

Tocotrienol in the Management of Nonalcoholic Fatty Liver Disease: A Systematic Review



Abstract
1. Introduction
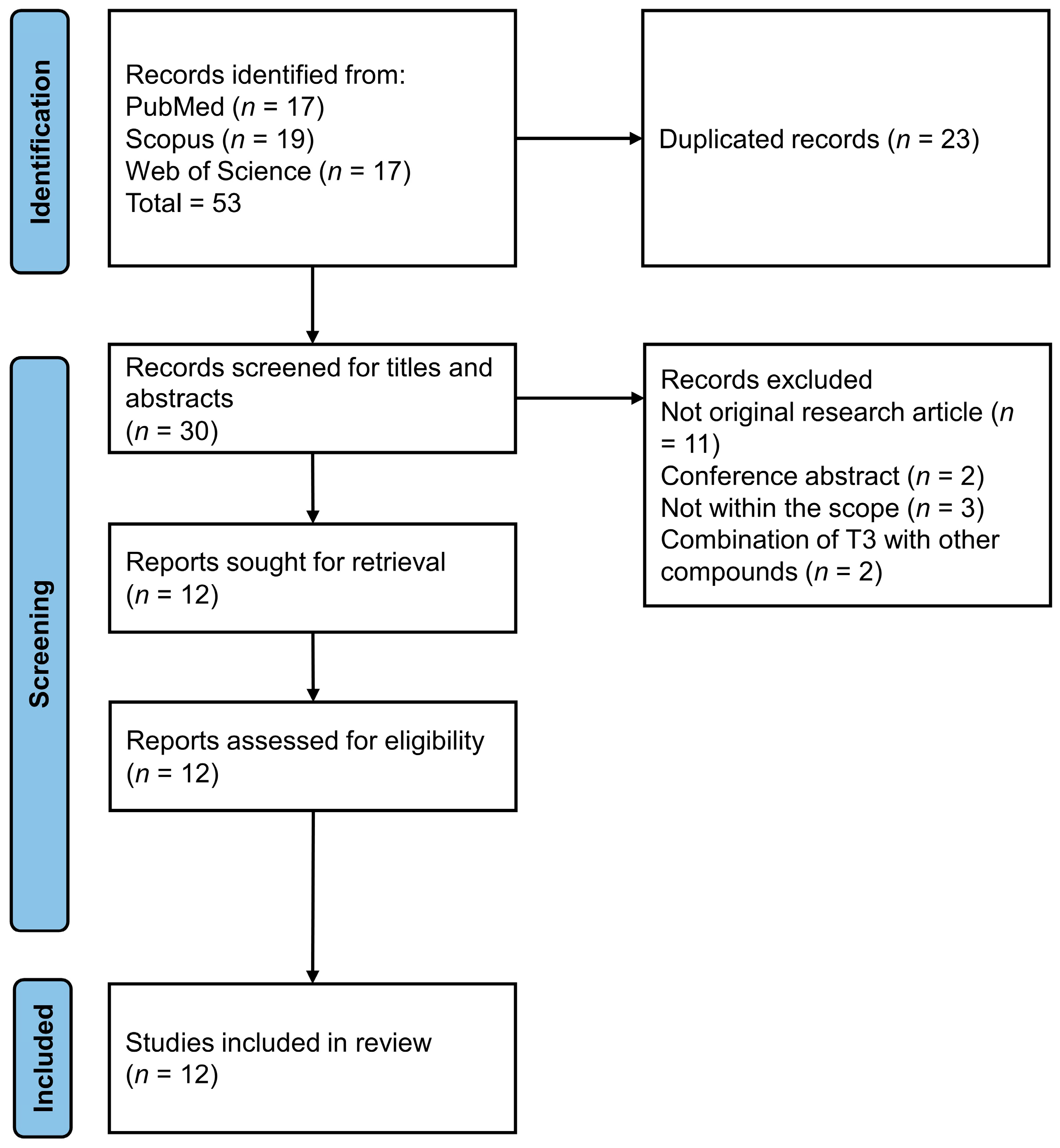
2. Methodology
3. Results

{kind=link}
{kind=link}
| Authors | Study Design/Model | Treatment | Major Findings |
|---|---|---|---|
| Preclinical studies | |||
| Yachi et al. (2013) [36] | Animal: Male SD-IGS rats (7 weeks old) Disease model: TNF-α/D-galactosamine-induced steatohepatitis (induction 2 or 5 h before sacrifice) | αTF (0.85% food mass), T3 (0.8% food mass) or mixed αTF + T3 (0.85% +0.8% food mass)-enriched high-fat diet for 4 weeks (oral) | ↓ liver TG content at 2 and 5 h in αTF + T3 group; ↔ in αTF and T3 alone group vs. negative control. ↑ liver MTP mRNA expression in αTF; ↔ liver MTP mRNA expression in T3 or αTF + T3 vs. negative control. ↓ plasma liver damage markers (AST, ALT) and TG in all treatment groups vs. negative control after 5 h. ↓ TBARS at 2 and 5 h in αTF + T3 group; ↔ in αTF or T3 vs. negative control. ↓ IL-1β at 2h in αTF and αTF + T3 group vs. negative control; ↔ IL-1β at 5h in all treated groups vs. negative control ↓ IL-6 at 2 h in all treated groups; at 5h in T3 and αTF + T3 groups vs. negative control ↓ liver TGF-β1 in αTF + T3 group vs. negative control at 2 h |
| Cells: Primary hepatocytes from male SD-IGS rats (3 weeks old) Treatment: Induction: TNFα (0.1, 0.5 nM) for 2–12 h before hepatocyte harvest. | Hepatocytes from vitamin-E-supplemented rats (doses as indicated above) | ↓ IL-1β and IL-6 mRNA expression in both αTF and αT3 after 8 h vs. negative control ↓ decline of MTP in αT3 vs. αTF and negative control. ↔ SMAD3/7/TGF-β mRNA expression in treatment vs. negative control. | |
| Allen et al. (2017) [33] | Animals: Male C57BL/6J mice (age: 6 weeks old) Disease model: High-fat-diet-induced steatohepatitis | 400 or 1600 mg/kg diet δT3 (or 28 or 112 mg/kg bw) for 14 weeks (oral) | ↑ glucose tolerance; ↓ hepatic steatosis (TG droplets and macrophage infiltration), serum TG, fat cell size and macrophage infiltration vs. negative control ↔ BW, fat pad weight vs. negative control ↓ mRNA and protein expression of pro-inflammatory adipokines (TNF-α, IL-6, MCP-1 & leptin); ↑ expression of anti-inflammatory adipokines (IL-10) for 400 mg/kg δT3 vs. negative control. ↑ markers of fatty acid oxidation (mRNA level of CPT1A/2 (1600 mg/kg only), Forkhead box A2) vs. negative control in adipose tissue ↑ markers of fatty acid oxidation (mRNA level of PPARα, PPAR δ (400 mg/kg only) and CPT2 (400 mg/kg only)); ↔ in CTP1A vs. negative control in the liver tissue. ↓ markers of fatty acid synthesis (mRNA level of fatty acid synthase and acetyl-CoA carboxylase-1) in adipose tissue vs. negative control ↓ markers of fatty acid synthesis (FASN, SCD-1 (400 mg/kg only), pyruvate kinase (1600 mg/kg only) and carbohydrate-responsive element-binding (1600 mg/kg only)) in liver tissue vs. negative control |
| Wong et al. (2017) [46] | Animals: Male Wistar rats (age: 9–10 weeks old) Disease model: HC- or HCHF-diet-induced metabolic syndrome | 85 mg/kg bw of αTF, αT3, γT3, δT3 for 8 weeks (oral) | ↓ total fat mass, abdominal circumference, adiposity index and retroperitoneal and epididymal fat pads vs. HCHF control in δT3 ↓ total cholesterol, NEFA, TG in δT3 vs. HCHF control, αTF, αT3 and γT3; ↓ plasma NEFA in αTF, αT3, γT3 vs. HCHF control. ↓ lipid droplets, portal inflammatory cell infiltration and ALT, AST level in all treated groups ↓ fasting plasma glucose levels, postprandial blood and glucose AUC in OGTT test in δT3 |
| Kim et al. (2018) [37] | Animals: Male C57BL/6J mice (age: 6 weeks old) Disease model: High fat (45%), cholesterol (0.2%) and sucrose (in drinking water) (HFCS)-diet-induced NAFLD/NASH | γT3 (0.1% food mass) for 12 weeks (oral) | ↓ body weight; ↑ daytime activity, energy expenditure and respiratory exchange ratio vs. negative control ↓ total cholesterol, LDL, fasting glucose, fasting insulin, plasma lactate and higher GSH/GSSG ratio and glucose clearance vs. negative control ↓ TG content and lipid droplets in liver vs. negative control ↓ pro-inflammatory cytokines mRNA expression (MCP-1, CD11c, TNF-α, NLRP3 and IL-1β) in liver ↓ de novo lipogenic gene expression (PPARγ, SREBP1C, FAS, DGAT2, SCD1 and LPL, DNL, ACC), ER stress protein expression (BiP, CHOP, p-JNK, peIF2α and p-p38) and fibrosis-related gene expression (α-SMA, TIMP-1, TGF- α, COL1a1 and HDAC9) in the liver vs. negative control ↓ mRNA expression (MCP1, IL-1β, IL-18 and IL-6) and protein expression (IκBα, p-p38 and p38) of inflammatory markers in the pancreas vs. negative control. ↓ mRNA expression (F4/80, MCP-1, TNF-α, IL-1β and IL-18) and protein expression (F4/80) of pro-inflammatory markers in the epididymal fat. |
| Animals: Male C57BL/6J mice (age: 6 weeks old) Disease model: methionine- and choline-deficient diet (MCD)-induced NAFLD/NASH | γT3 (0.1% food mass) for 5 weeks (oral) | ↔ total hepatic TG content; ↓ fibrosis vs. negative control ↓ hepatic stress and fibrosis with minimal impact on steatosis vs. negative control ↓ lipogenesis, ER stress, inflammation and fibrosis vs. negative control | |
| Animal: Male CHOP-knockout mice (age: 6 weeks old) Disease model: MCD-induced NAFLD/NASH | γT3 (0.1% food mass) for 5 weeks (oral) | ↑ protective effects of γT3 due to CHOP deletion vs. negative control ↓ hepatic fibrosis and inflammation vs. negative control | |
| Wong et al. (2020) [28] | Animals: Male Wistar rats (age: 12 weeks old) Disease model: HCHF-diet-induced NAFLD | 60 or 100 mg/kg bw of AnT3 (with 16% γT3 and 84% δT3) or pT3 (21.9% αTF, 24.7% αT3, 4.5% βT3, 36.9% γT3 and 12.0% δT3) for 12 weeks (oral) | ↓ liver index in 60 mg/kg pT3 ↓ liver TLR 2 and IL-10 expression and serum CRP level in all T3 treatments except 100 mg/kg pT3 vs. negative control ↔ TLR4 expression vs. negative control ↓ total NFκB in all T3 treatments except 60 mg/kg pT3 vs. negative control ↑ Cytoplasmic p-NFκB/total NFκB in the AnT3 groups vs. negative control ↓ MDA level and ↑ SOD activity in all T3 groups; ↑ GPx activity in AnT3 and 60 mg/kg pT3; ↑ GSH content in 60 mg/kg pT3 vs. negative control ↓ steatosis score in 100 mg/kg pT3 100 vs. negative control ↔ % of red oil stain with treatment vs. negative control |
| Goon et al. (2021) [41] | Animals: Male B6.Cg-LepOb/J mice (age: 8 weeks old) Disease model: NAFLD induced using genetically modified mice through HFD | 200 mg/kg TRF bw (72% T3 and 28% TF) or eTRF with palm kernel oil as the carrier (81% T3 and 19% TF) for 6 weeks (oral) | ↓ Bile acids, lysine, arachidonic acid and sphingolipids; ↑ xanthine and hypoxanthine vs. negative control ↑ FXR IHC score vs. carrier and negative control ↑ FXR expression in eTRF vs. negative control ↓ NAFLD activity score vs. negative control |
| Goon et al. (2022) [34] | Animals: ICR mice (age: 6 weeks old) Disease model: HFD-induced NAFLD | 200 mg/kg bw TRF (80% T3 and 20% TF) for 10 weeks (oral) | ↔ body weight, waist circumference, random blood glucose vs. negative control↓ NAFLD activity score vs. negative control ↓ steatosis of parenchymal, hepatocyte ballooning and lobular inflammation vs. negative control ↓ fatty liver macroscopically and microscopically vs. negative control |
| Noichi et al. (2022) [38] | Animals: Male C57BL/6J mice (age: 5 weeks old) Disease model: CDAHFD-induced NAFLD/NASH | αTF (4 mg fixed dose) and T3 (32% αT3, 5% βT3, 48% γT3 and 15% δT3; 4 mg fixed dose) for 2 weeks (oral) | ↔ liver weight, liver/body weight ratio, adipose tissue weight (epididymal, perirenal, mesenteric) vs. negative control ↑ TG in T3 group vs. negative control; ↑ ALT level in αTF vs. T3 and negative control; ↑AST level αTF vs. T3. ↓ F4/80, TNFα and Col4a1 in T3. ↔ TNF-α, Col1a1, α-SMA vs. negative control. ↓ MMP3 but not MMP13 in T3. ↓ dense collagen fibres vs. negative control |
| Human randomised controlled trials | |||
| Magosso et al. (2013) [42] | Participants: Adults of both genders (age: 35 yrs and above) Disease: mild untreated hypercholesterolaemia and ultrasound-proven NAFLD | Treatment: 200 mg mixed tocotrienol (61.5 mg αT3, 112.8 mg γT3, 25.7 mg δT3, 61.1 mg αTF) twice daily for 1 year (oral). Placebo control: content not disclosed. | Normalisation of hepatic echogenic response vs. placebo Worsening of steatotic grade in 2 cases in the placebo group, none in the T3 group. ↑ decline of ApoB vs. placebo ↑ circulating T3 level vs. placebo |
| Pervez et al. (2018) [43] | Participants: Adults of both genders (age: > 20 yrs) Disease: ultrasound-proven fatty liver disease | Treatment: 300 mg AnT3 (90% δT3 and 10% γT3) twice daily for 12 weeks (oral). Placebo control: sucrose | ↓ serum AST, hs-CRP, MDA and FLI score vs. placebo ↔ hepatic steatosis (ultrasound) vs. placebo. No adverse effect detected. |
| Pervez et al. (2020) [45] | Participants: Adults of both genders (age: > 20 yrs) Disease: ultrasound-proven fatty liver disease | Treatment: 300 mg AnT3 (90% δT3 and 10% γT3) twice daily for 24 weeks (oral). Placebo control: sucrose | ↓ FLI, HOMA-IR, hs-CRP, IL-6, TNF-α, MDA, AST, ALT. ↓ expression of miR-122-5p, miR-34a-5p and miR-375-3p No adverse events reported. |
| Pervez et al. (2022) [44] | Participants: Adults of both genders (age: 20–70 yrs) Disease: NAFLD | Treatment: 300 mg δT3 or 268 mg αTF twice daily for 24 and 48 weeks (oral). Placebo control: no | Improved FLI, liver-to-spleen CT attenuation ratio, HOMA-IR, serum hs-CRP and malondialdehyde ratio for both groups compared to baseline, but not significant between them. ↓ body weight, IL6, TNFα, leptin, cytokeratin-18 fragment M30; ↑ adiponectin in δT3 vs. αTF at 48 weeks |
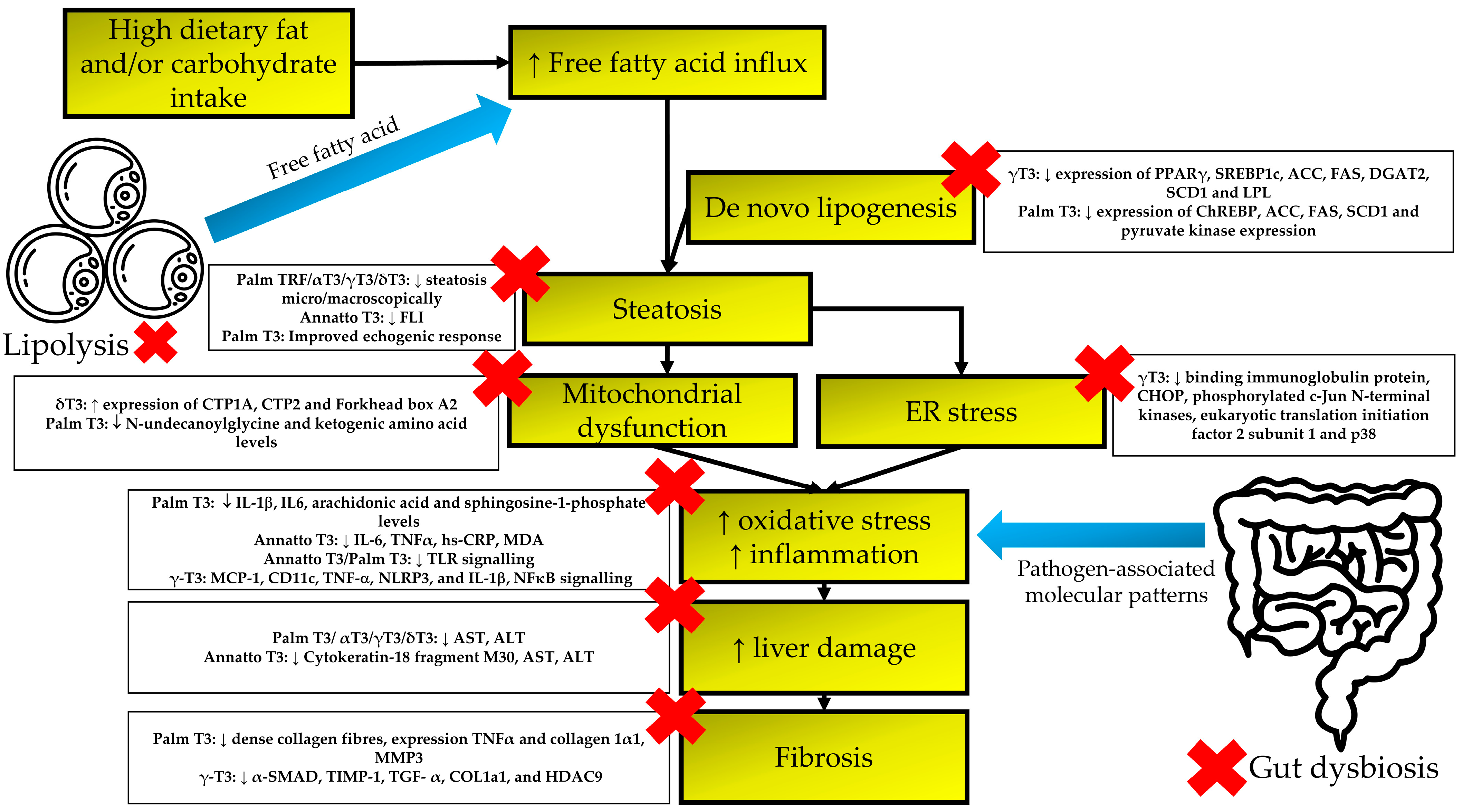
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviation
References
- Friedman, S.L.; Neuschwander-Tetri, B.A.; Rinella, M.; Sanyal, A.J. Mechanisms of NAFLD development and therapeutic strategies. Nat. Med. 2018, 24, 908–922. [Google Scholar] [CrossRef] [PubMed]
- Tomeno, W.; Imajo, K.; Takayanagi, T.; Ebisawa, Y.; Seita, K.; Takimoto, T.; Honda, K.; Kobayashi, T.; Nogami, A.; Kato, T.; et al. Complications of Non-Alcoholic Fatty Liver Disease in Extrahepatic Organs. Diagnostics 2020, 10, 912. [Google Scholar] [CrossRef]
- Mantovani, A.; Scorletti, E.; Mosca, A.; Alisi, A.; Byrne, C.D.; Targher, G. Complications, morbidity and mortality of nonalcoholic fatty liver disease. Metabolism 2020, 111, 154170. [Google Scholar] [CrossRef] [PubMed]
- Piazzolla, V.A.; Mangia, A. Noninvasive Diagnosis of NAFLD and NASH. Cells 2020, 9, 5. [Google Scholar] [CrossRef] [PubMed]
- Alberti, K.G.; Zimmet, P.; Shaw, J. Metabolic syndrome—A new world-wide definition. A Consensus Statement from the International Diabetes Federation. Diabet. Med. 2006, 23, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Grundy, S.M.; Brewer, H.B., Jr.; Cleeman, J.I.; Smith, S.C., Jr.; Lenfant, C.; National Heart, L.; Blood, I.; American Heart, A. Definition of metabolic syndrome: Report of the National Heart, Lung, and Blood Institute/American Heart Association conference on scientific issues related to definition. Arterioscler Thromb. Vasc. Biol. 2004, 24, e13–e18. [Google Scholar] [CrossRef]
- Godoy-Matos, A.F.; Silva Júnior, W.S.; Valerio, C.M. NAFLD as a continuum: From obesity to metabolic syndrome and diabetes. Diabetol. Metab. Syndr. 2020, 12, 60. [Google Scholar] [CrossRef]
- Ballestri, S.; Zona, S.; Targher, G.; Romagnoli, D.; Baldelli, E.; Nascimbeni, F.; Roverato, A.; Guaraldi, G.; Lonardo, A. Nonalcoholic fatty liver disease is associated with an almost twofold increased risk of incident type 2 diabetes and metabolic syndrome. Evidence from a systematic review and meta-analysis. J. Gastroenterol. Hepatol. 2016, 31, 936–944. [Google Scholar] [CrossRef]
- Riazi, K.; Azhari, H.; Charette, J.H.; Underwood, F.E.; King, J.A.; Afshar, E.E.; Swain, M.G.; Congly, S.E.; Kaplan, G.G.; Shaheen, A.-A. The prevalence and incidence of NAFLD worldwide: A systematic review and meta-analysis. Lancet Gastroenterol. Hepatol. 2022, 7, 851–861. [Google Scholar] [CrossRef]
- Saklayen, M.G. The Global Epidemic of the Metabolic Syndrome. Curr. Hypertens. Rep. 2018, 20, 12. [Google Scholar] [CrossRef]
- Paschos, P.; Paletas, K. Non alcoholic fatty liver disease and metabolic syndrome. Hippokratia 2009, 13, 9–19. [Google Scholar] [PubMed]
- Polyzos, S.A.; Kountouras, J.; Zavos, C. The multi-hit process and the antagonistic roles of tumor necrosis factor-alpha and adiponectin in non alcoholic fatty liver disease. Hippokratia 2009, 13, 127, author reply 128. [Google Scholar]
- Buzzetti, E.; Pinzani, M.; Tsochatzis, E.A. The multiple-hit pathogenesis of non-alcoholic fatty liver disease (NAFLD). Metabolism 2016, 65, 1038–1048. [Google Scholar] [CrossRef]
- Marchisello, S.; Di Pino, A.; Scicali, R.; Urbano, F.; Piro, S.; Purrello, F.; Rabuazzo, A.M. Pathophysiological, Molecular and Therapeutic Issues of Nonalcoholic Fatty Liver Disease: An Overview. Int. J. Mol. Sci. 2019, 20, 1948. [Google Scholar] [CrossRef]
- Teng, T.; Qiu, S.; Zhao, Y.; Zhao, S.; Sun, D.; Hou, L.; Li, Y.; Zhou, K.; Yu, X.; Yang, C.; et al. Pathogenesis and Therapeutic Strategies Related to Non-Alcoholic Fatty Liver Disease. Int. J. Mol. Sci. 2022, 23, 7841. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.; Pessin, J.E. Adipokines mediate inflammation and insulin resistance. Front. Endocrinol. 2013, 4, 71. [Google Scholar] [CrossRef] [PubMed]
- Attia, S.L.; Softic, S.; Mouzaki, M. Evolving Role for Pharmacotherapy in NAFLD/NASH. Clin. Transl. Sci. 2021, 14, 11–19. [Google Scholar] [CrossRef]
- Sumida, Y.; Yoneda, M.; Ogawa, Y.; Yoneda, M.; Okanoue, T.; Nakajima, A. Current and new pharmacotherapy options for non-alcoholic steatohepatitis. Expert Opin. Pharmacother. 2020, 21, 953–967. [Google Scholar] [CrossRef]
- Abdel-Maboud, M.; Menshawy, A.; Menshawy, E.; Emara, A.; Alshandidy, M.; Eid, M. The efficacy of vitamin E in reducing non-alcoholic fatty liver disease: A systematic review, meta-analysis, and meta-regression. Therap. Adv. Gastroenterol. 2020, 13, 1756284820974917. [Google Scholar] [CrossRef]
- Amanullah, I.; Khan, Y.H.; Anwar, I.; Gulzar, A.; Mallhi, T.H.; Raja, A.A. Effect of vitamin E in non-alcoholic fatty liver disease: A systematic review and meta-analysis of randomised controlled trials. Postgrad. Med. J. 2019, 95, 601. [Google Scholar] [CrossRef]
- Vadarlis, A.; Antza, C.; Bakaloudi, D.R.; Doundoulakis, I.; Kalopitas, G.; Samara, M.; Dardavessis, T.; Maris, T.; Chourdakis, M. Systematic review with meta-analysis: The effect of vitamin E supplementation in adult patients with non-alcoholic fatty liver disease. J. Gastroenterol. Hepatol. 2021, 36, 311–319. [Google Scholar] [CrossRef]
- Sato, K.; Gosho, M.; Yamamoto, T.; Kobayashi, Y.; Ishii, N.; Ohashi, T.; Nakade, Y.; Ito, K.; Fukuzawa, Y.; Yoneda, M. Vitamin E has a beneficial effect on nonalcoholic fatty liver disease: A meta-analysis of randomized controlled trials. Nutrition 2015, 31, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Chin, K.Y.; Pang, K.L.; Soelaiman, I.N. Tocotrienol and Its Role in Chronic Diseases. Adv. Exp. Med. Biol. 2016, 928, 97–130. [Google Scholar]
- Chin, K.-Y.; Ima-Nirwana, S. Vitamin E as an Antiosteoporotic Agent via Receptor Activator of Nuclear Factor Kappa-B Ligand Signaling Disruption: Current Evidence and Other Potential Research Areas. Evid.-Based Complement. Alternat. Med. 2012, 2012, 747020. [Google Scholar] [CrossRef]
- Frega, N.; Mozzon, M.; Bocci, F. Identification and estimation of tocotrienols in the annatto lipid fraction by gas chromatography-mass spectrometry. J. Amer. Oil Chem. Soc. 1998, 75, 1723–1727. [Google Scholar] [CrossRef]
- Chin, K.Y.; Ima-Nirwana, S. The biological effects of tocotrienol on bone: A review on evidence from rodent models. Drug Des. Dev. Ther. 2015, 9, 2049–2061. [Google Scholar] [CrossRef]
- Ahsan, H.; Ahad, A.; Siddiqui, W.A. A review of characterization of tocotrienols from plant oils and foods. J. Chem. Biol. 2015, 8, 45–59. [Google Scholar] [CrossRef]
- Wong, S.K.; Kamisah, Y.; Mohamed, N.; Muhammad, N.; Masbah, N.; Fahami, N.A.M.; Mohamed, I.N.; Shuid, A.N.; Saad, Q.M.; Abdullah, A.; et al. Potential Role of Tocotrienols on Non-Communicable Diseases: A Review of Current Evidence. Nutrients 2020, 12, E259. [Google Scholar] [CrossRef]
- Pang, K.L.; Chin, K.Y. The Role of Tocotrienol in Protecting Against Metabolic Diseases. Molecules 2019, 24, 923. [Google Scholar] [CrossRef]
- Wong, S.K.; Chin, K.-Y.; Suhaimi, F.H.; Ahmad, F.; Ima-Nirwana, S. The effects of palm tocotrienol on metabolic syndrome and bone loss in male rats induced by high-carbohydrate high-fat diet. J. Funct. Foods 2018, 44, 246–254. [Google Scholar] [CrossRef]
- Wong, S.K.; Chin, K.Y.; Suhaimi, F.H.; Ahmad, F.; Ima-Nirwana, S. Exploring the potential of tocotrienol from Bixa orellana as a single agent targeting metabolic syndrome and bone loss. Bone 2018, 116, 8–21. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef] [PubMed]
- Allen, L.; Ramalingam, L.; Menikdiwela, K.; Scoggin, S.; Shen, C.L.; Tomison, M.D.; Kaur, G.; Dufour, J.M.; Chung, E.; Kalupahana, N.S.; et al. Effects of delta-tocotrienol on obesity-related adipocyte hypertrophy, inflammation and hepatic steatosis in high-fat-fed mice. J. Nutr. Biochem. 2017, 48, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Goon, M.D.M.E.; Zulkanain, N.I.; Kadir, S.H.S.A.; Rahim, S.A.; Mazlan, M.; Latip, N.A.; Aziz, M.A.; Noor, N.M. Palm tocotrienol rich fraction with palm kernel oil supplementation prevents development of liver steatosis in high fat diet ICR mice. Transl. Gastroenterol. Hepatol. 2022, 7, 2. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.K.; Chin, K.Y.; Ahmad, F.; Ima-Nirwana, S. Regulation of inflammatory response and oxidative stress by tocotrienol in a rat model of non-alcoholic fatty liver disease. J. Funct. Foods 2020, 74, 104209. [Google Scholar] [CrossRef]
- Yachi, R.; Muto, C.; Ohtaka, N.; Aoki, Y.; Koike, T.; Igarashi, O.; Kiyose, C. Effects of tocotrienol on tumor necrosis factor-α/d-galactosamine-induced steatohepatitis in rats. J. Clin. Biochem. Nutr. 2013, 52, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Natarajan, S.K.; Chung, S. Gamma-Tocotrienol Attenuates the Hepatic Inflammation and Fibrosis by Suppressing Endoplasmic Reticulum Stress in Mice. Mol. Nutr. Food Res. 2018, 62, e1800519. [Google Scholar] [CrossRef]
- Noichi, J.; Ishiakawa, T.; Ichi, I.; Fujiwara, Y. Effect of tocotrienol on the primary progression of nonalcoholic steatohepatitis in a mouse model. J. Clin. Biochem. Nutr. 2022, 70, 140–146. [Google Scholar] [CrossRef]
- Rinella, M.E.; Green, R.M. The methionine-choline deficient dietary model of steatohepatitis does not exhibit insulin resistance. J. Hepatol. 2004, 40, 47–51. [Google Scholar] [CrossRef]
- Arab, J.P.; Arrese, M.; Trauner, M. Recent Insights into the Pathogenesis of Nonalcoholic Fatty Liver Disease. Annu. Rev. Pathol. Mech. Dis. 2018, 13, 321–350. [Google Scholar] [CrossRef]
- Goon, D.E.; Ab-Rahim, S.; Mohd Sakri, A.H.; Mazlan, M.; Tan, J.K.; Abdul Aziz, M.; Mohd Noor, N.; Ibrahim, E.; Sheikh Abdul Kadir, S.H. Untargeted serum metabolites profiling in high-fat diet mice supplemented with enhanced palm tocotrienol-rich fraction using UHPLC-MS. Sci. Rep. 2021, 11, 21001. [Google Scholar] [CrossRef] [PubMed]
- Magosso, E.; Ansari, M.A.; Gopalan, Y.; Shuaib, I.L.; Wong, J.W.; Khan, N.A.; Abu Bakar, M.R.; Ng, B.H.; Yuen, K.H. Tocotrienols for normalisation of hepatic echogenic response in nonalcoholic fatty liver: A randomised placebo-controlled clinical trial. Nutr. J. 2013, 12, 166. [Google Scholar] [CrossRef]
- Pervez, M.A.; Khan, D.A.; Ijaz, A.; Khan, S. Effects of Delta-tocotrienol Supplementation on Liver Enzymes, Inflammation, Oxidative stress and Hepatic Steatosis in Patients with Nonalcoholic Fatty Liver Disease. Turk. J. Gastroenterol. 2018, 29, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Pervez, M.A.; Khan, D.A.; Mirza, S.A.; Slehria, A.U.R.; Nisar, U.; Aamir, M. Comparison of delta-tocotrienol and alpha-tocopherol effects on hepatic steatosis and inflammatory biomarkers in patients with non-alcoholic fatty liver disease: A randomized double-blind active-controlled trial. Complement. Ther. Med. 2022, 70, 102866. [Google Scholar] [CrossRef] [PubMed]
- Pervez, M.A.; Khan, D.A.; Slehria, A.U.R.; Ijaz, A. Delta-tocotrienol supplementation improves biochemical markers of hepatocellular injury and steatosis in patients with nonalcoholic fatty liver disease: A randomized, placebo-controlled trial. Complement. Ther. Med. 2020, 52, 102494. [Google Scholar] [CrossRef]
- Wong, W.Y.; Ward, L.C.; Fong, C.W.; Yap, W.N.; Brown, L. Anti-inflammatory gamma- and delta-tocotrienols improve cardiovascular, liver and metabolic function in diet-induced obese rats. Eur. J. Nutr. 2017, 56, 133–150. [Google Scholar] [CrossRef]
- Van Herck, M.A.; Vonghia, L.; Francque, S.M. Animal Models of Nonalcoholic Fatty Liver Disease-A Starter’s Guide. Nutrients 2017, 9, 1072. [Google Scholar] [CrossRef]
- Sanders, F.W.; Griffin, J.L. De novo lipogenesis in the liver in health and disease: More than just a shunting yard for glucose. Biol. Rev. Camb. Philos. Soc. 2016, 91, 452–468. [Google Scholar] [CrossRef]
- Lee, Y.K.; Park, J.E.; Lee, M.; Hardwick, J.P. Hepatic lipid homeostasis by peroxisome proliferator-activated receptor gamma 2. Liver Res. 2018, 2, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Montandon, S.A.; Somm, E.; Loizides-Mangold, U.; de Vito, C.; Dibner, C.; Jornayvaz, F.R. Multi-technique comparison of atherogenic and MCD NASH models highlights changes in sphingolipid metabolism. Sci. Rep. 2019, 9, 16810. [Google Scholar] [CrossRef]
- Talley, J.; Mohiuddin, S. Biochemistry, Fatty Acid Oxidation. Available online: https://www.ncbi.nlm.nih.gov/books/NBK556002/ (accessed on 27 December 2022).
- Console, L.; Scalise, M.; Giangregorio, N.; Tonazzi, A.; Barile, M.; Indiveri, C. The Link Between the Mitochondrial Fatty Acid Oxidation Derangement and Kidney Injury. Front. Physiol. 2020, 11, 794. [Google Scholar] [CrossRef]
- Simões, I.C.M.; Fontes, A.; Pinton, P.; Zischka, H.; Wieckowski, M.R. Mitochondria in non-alcoholic fatty liver disease. Int. J. Biochem. Cell Biol. 2018, 95, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Prasun, P.; Ginevic, I.; Oishi, K. Mitochondrial dysfunction in nonalcoholic fatty liver disease and alcohol related liver disease. Transl. Gastroenterol. Hepatol. 2021, 6, 4. [Google Scholar] [CrossRef]
- Henkel, A.S. Unfolded Protein Response Sensors in Hepatic Lipid Metabolism and Nonalcoholic Fatty Liver Disease. Semin. Liver Dis. 2018, 38, 320–332. [Google Scholar] [CrossRef]
- Hrncir, T.; Hrncirova, L.; Kverka, M.; Hromadka, R.; Machova, V.; Trckova, E.; Kostovcikova, K.; Kralickova, P.; Krejsek, J.; Tlaskalova-Hogenova, H. Gut Microbiota and NAFLD: Pathogenetic Mechanisms, Microbiota Signatures, and Therapeutic Interventions. Microorganisms 2021, 9, 957. [Google Scholar] [CrossRef]
- Deguine, J.; Barton, G.M. MyD88: A central player in innate immune signaling. F1000Prime Rep. 2014, 6, 97. [Google Scholar] [CrossRef]
- Lingappan, K. NF-κB in Oxidative Stress. Curr. Opin. Toxicol. 2018, 7, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Saikh, K.U. MyD88 and beyond: A perspective on MyD88-targeted therapeutic approach for modulation of host immunity. Immunol. Res. 2021, 69, 117–128. [Google Scholar] [CrossRef]
- Leung, C.; Rivera, L.; Furness, J.B.; Angus, P.W. The role of the gut microbiota in NAFLD. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 412–425. [Google Scholar] [CrossRef]
- Chung, E.; Elmassry, M.M.; Kottapalli, P.; Kottapalli, K.R.; Kaur, G.; Dufour, J.M.; Wright, K.; Ramalingam, L.; Moustaid-Moussa, N.; Wang, R.; et al. Metabolic benefits of annatto-extracted tocotrienol on glucose homeostasis, inflammation, and gut microbiome. Nutr. Res. 2020, 77, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Farhana, L.; Sarkar, S.; Nangia-Makker, P.; Yu, Y.; Khosla, P.; Levi, E.; Azmi, A.; Majumdar, A.P.N. Natural agents inhibit colon cancer cell proliferation and alter microbial diversity in mice. PLoS ONE 2020, 15, e0229823. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Park, N.Y.; Jang, Y.; Ma, A.; Jiang, Q. Vitamin E γ-Tocotrienol Inhibits Cytokine-Stimulated NF-κB Activation by Induction of Anti-Inflammatory A20 via Stress Adaptive Response Due to Modulation of Sphingolipids. J. Immunol. 2015, 195, 126–133. [Google Scholar] [CrossRef]
- Yang, C.; Jiang, Q. Vitamin E δ-tocotrienol inhibits TNF-α-stimulated NF-κB activation by up-regulation of anti-inflammatory A20 via modulation of sphingolipid including elevation of intracellular dihydroceramides. J. Nutr. Biochem. 2019, 64, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Atia, A.; Alrawaiq, N.S.; Abdullah, A. Tocotrienols Activate Nrf2 Nuclear Translocation and Increase the Antioxidant- Related Hepatoprotective Mechanism in Mice Liver. Curr. Pharm. Biotechnol. 2021, 22, 1085–1098. [Google Scholar] [CrossRef]
- Chiang, J.Y. Bile acid metabolism and signaling. Compr. Physiol. 2013, 3, 1191–1212. [Google Scholar] [CrossRef]
- Ding, L.; Yang, L.; Wang, Z.; Huang, W. Bile acid nuclear receptor FXR and digestive system diseases. Acta Pharm. Sin. B 2015, 5, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Berlanga, A.; Guiu-Jurado, E.; Porras, J.A.; Auguet, T. Molecular pathways in non-alcoholic fatty liver disease. Clin. Exp. Gastroenterol. 2014, 7, 221–239. [Google Scholar] [CrossRef] [PubMed]
- Bataller, R.; Brenner, D.A. Liver fibrosis. J. Clin. Invest. 2005, 115, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Sufleţel, R.T.; Melincovici, C.S.; Gheban, B.A.; Toader, Z.; Mihu, C.M. Hepatic stellate cells—From past till present: Morphology, human markers, human cell lines, behavior in normal and liver pathology. Rom. J. Morphol. Embryol. 2020, 61, 615–642. [Google Scholar] [CrossRef]
- Hu, H.H.; Chen, D.Q.; Wang, Y.N.; Feng, Y.L.; Cao, G.; Vaziri, N.D.; Zhao, Y.Y. New insights into TGF-β/Smad signaling in tissue fibrosis. Chem. Biol. Interact. 2018, 292, 76–83. [Google Scholar] [CrossRef]
- Mahjabeen, W.; Khan, D.A.; Mirza, S.A.; Pervez, M.A. Effects of delta-tocotrienol supplementation on Glycemic Control, oxidative stress, inflammatory biomarkers and miRNA expression in type 2 diabetes mellitus: A randomized control trial. Phytother. Res. 2021, 35, 3968–3976. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Lim, Y. Tocotrienol-rich fraction supplementation reduces hyperglycemia-induced skeletal muscle damage through regulation of insulin signaling and oxidative stress in type 2 diabetic mice. J. Nutr. Biochem. 2018, 57, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Tao, C.; Sifuentes, A.; Holland, W.L. Regulation of glucose and lipid homeostasis by adiponectin: Effects on hepatocytes, pancreatic β cells and adipocytes. Best Pract. Res. Clin. Endocrinol. Metab. 2014, 28, 43–58. [Google Scholar] [CrossRef] [PubMed]
- Divella, R.; De Luca, R.; Abbate, I.; Naglieri, E.; Daniele, A. Obesity and cancer: The role of adipose tissue and adipo-cytokines-induced chronic inflammation. J. Cancer 2016, 7, 2346–2359. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Dalbeni, A. Treatments for NAFLD: State of Art. Int. J. Mol. Sci. 2021, 22, 2350. [Google Scholar] [CrossRef]
- Serbinova, E.; Kagan, V.; Han, D.; Packer, L. Free radical recycling and intramembrane mobility in the antioxidant properties of alpha-tocopherol and alpha-tocotrienol. Free Radic. Biol. Med. 1991, 10, 263–275. [Google Scholar] [CrossRef]
- Hasan, W.N.W.; Chin, K.Y.; Jolly, J.J.; Ghafar, N.A.; Soelaiman, I.N. Identifying Potential Therapeutics for Osteoporosis by Exploiting the Relationship between Mevalonate Pathway and Bone Metabolism. Endocr. Metab. Immune Disord. Drug Targets 2018, 18, 450–457. [Google Scholar] [CrossRef]
- Schierwagen, R.; Maybüchen, L.; Hittatiya, K.; Klein, S.; Uschner, F.E.; Braga, T.T.; Franklin, B.S.; Nickenig, G.; Strassburg, C.P.; Plat, J.; et al. Statins improve NASH via inhibition of RhoA and Ras. Am. J. Physiol.-Gastrointest. Liver Physiol. 2016, 311, G724–G733. [Google Scholar] [CrossRef]
- US Food and Drug Administration. GRN No. 307: Palm Oil-Derived Tocols with Tocotrienols and α-Tocopherol as the Principal Components. Available online: https://www.cfsanappsexternal.fda.gov/scripts/fdcc/?set=GRASNotices&id=307&sort=GRN_No&order=DESC&startrow=1&type=basic&search=tocotrienol (accessed on 29 October 2022).
- US Food and Drug Administration. GRN No. 471: Annatto Seed Extract. Available online: https://www.cfsanappsexternal.fda.gov/scripts/fdcc/?set=GRASNotices&id=471&sort=GRN_No&order=DESC&startrow=1&type=basic&search=tocotrienol (accessed on 29 October 2022).
- Ima-Nirwana, S.; Nurshazwani, Y.; Nazrun, A.S.; Norliza, M.; Norazlina, M. Subacute and subchronic toxicity studies of palm vitamin E in mice. J. Pharmacol. Toxicol. 2011, 6, 166–173. [Google Scholar] [CrossRef]
- Nakamura, H.; Furukawa, F.; Nishikawa, A.; Miyauchi, M.; Son, H.Y.; Imazawa, T.; Hirose, M. Oral toxicity of a tocotrienol preparation in rats. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2001, 39, 799–805. [Google Scholar] [CrossRef]
- Suleman, F.; Khan, D.A.; Pervez, M.A.; Aamir, M. Effects of delta-tocotrienol supplementation on glycaemic control in individuals with prediabetes: A randomized controlled study. J. Pak. Med. Assoc. 2022, 72, 4–7. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, A.A.; Khan, D.A.; Mahjabeen, W.; Silswal, N.; Qureshi, N. A Novel Mixture of δ-Tocotrienol, Vitamin D3, Resveratrol (NS3) Significantly Decreases Diabetes Biomarkers Including Inflammatory in People with Type 2 Diabetes. J. Diab. Clin. Stud. 2021, 5, 1–16. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chin, K.-Y.; Ekeuku, S.O.; Chew, D.C.H.; Trias, A. Tocotrienol in the Management of Nonalcoholic Fatty Liver Disease: A Systematic Review. Nutrients 2023, 15, 834. https://doi.org/10.3390/nu15040834
Chin K-Y, Ekeuku SO, Chew DCH, Trias A. Tocotrienol in the Management of Nonalcoholic Fatty Liver Disease: A Systematic Review. Nutrients. 2023; 15(4):834. https://doi.org/10.3390/nu15040834
Chicago/Turabian StyleChin, Kok-Yong, Sophia Ogechi Ekeuku, Deborah Chia Hsin Chew, and Anne Trias. 2023. "Tocotrienol in the Management of Nonalcoholic Fatty Liver Disease: A Systematic Review" Nutrients 15, no. 4: 834. https://doi.org/10.3390/nu15040834
APA StyleChin, K.-Y., Ekeuku, S. O., Chew, D. C. H., & Trias, A. (2023). Tocotrienol in the Management of Nonalcoholic Fatty Liver Disease: A Systematic Review. Nutrients, 15(4), 834. https://doi.org/10.3390/nu15040834