

Saponin and Fatty Acid Profiling of the Sea Cucumber Holothuria atra, α-Glucosidase Inhibitory Activity and the Identification of a Novel Triterpene Glycoside

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
General Experimental Procedures
2.2. Methods
2.2.1. Sea Cucumber Material
2.2.2. Preparation of Sea Cucumber H. atra Extract
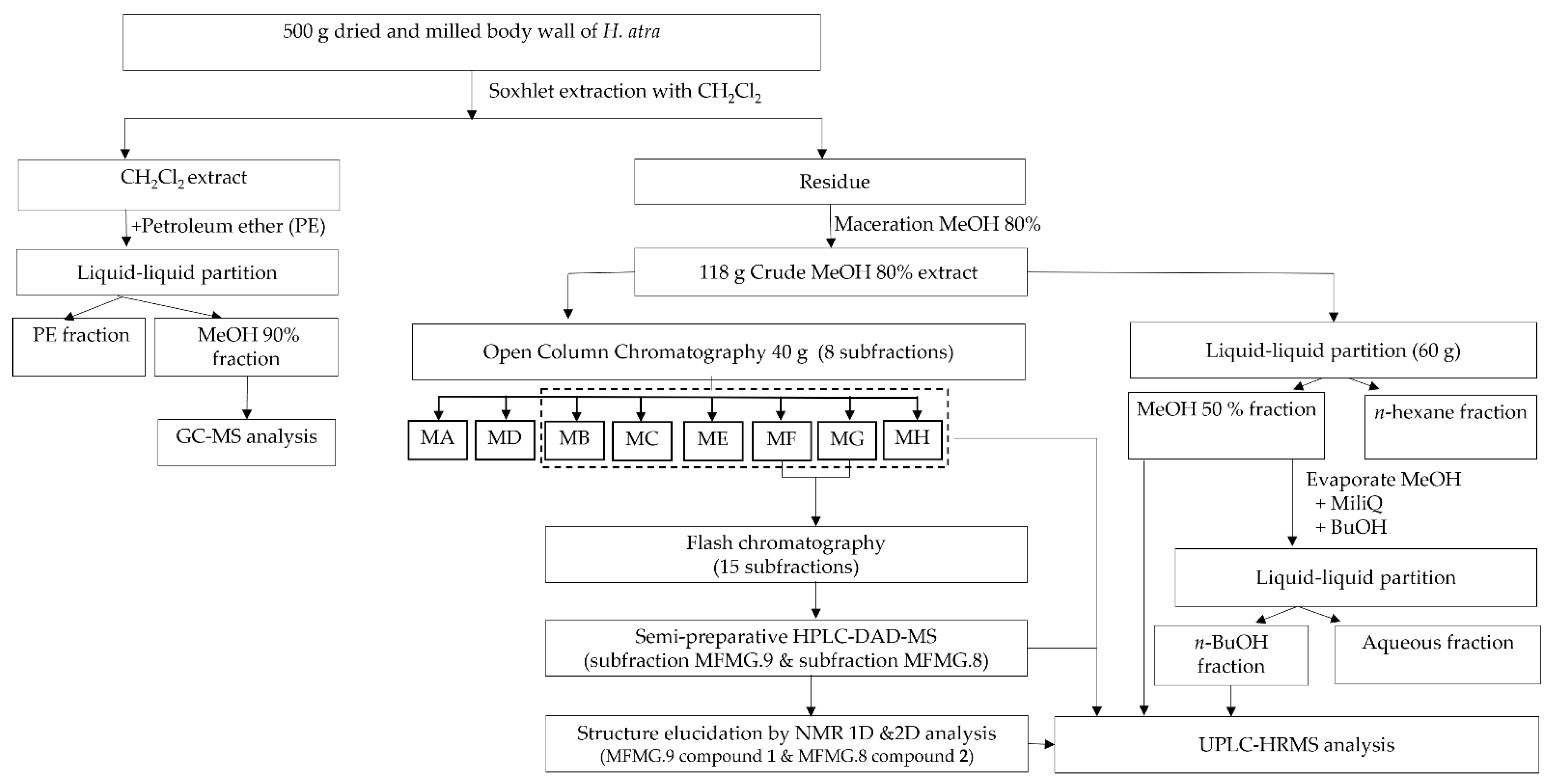
2.2.3. Fractionation of Triterpene Glycosides
2.2.4. Isolation of Triterpene Glycosides
2.2.5. Structure Elucidation Using 1D and 2D NMR
2.2.6. UPLC-HRMS Analysis
2.2.7. GC-MS Analysis
2.2.8. α-Glucosidase Inhibition Assay
3. Results
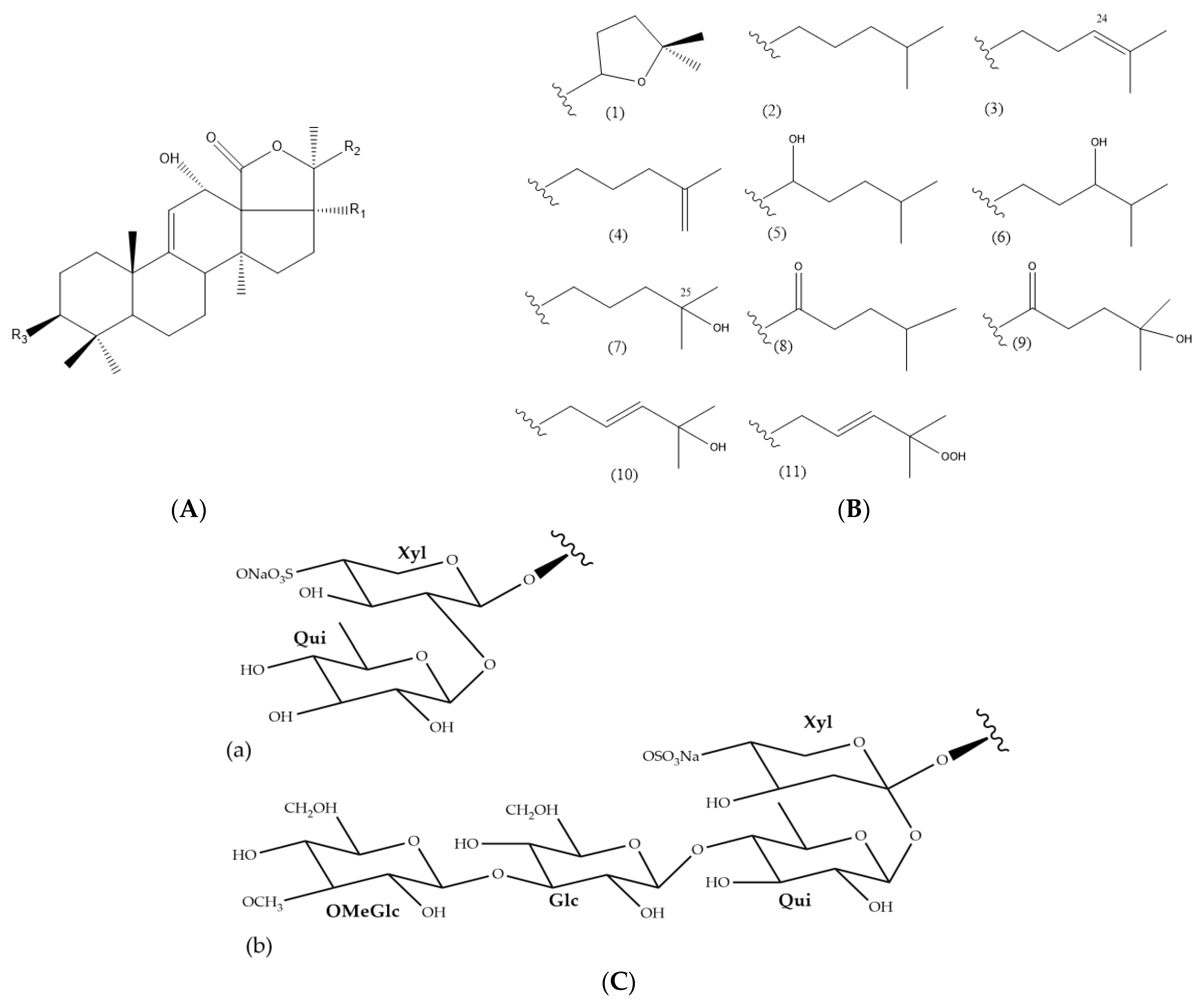
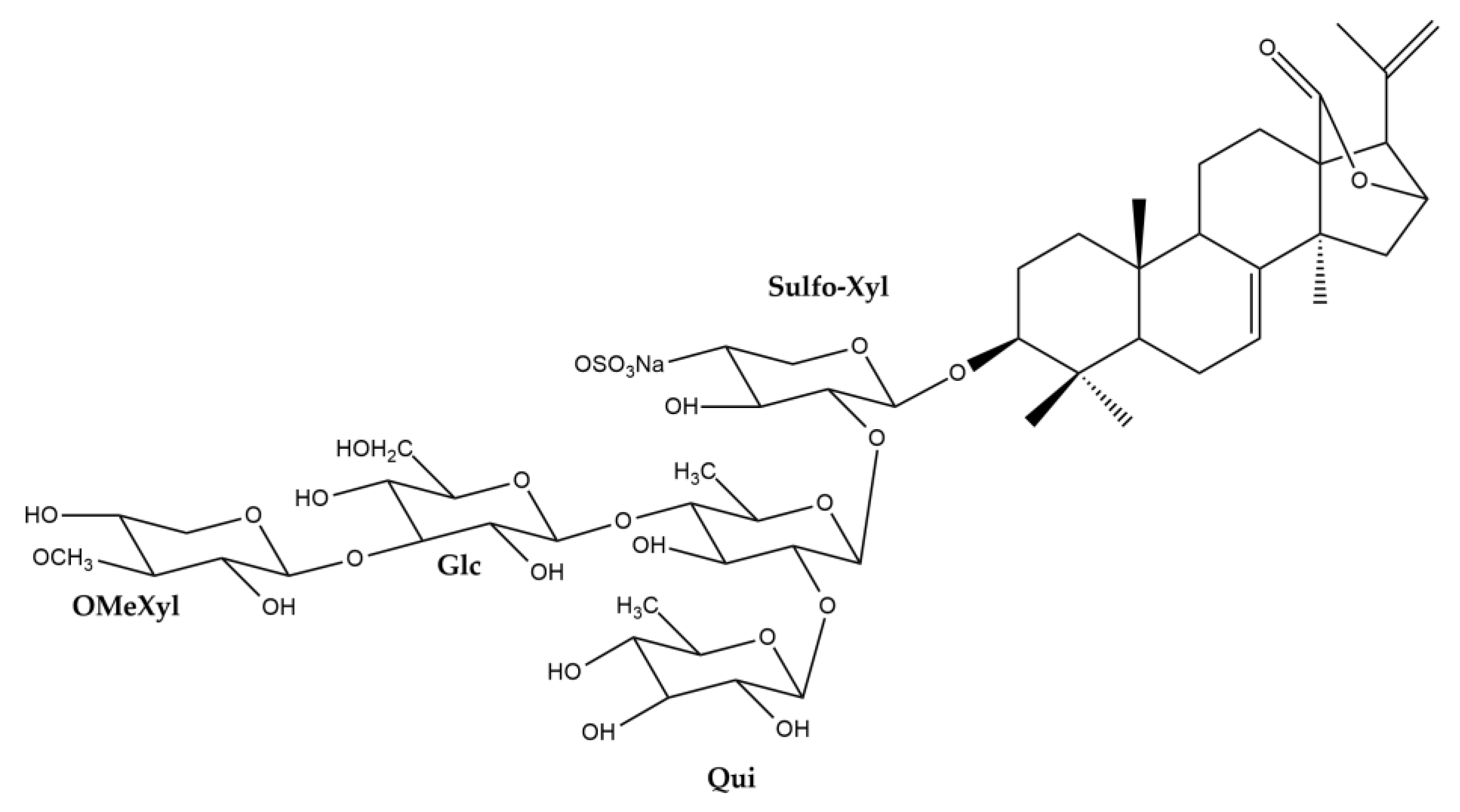
3.1. Tentative Identification of Triterpene Glycosides from H. atra Body Wall
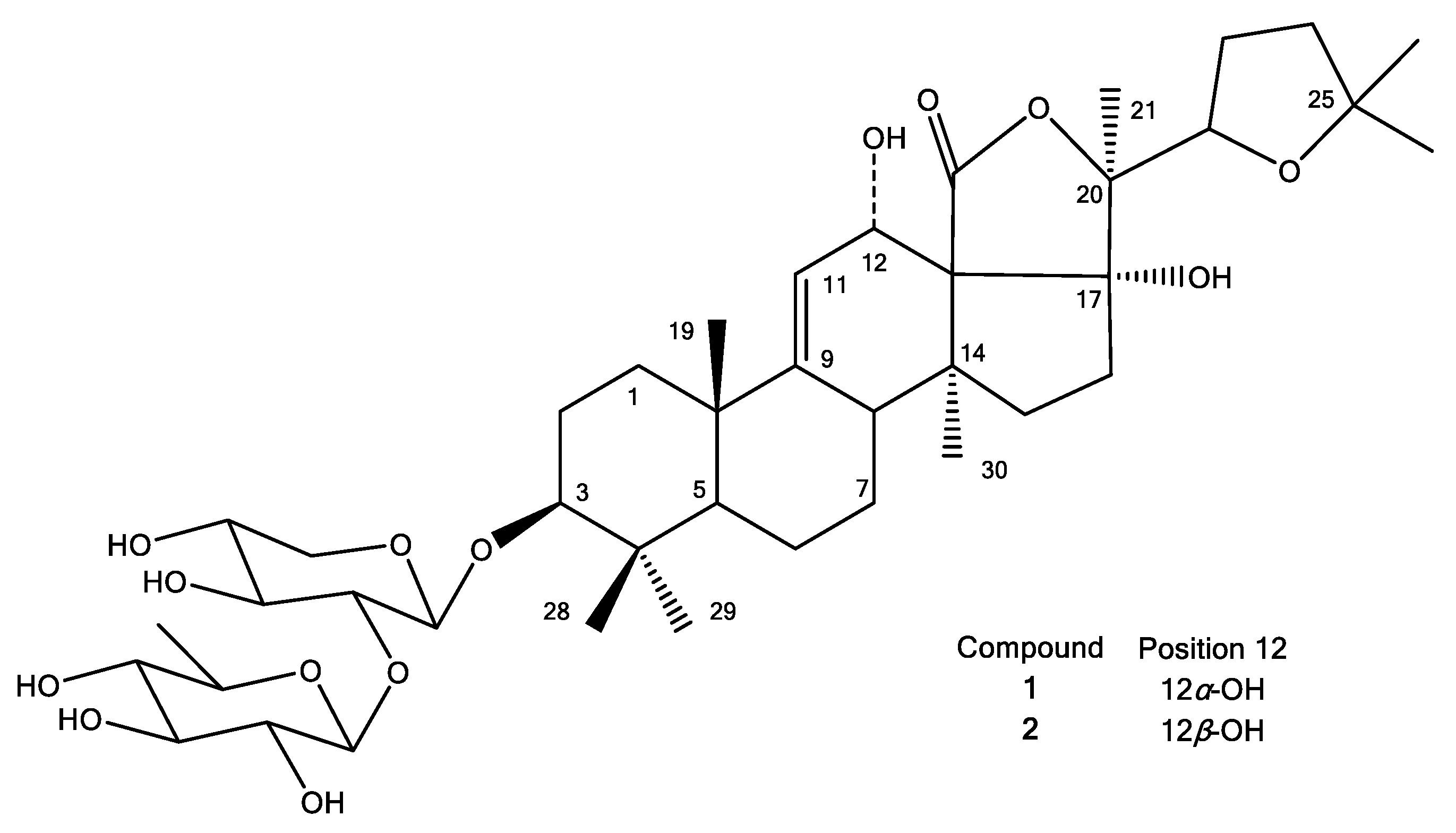
3.2. Purified Saponins from H. atra Body Wall
3.3. Fatty Acid Profile of the H. atra Body Wall
3.4. α-Glucosidase Inhibition Assay
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kinch, J.; Purcell, S.; Uthicke, S.; Friedman, K. Population Status, Fisheries and Trade of Sea Cucumbers in the Western Central Pacific. In Sea Cucumbers. A Global Review of Fisheries and Trade; Toral-Granda, V., Lovatelli, A., Vasconce, M., Eds.; FAO Fisheries and Aquaculture Technical Paper No. 516; FAO: Rome, Italy, 2008; pp. 7–55. [Google Scholar]
- Fabinyi, M. Historical, Cultural and Social Perspectives on Luxury Seafood Consumption in China. Environ. Conserv. 2012, 39, 83–92. [Google Scholar] [CrossRef]
- Yuan, W.H.; Yi, Y.H.; Xue, M.; Zhang, H.W.; La, M.P. Two Antifungal Active Triterpene Glycosides from Sea Cucumber Holothuria (Microthele) Axiloga. Chin. J. Nat. Med. 2008, 6, 105–108. [Google Scholar] [CrossRef]
- Yuan, W.H.; Yi, Y.H.; Tan, R.X.; Wang, Z.L.; Sun, G.Q.; Xue, M.; Zhang, H.W.; Tang, H.F. Antifungal Triterpene Glycosides from the Sea Cucumber Holothuria (Microthele) Axiloga. Planta Med. 2009, 75, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, J.; Castro, R.; Riguera, R. Holothurinosides: New Antitumor Non Sulphated Triterpenoid Glycosides from the Sea Cucumber Holothuria forskalii. Tetrahedron 1991, 47, 4753–4762. [Google Scholar] [CrossRef]
- Murniasih, T.; Putra, M.; Pangestuti, R. Antioxidant Capacities of Holothuria Sea Cucumbers. Ann. Bogor. 2015, 19, 21–26. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Yi, Y.H.; Tang, H.F. Bioactive Triterpene Glycosides from the Sea Cucumber Holothuria fuscocinerea. J. Nat. Prod. 2006, 69, 1492–1495. [Google Scholar] [CrossRef]
- Sun, G.Q.; Li, L.; Yi, Y.H.; Yuan, W.H.; Liu, B.S.; Weng, Y.Y.; Zhang, S.L.; Sun, P.; Wang, Z.L. Two New Cytotoxic Nonsulfated Pentasaccharide Holostane (=20-Hydroxylanostan-18-Oic Acid γ-Lactone) Glycosides from the Sea Cucumber Holothuria grisea. Helv. Chim. Acta 2008, 91, 1453–1460. [Google Scholar] [CrossRef]
- Wu, J.; Yi, Y.H.; Tang, H.F.; Wu, H.M.; Zhou, Z.R. Hillasides A and B, Two New Cytotoxic Triterpene Glycosides from the Sea Cucumber Holothuria hilla Lesson. J. Asian Nat. Prod. Res. 2007, 9, 609–615. [Google Scholar] [CrossRef]
- Wang, J.; Han, H.; Chen, X.; Yi, Y.; Sun, H. Cytotoxic and Apoptosis-Inducing Activity of Triterpene Glycosides from Holothuria scabra and Cucumaria frondosa against HepG2 Cells. Mar. Drugs 2014, 12, 4274–4290. [Google Scholar] [CrossRef]
- Kobayashi, M.; Hori, M.; Kan, K.; Yasuzawa, T.; Matsui, M.; Suzuki, S.; Kitagawa, I. Marine Natural Products. XXVII. Distribution of Lanostane-Type Triterpene Oligoglycoside in Ten Kinds of Okinawan Sea Cucumbers. Chem. Pharm. Bull. 1991, 39, 2282–2287. [Google Scholar] [CrossRef]
- Van Dyck, S.; Gerbaux, P.; Flammang, P. Qualitative and Quantitative Saponin Contents in Five Sea Cucumbers from the Indian Ocean. Mar. Drugs 2010, 8, 173–189. [Google Scholar] [CrossRef]
- Delia, J.T.; Herthel, L.W. Characterization of the Aglycones of the Toxic Principle of the Sea Cucumber Holothuria atra. Toxicon 1977, 15, 315–316. [Google Scholar] [CrossRef]
- Vincken, J.P.; Heng, L.; de Groot, A.; Gruppen, H. Saponins, Classification and Occurrence in the Plant Kingdom. Phytochemistry 2007, 68, 275–297. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.H.; Zou, Z.R.; Yi, Y.H.; Han, H.; Li, N.; Pan, M.X. Variegatusides: New Non-Sulphated Triterpene Glycosides from the Sea Cucumber Stichopus Variegates Semper. Mar. Drugs 2014, 12, 2004. [Google Scholar] [CrossRef]
- Van Dyck, S.; Caulier, G.; Todesco, M.; Gerbaux, P.; Fournier, I.; Wisztorski, M.; Flammang, P. The Triterpene Glycosides of Holothuria forskali: Usefulness and Efficiency as a Chemical Defense Mechanism against Predatory Fish. J. Exp. Biol. 2011, 214, 1347–1356. [Google Scholar] [CrossRef] [PubMed]
- Garneau, F.-X.; Simard, J.; Harvey, O.; Apsimon, J.W.; Girard, M. The Structure of Psoluthurin A, the Major Triterpene Glycoside of the Sea Cucumber Psolus fabricii. Can. J. Chem. 1983, 61, 1465–1471. [Google Scholar] [CrossRef]
- Kicha, A.A.; Ivanchina, N.V.; Kalinovsky, A.I.; Dmitrenok, P.S.; Stonik, V.A. Sulfated Steroid Compounds from the Starfish Aphelasterias japonica of the Kuril Population. Russ. Chem. Bull. 2001, 50, 724–727. [Google Scholar] [CrossRef]
- Kitagawa, I.; Kobayashi, M. On the Structure of the Major Saponin from the Starfish Acanthaster planci. Tetrahedron Lett. 1977, 18, 859–862. [Google Scholar] [CrossRef]
- Kitagawa, I.; Kobayashi, M. Steroidal Saponins from the Starfish Acanthaster planci L. (Crown of Thorns). Chem. Pharm. Bull. 1978, 26, 1864–1873. [Google Scholar] [CrossRef]
- Demeyer, M.; De Winter, J.; Caulier, G.; Eeckhaut, I.; Flammang, P.; Gerbaux, P. Molecular Diversity and Body Distribution of Saponins in the Sea Star Asterias rubens by Mass Spectrometry. Comp. Biochem. Physiol.-B Biochem. Mol. Biol. 2014, 168, 1–11. [Google Scholar] [CrossRef]
- Calabro, K.; Kalahroodi, E.L.; Rodrigues, D.; Díaz, C.; De La Cruz, M.; Cautain, B.; Laville, R.; Reyes, F.; Pérez, T.; Soussi, B.; et al. Poecillastrosides, Steroidal Saponins from the Mediterranean Deep-Sea Sponge Poecillastra compressa (Bowerbank, 1866). Mar. Drugs 2017, 15, 199. [Google Scholar] [CrossRef]
- Colorado, J.; Muñoz, D.; Marquez, D.; Marquez, M.E.; Lopez, J.; Thomas, O.P.; Martinez, A. Ulososides and Urabosides—Triterpenoid Saponins from the Caribbean Marine Sponge Ectyoplasia ferox. Molecules 2013, 18, 2598–2610. [Google Scholar] [CrossRef]
- Kubanek, J.; Pawlik, J.R.; Eve, T.M.; Fenical, W. Triterpene Glycosides Defend the Caribbean Reef Sponge Erylus Formosus from Predatory Fishes. Mar. Ecol. Prog. Ser. 2000, 207, 69–77. [Google Scholar] [CrossRef]
- Oleszek, W.; Bialy, Z. Chromatographic Determination of Plant Saponins—An Update (2002–2005). J. Chromatogr. A 2006, 1112, 78–91. [Google Scholar] [CrossRef]
- Kalinin, V.I.; Silchenko, A.S.; Avilov, S.A.; Stonik, V.A.; Smirnov, A.V. Sea Cucumbers Triterpene Glycosides, the Recent Progress in Structural Elucidation and Chemotaxonomy. Phytochem. Rev. 2005, 4, 221–236. [Google Scholar] [CrossRef]
- Silchenko, A.S.; Kalinovsky, A.I.; Avilov, S.A.; Andryjaschenko, P.V.; Dmitrenok, P.S.; Martyyas, E.A.; Kalinin, V.I. Triterpene Glycosides from the Sea Cucumber Eupentacta fraudatrix. Structure and Biological Action of Cucumariosides I1, I3, I4, Three New Minor Disulfated Pentaosides. Nat. Prod. Commun. 2013, 8, 1053–1058. [Google Scholar] [CrossRef] [PubMed]
- Antonov, A.S.; Avilov, S.A.; Kalinovsky, A.I.; Anastyuk, S.D.; Dmitrenok, P.S.; Evtushenko, E.V.; Kalinin, V.I.; Smirnov, A.V.; Taboada, S.; Ballesteros, M.; et al. Triterpene Glycosides from Antarctic Sea Cucumbers. 1. Structure of Liouvillosides A1, A2, A3, B1, and B2 from the Sea Cucumber Staurocucumis liouvillei: New Procedure for Separation of Highly Polar Glycoside Fractions and Taxonomic Revision. J. Nat. Prod. 2008, 71, 1677–1685. [Google Scholar] [CrossRef]
- Antonov, A.S.; Avilov, S.A.; Kalinovsky, A.I.; Dmitrenok, P.S.; Kalinin, V.I.; Taboada, S.; Ballesteros, M.; Avila, C. Triterpene Glycosides from Antarctic Sea Cucumbers III. Structures of Liouvillosides A4 and A5, Two Minor Disulphated Tetraosides Containing 3-O-Methylquinovose as Terminal Monosaccharide Units from the Sea Cucumber Staurocucumis liouvillei (Vaney). Nat. Prod. Res. 2011, 25, 1324–1333. [Google Scholar] [CrossRef]
- Bahrami, Y.; Franco, C.M.M. Structure Elucidation of New Acetylated Saponins, Lessoniosides A, B, C, D, and E, and Non-Acetylated Saponins, Lessoniosides F and G, from the Viscera of the Sea Cucumber Holothuria lessoni. Mar. Drugs 2015, 13, 597–617. [Google Scholar] [CrossRef] [PubMed]
- Stonik, V.A.; Kalinin, V.I.; Avilov, S.A. Toxins from Sea Cucumber (Holothuroids): Chemical Structures, Properties, Taxonomic Distribution, Biosynthesis and Evolution. J. Nat. Toxins 1999, 8, 235–247. [Google Scholar]
- Puspitasari, Y.E.; De Bruyne, T.; Foubert, K.; Aulanni’am, A.; Pieters, L.; Hermans, N.; Tuenter, E. Holothuria Triterpene Glycosides: A Comprehensive Guide for Their Structure Elucidation and Critical Appraisal of Reported Compounds. Phytochem. Rev. 2021, 21, 1315–1358, Erratum in Phytochem. Rev. 2022, 21, 1359–1378. [Google Scholar] [CrossRef]
- Bonham, K.; Held, E.E. Ecological Observations on the Sea Cucumbers Holothuria atra and H. leucospilota at Rongelap Atoll, Marshall Islands. Pac. Sci. 1963, 17, 305–314. [Google Scholar]
- Hartati, R.; Widianingsih; Trianto, A.; Zainuri, M.; Ambariyanto, A. The Abundance of Prospective Natural Food for Sea Cucumber Holothuria atra at Karimunjawa Island Waters, Jepara, Indonesia. Biodiversitas 2017, 18, 947–953. [Google Scholar] [CrossRef]
- Stonik, V.A.; Chumak, A.D.; Isakov, V.V.; Belogortseva, N.I.; Chirva, V.Y.; Elyakov, G.B. Glycosides of Marine Invertebrates. VII. Structure of Holothurin B from Holothuria atra. Chem. Nat. Compd. 1979, 15, 453–457. [Google Scholar] [CrossRef]
- Omran, N.E.; Salem, H.K.; Eissa, S.H.; Kabbash, A.M.; Kandeil, M.A.; Salem, M.A. Chemotaxonomic Study of the Most Abundant Egyptian Sea-Cucumbers Using Ultra-Performance Liquid Chromatography (UPLC) Coupled to High-Resolution Mass Spectrometry (HRMS). Chemoecology 2020, 30, 35–48. [Google Scholar] [CrossRef]
- Grauso, L.; Yegdaneh, A.; Sharifi, M.; Mangoni, A.; Zolfaghari, B.; Lanzotti, V. Molecular Networking-Based Analysis of Cytotoxic Saponins from Sea Cucumber Holothuria atra. Mar. Drugs 2019, 17, 86. [Google Scholar] [CrossRef]
- Shahinozzaman, M.; Ishii, T.; Takano, R.; Halim, M.A.; Hossain, M.A.; Tawata, S. Cytotoxic Desulfated Saponin from Holothuria atra Predicted to Have High Binding Affinity to the Oncogenic Kinase PAK1: A Combined in Vitro and in Silico Study. Sci. Pharm. 2018, 86, 32. [Google Scholar] [CrossRef]
- Kiew, P.L.; Don, M.M. Jewel of the Seabed: Sea Cucumbers as Nutritional and Drug Candidates. Int. J. Food Sci. Nutr. 2012, 63, 616–636. [Google Scholar] [CrossRef]
- Hashim, R. Sea Cucumbers A Malaysian Heritage; International Islamic University Malaysia: Pahang, Malaysia, 2007. [Google Scholar]
- Fu, X.; Wen, M.; Han, X.; Yanagita, T.; Xue, Y.; Wang, J.; Xue, C.; Wang, Y. Effect and Potential Mechanism of Action of Sea Cucumber Saponins on Postprandial Blood Glucose in Mice. Biosci. Biotechnol. Biochem. 2016, 80, 1081–1087. [Google Scholar] [CrossRef]
- El Barky, A.R.; Hussein, S.A.; Alm-Eldeen, A.A.; Hafez, Y.A.; Mohamed, T.M.; El Barky, A.R.; Hussein, S.A.; Alm-Eldeen, A.A.; Hafez, Y.A.; Mohamed, T.M. Anti-Diabetic Activity of Holothuria thomasi Saponin. Biomed. Pharmacother. 2016, 84, 1472–1487. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Um, B.H.; Kim, S.M. Two Unsaturated Fatty Acids with Potent α-Glucosidase Inhibitory Activity Purified from the Body Wall of Sea Cucumber (Stichopus japonicus). J. Food Sci. 2011, 76, 208–214. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Kim, S.M. α-Glucosidase Inhibitory Activities of Fatty Acids Purified from the Internal Organ of Sea Cucumber Stichopus japonicas. J. Food Sci. 2015, 80, H841–H847. [Google Scholar] [CrossRef]
- Fredalina, B.D.; Ridzwan, B.H.; Abidin, A.A.Z.; Kaswandi, M.A.; Zaiton, H.; Zali, I.; Kittakoop, P.; Jais, A.M.M. Fatty Acid Compositions in Local Sea Cucumber. Gen. Pharmacol. Vasc. Syst. 1999, 33, 337–340. [Google Scholar] [CrossRef]
- Ridzwan, B.H.; Hanita, M.H.; Nurzafirah, M.; Norshuhadaa, M.P.S.; Hanis, Z.F. Free Fatty Acids Composition in Lipid Extracts of Several Sea Cucumbers Species from Malaysia. Int. J. Biosci. Biochem. Bioinform. 2014, 4, 204–207. [Google Scholar] [CrossRef]
- Barber, E.; Houghton, M.J.; Williamson, G. Flavonoids as Human Intestinal α-Glucosidase Inhibitors. Foods 2021, 10, 1939. [Google Scholar] [CrossRef]
- Shah, A.B.; Yoon, S.; Kim, J.H.; Zhumanova, K.; Ban, Y.J.; Lee, K.W.; Park, K.H. Effectiveness of Cyclohexyl Functionality in Ugonins from Helminthostachys zeylanica to PTP1B and α-Glucosidase Inhibitions. Int. J. Biol. Macromol. 2020, 165, 1822–1831. [Google Scholar] [CrossRef] [PubMed]
- Putram, N.M.; Setyaningsih, I.; Tarman, K.; Nursid, M. Aktivitas Antikanker Dari Fraksi Aktif Teripang. J. Pengolah. Has. Perikan. Indones. 2017, 20, 53–62. [Google Scholar] [CrossRef]
- El Barky, A.; Hussein, S.A. Saponins-and-Their-Potential-Role-in-Diabetes-Mellitus. Diabetes Manag. 2017, 7, 148–158. [Google Scholar]
- Barky, E.A.; Ali, E.M.M. Mohamed Marine Sea Cucumber Saponins and Diabetes. Austin Pancreat Disord 2017, 1, 1–7. [Google Scholar]
- Zhang, Y.; Hou, H.; Fan, Y.; Zhang, F.; Li, B.; Xue, C. Effect of Moisture Status on the Stability of Thermal Gels from the Body Wall of Sea Cucumbers (Apostichopus japonicus). LWT-Food Sci. Technol. 2016, 74, 294–302. [Google Scholar] [CrossRef]
- Sottorff, I.; Aballay, A.; Hernández, V.; Roa, L.; Muñoz, L.X.; Silva, M.; Becerra, J.; Astuya, A. Characterization of Bioactive Molecules Isolated from Sea Cucumber Athyonidium chilensis. Rev. Biol. Mar. Oceanogr. 2013, 48, 23–35. [Google Scholar] [CrossRef]
- Rivera-Mondragón, A.; Bijttebier, S.; Tuenter, E.; Custers, D.; Ortíz, O.O.; Pieters, L.; Caballero-George, C.; Apers, S.; Foubert, K. Phytochemical Characterization and Comparative Studies of Four Cecropia Species Collected in Panama Using Multivariate Data Analysis. Sci. Rep. 2019, 9, 1763. [Google Scholar] [CrossRef] [PubMed]
- Dendooven, E.; Foubert, K.; Goossens, A.; Gilles, P.; De Borggraeve, W.; Pieters, L.; Lambert, J.; Aerts, O. Concomitant Positive Patch Test Reactions in FreeStyle-allergic Patients Sensitized to Isobornyl Acrylate. Contact Dermat. 2021, 84, 166–174. [Google Scholar] [CrossRef]
- Su, C.H.; Hsu, C.H.; Ng, L.T. Inhibitory Potential of Fatty Acids on Key Enzymes Related to Type 2 Diabetes. BioFactors 2013, 39, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Trinh, B.T.D.; Staerk, D.; Jager, A.K. Screening for Potential α-Glucosidase and α-Amylase Inhibitory Constituents from Selected Vietnames Plants Used to Treat Type 2 Diabetes. J. Ethnopharmacol. 2016, 186, 189–195. [Google Scholar] [CrossRef]
- Bahrami, Y.; Zhang, W.; Franco, C.; Seafood, A.; Building, M.O.; Park, S.; Biology, M. Discovery of Novel Saponins from the Viscera of the Sea Cucumber Holothuria lessoni. Mar. Drugs 2014, 12, 2633–2667. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, Y.; Zhang, W.; Chataway, T.; Franco, C.; Seafood, A.; Building, M.O.; Park, S.; Biology, M.; Facility, F.P. Structural Elucidation of Novel Saponins in the Sea Cucumber Holothuria lessoni. Mar. Drugs 2014, 12, 4439–4473. [Google Scholar] [CrossRef]
- Caulier, G.; Flammang, P.; Gerbaux, P.; Eeckhaut, I. When a Repellent Becomes an Attractant: Harmful Saponins Are Kairomones Attracting the Symbiotic Harlequin Crab. Sci. Rep. 2013, 3, 2639. [Google Scholar] [CrossRef]
- Caulier, G.; Flammang, P.; Rakotorisoa, P.; Gerbaux, P.; Demeyer, M.; Eeckhaut, I. Preservation of the Bioactive Saponins of Holothuria scabra through the Processing of Trepang. Cah. Biol. Mar. 2013, 54, 685–690. [Google Scholar]
- Han, H.; Yi, Y.; Xu, Q.; La, M.; Zhang, H. Two New Cytotoxic Triterpene Glycosides from the Sea Cucumber Holothuria scabra. Planta Med. 2009, 75, 1608–1612. [Google Scholar] [CrossRef]
- Yuan, W.; Yi, Y.; Tang, H.; Xue, M.; Wang, Z.; Sun, G.; Zhang, W.; Liu, B.; Li, L.; Sun, P. Two New Holostan-Type Triterpene Glycosides from the Sea Cucumber Bohadschia marmorata Jaeger. Chem. Pharm. Bull. 2008, 56, 1207–1211. [Google Scholar] [CrossRef]
- Han, H.; Yi, Y.H.; Li, L.; Wang, X.H.; Liu, B.S.; Sun, P.; Pan, M.X. A New Triterpene Glycoside from Sea Cucumber Holothuria leucospilota. Chin. Chem. Lett. 2006, 18, 161–164. [Google Scholar] [CrossRef]
- Dang, N.H.; Van Thanh, N.; Van Kiem, P.; Huong, L.M.; Van Minh, C.; Kim, Y.H. Two New Triterpene Glycosides from the Vietnamese Sea Cucumber Holothuria scabra. Arch. Pharm. Res. 2007, 30, 1387–1391. [Google Scholar] [CrossRef]
- Hoang, L.; Le Thi, V.; Tran Thi Hong, H.; Nguyen Van, T.; Nguyen Xuan, C.; Nguyen Hoai, N.; Do Cong, T.; Ivanchina, N.V.; Do Thi, T.; Dmitrenok, P.S.; et al. Triterpene Glycosides from the Vietnamese Sea Cucumber Holothuria edulis. Nat. Prod. Res. 2020, 34, 1061–1067. [Google Scholar] [CrossRef] [PubMed]
- Oleinikova, G.K.; Kuznetsova, T.A.; Ivanova, N.S.; Kalinovskii, A.I.; Rovnykh, N.V.; Elyakov, G.B. Glycosides of Marine Invertebrates. XV. A New Triterpene Glycoside—Holothurin A1—From Caribbean Holothurians of the Family Holothuriidae. Chem. Nat. Compd. 1982, 18, 430–434. [Google Scholar] [CrossRef]
- Han, H.; Li, L.; Yi, Y.; Wang, X.; Pan, M. Triterpene Glycosides from Sea Cucumber Holothuria scabra with Cytotoxic Activity. Chin. Herb. Med. 2012, 4, 183–188. [Google Scholar] [CrossRef]
- Assawasuparerk, K.; Rawangchue, T.; Phonarknguen, R. Scabraside D Derived from Sea Cucumber Induces Apoptosis and Inhibits Metastasis via INOS and STAT-3 Expression in Human Cholangiocarcinoma Xenografts. Asian Pac. J. Cancer Prev. 2016, 17, 2151–2157. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Yi, Y.H.; Li, L.; Gui, M.; Tang, H.F.; Zhang, D.Z.; Zhang, S.L. 17-Dehydroxyholothurin A, a New Sulfated Triterpene Glycoside from Sea Cucumber Holothuria Impatiens. Chin. Chem. Lett. 2006, 17, 1454–1456. [Google Scholar]
- Han, H.; Yi, Y.H.; Li, L.; Liu, B.S.; Zhang, H.-W. Antifungal Active Triterpene Glycosides from Sea Cucumber Holothuria scabra. Acta Pharm. Sin. 2009, 44, 620–624. [Google Scholar]
- Han, H.; Yi, Y.H.; Li, L.; Liu, B.S.; Pan, M.X.; Yan, B.; Wang, X.H. Triterpene Glycosides from Sea Cucumber Holothuria leucospilota. Chin. J. Nat. Med. 2009, 7, 346–350. [Google Scholar] [CrossRef]
- Silchenko, A.S.; Stonik, V.A.; Avilov, S.A.; Kalinin, V.I.; Kalinovsky, A.I.; Zakharenko, A.M.; Smirnov, A.V.; Mollo, E.; Cimino, G. Holothurins B2, B3, and B4, New Triterpene Glycosides from Mediterranean Sea Cucumbers of the Genus Holothuria. J. Nat. Prod. 2005, 68, 564–567. [Google Scholar] [CrossRef]
- Bahrami, Y.; Franco, C.M.M.M.; Benkendorff, K. Acetylated Triterpene Glycosides and Their Biological Activity from Holothuroidea Reported in the Past Six Decades. Mar. Drugs 2016, 14, 147. [Google Scholar] [CrossRef] [PubMed]
- Van Thanh, N.; Dang, N.H.; Van Kiem, P.; Cuong, N.X.; Huong, H.T.; Minh, C. Van A New Triterpene Glycoside from the Sea Cucumber Holothuria scabra Collected in Vietnam. ASEAN J. Sci. Technol. Dev. 2006, 23, 253–259. [Google Scholar] [CrossRef]
- Radhika, P.; Anjaneyulu, V.; Subba Rao, P.V.; Makarieva, T.N.; Kalinovosky, A.I. Chemical Examination of the Echinoderms of Indian Ocean: The Triterpene Glycosides of the Sea Cucumbers: Holothuria nobilis, Bohadschia aff. tenuissima and Actinopyga mauritana from Lakshadweep, Andaman and Nicobar Islands. Indian J. Chem.-Sect. B Org. Med. Chem. 2002, 41, 1276–1282. [Google Scholar]
- Kuznetsova, T.A.; Anisimov, M.M.; Popov, A.M.; Baranova, S.I.; Afiyatullov, S.S.; Kapustina, I.I.; Antonov, A.S.; Elyakov, G.B. A Comparative Study in Vitro of Physiological Activity of Triterpene Glycosides of Marine Invertebrates of Echinoderm Type. Comp. Biochem. Physiol. Part C Comp. 1982, 73, 41–43. [Google Scholar] [CrossRef] [PubMed]
- Sroyraya, M.; Kaewphalug, W.; Anantachoke, N.; Poomtong, T.; Sobhon, P.; Srimongkol, A.; Suphamungmee, W. Saponins Enriched in the Epidermal Layer of Holothuria leucospilota Body Wall. Microsc. Res. Tech. 2018, 81, 1182–1190. [Google Scholar] [CrossRef] [PubMed]
- Bondoc, K.G.V.; Lee, H.; Cruz, L.J.; Lebrilla, C.B.; Juinio-Meñez, M.A. Chemical Fingerprinting and Phylogenetic Mapping of Saponin Congeners from Three Tropical Holothurian Sea Cucumbers. Comp. Biochem. Physiol.-B Biochem. Mol. Biol. 2013, 166, 182–193. [Google Scholar] [CrossRef]
- Caulier, G.; Gerbaux, P.; Eeckhaut, I.; Flammang, P.; Caulier, G.; Van Dyck, S.; Gerbaux, P.; Eeckhaut, I.; Flammang, P. Review of Saponin Diversity in Sea Cucumbers Belonging to the Family Holothuriidae. SPC Beche-De-Mer Inf. Bull. 2011, 31, 48–54. [Google Scholar]
- Oleinikova, G.K.; Kuznetsova, T.A.; Rovnykh, N.V.; Kalinovskii, A.I.; Elyakov, G.B. Glycosides of Marine Invertebrates. XVIII. Holothurin A2 from the Caribbean Holothurian Holothuria floridana. Chem. Nat. Compd. 1982, 18, 501–502. [Google Scholar] [CrossRef]
- Yu, S.; Ye, X.; Huang, H.; Peng, R.; Su, Z.; Lian, X.Y.; Zhang, Z. Bioactive Sulfated Saponins from Sea Cucumber Holothuria moebii. Planta Med. 2015, 81, 152–159. [Google Scholar] [CrossRef]
- Elyakov, G.B.; Kalinovskaya, N.I.; Kalinovskii, A.I.; Stonik, V.A.; Kuznetsova, T.A. Glycosides of Marine Invertebrates. XIII. New Holothurinogenins of Holothurin B1 from Holothuria floridana. Chem. Nat. Compd. 1982, 18, 298–302. [Google Scholar] [CrossRef]
- Kitagawa, I.; Akutsu, H.; Kyogoku, Y.; Zubrica, H. Structure of Holothurin B A Pharmacologically Active Triterpene-Oligoglycoside From The Sea Cucumber Holothuria leucospilota Brandt. Tetrahedron Lett. 1978, 3385, 985–988. [Google Scholar] [CrossRef]
- Kitagawa, I.; Nishino, T.; Kobayashi, M.; Matsuno, T.; Akutsu, H.; Kyogoku, Y. Marine Natural Products VII Bioactive Triterpene-Oligoglycosides from the Sea Cucumber Holothuria leucospilota Brandt (1) Structure of Holothurin B. Chem. Pharm. Bull. 1981, 29, 1942–1950. [Google Scholar] [CrossRef]
- Oleinikova, G.K.; Kuznetsova, T.A. Glycosides of the Holothurian Holothuria atra. Chem. Nat. Compd. 1987, 22, 617. [Google Scholar] [CrossRef]
- Putra, Y.; Soffa, F.B.; Firdaus, M.; Pangestuti, R.; Siahaan, E.A. Determination of Fatty Acid Profiles and Bioactive Properties of Body Wall and Viscera of Holothuria atra Collected from Lombok Island, Indonesia. IOP Conf. Ser. Earth Environ. Sci. 2022, 1119, 012052. [Google Scholar] [CrossRef]
- Maier, M.S. Biological Activities of Sulfated Glycosides from Echinoderms. Stud. Nat. Prod. Chem. 2008, 35, 311–354. [Google Scholar] [CrossRef]
- Honey-Escandon, M.; Arreguin-Espinosa, R.; Solis-Marin, F.A.; Samyn, Y. Biological and Taxonomic Perspective of Triterpenoid Glycosides of Sea Cucumbers of the Family Holothuriidae (Echinodermata, Holothuroidea). Comp. Biochem. Physiol. Part-B Biochem. Mol. Biol. 2015, 180, 16–39. [Google Scholar] [CrossRef]
- Zhang, J.-J. Extraction, Isolation, and Structure Elucidation of Two New Triterpene Glycosides from Sea Cucumber Holothuria nobilis. Chin. Tradit. Herb. Drugs 2011, 24, 1467–1472. [Google Scholar]
- Van Dyck, S.; Gerbaux, P.; Flammang, P. Elucidation of Molecular Diversity and Body Distribution of Saponins in the Sea Cucumber Holothuria forskali (Echinodermata) by Mass Spectrometry. Comp. Biochem. Physiol.-B 2009, 152, 124–134. [Google Scholar] [CrossRef]
- Han, H.; Zhang, W.; Yi, Y.H.; Liu, B.S.; Pan, M.X.; Wang, X.H. A Novel Sulfated Holostane Glycoside from Sea Cucumber Holothuria leucospilota. Chem. Biodivers. 2010, 7, 1764–1769. [Google Scholar] [CrossRef] [PubMed]
- Avilov, S.A.; Antonov, A.S.; Drozdova, O.A.; Kalinin, V.I.; Kalinovsky, A.I.; Stonik, V.A.; Riguera, R.; Lenis, L.A.; Jime, C. Triterpene Glycosides from the Far-Eastern Sea Cucumber Pentamera calcigera. 1. Monosulfated Glycosides and Cytotoxicity of Their Unsulfated Derivatives. J. Nat. Prod. 2000, 63, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Assawasuparerk, K.; Vanichviriyakit, R.; Chotwiwatthanakun, C.; Nobsathian, S.; Rawangchue, T.; Wittayachumnankul, B. Scabraside D Extracted from Holothuria Scabra Induces Apoptosis and Inhibits Growth of Human Cholangiocarcinoma Xenografts in Mice. Asian Pac. J. Cancer Prev. 2016, 17, 511–517. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Number | Compound Name | RT (min) | Molecular Formula | Nominal Mass | Ion | Measured m/z | Error (ppm) | Fragmentation Pattern | Reference | Sample |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 17-hydroxyfuscocineroside B (=scabraside B) * or 25-hydroxyfuscocineroside B * | 7.56 | C54H85O27Sna | 1220 | [M-Na]- | 1197.4982 | −1.67 | 875.3718 [M-Na-322 (MeGlc+Qui)]- | [58,59,60,61,62,63] | c, f |
| 2 | Leucospilotaside A * | 7.87 | C41H63O18Sna | 898 | [M-Na]- | 875.371 | 4.66 | 729.3176 [M-Na-146(Qui)]- 755.2563 [M-Na-120(NaHSO4)]- | [58,64] | c, d, e, f, g |
| 3 | Holothurin A3 * or Holothurin A5 (II)* | 8.53 | C54H85O28Sna | 1236 | [M-Na]- | 1213.4944 | −0.66 | 875.3729 [M-Na-338 (MeGlc+Glc)]- | [65,66] | d |
| 4 | Holothurin A1 *, or Holothurin A4 * or Scabraside D * | 9.06 | C54H87O27Sna | 1222 | [M-Na]- | 1199.5132 | −1.66 | 861.3885 [M-Na-338 (MeGlc+Glc)]- 843.3527 [M-Na-356 (MeGlc+Glc+H2O)]- | [58,65,67,68,69] | d, f |
| 5 | Calcigeroside B | 9.59 | C54H83O27Sna | 1218 | [M-Na]- | 1195.486 | 1.51 | Not observed | [37] | d, f |
| 6 | 17-dehydroxyholothurin A * (=fuscocineroside C *), or Scabraside A *, or 24-dehydroechinoside A *, or Fuscocineroside B * | 9.94 | C54H85O26Sna | 1204 | [M-Na]- | 1181.5032 | −1.69 | Not observed | [7,11,37,58,60,61,68,70,71] | e, g |
| 7 | Holothurin B2 | 10.27 | C41H65O17Sna | 884 | [M-Na]- | 861.3938 | −0.46 | 715.7407 [M-Na-146(Qui)]- | [72,73] | c, f, g |
| 8 | Holothurin B, or Holothurin B4, or Nobiliside II (=ananaside C) * | 10.70 | C41H63O17Sna | 882 | [M-Na]- | 859.3801 | 1.75 | 713.3293 [M-Na-146(Qui)]- | [11,12,37,58,66,73,74,75,76,77,78] | a, b, c, d, f, g |
| 9 | Echinoside A (=Holothurin A2) | 10.89 | C54H87O26Sna | 1206 | [M-Na]- | 1183.5178 | −2.03 | 1165.5055 [M-Na-18(H2O)]- | [60,71,76,79,80,81] | d, e, f |
| 10 | Holothurin B3 or 24-dehydroechinoside B * | 11.02 | C41H63O16Sna | 866 | [M-Na]- | 843.3819 | −2.13 | 697.3291 [M-Na-146(Qui)]- | [12,73,74,82] | c, d, e, f, g |
| 11 | Echinoside B (=Holothurin B1) | 12.49 | C41H65O16Sna | 868 | [M-Na]- | 845.3994 | 0.11 | 827.3956 [M-Na-18(H2O)]- 695.4776 [M-Na-150(Xyl)]- 725.5139 [M-Na-120(NaHSO4)]- | [12,36,67,72,77,78,83] | c, d, e, f, g |
| Compound Number | Compound Name | Sapogenin* | R1 * | R2 * | R3 * |
|---|---|---|---|---|---|
| 1 | 17-hydroxyfuscocineroside B (=scabraside B) or | A | OH | 8 | b |
| 25-hydroxyfuscocineroside B | A | H | 9 | b | |
| 2 | Leucospilotaside A | A | OH | 9 | a |
| Holothurin A3 or | A | OH | 9 | b | |
| 3 | Holothurin A5 | A | OH | 11 | b |
| 4 | Holothurin A1 or | A | OH | 5 | b |
| Holothurin A4 or | A | OH | 6 | b | |
| Scabraside D | A | OH | 7 | b | |
| 5 | Calcigeroside B | Atypical | |||
| 6 | 17-dehydroxyholothurin A (=fuscocineroside C) or | A | H | 1 | b |
| Scabraside A or | A | OH | 4 | b | |
| 24-dehydroechinoside A or | A | OH | 3 | b | |
| Fuscocineroside B | A | H | 1 | b | |
| 7 | Holothurin B2 | A | OH | 5 | a |
| 8 | Holothurin B, or | A | OH | 1 | a |
| Holothurin B4, or | A | OH | 10 | a | |
| Nobiliside II (=ananaside C) | Atypical | 1 | a | ||
| 9 | Echinoside A (=Holothurin A2) | A | OH | 2 | b |
| 10 | Holothurin B3 or | A | H | 1 | a |
| 24-dehydroechinoside B | A | OH | 3 | a | |
| 11 | Echinoside B (Holothurin B1) | A | OH | 2 | a |
| 1 | 2 | |||||
|---|---|---|---|---|---|---|
| δ 13C (ppm) | δ1H (ppm) | δ 13C (ppm) | δ 1H (ppm) | |||
| 1 | 37.3 | 1.85/1.51 | (1H */1H *) | 36.5 | 1.71/1.47 | (1H */1H *) |
| 2 | 27.7 | 1.97/1.79 | (1H */1H *) | 27.2 | 2.18/1.93 | (1H */1H *) |
| 3 | 90.3 | 3.13 | (1H, d, 11.6) | 89.0 | 3.24 | (1H *) |
| 4 | 40.8 | 40.3 | ||||
| 5 | 53.8 | 0.98 | (1H, d, 11.2) | 53.3 | 0.98 | (1H, d, 11.7) |
| 6 | 22.0 | 1.77/1.57 | (1H */1H *) | 21.4 | 1.72/1.59 | (1H */1H *) |
| 7 | 29.0 | 1.78/1.46 | (1H */1H *) | 27.7 | 1.70 | (2H *) |
| 8 | 41.9 | 3.01 | (1H *) | 40.8 | 3.26 | (1H *) |
| 9 | 155.6 | 151.2 | ||||
| 10 | 40.6 | 39.5 | ||||
| 11 | 115.4 | 5.38 | (1H, d, 4.8) | 119.5 | 5.62 | (1H, br s) |
| 12 | 72.5 | 4.53 | (1H *) | 66.9 | 5.25 | (1H, br s) |
| 13 | 59.7 | 65.3 | ||||
| 14 | 46.5 | 48.7 | ||||
| 15 | 37.5 | 1.79/1.16 | (1H */1H *) | 37.0 | 1.75/1.40 | (1H */1H *) |
| 16 | 35.9 | 2.80/2.07 | (1H, dd, 15.1; 8.5/1H *) | 37.3 | 3.11/2.35 | (1H, dd, 14.5;7.2/1H, m) |
| 17 | 90.5 | 87.4 | ||||
| 18 | 176.2 | N.O. | ||||
| 19 | 22.8 | 1.15 | (3H, s) | 22.0 | 1.40 | (3H, s) |
| 20 | 87.6 | 87.4 | ||||
| 21 | 18.7 | 1.50 | (3H, s) | 16.9 | 2.04 | (3H, s) |
| 22 | 81.5 | 4.20 | (1H, t, 7.4) | 81.7 | 4.27 | (1H *) |
| 23 | 28.6 | 2.04 | (2H*) | 28.4 | 2.00 | (2H*) |
| 24 | 39.2 | 1.78 | (2H*) | 38.6 | 1.56 | (2H*) |
| 25 | 82.9 | 81.5 | ||||
| 26 | 28.9 | 1.30 | (3H, s) | 28.6 | 1.02 | (3H, s) |
| 27 | 27.6 | 1.25 | (3H, s) | 27.7 | 1.13 | (3H, s) |
| 28 | 17.1 | 0.92 | (3H, s) | 16.9 | 2.04 | (3H, s) |
| 29 | 28.6 | 1.09 | (3H, s) | 28.2 | 1.31 | (3H, s) |
| 30 | 20.4 | 1.31 | (3H, s) | 19.9 | 1.37 | (3H, s) |
| 1’ | 106.0 | 4.42 | (1H, d, 6.8) | 105.8 | 4.78 | (1H, d, 7.3) |
| 2’ | 83.0 | 3.47 | (1H *) | 84.1 | 4.08 | (1H *) |
| 3’ | 77.8 | 3.52 | (1H *) | 78.2 | 4.16 | (1H *) |
| 4’ | 71.1 | 3.52 | (1H *) | 71.0 | 4.14 | (1H *) |
| 5’ | 66.4 | 3.86/3.21 | (1H, dd, 11.5; 4.0/1H *) | 66.8 | 4.24/3.61 | (1H */1H, t, 10.1) |
| 1’’ | 105.6 | 4.55 | (1H) | 106.4 | 5.18 | (1H *) |
| 2’’ | 76.8 | 3.25 | (1H *) | 77.3 | 4.08 | (1H *) |
| 3’’ | 77.5 | 3.33 | (1H *) | 77.9 | 4.10 | (1H *) |
| 4’’ | 77.0 | 2.99 | (1H, t, 9.0) | 76.8 | 3.70 | (1H *) |
| 5’’ | 73.7 | 3.29 | (1H *) | 73.6 | 3.78 | (1H *) |
| 6’’ | 18.2 | 1.27 | (3H, s) | 18.8 | 1.65 | (3H, s) |
| Compound | Molecular Formula |
|---|---|
| Myristic acid | C14H28O2/C14:0 |
| Pentadecanoic acid | C15H30O2/C15:0 |
| Palmitoleic acid | C16H30O2/C16:1 ω-7 |
| Palmitic acid | C16H32O2/C16:0 |
| Stearic acid | C18H36O2/C18:0 |
| Arachidonic acid | C20H32O2/C20:4n-6 ω-6 |
| Eicosapentaenoic acid | C20H30O2/C20:5n-3 ω-3 |
| Arachidic acid | C20H40O2/C20:0 |
| Heneicosanoic acid * | C21H42O2/C21:0 |
| Behenic acid | C22H44O2/C22:0 |
| Nervonic acid * | C24H46O2/C24:1n-9 ω9 |
| Sample | IC50 (mg/mL) |
|---|---|
| Crude extract | >20 |
| MB subfraction | >10 |
| MC subfraction | >10 |
| ME subfraction | 0.469 ± 0.029 c,d |
| MF subfraction | 1.599 ± 0.361 b |
| MG subfraction | 0.181 ± 0.006 d |
| Hex fraction | >20 |
| Me fraction | >20 |
| Bu fraction | 0.779 ± 0.066 c |
| Aq fraction | > 20 |
| Me90 fraction | 0.158 ± 0.002 d |
| Acarbose | 2.340 ± 0.044 a |
| Sample | IC50 (mM) |
|---|---|
| Palmitoleic acid | 0.320 ± 0.032 b |
| Arachidonic acid | 0.596 ± 0.044 b |
| Eicosapentaenoic acid | 0.541 ± 0.039 b |
| Acarbose | 3.533 ± 0.462 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puspitasari, Y.E.; Tuenter, E.; Foubert, K.; Herawati, H.; Hariati, A.M.; Aulanni’am, A.; Pieters, L.; De Bruyne, T.; Hermans, N. Saponin and Fatty Acid Profiling of the Sea Cucumber Holothuria atra, α-Glucosidase Inhibitory Activity and the Identification of a Novel Triterpene Glycoside. Nutrients 2023, 15, 1033. https://doi.org/10.3390/nu15041033
Puspitasari YE, Tuenter E, Foubert K, Herawati H, Hariati AM, Aulanni’am A, Pieters L, De Bruyne T, Hermans N. Saponin and Fatty Acid Profiling of the Sea Cucumber Holothuria atra, α-Glucosidase Inhibitory Activity and the Identification of a Novel Triterpene Glycoside. Nutrients. 2023; 15(4):1033. https://doi.org/10.3390/nu15041033
Chicago/Turabian StylePuspitasari, Yunita Eka, Emmy Tuenter, Kenn Foubert, Herawati Herawati, Anik Martinah Hariati, Aulanni’am Aulanni’am, Luc Pieters, Tess De Bruyne, and Nina Hermans. 2023. "Saponin and Fatty Acid Profiling of the Sea Cucumber Holothuria atra, α-Glucosidase Inhibitory Activity and the Identification of a Novel Triterpene Glycoside" Nutrients 15, no. 4: 1033. https://doi.org/10.3390/nu15041033
APA StylePuspitasari, Y. E., Tuenter, E., Foubert, K., Herawati, H., Hariati, A. M., Aulanni’am, A., Pieters, L., De Bruyne, T., & Hermans, N. (2023). Saponin and Fatty Acid Profiling of the Sea Cucumber Holothuria atra, α-Glucosidase Inhibitory Activity and the Identification of a Novel Triterpene Glycoside. Nutrients, 15(4), 1033. https://doi.org/10.3390/nu15041033