
Adipocyte and Adipokines Promote a Uterine Leiomyoma Friendly Microenvironment



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells Isolation and Culture
2.2. Coculture and Treatment
2.3. Multiplex Cytokine Array
2.4. Western Blot Analyses
2.5. Immunocytochemistry
2.6. Statistical Analysis
3. Results
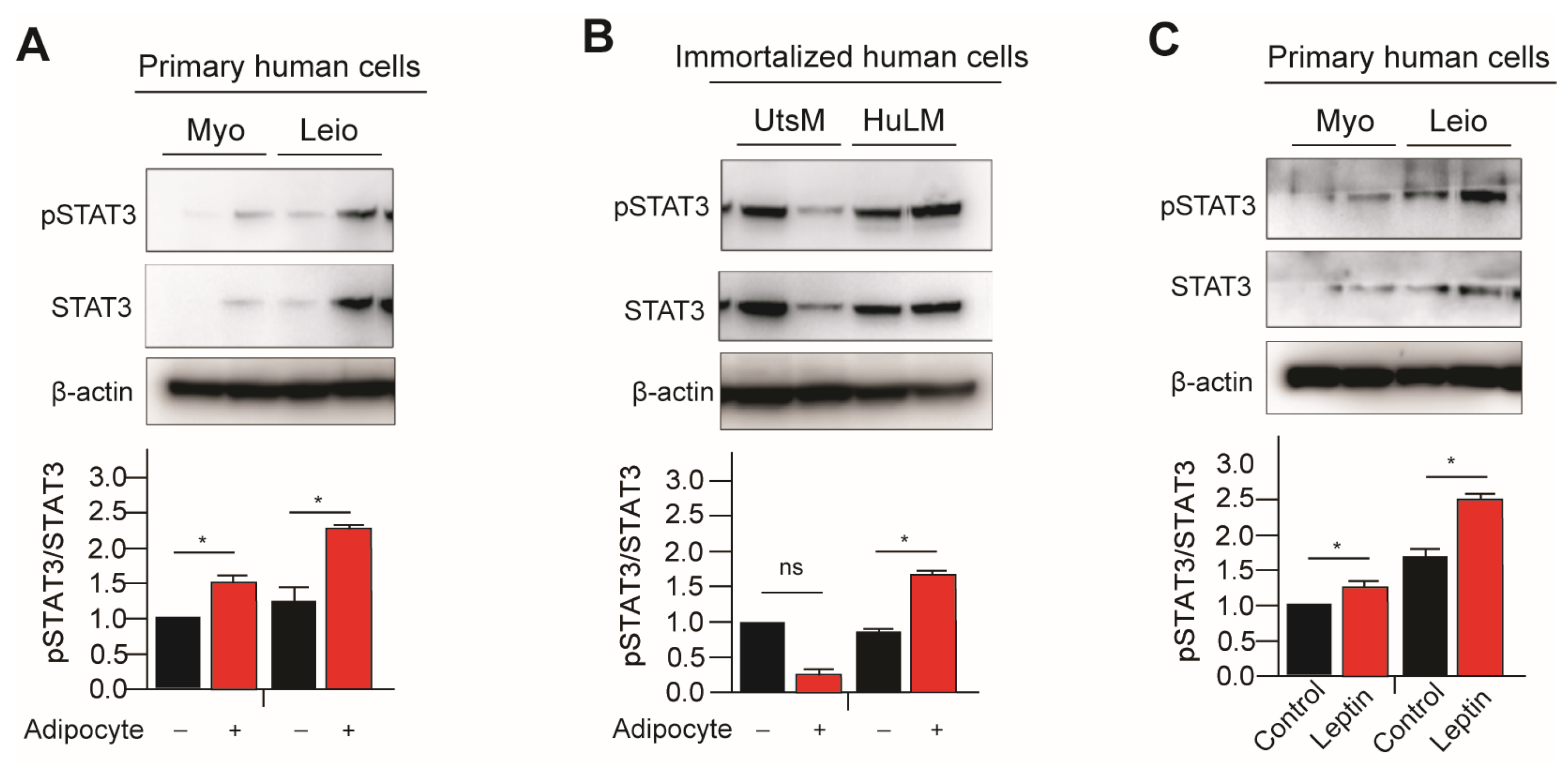
3.1. Adipocyte Coculture and Leptin Treatment with Myometrium and Leiomyomas Cells Activates JAK2/STAT3 Signaling Pathways
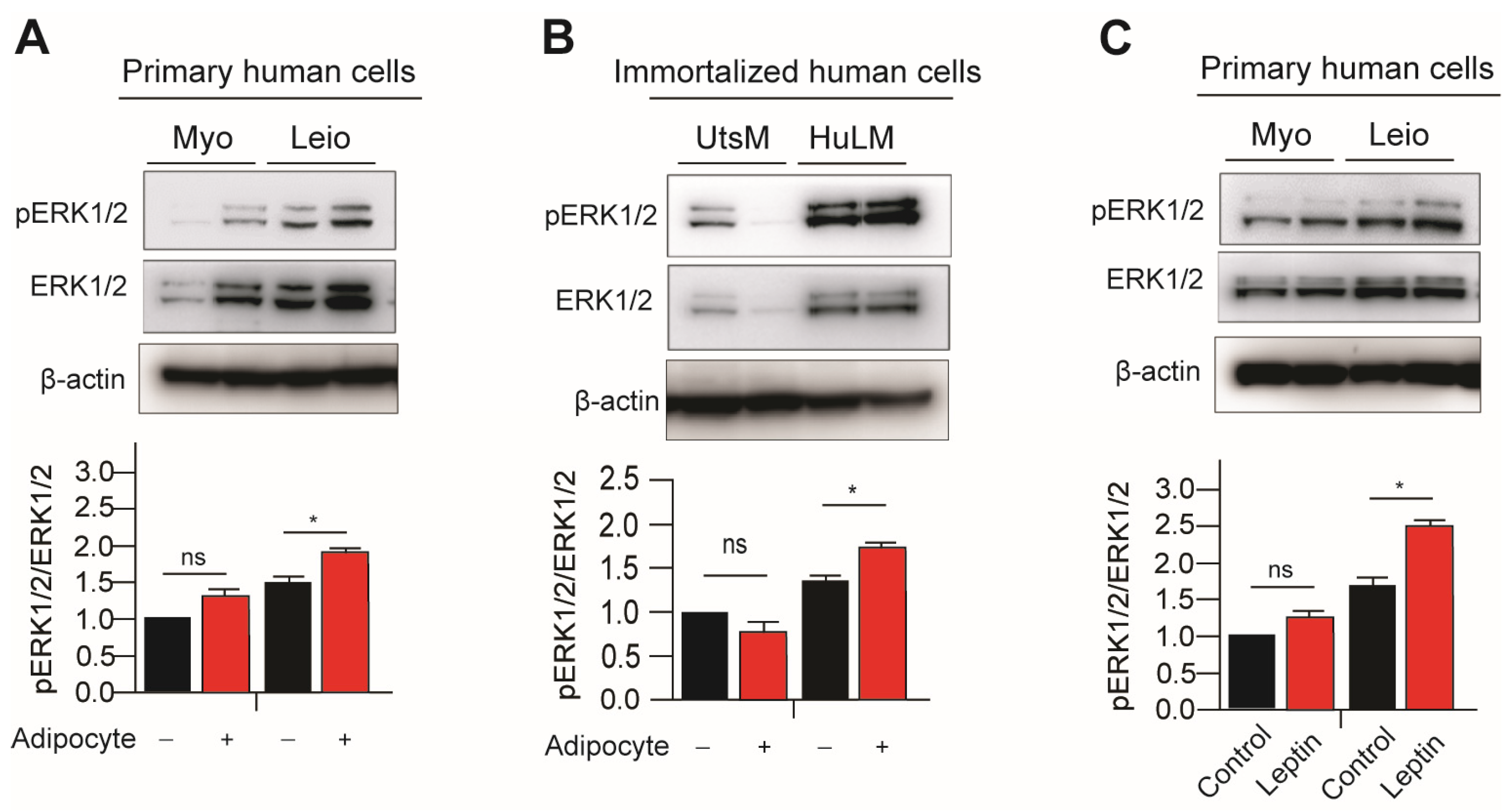
3.2. Leptin Treatment and Adipocyte Coculture with Myometrium and Leiomyomas Cells Activates MAPK/ERK Signaling Pathways
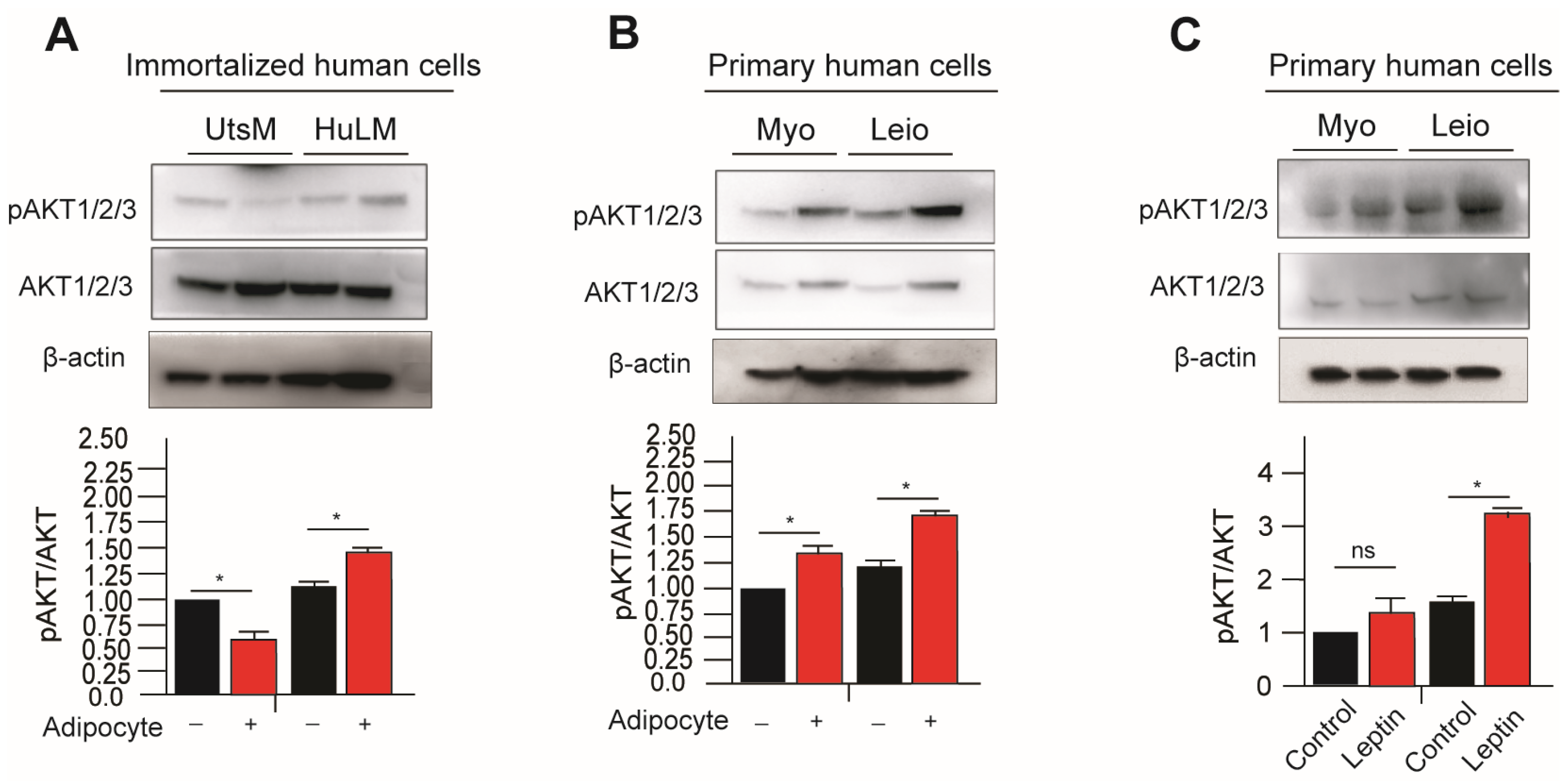
3.3. Leptin Treatment and Adipocyte Coculture Further Activates PI3K/AKT Signaling in Myometrium and Leiomyoma Cells
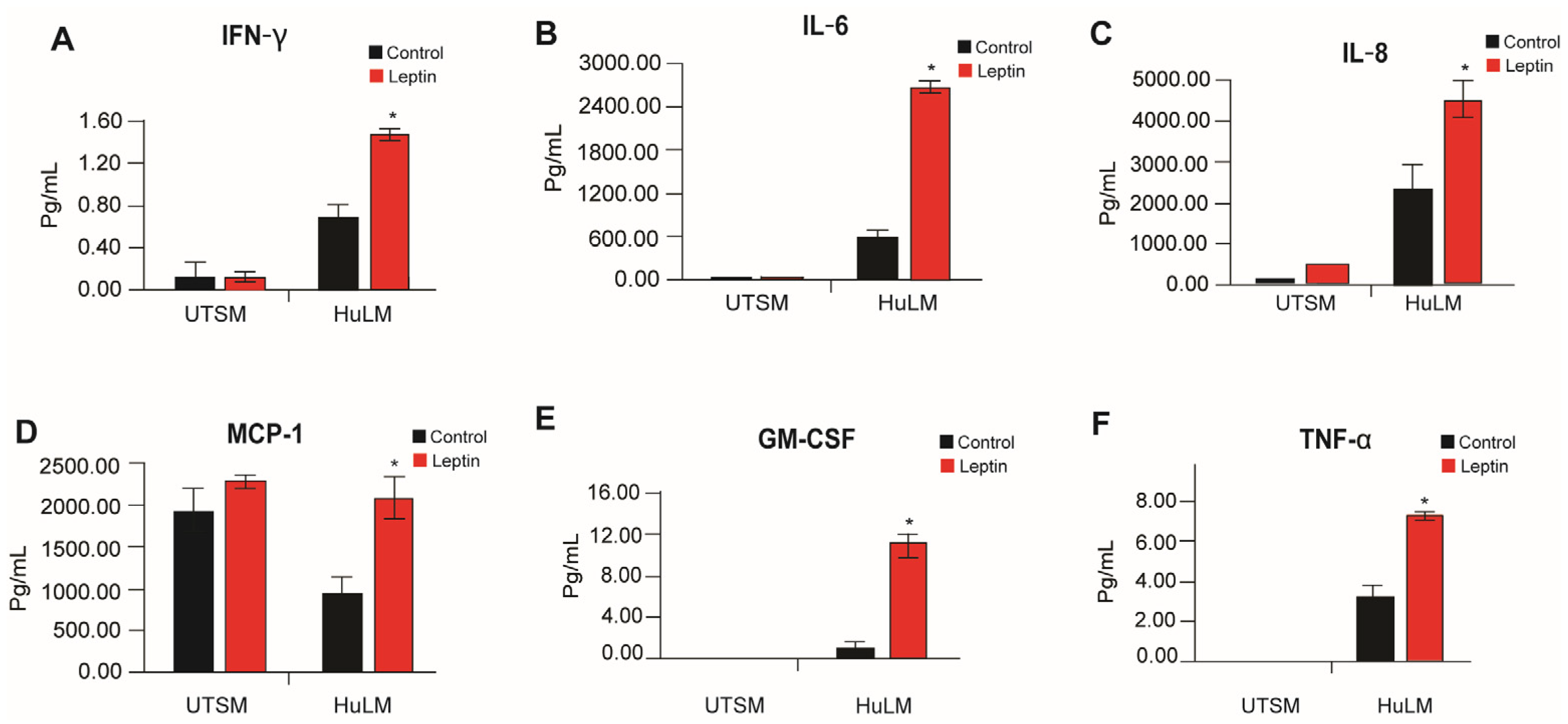
3.4. Effect of Leptin on Pro-Inflammatory Cytokines in Uterine Myometrium and Leiomyoma Cells
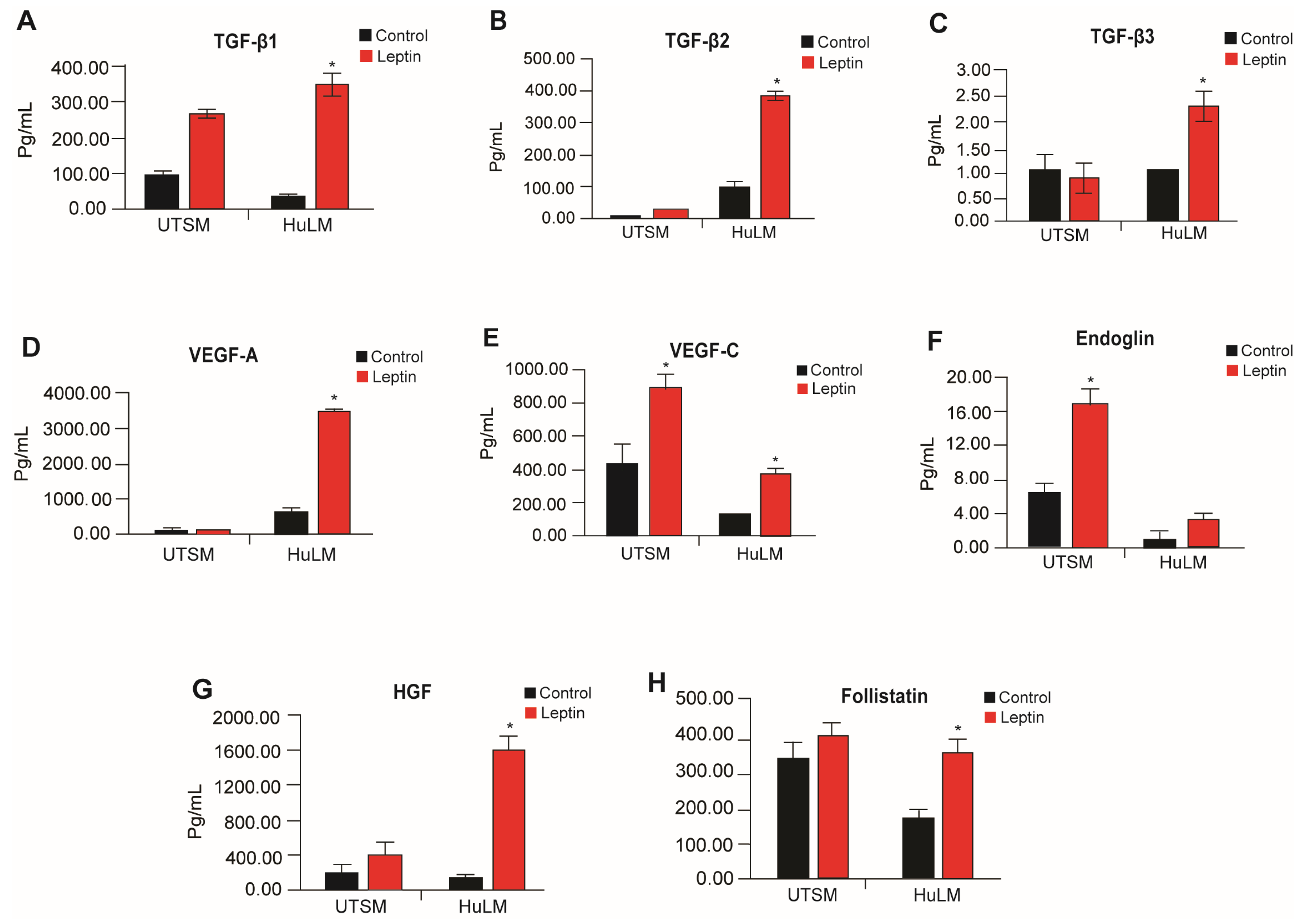
3.5. Effect of Leptin on Angiogenic and Pro-Fibrotic Mediators in Uterine Leiomyoma and Myometrium Cells
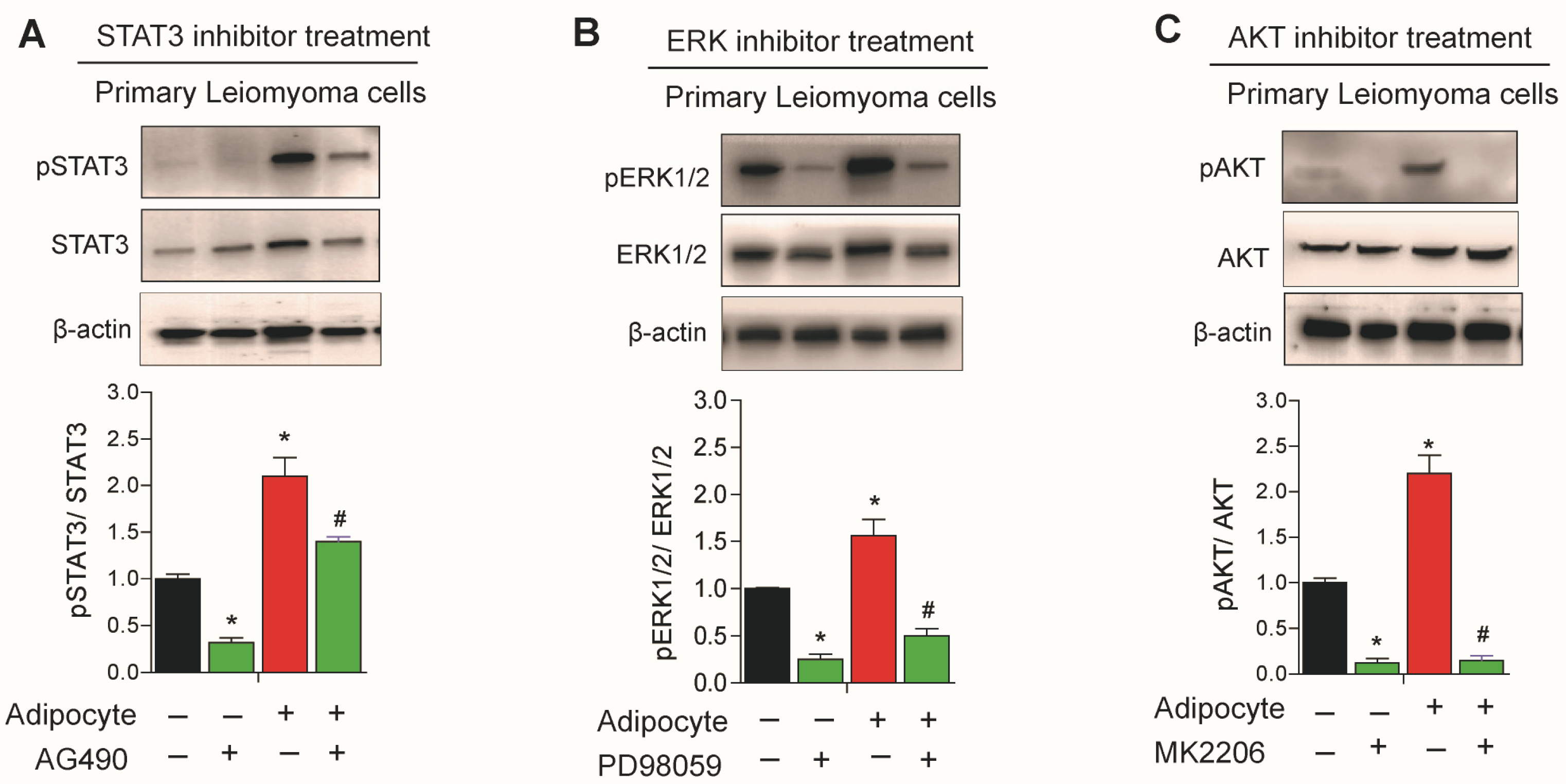
3.6. Effect of STAT3, ERK, and AKT Inhibitors on Leiomyoma Cells Cocultured with Adipocytes
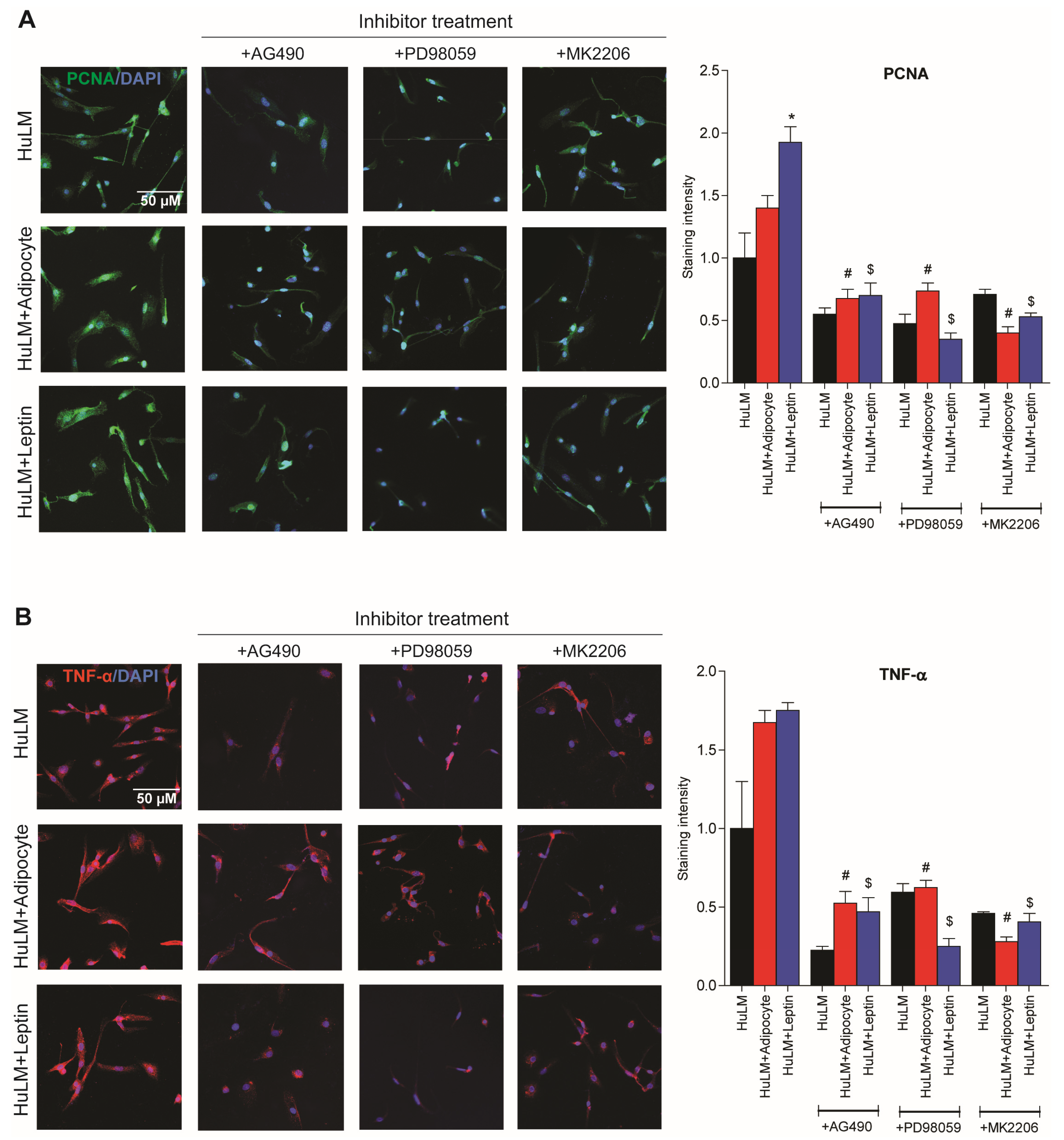
3.7. Effect of STAT3, ERK, and AKT Inhibitors on PCNA and TNF-α Expression
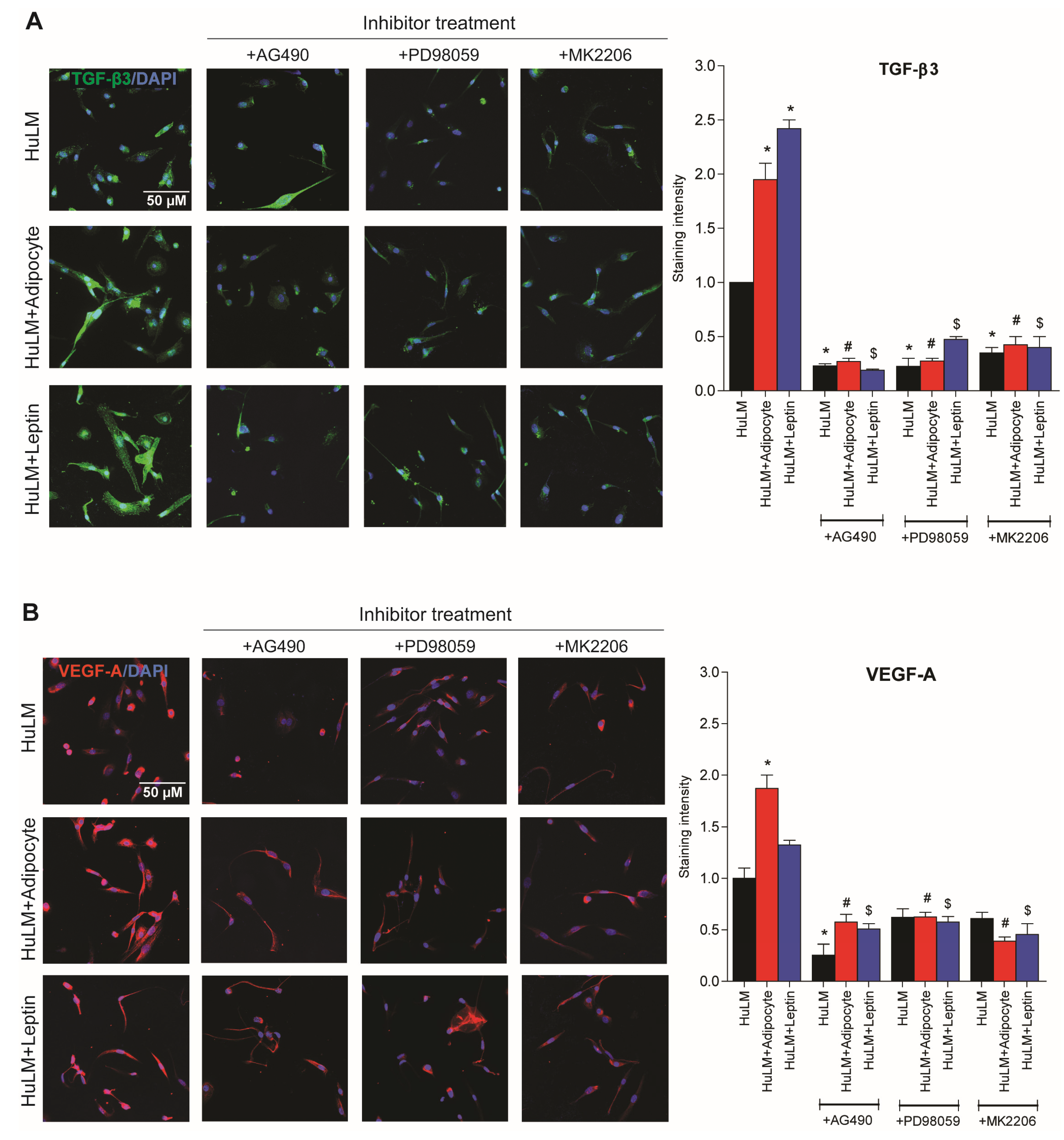
3.8. Effect of STAT3, ERK, and AKT Inhibitors on TGF-β3 and VEGF-A Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baird, D.D.; Dunson, D.B.; Hill, M.C.; Cousins, D.; Schectman, J.M. High Cumulative Incidence of Uterine Leiomyoma in Black and White Women: Ultrasound Evidence. Am. J. Obstet. Gynecol. 2003, 188, 100–107. [Google Scholar] [CrossRef]
- Cardozo, E.R.; Clark, A.D.; Banks, N.K.; Henne, M.B.; Stegmann, B.J.; Segars, J.H. The Estimated Annual Cost of Uterine Leiomyomata in the United States. Am. J. Obstet. Gynecol. 2012, 206, 211.e1–211.e9. [Google Scholar] [CrossRef]
- Booth, A.; Magnuson, A.; Fouts, J.; Foster, M. Adipose Tissue, Obesity and Adipokines: Role in Cancer Promotion. Horm. Mol. Biol. Clin. Investig. 2015, 21, 57–74. [Google Scholar] [CrossRef]
- AlAshqar, A.; Patzkowsky, K.; Afrin, S.; Wild, R.; Taylor, H.S.; Borahay, M.A. Cardiometabolic Risk Factors and Benign Gynecologic Disorders. Obstet. Gynecol. Surv. 2019, 74, 661–673. [Google Scholar] [CrossRef]
- He, Y.; Zeng, Q.; Li, X.; Liu, B.; Wang, P. The Association between Subclinical Atherosclerosis and Uterine Fibroids. PLoS ONE 2013, 8, e57089. [Google Scholar] [CrossRef]
- Sato, F.; Nishi, M.; Kudo, R.; Miyake, H. Body Fat Distribution and Uterine Leiomyomas. J. Epidemiol. 1998, 8, 176–180. [Google Scholar] [CrossRef]
- Qin, H.; Lin, Z.; Vásquez, E.; Luan, X.; Guo, F.; Xu, L. Association between Obesity and the Risk of Uterine Fibroids: A Systematic Review and Meta-Analysis. J. Epidemiol. Community Health 2021, 75, 197–204. [Google Scholar] [CrossRef]
- Peristats|March of Dimes. Available online: https://www.marchofdimes.org/peristats/ (accessed on 3 October 2022).
- Reschke, L.; Afrin, S.; El Sabah, M.; Charewycz, N.; Miyashita-Ishiwata, M.; Borahay, M.A. Leptin Induces Leiomyoma Cell Proliferation and Extracellular Matrix Deposition via JAK2/STAT3 and MAPK/ERK Pathways. FS Sci. 2022, 3, 383–391. [Google Scholar] [CrossRef]
- Afrin, S.; El Sabah, M.; Manzoor, A.; Miyashita-Ishiwata, M.; Reschke, L.; Borahay, M.A. Adipocyte Coculture Induces a Pro-Inflammatory, Fibrotic, Angiogenic, and Proliferative Microenvironment in Uterine Leiomyoma Cells. Biochim. Biophys. Acta Mol. Basis Dis. 2023, 1869, 166564. [Google Scholar] [CrossRef]
- Afrin, S.; Islam, M.S.; Patzkowsky, K.; Malik, M.; Catherino, W.H.; Segars, J.H.; Borahay, M.A. Simvastatin Ameliorates Altered Mechanotransduction in Uterine Leiomyoma Cells. Am. J. Obstet. Gynecol. 2020, 223, e1–e733. [Google Scholar] [CrossRef]
- Afrin, S.; El Sabeh, M.; Islam, M.S.; Miyashita-Ishiwata, M.; Malik, M.; Catherino, W.H.; Akimzhanov, A.M.; Boehning, D.; Yang, Q.; Al-Hendy, A.; et al. Simvastatin Modulates Estrogen Signaling in Uterine Leiomyoma via Regulating Receptor Palmitoylation, Trafficking and Degradation. Pharmacol. Res. 2021, 172, 105856. [Google Scholar] [CrossRef]
- Carney, S.A.; Tahara, H.; Swartz, C.D.; Risinger, J.I.; He, H.; Moore, A.B.; Haseman, J.K.; Barrett, J.C.; Dixon, D. Immortalization of Human Uterine Leiomyoma and Myometrial Cell Lines after Induction of Telomerase Activity: Molecular and Phenotypic Characteristics. Lab. Investig. 2002, 82, 719–728. [Google Scholar] [CrossRef]
- Nair, S.; Al-Hendy, A. Adipocytes Enhance the Proliferation of Human Leiomyoma Cells Via TNF-α Proinflammatory Cytokine. Reprod. Sci. 2011, 18, 1186–1192. [Google Scholar] [CrossRef]
- Richardson, M.A.; Berg, D.T.; Johnston, P.A.; McClure, D.; Grinnell, B.W. Human Liposarcoma Cell Line, SW872, Secretes Cholesteryl Ester Transfer Protein in Response to Cholesterol. J. Lipid Res. 1996, 37, 1162–1166. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Borahay, M.A.; Al-Hendy, A.; Kilic, G.S.; Boehning, D. Signaling Pathways in Leiomyoma: Understanding Pathobiology and Implications for Therapy. Mol. Med. Camb. Mass 2015, 21, 242–256. [Google Scholar] [CrossRef]
- Crespi, E.; Bottai, G.; Santarpia, L. Role of Inflammation in Obesity-Related Breast Cancer. Curr. Opin. Pharmacol. 2016, 31, 114–122. [Google Scholar] [CrossRef]
- Orciani, M.; Caffarini, M.; Biagini, A.; Lucarini, G.; Delli Carpini, G.; Berretta, A.; Di Primio, R.; Ciavattini, A. Chronic Inflammation May Enhance Leiomyoma Development by the Involvement of Progenitor Cells. Stem Cells Int. 2018, 2018, 1716246. [Google Scholar] [CrossRef]
- Divella, R.; De Luca, R.; Abbate, I.; Naglieri, E.; Daniele, A. Obesity and Cancer: The Role of Adipose Tissue and Adipo-Cytokines-Induced Chronic Inflammation. J. Cancer 2016, 7, 2346–2359. [Google Scholar] [CrossRef]
- Doerstling, S.S.; O’Flanagan, C.H.; Hursting, S.D. Obesity and Cancer Metabolism: A Perspective on Interacting Tumor–Intrinsic and Extrinsic Factors. Front. Oncol. 2017, 7, 216. [Google Scholar] [CrossRef]
- Ghasemi, A.; Saeidi, J.; Azimi-Nejad, M.; Hashemy, S.I. Leptin-Induced Signaling Pathways in Cancer Cell Migration and Invasion. Cell. Oncol. 2019, 42, 243–260. [Google Scholar] [CrossRef]
- Park, H.-K.; Ahima, R.S. Leptin Signaling. F1000Prime Rep. 2014, 6, 73. [Google Scholar] [CrossRef]
- Saxena, N.K.; Sharma, D.; Ding, X.; Lin, S.; Marra, F.; Merlin, D.; Anania, F.A. Concomitant Activation of the JAK/STAT, PI3K/AKT, and ERK Signaling Is Involved in Leptin-Mediated Promotion of Invasion and Migration of Hepatocellular Carcinoma Cells. Cancer Res. 2007, 67, 2497–2507. [Google Scholar] [CrossRef]
- Frühbeck, G. Intracellular Signalling Pathways Activated by Leptin. Biochem. J. 2006, 393, 7–20. [Google Scholar] [CrossRef]
- Haque, I.; Ghosh, A.; Acup, S.; Banerjee, S.; Dhar, K.; Ray, A.; Sarkar, S.; Kambhampati, S.; Banerjee, S.K. Leptin-Induced ER-α-Positive Breast Cancer Cell Viability and Migration Is Mediated by Suppressing CCN5-Signaling via Activating JAK/AKT/STAT-Pathway. BMC Cancer 2018, 18, 99. [Google Scholar] [CrossRef]
- Islam, M.S.; Ciavattini, A.; Petraglia, F.; Castellucci, M.; Ciarmela, P. Extracellular Matrix in Uterine Leiomyoma Pathogenesis: A Potential Target for Future Therapeutics. Hum. Reprod. Update 2018, 24, 59–85. [Google Scholar] [CrossRef]
- Ciebiera, M.; Włodarczyk, M.; Wrzosek, M.; Wojtyła, C.; Błażej, M.; Nowicka, G.; Łukaszuk, K.; Jakiel, G. TNF-α Serum Levels Are Elevated in Women with Clinically Symptomatic Uterine Fibroids. Int. J. Immunopathol. Pharmacol. 2018, 32, 2058738418779461. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Afrin, S.; Ramaiyer, M.; Begum, U.A.M.; Borahay, M.A. Adipocyte and Adipokines Promote a Uterine Leiomyoma Friendly Microenvironment. Nutrients 2023, 15, 715. https://doi.org/10.3390/nu15030715
Afrin S, Ramaiyer M, Begum UAM, Borahay MA. Adipocyte and Adipokines Promote a Uterine Leiomyoma Friendly Microenvironment. Nutrients. 2023; 15(3):715. https://doi.org/10.3390/nu15030715
Chicago/Turabian StyleAfrin, Sadia, Malini Ramaiyer, Umme Aoufa Mafruha Begum, and Mostafa A. Borahay. 2023. "Adipocyte and Adipokines Promote a Uterine Leiomyoma Friendly Microenvironment" Nutrients 15, no. 3: 715. https://doi.org/10.3390/nu15030715
APA StyleAfrin, S., Ramaiyer, M., Begum, U. A. M., & Borahay, M. A. (2023). Adipocyte and Adipokines Promote a Uterine Leiomyoma Friendly Microenvironment. Nutrients, 15(3), 715. https://doi.org/10.3390/nu15030715