
Iodide Excess Inhibits Thyroid Hormone Synthesis Pathway Involving XBP1-Mediated Regulation

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Treatments
2.2. Culture and Treatment of Cells and Organoids
2.3. Immunohistochemical and Whole-Mount Immunofluorescence Staining
2.4. Measurement of mRNA and Protein Expression
2.5. Transfection
2.6. ChIP-PCR Assay
2.7. Statistical Analysis
3. Results
3.1. Excessive Iodide Intake Downregulates Genes Involved in TH Synthesis in Mouse Thyroid
3.2. Sodium Iodide Has No Effect on Genes Involved in TH Synthesis in Cultured Cell Lines
3.3. Three-Dimensional Culture Enables Thyrocytes to Form Organoids with Follicle-like Structures
3.4. Three-Dimensional Cultured Murine Thyroid Organoids Retain the Thyroid Function
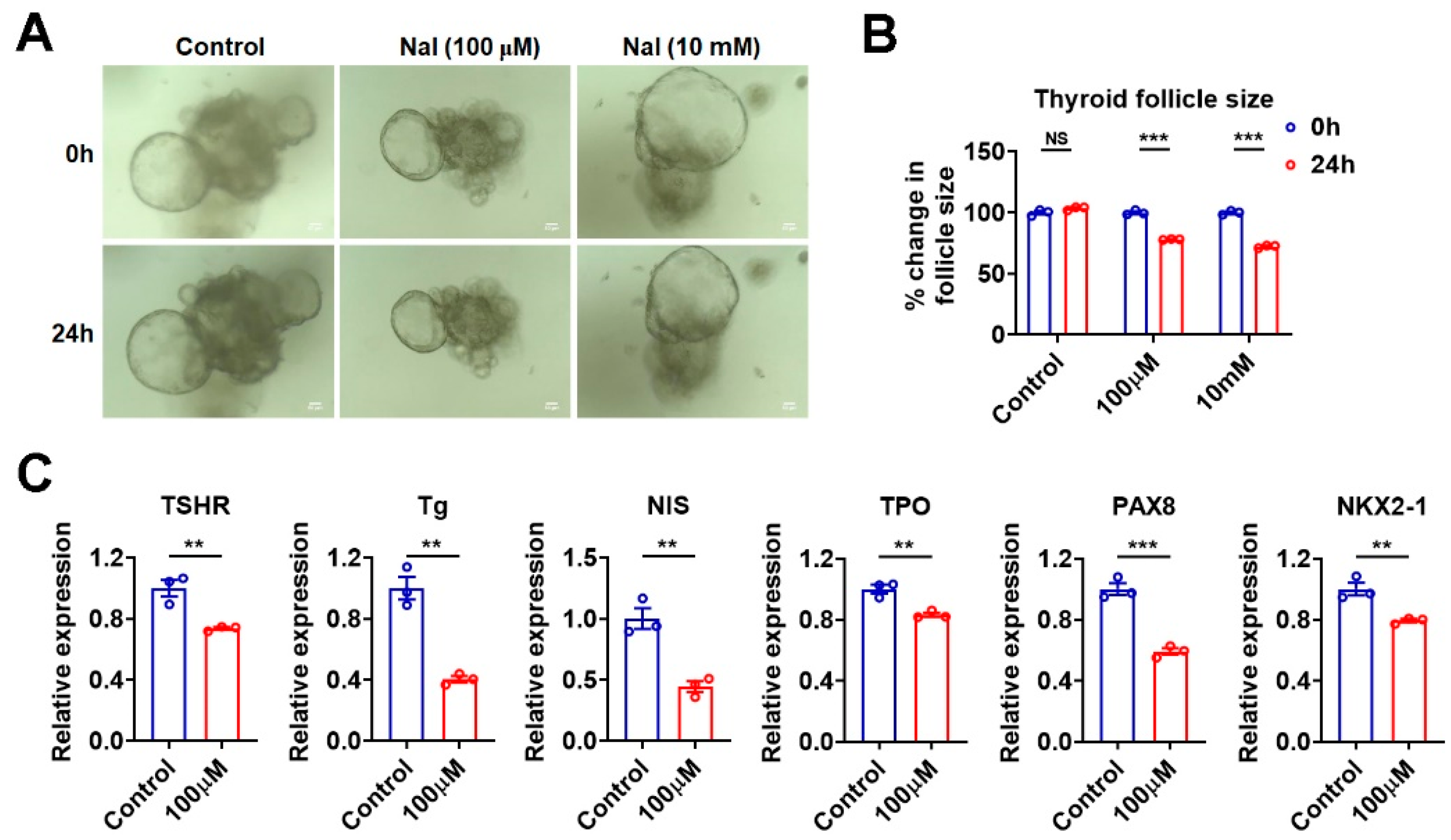
3.5. Effects of Iodide Excess on Genes Involved in TH Synthesis Can Be Mimicked in Murine Thyroid Organoids
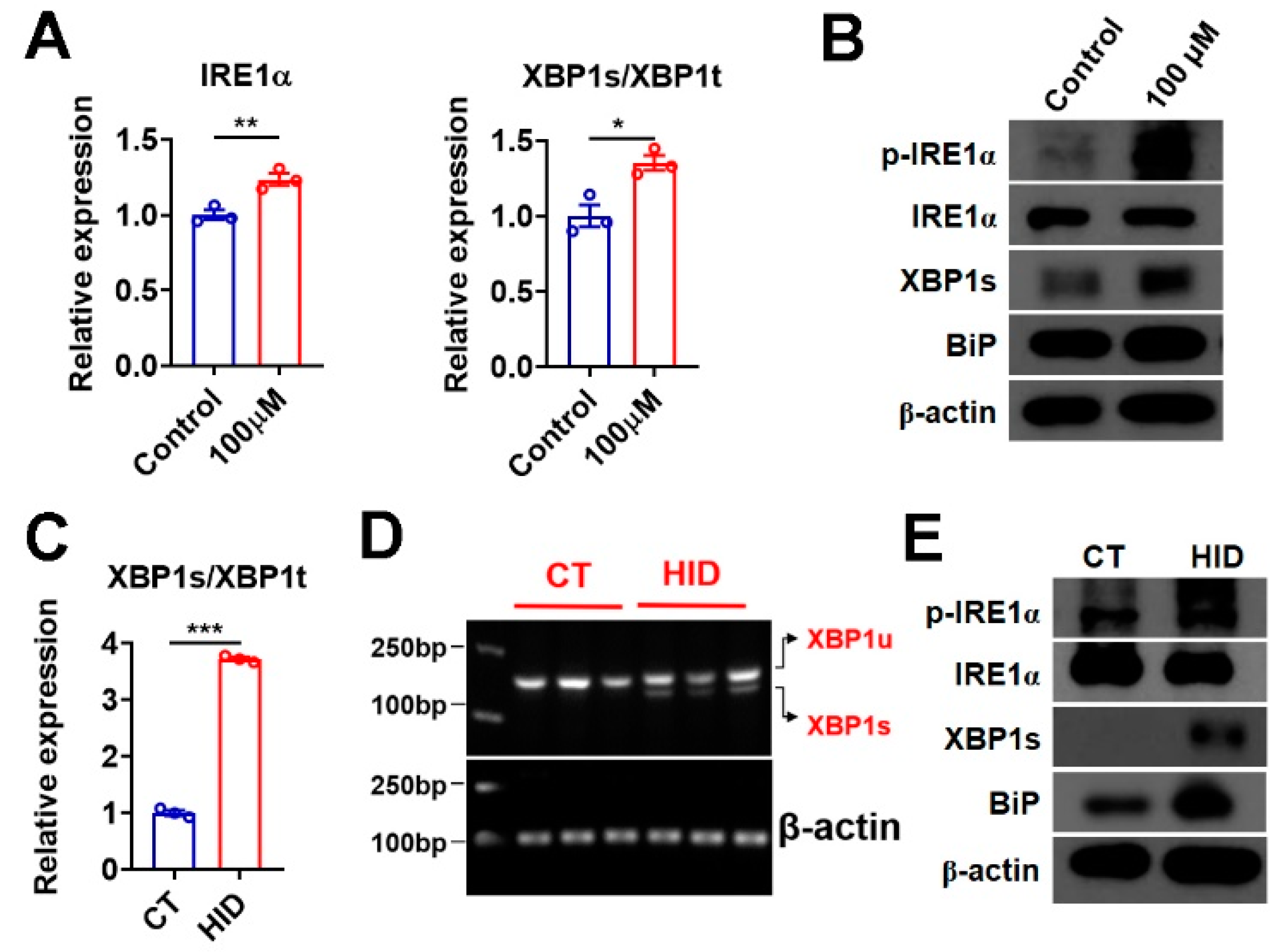
3.6. Iodide Excess Activates XBP1 in Murine Thyroid and Thyroid Organoids
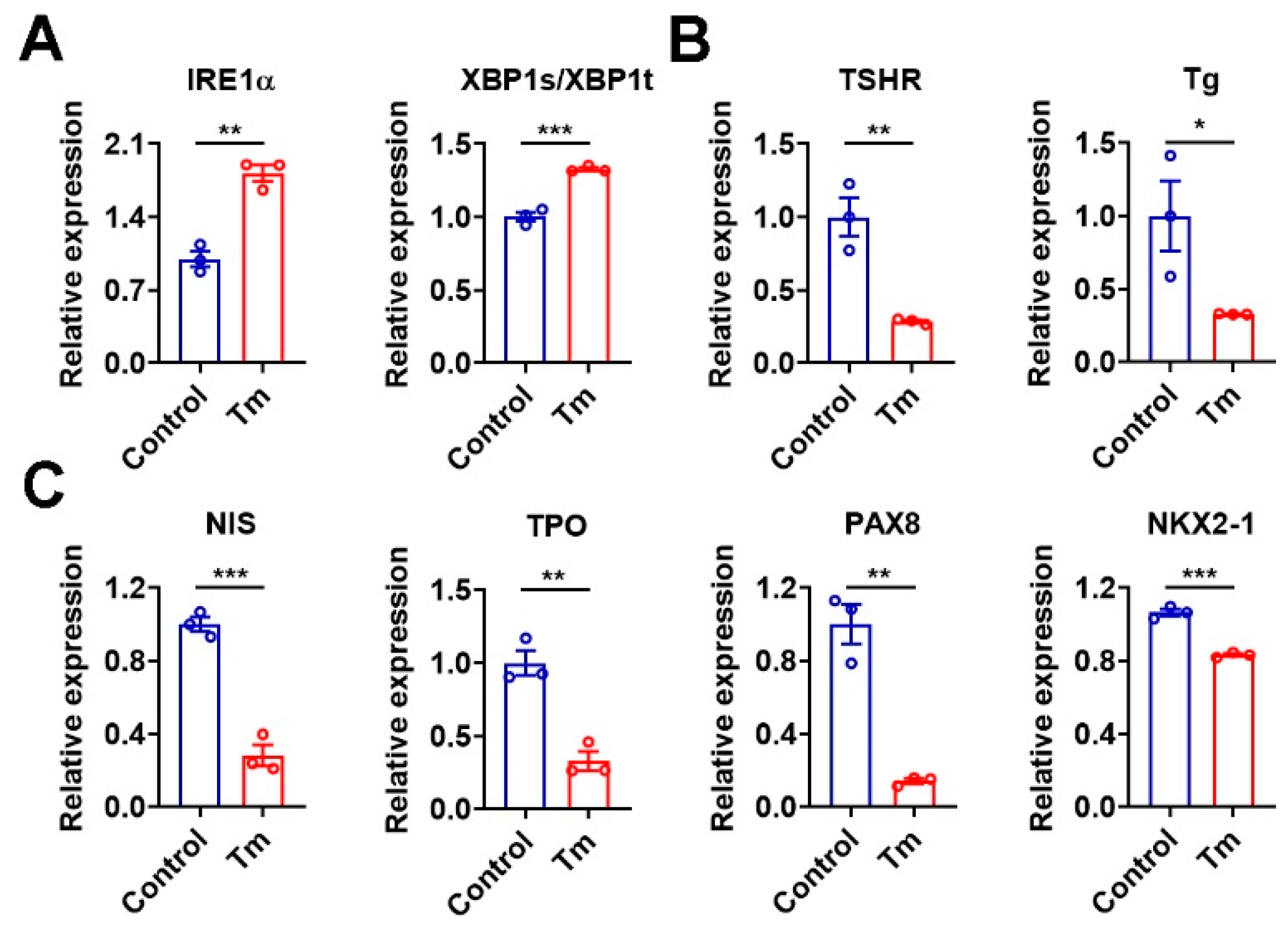
3.7. Activation of XBP1 Mimics the Iodide Excess Effect on Genes Related to TH Synthesis in Murine Thyroid Organoids
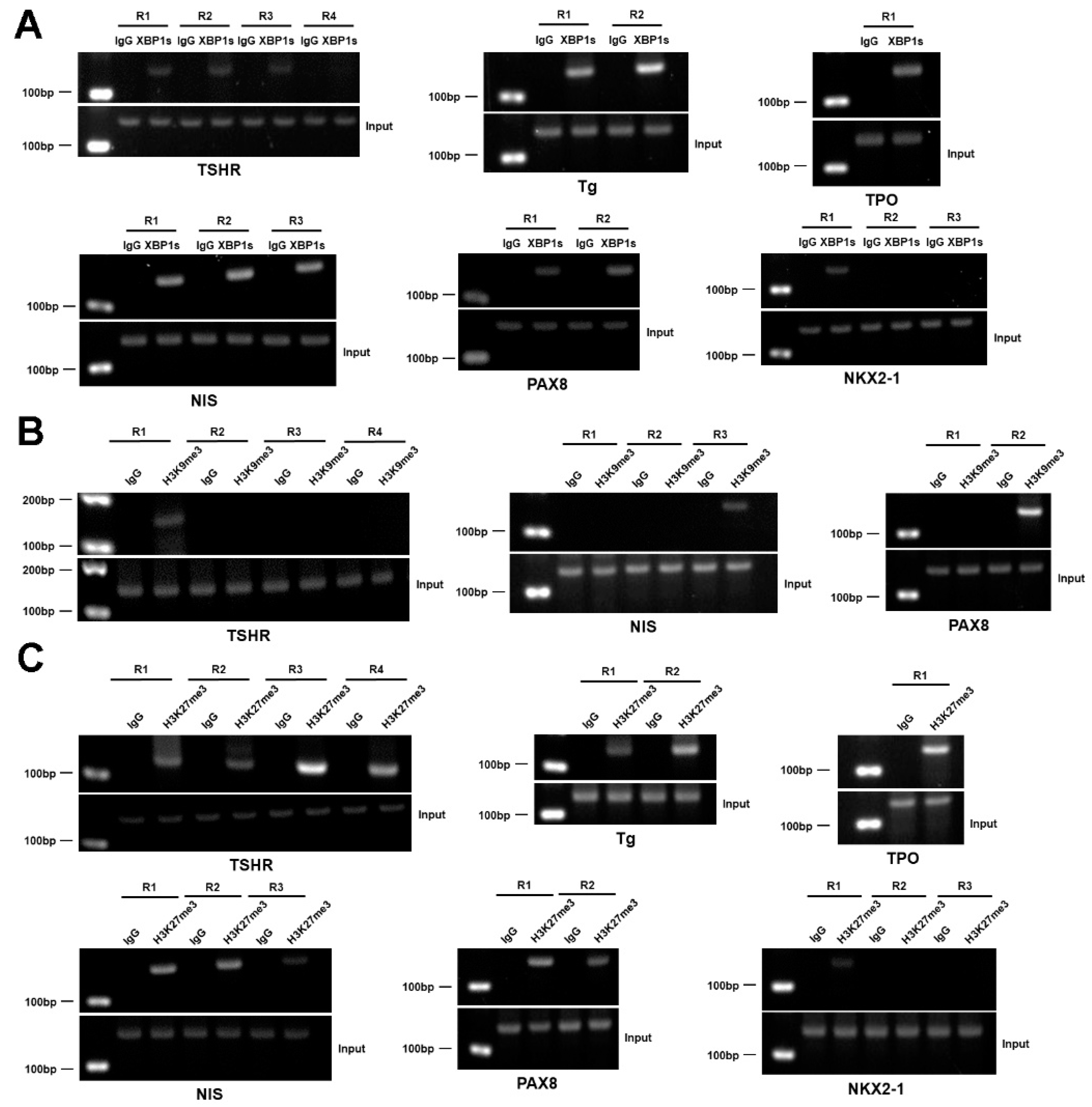
3.8. XBP1 Transcriptionally Represses the Genes Involved in the Synthesis of TH
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Markou, K.; Georgopoulos, N.; Kyriazopoulou, V.; Vagenakis, A.G. Iodine-Induced hypothyroidism. Thyroid 2001, 11, 501–510. [Google Scholar] [CrossRef]
- Pearce, E.N.; Lazarus, J.H.; Moreno-Reyes, R.; Zimmermann, M.B. Consequences of iodine deficiency and excess in pregnant women: An overview of current knowns and unknowns. Am. J. Clin. Nutr. 2016, 104 (Suppl. 3), 918S–923S. [Google Scholar]
- Leung, A.M.; Braverman, L. Consequences of excess iodine. Nat. Rev. Endocrinol. 2014, 10, 136–142. [Google Scholar] [CrossRef]
- Yao, X.; Li, M.; He, J.; Zhang, G.; Wang, M.; Ma, J.; Li, L. Effect of early acute high concentrations of iodide exposure on mitochondrial superoxide production in FRTL cells. Free Radic. Biol. Med. 2012, 52, 1343–1352. [Google Scholar]
- Serrano-Nascimento, C.; Nicola, J.P.; da Silva Teixeira, S.; Poyares, L.L.; Lellis-Santos, C.; Bordin, S.; Nunes, M.T. Excess iodide downregulates Na(+)/I(−) symporter gene transcription through activation of PI3K/Akt pathway. Mol. Cell. Endocrinol. 2016, 426, 73–90. [Google Scholar]
- Eng, P.H.; Cardona, G.R.; Fang, S.L.; Previti, M.; Alex, S.; Carrasco, N.; Braverman, L.E. Escape from the acute Wolff-Chaikoff effect is associated with a decrease in thyroid sodium/iodide symporter messenger ribonucleic acid and protein. Endocrinology 1999, 140, 3404–3410. [Google Scholar] [CrossRef]
- Schwarz, D.S.; Blower, M.D. The endoplasmic reticulum: Structure, function and response to cellular signaling. Cell. Mol. Life Sci. 2016, 73, 79–94. [Google Scholar] [CrossRef]
- Chen, S.; Novick, P.; Ferro-Novick, S. ER structure and function. Curr. Opin. Cell. Biol. 2013, 25, 428–433. [Google Scholar]
- Bravo, R.; Parra, V.; Gatica, D.; Rodriguez, A.E.; Torrealba, N.; Paredes, F.; Lavandero, S. Endoplasmic reticulum and the unfolded protein response: Dynamics and metabolic integration. Int. Rev. Cell. Mol. Biol. 2013, 301, 215–290. [Google Scholar]
- Hetz, C.; Zhang, K.; Kaufman, R.J. Mechanisms, regulation and functions of the unfolded protein response. Nat. Rev. Mol. Cell. Biol. 2020, 21, 421–438. [Google Scholar]
- Hetz, C.; Chevet, E.; Harding, H.P. Targeting the unfolded protein response in disease. Nat. Rev. Drug Discov. 2013, 12, 703–719. [Google Scholar] [CrossRef]
- Leonardi, A.; Vito, P.; Mauro, C.; Pacifico, F.; Ulianich, L.; Consiglio, E.; Di Jeso, B. Endoplasmic reticulum stress causes thyroglobulin retention in this organelle and triggers activation of nuclear factor-kappa B via tumor necrosis factor receptor-associated factor 2. Endocrinology 2002, 143, 2169–2177. [Google Scholar]
- Baryshev, M.; Sargsyan, E.; Wallin, G.; Lejnieks, A.; Furudate, S.; Hishinuma, A.; Mkrtchian, S. Unfolded protein response is involved in the pathology of human congenital hypothyroid goiter and rat non-goitrous congenital hypothyroidism. J. Mol. Endocrinol. 2004, 32, 903–920. [Google Scholar] [CrossRef]
- Wen, G.; Eder, K.; Ringseis, R. Resveratrol Alleviates the Inhibitory Effect of Tunicamycin-Induced Endoplasmic Reticulum Stress on Expression of Genes Involved in Thyroid Hormone Synthesis in FRTL-5 Thyrocytes. Int. J. Mol. Sci. 2021, 22, 4373. [Google Scholar]
- Wen, G.; Ringseis, R.; Eder, K. Endoplasmic reticulum stress inhibits expression of genes involved in thyroid hormone synthesis and their key transcriptional regulators in FRTL-5 thyrocytes. PLoS ONE 2017, 12, e0187561. [Google Scholar] [CrossRef]
- Saito, Y.; Onishi, N.; Takami, H.; Seishima, R.; Inoue, H.; Hirata, Y.; Nagano, O. Development of a functional thyroid model based on an organoid culture system. Biochem. Biophys. Res. Commun. 2018, 497, 783–789. [Google Scholar]
- Xiao, H.; Liang, J.; Liu, S.; Zhang, Q.; Xie, F.; Kong, X.; Liu, T. Proteomics and Organoid Culture Reveal the Underlying Pathogenesis of Hashimoto’s Thyroiditis. Front. Immunol. 2021, 12, 784975. [Google Scholar]
- van der Vaart, J.; Bosmans, L.; Sijbesma, S.F.; Knoops, K.; van de Wetering, W.J.; Otten, H.G.; Clevers, H. Adult mouse and human organoids derived from thyroid follicular cells and modeling of Graves’ hyperthyroidism. Proc. Natl. Acad. Sci. USA 2021, 118, e2117017118. [Google Scholar] [CrossRef]
- Yao, X.; Hou, S.; Zhang, D.; Xia, H.; Wang, Y.C.; Jiang, J.; Ying, H. Regulation of fatty acid composition and lipid storage by thyroid hormone in mouse liver. Cell Biosci. 2014, 4, 38. [Google Scholar]
- Rios, A.C.; Capaldo, B.D.; Vaillant, F.; Pal, B.; van Ineveld, R.; Dawson, C.A.; Visvader, J.E. Intraclonal Plasticity in Mammary Tumors Revealed through Large-Scale Single-Cell Resolution 3D Imaging. Cancer Cell 2019, 35, 953. [Google Scholar]
- Lemoine, N.R.; Mayall, E.S.; Jones, T.; Sheer, D.; McDermid, S.; Kendall-Taylor, P.; Wynford-Thomas, D. Characterisation of human thyroid epithelial cells immortalised in vitro by simian virus 40 DNA transfection. Br. J. Cancer 1989, 60, 897–903. [Google Scholar] [CrossRef]
- Fabien, N.; Fusco, A.; Santoro, M.; Barbier, Y.; Dubois, P.M.; Paulin, C. Description of a human papillary thyroid carcinoma cell line. Morphologic study and expression of tumoral markers. Cancer 1994, 73, 2206–2212. [Google Scholar] [CrossRef]
- Rossi, G.; Manfrin, A.; Lutolf, M.P. Progress and potential in organoid research. Nat. Rev. Genet. 2018, 19, 671–687. [Google Scholar] [CrossRef]
- Takasu, N.; Handa, Y.; Kawaoi, A.; Shimizu, Y.; Yamada, T. Effects of iodide on thyroid follicle structure and electrophysiological potentials of cultured thyroid cells. Endocrinology 1985, 117, 71–76. [Google Scholar] [CrossRef]
- Zhang, T.; Cooper, S.; Brockdorff, N. The interplay of histone modifications—Writers that read. EMBO Rep. 2015, 16, 1467–1481. [Google Scholar]
- Leoni, S.G.; Galante, P.A.; Ricarte-Filho JC, M.; Kimura, E.T. Differential gene expression analysis of iodide-treated rat thyroid follicular cell line PCCl3. Genomics 2008, 91, 356–366. [Google Scholar] [CrossRef]
- Serrano-Nascimento, C.; Calil-Silveira, J.; Goulart-Silva, F.; Nunes, M.T. New insights about the posttranscriptional mechanisms triggered by iodide excess on sodium/iodide symporter (NIS) expression in PCCl3 cells. Mol. Cell. Endocrinol. 2012, 349, 154–161. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sequence | Band Sizes |
|---|---|---|
| tdTomato P1 | CTCTGCTGCCTCCTGGCTTCT | Wild type = 330 bp tdTomato = 250 bp |
| tdTomato P2 | CGAGGCGGATCACAAGCAATA | |
| tdTomato P3 | TCAATGGGCGGGGGTCGTT | |
| TPO-Cre F | CTAGCATCTTGACGGGCTATC | TPO = 130 bp |
| TPO-Cre R | AGGCAAATTTTGGTGTACGG |
| Primer | Sequence | |
|---|---|---|
| mNIS | GCTCAGTCTCGCTCAAAACC | CGTGTGACAGGCCACATAAC |
| mTSHR | CTCTCTTACCCGAGCCACTG | TTGTCACCCGGATCTTCTTC |
| mTPO | TGACTTCCAGGAGCACACAG | GCAAGTTCAGTGATGCCAGA |
| mTg | TGTCCCACCAAGTGTGAAAA | CCAAGGAAAGCTTGTTCAGC |
| mPAX8 | CAGAAGGCGTTTGTGACAATGA | TGCACTTTGGTCCGGATGAT |
| mNKX2-1 | TCCAGCCTATCCCATCTGAACT | CAAGCGCATCTCACGTCTCA |
| mIRE1α | CGCACATGGCAGGATCAAG | TGCCCACTGCCAGCTTCT |
| mXBP1t | TGGCCGGGTCTGCTGAGTCCG | GTCCATGGGAAGATGTTCTGG |
| mXBP1s | CTGAGTCCGAATCAGGTGCAG | GTCCATGGGAAGATGTTCTGG |
| mXBP1 splicing | ACACGCTTGGGAATGGACAC | CCATGGGAAGATGTTCTGGG |
| mBip | ACTTGGGGACCACCTATTCCT | ATCGCCAATCAGACGCTCC |
| mATF4 | CCTGAACAGCGAAGTGTTGG | TGGAGAACCCATGAGGTTTCAA |
| mCHOP | CTGGAAGCCTGGTATGAGGAT | CAGGGTCAAGAGTAGTGAAGGT |
| mATF6 | ATTCTCAGCTGATGGCTGTCCAGT | TGGTAACTTCCAGGCGAAGCGTAA |
| mHerpud1 | CCAAAGCAGGAAAAGCGACA | GGATTCAGCACCCTTTGTGC |
| mβ-actin | TACCACCATGTACCCAGGCA | CTCAGGAGGAGCAATGATCTTGAT |
| hNIS | CTCTGCGGGACTTTGCAG | ATCACCACGACCTGGAAC |
| hTSHR | AGCCACTGCTGTGCTTTTAAG | CCAAAACCAATGATCTCATCC |
| hTPO | GTCTGTCACGCTGGTTATGG | CAATCACTCCGCTTGTTGGC |
| hTg | CAGCAACCAGCTTTGGTCAC | AAGAGAACAGGTCGTGCTGG |
| hXBP1t | GACGGGACCCCTAAAGTTCTG | CTTCTTTCGATCTCTGGCAGTC |
| hXBP1s | CTGAGTCCGAATCAGGTGCAG | ATCCATGGGGAGATGTTCTGG |
| hXBP1 splicing | CCTGGTTGCTGAAGAGGAGG | CCATGGGGAGATGTTCTGGAG |
| hβ-actin | GATCATTGCTCCTCCTGAGC | ACTCCTGCTTGCTGATCCAC |
| Primer | Sequence | |
|---|---|---|
| shScramble | CCGGTCCTAAGGTTAAGTCGCCCTCGC | AATTCAAAAACCTAAGGTTAAGTCGCCC |
| GAGCGAGGGCGACTTAACCTTAGGTTT | TCGCTCGAGCGAGGGCGACTTAACCTTA | |
| TTG | GGA | |
| shXBP1-1 | CCGGAGATCGAAAGAAGGCTCGAATCT | AATTCAAAAAAGATCGAAAGAAGGCTCG |
| CGAGATTCGAGCCTTCTTTCGATCTTTTT | AATCTCGAGATTCGAGCCTTCTTTCGAT | |
| TG | CT | |
| shXBP1-2 | CCGGGCGGTATTGACTCTTCAGATTCTC | AATTCAAAAAGCGGTATTGACTCTTCAG |
| GAGAATCTGAAGAGTCAATACCGCTTTT | ATTCTCGAGAATCTGAAGAGTCAATACC | |
| TG | GC | |
| shXBP1-3 | CCGGGAACAGCAAGTGGTAGATTTACT | AATTCAAAAAGAACAGCAAGTGGTAGAT |
| CGAGTAAATCTACCACTTGCTGTTCTTTT | TTACTCGAGTAAATCTACCACTTGCTGT | |
| TG | TC | |
| Primer | Sequence | |
|---|---|---|
| TSHR R1 | CTGCCATTCCCGATTCTTCC | CTGCACCCCTGGAGGATTCT |
| TSHR R2 | CAGTCGACTCAACCACCGGA | AACTTCAGCTCCTCCTGGCC |
| TSHR R3 | GAAGTTCTGCAGGACATTGG | CCCTCCTTTCCTCCTCTTTA |
| TSHR R4 | TGGAGAGAACTAATGGGAGG | GCAACCTTGACCCAAACAAG |
| Tg R1 | AAGGTGATAGAATTGCCCCA | TACTTTGGGTGAAGTGTGTG |
| Tg R2 | CATGGTGAAGAGGAGGAGAA | AATGCAGGTTGGAGGCTAGT |
| NIS R1 | AGAGAAGGGCCCTGAGATGA | ATGAACCCGAATAAGCGGAG |
| NIS R2 | GCACTTATCATGTACCAGGC | GGGAGACATTCAGAGCTCAA |
| NIS R3 | CTAGGTCTGGAGGCGGAGTC | TTCCGCTGTCCCATGCAGCC |
| TPO R1 | TGAGGAGAGACGCAGGGAAT | AGTCAGTGTGTTCCAGTGCA |
| PAX8 R1 | CTCACAATCTTACGTGTACC | TTAAGAAGGGGTCCACTGGT |
| PAX8 R2 | CAAGGAGCAAGAAAGAGGGC | GAGCAGCACCTGAAGCCATT |
| NKX2-1 R1 | AACCCTCTTCCTCCTAAGCA | TCTTGCTTCCCGGGAATAAC |
| NKX2-1 R2 | AACAGACAGACGGGCACTCA | ACCCTCGCCCATCTCCCAGA |
| NKX2-1 R3 | TTTGTTCGAGGTGGGGCAAC | TTTTTCAGTGGGCCACCCCC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, J.; Shen, S.; Yan, Y.; Liu, L.; Luo, R.; Liu, S.; Wu, Y.; Li, Y.; Jiang, J.; Ying, H. Iodide Excess Inhibits Thyroid Hormone Synthesis Pathway Involving XBP1-Mediated Regulation. Nutrients 2023, 15, 887. https://doi.org/10.3390/nu15040887
Yu J, Shen S, Yan Y, Liu L, Luo R, Liu S, Wu Y, Li Y, Jiang J, Ying H. Iodide Excess Inhibits Thyroid Hormone Synthesis Pathway Involving XBP1-Mediated Regulation. Nutrients. 2023; 15(4):887. https://doi.org/10.3390/nu15040887
Chicago/Turabian StyleYu, Jing, Siyi Shen, Ying Yan, Lingxiao Liu, Rongkui Luo, Shengnan Liu, Yuting Wu, Yuying Li, Jingjing Jiang, and Hao Ying. 2023. "Iodide Excess Inhibits Thyroid Hormone Synthesis Pathway Involving XBP1-Mediated Regulation" Nutrients 15, no. 4: 887. https://doi.org/10.3390/nu15040887
APA StyleYu, J., Shen, S., Yan, Y., Liu, L., Luo, R., Liu, S., Wu, Y., Li, Y., Jiang, J., & Ying, H. (2023). Iodide Excess Inhibits Thyroid Hormone Synthesis Pathway Involving XBP1-Mediated Regulation. Nutrients, 15(4), 887. https://doi.org/10.3390/nu15040887