
Curcuma aromatica Salisb. Protects from Acetaminophen-Induced Hepatotoxicity by Regulating the Sirt1/HO-1 Signaling Pathway
 , ,
, ,  and
and
Abstract

1. Introduction
2. Materials and Methods
2.1. Primary Mouse Hepatocytes
2.2. Preparation of CAS Extract
2.3. APAP-Induced Liver Injury and CAS Treatment
2.4. Flow Cytometry
2.5. Immunocytochemistry
2.6. In Vivo Model of APAP-Induced Liver Injury
2.7. Serum Analyses
2.8. Histology
2.9. ELISA
2.10. Reverse Transcription Quantitative PCR (RT-qPCR)
2.11. Statistics
3. Results
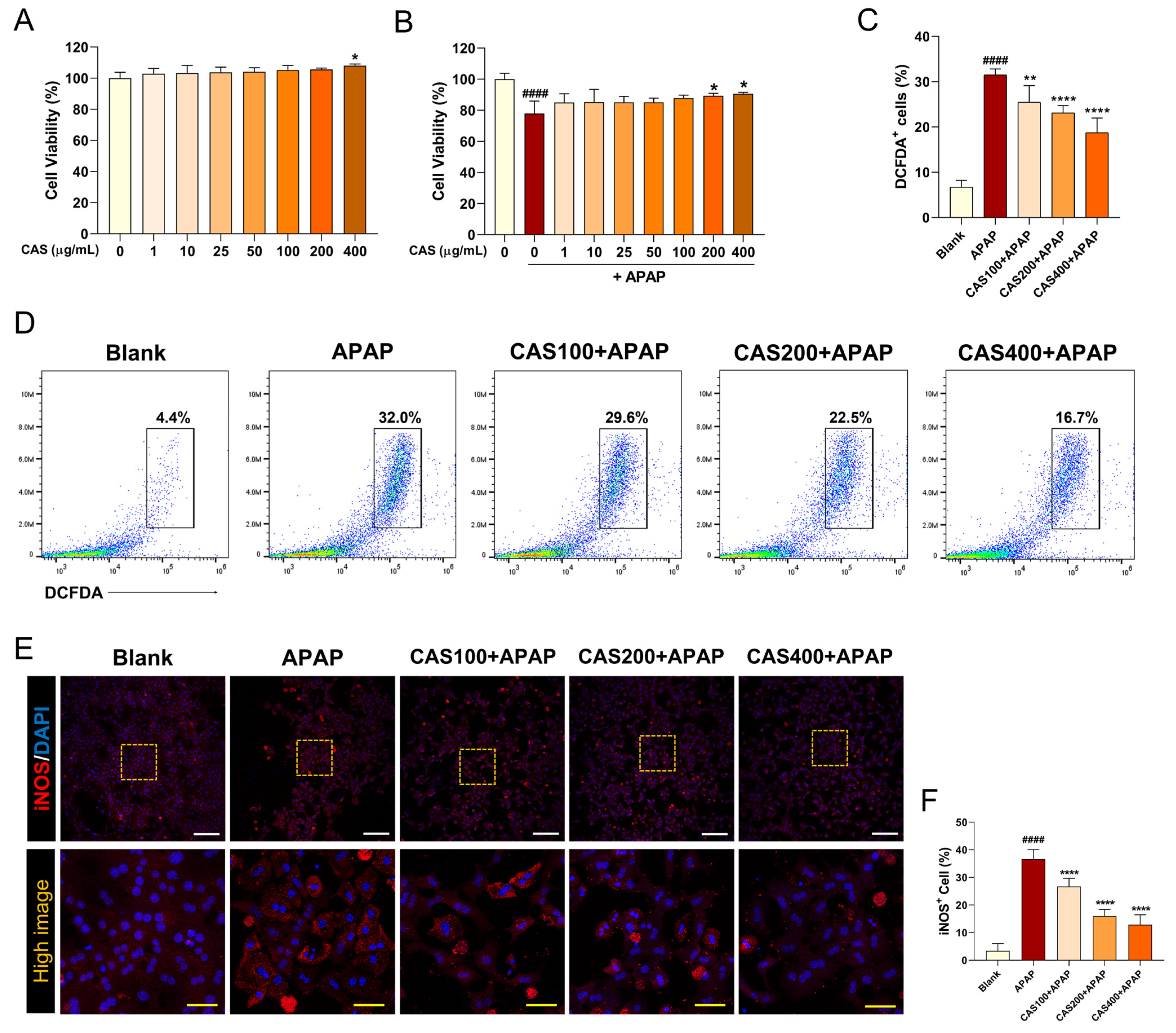
3.1. CAS Suppresses APAP-Induced Cell Viability Reduction and ROS Generation in Primary Mouse Hepatocytes
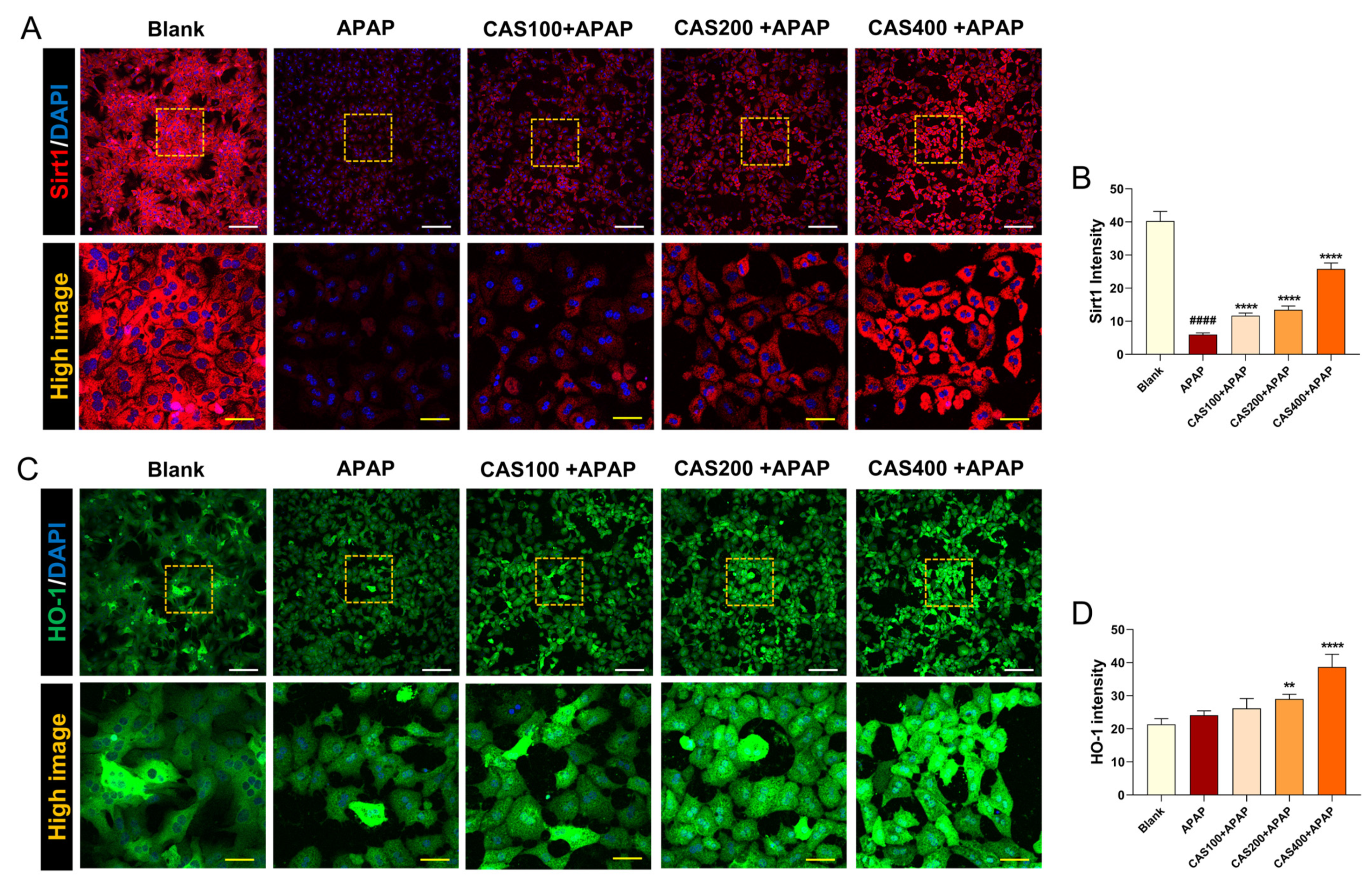
3.2. CAS Protects Cultured Hepatocytes from APAP Hepatotoxicity by Activating the Sirt1/HO-1 Signaling Pathway
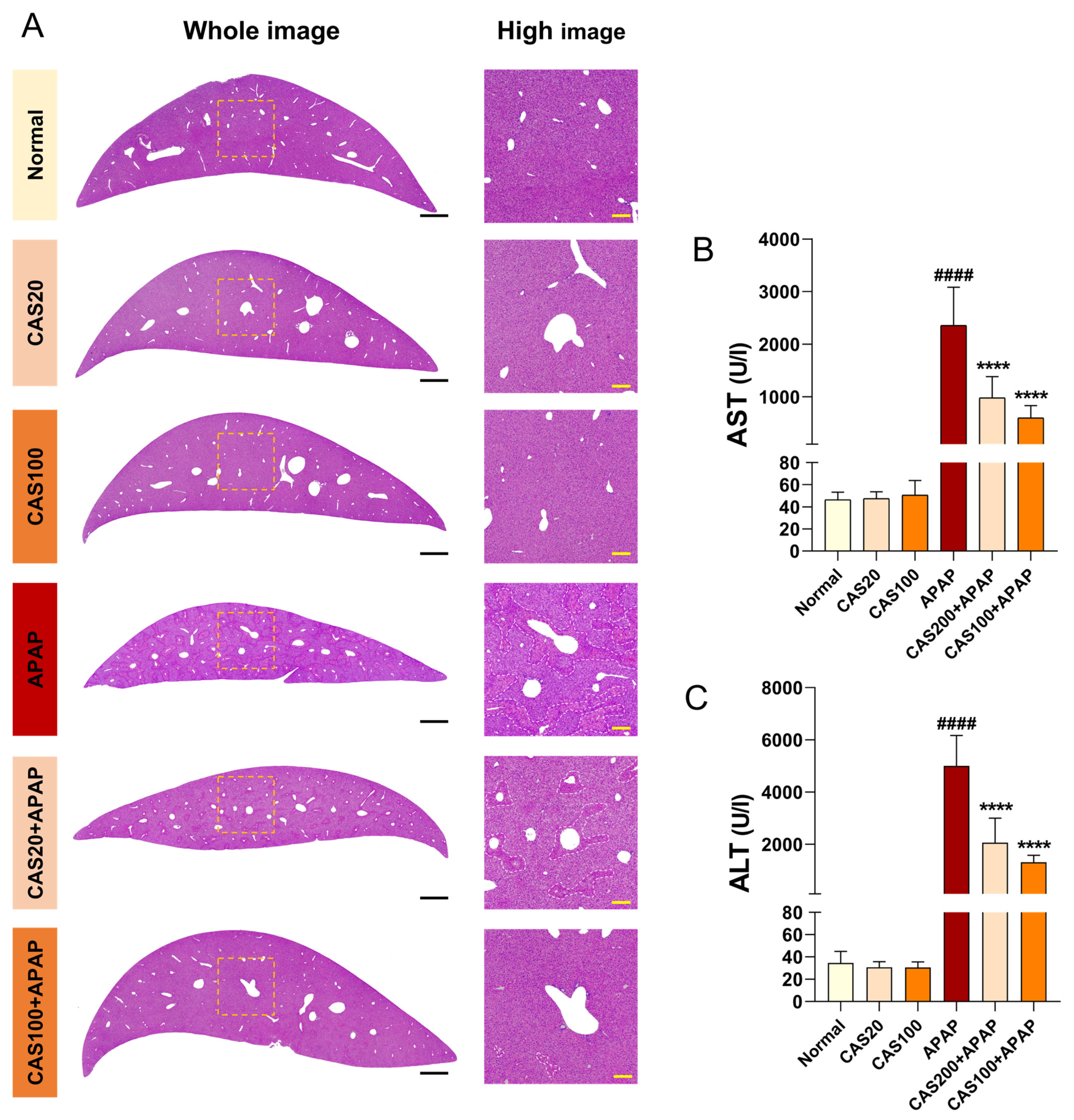
3.3. CAS Protects the Liver from APAP-Induced liver Injury in Mice
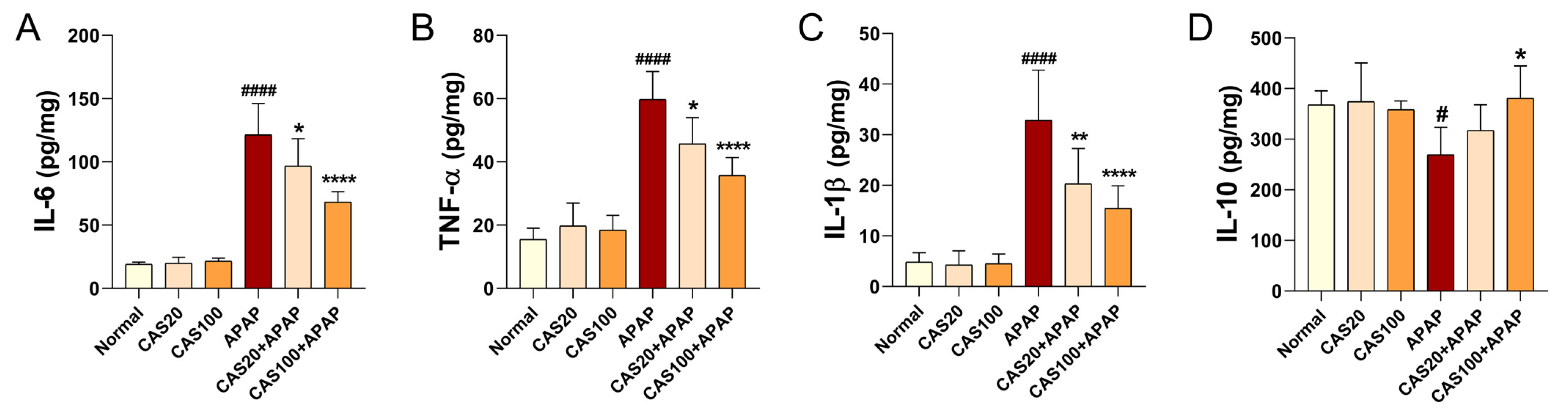
3.4. CAS Ameliorates APAP-Induced Liver Inflammation in Mice
3.5. CAS Alleviates APAP-Induced Oxidative Damage and Glutathione Depletion by Activating Sirt1/HO-1 mRNA Expression in Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jaeschke, H. Acetaminophen: Dose-Dependent Drug Hepatotoxicity and Acute Liver Failure in Patients. Dig. Dis. 2015, 33, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Mossanen, J.C.; Tacke, F. Acetaminophen-induced acute liver injury in mice. Lab. Anim. 2015, 49, 30–36. [Google Scholar] [CrossRef]
- Larson, A.M.; Polson, J.; Fontana, R.J.; Davern, T.J.; Lalani, E.; Hynan, L.S.; Reisch, J.S.; Schiodt, F.V.; Ostapowicz, G.; Shakil, A.O.; et al. Acetaminophen-induced acute liver failure: Results of a United States multicenter, prospective study. Hepatology 2005, 42, 1364–1372. [Google Scholar] [CrossRef]
- Reddy, K.R.; Ellerbe, C.; Schilsky, M.; Stravitz, R.T.; Fontana, R.J.; Durkalski, V.; Lee, W.M.; Acute Liver Failure Study Group. Determinants of outcome among patients with acute liver failure listed for liver transplantation in the United States. Liver Transpl. 2016, 22, 505–515. [Google Scholar] [CrossRef]
- Du, K.; Ramachandran, A.; Jaeschke, H. Oxidative stress during acetaminophen hepatotoxicity: Sources, pathophysiological role and therapeutic potential. Redox Biol. 2016, 10, 148–156. [Google Scholar] [CrossRef]
- Arauz, J.; Ramos-Tovar, E.; Muriel, P. Redox state and methods to evaluate oxidative stress in liver damage: From bench to bedside. Ann. Hepatol. 2016, 15, 160–173. [Google Scholar] [CrossRef]
- Muriel, P.; Gordillo, K.R. Role of Oxidative Stress in Liver Health and Disease. Oxid. Med. Cell Longev. 2016, 2016, 9037051. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Jia, Z.; Pan, M.H.; Anandh Babu, P.V. Natural Products for the Prevention of Oxidative Stress-Related Diseases: Mechanisms and Strategies. Oxid. Med. Cell Longev. 2016, 2016, 4628502. [Google Scholar] [CrossRef] [PubMed]
- Park, M.N.; Rahman, M.A.; Rahman, M.H.; Kim, J.W.; Choi, M.; Kim, J.W.; Choi, J.; Moon, M.; Ahmed, K.R.; Kim, B. Potential Therapeutic Implication of Herbal Medicine in Mitochondria-Mediated Oxidative Stress-Related Liver Diseases. Antioxidants 2022, 11, 2041. [Google Scholar] [CrossRef]
- Tawre, M.S.; Shiledar, A.; Satpute, S.K.; Ahire, K.; Ghosh, S.; Pardesi, K. Synergistic and antibiofilm potential of Curcuma aromatica derived silver nanoparticles in combination with antibiotics against multidrug-resistant pathogens. Front Chem. 2022, 10, 1029056. [Google Scholar] [CrossRef]
- Kulyal, P.; Acharya, S.; Ankari, A.B.; Kokkiripati, P.K.; Tetali, S.D.; Raghavendra, A.S. Variable Secondary Metabolite Profiles Across Cultivars of Curcuma longa L. and C. aromatica Salisb. Front. Pharmacol. 2021, 12, 659546. [Google Scholar] [CrossRef] [PubMed]
- Rajagopal, K.; Varakumar, P.; Baliwada, A.; Byran, G. Activity of phytochemical constituents of Curcuma longa (turmeric) and Andrographis paniculata against coronavirus (COVID-19): An in silico approach. Futur. J. Pharm. Sci. 2020, 6, 104. [Google Scholar] [CrossRef]
- Kocaadam, B.; Sanlier, N. Curcumin, an active component of turmeric (Curcuma longa), and its effects on health. Crit. Rev. Food Sci. Nutr. 2017, 57, 2889–2895. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Rayess, Y.E.; Rizk, A.A.; Sadaka, C.; Zgheib, R.; Zam, W.; Sestito, S.; Rapposelli, S.; Neffe-Skocinska, K.; Zielinska, D.; et al. Turmeric and Its Major Compound Curcumin on Health: Bioactive Effects and Safety Profiles for Food, Pharmaceutical, Biotechnological and Medicinal Applications. Front. Pharmacol. 2020, 11, 01021. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Nie, Y.; Luo, M.; Chen, Z.; He, B. Natural Compounds: A Potential Treatment for Alcoholic Liver Disease? Front. Pharmacol. 2021, 12, 694475. [Google Scholar] [CrossRef]
- Gu, J.; Sun, R.; Tang, D.; Liu, F.; Chang, X.; Wang, Q. Astragalus mongholicus Bunge-Curcuma aromatica Salisb. suppresses growth and metastasis of colorectal cancer cells by inhibiting M2 macrophage polarization via a Sp1/ZFAS1/miR-153-3p/CCR5 regulatory axis. Cell Biol. Toxicol. 2022, 38, 679–697. [Google Scholar] [CrossRef]
- Farghali, H.; Kemelo, M.K.; Canova, N.K. SIRT1 Modulators in Experimentally Induced Liver Injury. Oxid. Med. Cell Longev. 2019, 2019, 8765954. [Google Scholar] [CrossRef]
- Wojnarova, L.; Kutinova Canova, N.; Farghali, H.; Kucera, T. Sirtuin 1 modulation in rat model of acetaminophen-induced hepatotoxicity. Physiol. Res. 2015, 64, S477–S487. [Google Scholar] [CrossRef]
- Chang, H.C.; Guarente, L. SIRT1 and other sirtuins in metabolism. Trends Endocrinol. Metab. 2014, 25, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Yan, T.; Huang, J.; Nisar, M.F.; Wan, C.; Huang, W. The Beneficial Roles of SIRT1 in Drug-Induced Liver Injury. Oxid. Med. Cell Longev. 2019, 2019, 8506195. [Google Scholar] [CrossRef]
- Rada, P.; Pardo, V.; Mobasher, M.A.; Garcia-Martinez, I.; Ruiz, L.; Gonzalez-Rodriguez, A.; Sanchez-Ramos, C.; Muntane, J.; Alemany, S.; James, L.P.; et al. SIRT1 Controls Acetaminophen Hepatotoxicity by Modulating Inflammation and Oxidative Stress. Antioxid. Redox Signal. 2018, 28, 1187–1208. [Google Scholar] [CrossRef]
- Cui, X.; Chen, Q.; Dong, Z.; Xu, L.; Lu, T.; Li, D.; Zhang, J.; Zhang, M.; Xia, Q. Inactivation of Sirt1 in mouse livers protects against endotoxemic liver injury by acetylating and activating NF-kappaB. Cell Death Dis. 2016, 7, e2403. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, J.; Hu, C.; Wang, T.; Lu, J.; Wu, C.; Chen, L.; Jin, M.; Ji, G.; Cao, Q.; et al. Apigenin Prevents Acetaminophen-Induced Liver Injury by Activating the SIRT1 Pathway. Front. Pharmacol. 2020, 11, 514. [Google Scholar] [CrossRef] [PubMed]
- Singh, C.K.; Chhabra, G.; Ndiaye, M.A.; Garcia-Peterson, L.M.; Mack, N.J.; Ahmad, N. The Role of Sirtuins in Antioxidant and Redox Signaling. Antioxid. Redox Signal. 2018, 28, 643–661. [Google Scholar] [CrossRef] [PubMed]
- McGill, M.R.; Hinson, J.A. The development and hepatotoxicity of acetaminophen: Reviewing over a century of progress. Drug Metab. Rev. 2020, 52, 472–500. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.M. Drug-induced acute liver failure. Clin. Liver Dis. 2013, 17, 575–586. [Google Scholar] [CrossRef]
- Teschke, R.; Eickhoff, A. Herbal hepatotoxicity in traditional and modern medicine: Actual key issues and new encouraging steps. Front. Pharmacol. 2015, 6, 72. [Google Scholar] [CrossRef]
- Gu, J.; Sun, R.; Wang, Q.; Liu, F.; Tang, D.; Chang, X. Standardized Astragalus Mongholicus Bunge-Curcuma Aromatica Salisb. Extract Efficiently Suppresses Colon Cancer Progression through Gut Microbiota Modification in CT26-Bearing Mice. Front. Pharmacol. 2021, 12, 714322. [Google Scholar] [CrossRef]
- Lee, H.Y.; Kim, S.W.; Lee, G.H.; Choi, M.K.; Jung, H.W.; Kim, Y.J.; Kwon, H.J.; Chae, H.J. Turmeric extract and its active compound, curcumin, protect against chronic CCl4-induced liver damage by enhancing antioxidation. BMC Complement. Altern. Med. 2016, 16, 316. [Google Scholar] [CrossRef]
- Quiles, J.L.; Mesa, M.D.; Ramirez-Tortosa, C.L.; Aguilera, C.M.; Battino, M.; Gil, A.; Ramirez-Tortosa, M.C. Curcuma longa extract supplementation reduces oxidative stress and attenuates aortic fatty streak development in rabbits. Arter. Thromb. Vasc. Biol. 2002, 22, 1225–1231. [Google Scholar] [CrossRef] [PubMed]
- Kheradpezhouh, E.; Panjehshahin, M.R.; Miri, R.; Javidnia, K.; Noorafshan, A.; Monabati, A.; Dehpour, A.R. Curcumin protects rats against acetaminophen-induced hepatorenal damages and shows synergistic activity with N-acetyl cysteine. Eur. J. Pharmacol. 2010, 628, 274–281. [Google Scholar] [CrossRef] [PubMed]
- McGill, M.R.; Sharpe, M.R.; Williams, C.D.; Taha, M.; Curry, S.C.; Jaeschke, H. The mechanism underlying acetaminophen-induced hepatotoxicity in humans and mice involves mitochondrial damage and nuclear DNA fragmentation. J. Clin. Investig. 2012, 122, 1574–1583. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Qian, K.; Xiong, J.; Ma, K.; Wang, A.; Zou, Y. Curcumin alleviates lipopolysaccharide induced sepsis and liver failure by suppression of oxidative stress-related inflammation via PI3K/AKT and NF-kappaB related signaling. Biomed. Pharmacother. 2016, 83, 302–313. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri-Fard, S.; Shoorei, H.; Bahroudi, Z.; Hussen, B.M.; Talebi, S.F.; Taheri, M.; Ayatollahi, S.A. Nrf2-Related Therapeutic Effects of Curcumin in Different Disorders. Biomolecules 2022, 12, 82. [Google Scholar] [CrossRef]
- Ghareghomi, S.; Rahban, M.; Moosavi-Movahedi, Z.; Habibi-Rezaei, M.; Saso, L.; Moosavi-Movahedi, A.A. The Potential Role of Curcumin in Modulating the Master Antioxidant Pathway in Diabetic Hypoxia-Induced Complications. Molecules 2021, 26, 7658. [Google Scholar] [CrossRef]
- Ungurianu, A.; Zanfirescu, A.; Margina, D. Regulation of Gene Expression through Food-Curcumin as a Sirtuin Activity Modulator. Plants 2022, 11, 1741. [Google Scholar] [CrossRef]
- Xiao, Y.; Xia, J.; Wu, S.; Lv, Z.; Huang, S.; Huang, H.; Su, X.; Cheng, J.; Ke, Y. Curcumin Inhibits Acute Vascular Inflammation through the Activation of Heme Oxygenase-1. Oxid. Med. Cell Longev. 2018, 2018, 3295807. [Google Scholar] [CrossRef]
- Farzaei, M.H.; Zobeiri, M.; Parvizi, F.; El-Senduny, F.F.; Marmouzi, I.; Coy-Barrera, E.; Naseri, R.; Nabavi, S.M.; Rahimi, R.; Abdollahi, M. Curcumin in Liver Diseases: A Systematic Review of the Cellular Mechanisms of Oxidative Stress and Clinical Perspective. Nutrients 2018, 10, 855. [Google Scholar] [CrossRef]
- Kemelo, M.K.; Kutinova Canova, N.; Horinek, A.; Farghali, H. Sirtuin-activating compounds (STACs) alleviate D-galactosamine/lipopolysaccharide-induced hepatotoxicity in rats: Involvement of sirtuin 1 and heme oxygenase 1. Physiol. Res. 2017, 66, 497–505. [Google Scholar] [CrossRef]
- Kemelo, M.K.; Pierzynova, A.; Kutinova Canova, N.; Kucera, T.; Farghali, H. The involvement of sirtuin 1 and heme oxygenase 1 in the hepatoprotective effects of quercetin against carbon tetrachloride-induced sub-chronic liver toxicity in rats. Chem. Biol. Interact. 2017, 269, 1–8. [Google Scholar] [CrossRef]
- Hall, P.; Cash, J. What is the real function of the liver ‘function’ tests? Ulster Med. J. 2012, 81, 30–36. [Google Scholar]
- Hinson, J.A.; Pohl, L.R.; Monks, T.J.; Gillette, J.R. Acetaminophen-induced hepatotoxicity. Life Sci. 1981, 29, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Vairetti, M.; Di Pasqua, L.G.; Cagna, M.; Richelmi, P.; Ferrigno, A.; Berardo, C. Changes in Glutathione Content in Liver Diseases: An Update. Antioxidants 2021, 10, 364. [Google Scholar] [CrossRef] [PubMed]
- Iwakiri, Y. Nitric oxide in liver fibrosis: The role of inducible nitric oxide synthase. Clin. Mol. Hepatol. 2015, 21, 319–325. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | 5′-3′ | Primer Sequence |
|---|---|---|
| IL-6 | Forward | CCACCCACAACAGACCAGTA |
| Reverse | GGAACTCCAGAAGACCAGAGC | |
| TNF-α | Forward | CCGACTACGTGCTCCTCACC |
| Reverse | CTCCAAAGTAGACCTGCCCG | |
| IL-1β | Forward | TTGCTTCCAAGCCCTTGACT |
| Reverse | GGTCGTCATCATCCCACGAG | |
| IL-10 | Forward | TAACTGCACCCACTTCCCAG |
| Reverse | AGGCTTGGCAACCCAAGTAA | |
| iNOS | Forward | ATGGCTTGCCCCTGGAAGTT |
| Reverse | TGTTGGGCTGGGAATAGCAC | |
| COX2 | Forward | CTCAGCCATGCAGCAAATCC |
| Reverse | GGGTGGGCTTCAGCAGTAAT | |
| Sirt1 | Forward | AGGGAACCTCTGCCTCATCT |
| Reverse | TGGCATACTCGCCACCTAAC | |
| HO-1 | Forward | CCCACCAAGTTCAAACAGCTC |
| Reverse | AGGAAGGCGGTCTTAGCCTC | |
| GAPDH | Forward | CCCCCAATGTATCCGTTGTG |
| Reverse | TAGCCCAGGATGCCCTTTAGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.; Hong, J.; Lee, J.; Jeon, W.; Yeo, C.; Lee, Y.; Baek, S.; Ha, I. Curcuma aromatica Salisb. Protects from Acetaminophen-Induced Hepatotoxicity by Regulating the Sirt1/HO-1 Signaling Pathway. Nutrients 2023, 15, 808. https://doi.org/10.3390/nu15040808
Kim H, Hong J, Lee J, Jeon W, Yeo C, Lee Y, Baek S, Ha I. Curcuma aromatica Salisb. Protects from Acetaminophen-Induced Hepatotoxicity by Regulating the Sirt1/HO-1 Signaling Pathway. Nutrients. 2023; 15(4):808. https://doi.org/10.3390/nu15040808
Chicago/Turabian StyleKim, Hyunseong, Jinyoung Hong, Junseon Lee, Wanjin Jeon, Changhwan Yeo, Yoonjae Lee, Seungho Baek, and Inhyuk Ha. 2023. "Curcuma aromatica Salisb. Protects from Acetaminophen-Induced Hepatotoxicity by Regulating the Sirt1/HO-1 Signaling Pathway" Nutrients 15, no. 4: 808. https://doi.org/10.3390/nu15040808
APA StyleKim, H., Hong, J., Lee, J., Jeon, W., Yeo, C., Lee, Y., Baek, S., & Ha, I. (2023). Curcuma aromatica Salisb. Protects from Acetaminophen-Induced Hepatotoxicity by Regulating the Sirt1/HO-1 Signaling Pathway. Nutrients, 15(4), 808. https://doi.org/10.3390/nu15040808