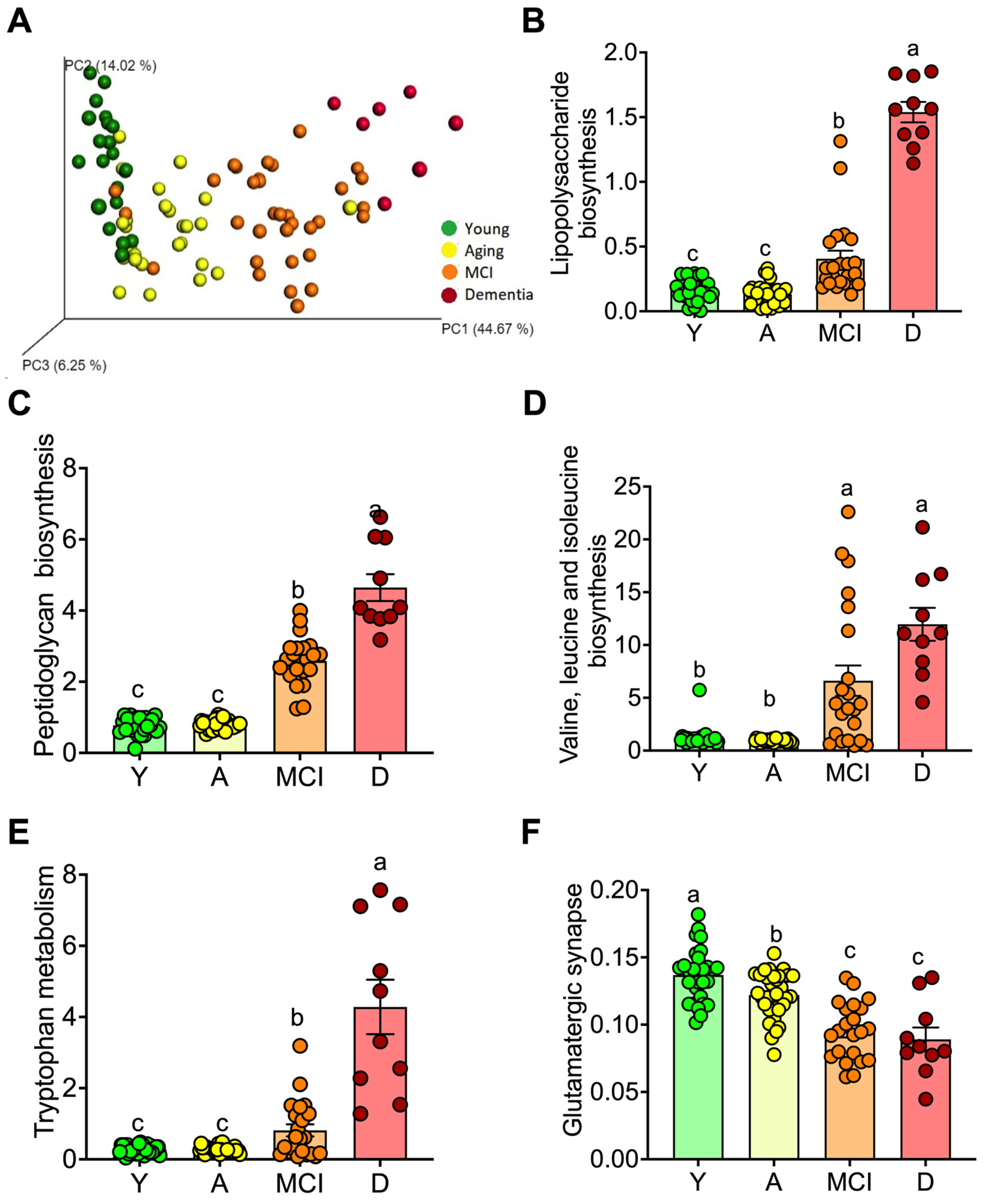
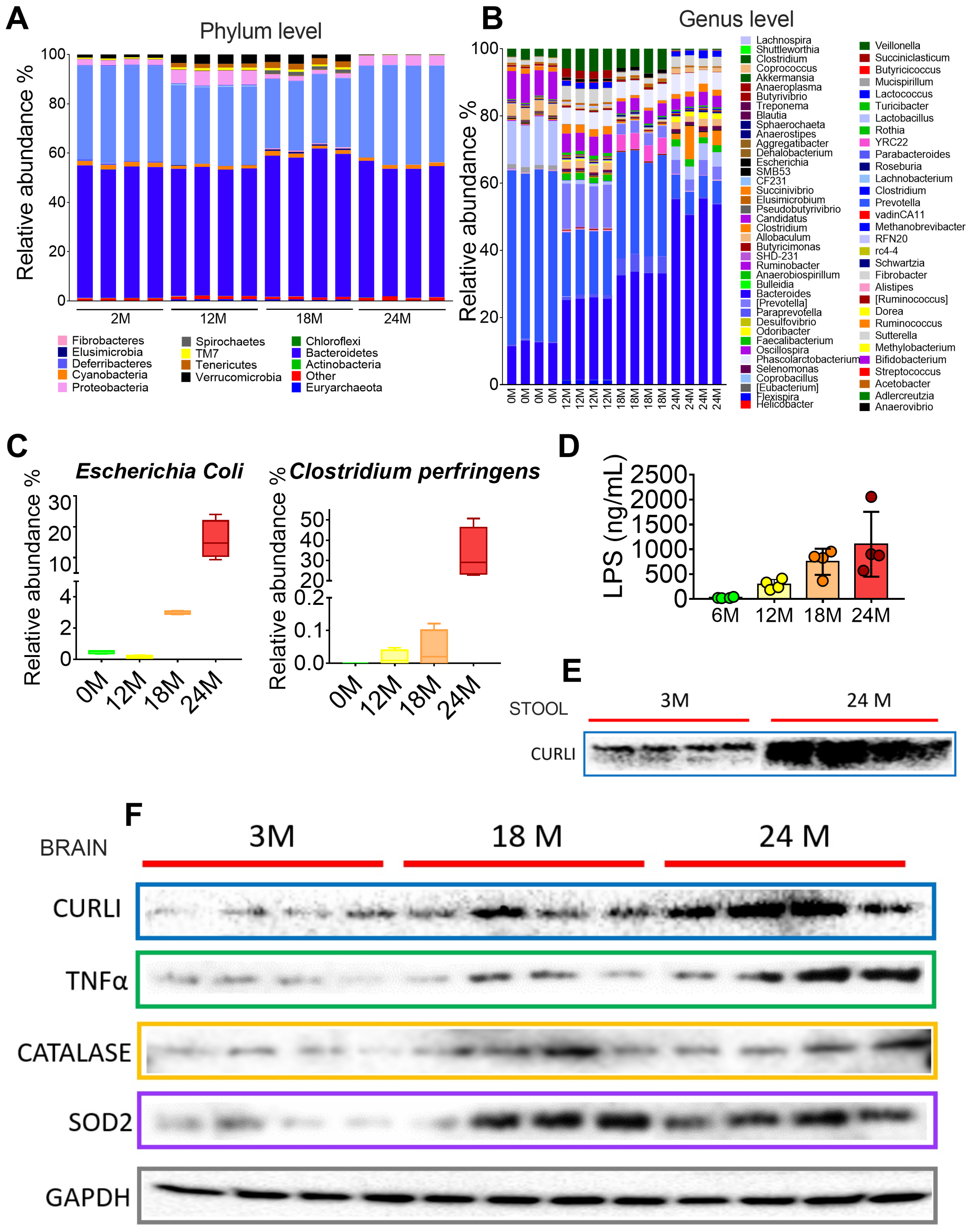
3.3. The Gut Microbiota throughout Life in Humans
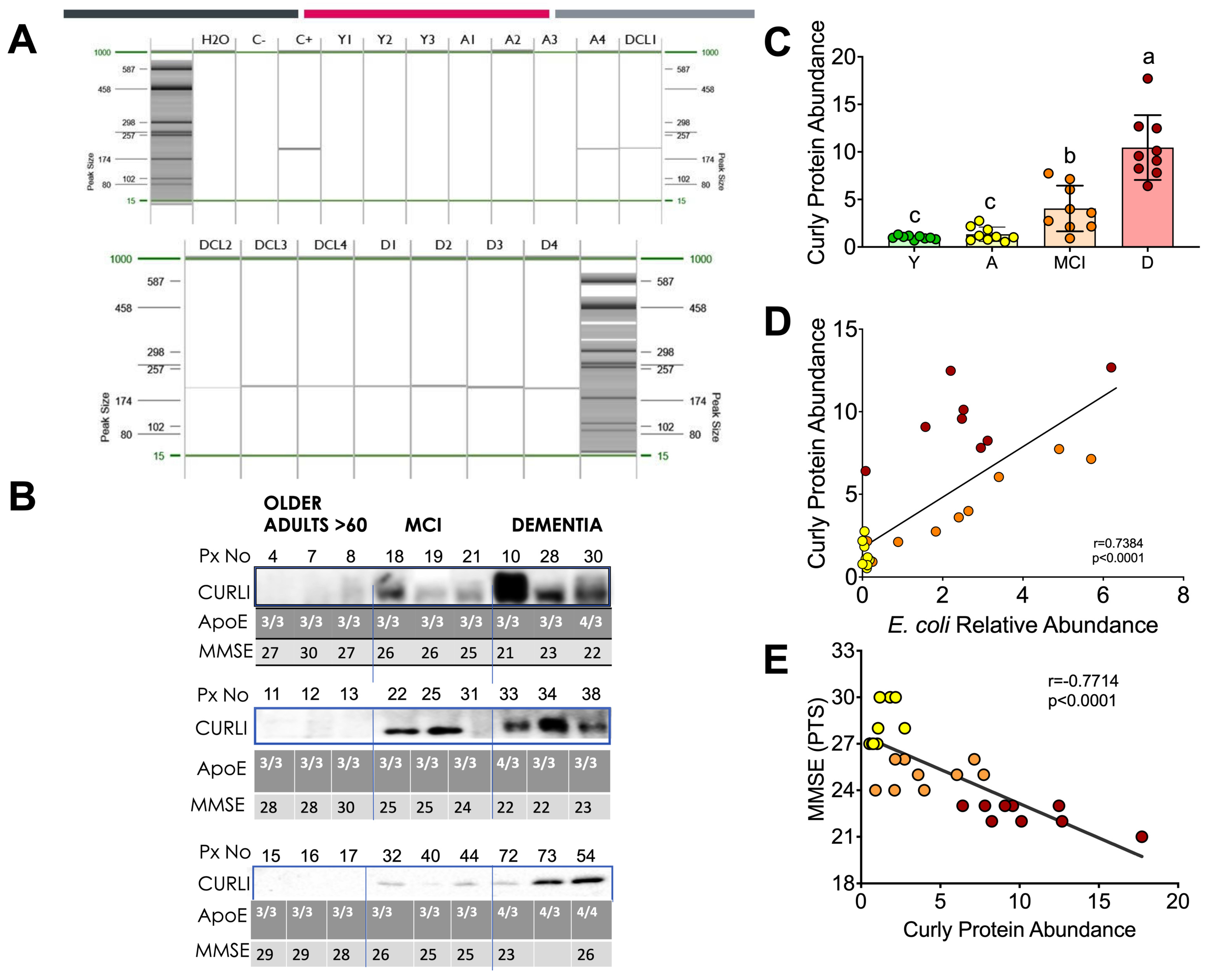
3.4. Curli Protein during Different Periods of Life in Humans
3.5. Gut Microbiota during Different Periods of Life in Rats
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Livingston, G.; Sommerlad, A.; Orgeta, V.; Costafreda, S.G.; Huntley, J.; Ames, D.; Ballard, C.; Banerjee, S.; Burns, A.; Cohen-Mansfield, J.; et al. Dementia prevention, intervention, and care. Lancet 2017, 390, 2673–2734. [Google Scholar] [CrossRef] [PubMed]
- Prince, M.J.; Wu, F.; Guo, Y.; Gutierrez Robledo, L.M.; O’Donnell, M.; Sullivan, R.; Yusuf, S. The burden of disease in older people and implications for health policy and practice. Lancet 2015, 385, 549–562. [Google Scholar] [CrossRef] [PubMed]
- Ferrucci, L.; Fabbri, E. Inflammageing: Chronic inflammation in ageing, cardiovascular disease, and frailty. Nat. Rev. Cardiol. 2018, 15, 505–522. [Google Scholar] [CrossRef] [PubMed]
- Alzheimer’s, A. 2015 Alzheimer’s disease facts and figures. Alzheimer’s Dement. 2015, 11, 332–384. [Google Scholar] [CrossRef]
- Lee, J.W.; Lee, Y.K.; Yuk, D.Y.; Choi, D.Y.; Ban, S.B.; Oh, K.W.; Hong, J.T. Neuro-inflammation induced by lipopolysaccharide causes cognitive impairment through enhancement of beta-amyloid generation. J. Neuroinflamm. 2008, 5, 37. [Google Scholar] [CrossRef]
- Pistollato, F.; Sumalla Cano, S.; Elio, I.; Masias Vergara, M.; Giampieri, F.; Battino, M. Role of gut microbiota and nutrients in amyloid formation and pathogenesis of Alzheimer disease. Nutr. Rev. 2016, 74, 624–634. [Google Scholar] [CrossRef]
- Schwartz, K.; Boles, B.R. Microbial amyloids--functions and interactions within the host. Curr. Opin. Microbiol. 2013, 16, 93–99. [Google Scholar] [CrossRef]
- Tapiola, T.; Alafuzoff, I.; Herukka, S.K.; Parkkinen, L.; Hartikainen, P.; Soininen, H.; Pirttila, T. Cerebrospinal fluid beta-amyloid 42 and tau proteins as biomarkers of Alzheimer-type pathologic changes in the brain. Arch. Neurol. 2009, 66, 382–389. [Google Scholar] [CrossRef]
- Zhu, S.; Jiang, Y.; Xu, K.; Cui, M.; Ye, W.; Zhao, G.; Jin, L.; Chen, X. The progress of gut microbiome research related to brain disorders. J. Neuroinflamm. 2020, 17, 25. [Google Scholar] [CrossRef]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The Microbiota-Gut-Brain Axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef]
- Tillisch, K. The effects of gut microbiota on CNS function in humans. Gut Microbes 2014, 5, 404–410. [Google Scholar] [CrossRef]
- Lambert, J.C.; Ibrahim-Verbaas, C.A.; Harold, D.; Naj, A.C.; Sims, R.; Bellenguez, C.; DeStafano, A.L.; Bis, J.C.; Beecham, G.W.; Grenier-Boley, B.; et al. Meta-analysis of 74,046 individuals identifies 11 new susceptibility loci for Alzheimer’s disease. Nat. Genet. 2013, 45, 1452–1458. [Google Scholar] [CrossRef]
- Reitz, C.; Brayne, C.; Mayeux, R. Epidemiology of Alzheimer disease. Nat. Rev. Neurol. 2011, 7, 137–152. [Google Scholar] [CrossRef] [PubMed]
- Daulatzai, M.A. Role of stress, depression, and aging in cognitive decline and Alzheimer’s disease. Curr. Top. Behav. Neurosci. 2014, 18, 265–296. [Google Scholar] [CrossRef]
- Zhou, Y.; Smith, D.R.; Hufnagel, D.A.; Chapman, M.R. Experimental manipulation of the microbial functional amyloid called curli. Methods Mol. Biol. 2013, 966, 53–75. [Google Scholar] [CrossRef] [PubMed]
- Biagi, E.; Candela, M.; Fairweather-Tait, S.; Franceschi, C.; Brigidi, P. Aging of the human metaorganism: The microbial counterpart. Age 2012, 34, 247–267. [Google Scholar] [CrossRef] [PubMed]
- Medina-Vera, I.; Sanchez-Tapia, M.; Noriega-Lopez, L.; Granados-Portillo, O.; Guevara-Cruz, M.; Flores-Lopez, A.; Avila-Nava, A.; Fernandez, M.L.; Tovar, A.R.; Torres, N. A dietary intervention with functional foods reduces metabolic endotoxaemia and attenuates biochemical abnormalities by modifying faecal microbiota in people with type 2 diabetes. Diabetes Metab. 2019, 45, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Langille, M.G.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef] [PubMed]
- O’Mahony, S.M.; Clarke, G.; Borre, Y.E.; Dinan, T.G.; Cryan, J.F. Serotonin, tryptophan metabolism and the brain-gut-microbiome axis. Behav. Brain Res. 2015, 277, 32–48. [Google Scholar] [CrossRef]
- Conway, M.E. Alzheimer’s disease: Targeting the glutamatergic system. Biogerontology 2020, 21, 257–274. [Google Scholar] [CrossRef]
- Fischer, R.; Maier, O. Interrelation of oxidative stress and inflammation in neurodegenerative disease: Role of TNF. Oxidative Med. Cell. Longev. 2015, 2015, 610813. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Multisectoral Action for a Life Course Approach to Healthy Ageing: Draft Global Strategy and Plan of Action on Ageing and Health; World Health Organization: Geneva, Switzerland, 2016; Available online: http://apps.who.int/gb/ebwha/pdf_les/WHA69/A69_17 (accessed on 11 January 2023).
- Franceschi, C. Inflammaging as a major characteristic of old people: Can it be prevented or cured? Nutr. Rev. 2007, 65, S173–S176. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, K.; Mulak, A. Brain-Gut-Microbiota Axis in Alzheimer’s Disease. J. Neurogastroenterol. Motil. 2019, 25, 48–60. [Google Scholar] [CrossRef]
- Hu, X.; Wang, T.; Jin, F. Alzheimer’s disease and gut microbiota. Sci. China Life Sci. 2016, 59, 1006–1023. [Google Scholar] [CrossRef]
- Sokol, H.; Pigneur, B.; Watterlot, L.; Lakhdari, O.; Bermudez-Humaran, L.G.; Gratadoux, J.J.; Blugeon, S.; Bridonneau, C.; Furet, J.P.; Corthier, G.; et al. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc. Natl. Acad. Sci. USA 2008, 105, 16731–16736. [Google Scholar] [CrossRef]
- Chen, S.G.; Stribinskis, V.; Rane, M.J.; Demuth, D.R.; Gozal, E.; Roberts, A.M.; Jagadapillai, R.; Liu, R.; Choe, K.; Shivakumar, B.; et al. Exposure to the Functional Bacterial Amyloid Protein Curli Enhances Alpha-Synuclein Aggregation in Aged Fischer 344 Rats and Caenorhabditis elegans. Sci. Rep. 2016, 6, 34477. [Google Scholar] [CrossRef] [PubMed]
- Fang, P.; Kazmi, S.A.; Jameson, K.G.; Hsiao, E.Y. The Microbiome as a Modifier of Neurodegenerative Disease Risk. Cell Host Microbe 2020, 28, 201–222. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Health | Aging | MCI | Dementia | p | |
|---|---|---|---|---|---|
| n = 29 | n = 42 | n = 32 | n = 13 | ||
| Biochemical | |||||
| Glucose, mg/dL | 85.3 ± 10.5 | 98.2 ± 14.3 | 116 ± 57.3 | 121 ± 58.6 | 0.01 |
| Triglycerides, mg/dL | 89.9 ± 33 | 150 ± 104 | 143 ± 103 | 150 ± 42 | 0.02 |
| Total cholesterol, mg/dL | 167 ± 35.1 | 192 ± 44 | 181 ± 43 | 174 ± 34 | 0.11 |
| Demographics | |||||
| Age, years | 31.1 ± 9.7 | 66.2 ± 7.0 | 74.3 ± 7.80 | 68.6 ± 7.0 | 0.0001 |
| Female sex, n (%) | 19 (65.5%) | 28 (66.6%) | 16 (50%) | 11 (84.6%) | 0.15 |
| Education, years | 15.2 ± 2.10 | 12.9 ± 4.90 | 11.0 ± 5.18 | 11.5 ± 5.0 | 0.031 |
| Body mass index, kg/m2 | 22.6 ± 1.72 | 28.2 ± 4.43 | 27.7 ± 4.31 | 25.7 ± 3.8 | 0.001 |
| Body fat mass, % | 29.2 ± 8.4 | 33.2 ± 9.4 | 35.7 ± 8.5 | 45.0 ± 5.9 | 0.03 |
| Visceral fat mass, cm2 | 68.3 ± 18.9 | 118 ± 27.0 | 95.9 ± 24.1 | 130 ± 34.1 | 0.0001 |
| Skeletal muscle mass, kg | 43.8 ± 10.5 | 22.5 ± 6.0 | 21.2 ± 4.5 | 14.6 ± 1.9 | 0.0001 |
| Hypertension, n (%) | 17 (40.5%) | 15 (46.9%) | 4 (30.8%) | 0.777 | |
| Diabetes mellitus, n (%) | 1 (2.4%) | 8 (25.0%) | 2 (15.4%) | 0.033 | |
| Dyslipidemia, n (%) | 10 (23.8%) | 10 (31.3%) | 2 (15.4%) | 0.881 | |
| CKD, n (%) | 1 (2.4%) | 1 (3.1%) | 0 | 0.871 | |
| IHD, n (%) | 1 (2.4%) | 3 (9.4%) | 1 (7.7%) | 0.424 | |
| Smoking habit, n (%) | 9 (21.4%) | 9 (28.1%) | 3 (23.1%) | 0.671 | |
| Alcohol consumption, n (%) | 8 (19.0%) | 6 (18.8%) | 2 (15.4%) | 0.870 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Tapia, M.; Mimenza-Alvarado, A.; Granados-Domínguez, L.; Flores-López, A.; López-Barradas, A.; Ortiz, V.; Pérez-Cruz, C.; Sánchez-Vidal, H.; Hernández-Acosta, J.; Ávila-Funes, J.A.; et al. The Gut Microbiota–Brain Axis during Aging, Mild Cognitive Impairment and Dementia: Role of Tau Protein, β-Amyloid and LPS in Serum and Curli Protein in Stool. Nutrients 2023, 15, 932. https://doi.org/10.3390/nu15040932
Sánchez-Tapia M, Mimenza-Alvarado A, Granados-Domínguez L, Flores-López A, López-Barradas A, Ortiz V, Pérez-Cruz C, Sánchez-Vidal H, Hernández-Acosta J, Ávila-Funes JA, et al. The Gut Microbiota–Brain Axis during Aging, Mild Cognitive Impairment and Dementia: Role of Tau Protein, β-Amyloid and LPS in Serum and Curli Protein in Stool. Nutrients. 2023; 15(4):932. https://doi.org/10.3390/nu15040932
Chicago/Turabian StyleSánchez-Tapia, Mónica, Alberto Mimenza-Alvarado, Lizbeth Granados-Domínguez, Adriana Flores-López, Adriana López-Barradas, Victor Ortiz, Claudia Pérez-Cruz, Hilda Sánchez-Vidal, Julieta Hernández-Acosta, José Alberto Ávila-Funes, and et al. 2023. "The Gut Microbiota–Brain Axis during Aging, Mild Cognitive Impairment and Dementia: Role of Tau Protein, β-Amyloid and LPS in Serum and Curli Protein in Stool" Nutrients 15, no. 4: 932. https://doi.org/10.3390/nu15040932
APA StyleSánchez-Tapia, M., Mimenza-Alvarado, A., Granados-Domínguez, L., Flores-López, A., López-Barradas, A., Ortiz, V., Pérez-Cruz, C., Sánchez-Vidal, H., Hernández-Acosta, J., Ávila-Funes, J. A., Guevara-Cruz, M., Tovar, A. R., & Torres, N. (2023). The Gut Microbiota–Brain Axis during Aging, Mild Cognitive Impairment and Dementia: Role of Tau Protein, β-Amyloid and LPS in Serum and Curli Protein in Stool. Nutrients, 15(4), 932. https://doi.org/10.3390/nu15040932