
Monitoring Yellow Mealworm (Tenebrio molitor) as a Potential Novel Allergenic Food: Effect of Food Processing and Matrix





Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Preparation of Reference and Validation Model Mixtures
2.3. DNA Extraction
2.4. Selection of Yellow Mealworm Genetic Markers
2.5. Qualitative PCR
2.6. Real-Time PCR
2.7. Statistical Analysis
3. Results
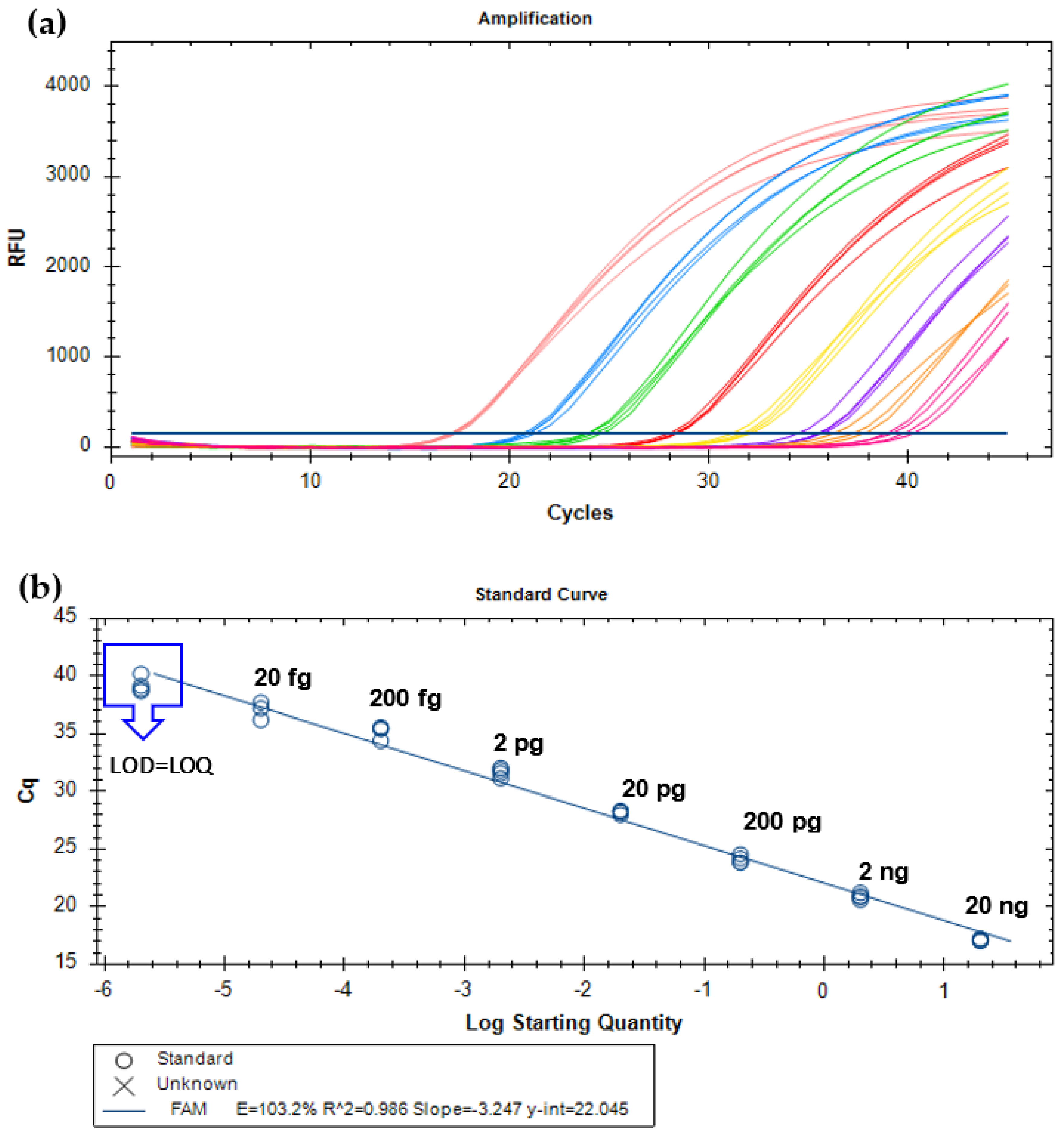
3.1. Specificity and Sensitivity Evaluation
3.2. Development of Real-Time PCR Systems
3.3. Validation of the Method
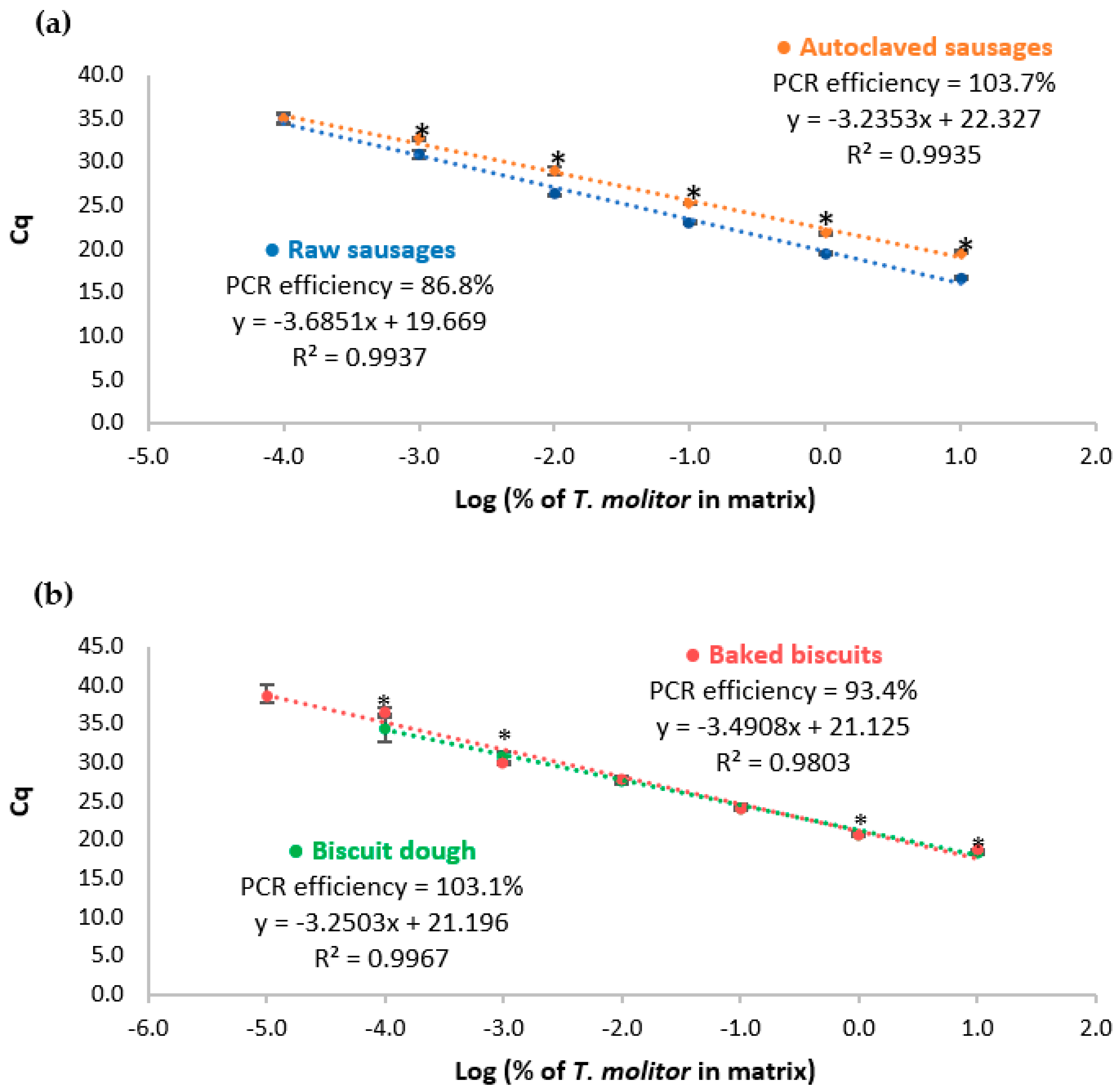
3.4. Thermal Processing and Matrix Effects
3.5. Analysis of Commercial Samples
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pali-Schöll, I.; Verhoeckx, K.; Mafra, I.; Bavaro, S.L.; Clare Mills, E.N.; Monaci, L. Allergenic and novel food proteins: State of the art and challenges in the allergenicity assessment. Trends Food Sci. Technol. 2019, 84, 45–48. [Google Scholar] [CrossRef]
- Patel, S.; Suleria, H.A.R.; Rauf, A. Edible insects as innovative foods: Nutritional and functional assessments. Trends Food Sci. Technol. 2019, 86, 352–359. [Google Scholar] [CrossRef]
- Bessa, L.W.; Pieterse, E.; Sigge, G.; Hoffman, L.C. Insects as human food; from farm to fork. J. Sci. Food Agric. 2020, 100, 5017–5022. [Google Scholar] [CrossRef] [PubMed]
- De Marchi, L.; Wangorsch, A.; Zoccatelli, G. Allergens from edible insects: Cross-reactivity and effects of processing. Curr. Allergy Asthma Rep. 2021, 21, 35. [Google Scholar] [CrossRef] [PubMed]
- Mishyna, M.; Chen, J.; Benjamin, O. Sensory attributes of edible insects and insect-based foods—Future outlooks for enhancing consumer appeal. Trends Food Sci. Technol. 2020, 95, 141–148. [Google Scholar] [CrossRef]
- van der Fels-Klerx, H.J.; Camenzuli, L.; Belluco, S.; Meijer, N.; Ricci, A. Food safety issues related to uses of insects for feeds and foods. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1172–1183. [Google Scholar] [CrossRef] [PubMed]
- Regulation (EU)2015/2283 of the European Parliament and of the Council of 25 November 2015 on novel foods, amending Regulation (EU) No 1169/2011 of the European Parliament and of the Council and repealing Regulation (EC) No 258/97 of the European Parliament and of the Council and Commission Regulation (EC) No 1852/2001. Off. J. Eur. Union 2015, L 327, 1–22.
- Turck, D.; Bresson, J.L.; Burlingame, B.; Dean, T.; Fairweather-Tait, S.; Heinonen, M.; Hirsch-Ernst, K.I.; Mangelsdorf, I.; McArdle, H. Guidance on the preparation and presentation of an application for authorisation of a novel food in the context of Regulation (EU) 2015/2283. Efsa J. 2016, 14, e04594. [Google Scholar] [CrossRef]
- van der Spiegel, M.; Noordam, M.; van der Fels-Klerx, H. Safety of novel protein sources (insects, microalgae, seaweed, duckweed, and rapeseed) and legislative aspects for their application in food and feed production. Compr. Rev. Food Sci. Food Saf. 2013, 12, 662–678. [Google Scholar] [CrossRef]
- van Huis, A. Edible insects are the future? Proc. Nutr. Soc. 2016, 75, 294–305. [Google Scholar] [CrossRef]
- Turck, D.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.; Kearney, J.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.; Naska, A.; Pelaez, C. Safety of dried yellow mealworm (Tenebrio molitor larva) as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2021, 19, 6343. [Google Scholar] [CrossRef]
- de Gier, S.; Verhoeckx, K. Insect (food) allergy and allergens. Mol. Immunol. 2018, 100, 82–106. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, J.C.; Cunha, L.M.; Sousa-Pinto, B.; Fonseca, J. Allergic risks of consuming edible insects: A systematic review. Mol. Nutr. Food Res. 2018, 62, 1700030. [Google Scholar] [CrossRef]
- Barre, A.; Pichereaux, C.; Velazquez, E.; Maudouit, A.; Simplicien, M.; Garnier, L.; Bienvenu, F.; Bienvenu, J.; Burlet-Schiltz, O.; Auriol, C.; et al. Insights into the allergenic potential of the edible yellow mealworm (Tenebrio molitor). Foods 2019, 8, 515. [Google Scholar] [CrossRef]
- Debode, F.; Marien, A.; Gérard, A.; Francis, F.; Fumière, O.; Berben, G. Development of real-time PCR tests for the detection of Tenebrio molitor in food and feed. Food Addit. Contam. Part A 2017, 34, 1421–1426. [Google Scholar] [CrossRef] [PubMed]
- Köppel, R.; Schum, R.; Habermacher, M.; Sester, C.; Piller, L.E.; Meissner, S.; Pietsch, K. Multiplex real-time PCR for the detection of insect DNA and determination of contents of Tenebrio molitor, Locusta migratoria and Achaeta domestica in food. Eur. Food Res. Technol. 2019, 245, 559–567. [Google Scholar] [CrossRef]
- Tramuta, C.; Gallina, S.; Bellio, A.; Bianchi, D.M.; Chiesa, F.; Rubiola, S.; Romano, A.; Decastelli, L. A set of multiplex polymerase chain reactions for genomic detection of nine edible insect species in foods. J. Insect Sci. 2018, 18, 3. [Google Scholar] [CrossRef]
- Barre, A.; Pichereaux, C.; Simplicien, M.; Burlet-Schiltz, O.; Benoist, H.; Rougé, P. A proteomic- and bioinformatic-based identification of specific allergens from edible insects: Probes for future detection as food ingredients. Foods 2021, 10, 280. [Google Scholar] [CrossRef]
- Francis, F.; Mazzucchelli, G.; Baiwir, D.; Debode, F.; Berben, G.; Caparros Megido, R. Proteomics based approach for edible insect fingerprinting in novel food: Differential efficiency according to selected model species. Food Control 2020, 112, 107135. [Google Scholar] [CrossRef]
- Fajardo, V.; Gonzalez, I.; Martin, I.; Rojas, M.; Hernandez, P.E.; Garcia, T.; Martin, R. Real-time PCR for detection and quantification of red deer (Cervus elaphus), fallow deer (Dama dama), and roe deer (Capreolus capreolus) in meat mixtures. Meat Sci. 2008, 79, 289–298. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed]
- ENGL. Definition of Minimum Performance Requirements for Analytical methods of GMO Testing; European Network of GMO Laboratories, Joint Research Center, EURL: Ispra, Italy, 2015; Available online: http://gmo-crl.jrc.ec.europa.eu/doc/MPR%20Report%20Application%2020_10_2015.pdf (accessed on 30 September 2022).
- Costa, J.; Fernandes, T.J.R.; Villa, C.; Oliveira, M.B.P.P.; Mafra, I. Advances in food allergen analysis. In Food Safety: Innovative Analytical Tools for Safety Assessment; Spizzirri, G., Cirillo, G., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2017; pp. 305–360. [Google Scholar] [CrossRef]
- Broekman, H.; Verhoeckx, K.C.; den Hartog Jager, C.F.; Kruizinga, A.G.; Pronk-Kleinjan, M.; Remington, B.C.; Bruijnzeel-Koomen, C.A.; Houben, G.F.; Knulst, A.C. Majority of shrimp-allergic patients are allergic to mealworm. J. Allergy Clin. Immunol. 2016, 137, 1261–1263. [Google Scholar] [CrossRef] [PubMed]
- Freye, H.B.; Esch, R.E.; Litwin, C.M.; Sorkin, L. Anaphylaxis to the ingestion and inhalation of Tenebrio molitor (mealworm) and Zophobas morio (superworm). Allergy Asthma Proc. 1996, 17, 215. [Google Scholar] [CrossRef]
- German Federal Institute for Risk Assessment (BfR); National Reference Laboratory for Animal Protein in Feed; NRL-AP; Garino, C.; Zagon, J.; Braeuning, A. Insects in food and feed—Allergenicity risk assessment and analytical detection. EFSA J. 2019, 17, e170907. [Google Scholar] [CrossRef]
- Garino, C.; Mielke, H.; Knüppel, S.; Selhorst, T.; Broll, H.; Braeuning, A. Quantitative allergenicity risk assessment of food products containing yellow mealworm (Tenebrio molitor). Food Chem. Toxicol. 2020, 142, 111460. [Google Scholar] [CrossRef]
- Costa, J.; Oliveira, M.B.P.; Mafra, I. Effect of thermal processing on the performance of the novel single-tube nested real-time PCR for the detection of walnut allergens in sponge cakes. Food Res. Int. 2013, 54, 1722–1729. [Google Scholar] [CrossRef]
- Villa, C.; Costa, J.; Mafra, I. Detection and quantification of milk ingredients as hidden allergens in meat products by a novel specific real-time PCR method. Biomolecules 2019, 9, 804. [Google Scholar] [CrossRef]
- Costa, J.; Villa, C.; Grazina, L.; Mafra, I. Single-tube nested real-time PCR versus normalised real-time PCR for the quantification of allergenic cashew nut in foods: Impact of thermal processing and matrix. Food Chem. 2022, 397, 133778. [Google Scholar] [CrossRef]
- Costa, J.; Amaral, J.S.; Grazina, L.; Oliveira, M.B.P.P.; Mafra, I. Matrix-normalised real-time PCR approach to quantify soybean as a potential food allergen as affected by thermal processing. Food Chem. 2017, 221, 1843–1850. [Google Scholar] [CrossRef]
- Jantzen da Silva Lucas, A.; Menegon de Oliveira, L.; da Rocha, M.; Prentice, C. Edible insects: An alternative of nutritional, functional and bioactive compounds. Food Chem. 2020, 311, 126022. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Target Region | Primers | Sequence (5′→3′) | Amplicon (bp) | Annealing Temperature (°C) | Accession No. (NCBI)/ Reference |
|---|---|---|---|---|---|
| Nuclear 18S rRNA | 18SEU-F 18SEU-R | TCTGCCCTATCAACTTTCGATGG TAATTTGCGCGCCTGCTG | 140 | 60 | [20] |
| NADH dehydrogenase subunit 2 | YMW_ND2-F YMW_ND2-R | ACGTCTTTTCTACAGAAGCATC GGGGTTAATGAATTCGTTTGTTA | 113 | 56 | KP994554.1/ |
| Cytochrome C oxidase subunit 1 | YMW_COX1-F YMW_COX1-R | CCCATGGAGGAGCATCTGTC TGCCCTGTGGTCGTATGTTGA | 117 | 60 | KP994554.1 |
| NADH dehydrogenase subunit 5 | YMW_ND5-F YMW_ND5-R | AATTGACCAGCCTCCCCATAAA CTACACTTAGTCAGCTTGGTCT | 180 | 56 | KP994554.1 |
| Tropomyosin mRNA | YMW_TM-F YMW_TM-R | ATGCTCAAGACAAGGCCGATGC TCCTTGTTGGCTTGTTCCAGGT | 160 | 59 | MK425158.1/ |
| Cytochrome b | YMW_CytB-F YMW_CytB-R YMW_CytB-P | CCTTACCTAGGAACAACAATTGT GATTGTTTGATCCTGTTTGGTGT FAM-TTGAGGAGGATTTGCTGTAGACAATGCA-BHQ1 | 160 | 59 | KP994554.1 |
| E Cadherin mRNA | YMW_CAD-F YMW_CAD-R | TGGTTGATCGGTGTTTCGGTA AATCCCGACTCTTGCGACC | 137 | 61 | DQ988044 |
| Parameter | Sausages | Biscuits | ||
|---|---|---|---|---|
| Raw | Autoclaved | Raw | Baked | |
| Correlation coefficient (R2) | 0.9937 | 0.9935 | 0.9967 | 0.9803 |
| Slope | −3.6851 | −3.2353 | −3.2503 | −3.4908 |
| PCR efficiency (%) | 86.8 | 103.7 | 103.1 | 93.4 |
| Relative LOD/LOQ (mg/kg) | 1.0 | 1.0 | 1.0 | 0.1 |
| Blind Mixtures | Actual T. molitor Content (mg/kg) | Estimated T. molitor Content (Mean ± SD, mg/kg) a | CV b (%) | Bias c (%) |
|---|---|---|---|---|
| Sausages | ||||
| Raw | ||||
| A | 40,000 | 47,249 ± 6034 | 15.1 | 18.1 |
| B | 20,000 | 25,912 ± 2009 | 10.0 | −22.8 |
| C | 4000 | 4914 ± 291.9 | 7.3 | 22.9 |
| D | 2000 | 2230 ± 216.4 | 10.8 | 11.5 |
| E | 400 | 490.8 ± 61.1 | 15.3 | 22.7 |
| F | 40 | 131.9 ± 33.3 | 83.2 | 229.8 |
| Autoclaved | ||||
| G | 40,000 | 46,907 ± 2666 | 6.7 | 17.3 |
| H | 20,000 | 21,445 ± 1238 | 6.2 | 7.2 |
| I | 4000 | 3003 ± 126.8 | 3.2 | −24.9 |
| J | 2000 | 2175 ± 237.3 | 11.9 | 8.8 |
| K | 400 | 330.4 ± 50.3 | 12.6 | −17.4 |
| L | 40 | 27.80 ± 4.04 | 10.1 | −30.5 |
| Biscuits | ||||
| Raw | ||||
| M | 40,000 | 39,620 ± 3674 | 9.2 | −1.0 |
| n | 4000 | 3146 ± 324.7 | 8.1 | −21.4 |
| O | 400 | 356.7 ± 29.2 | 7.3 | −10.8 |
| p | 40 | 31.62 ± 12.91 | 32.3 | −21.0 |
| Oven-baked | ||||
| Q | 40,000 | 45,322 ± 2359 | 5.2 | 13.3 |
| R | 4000 | 5623 ± 276.7 | 4.9 | 40.6 |
| S | 400 | 311.0 ± 23.3 | 7.5 | −22.3 |
| T | 40 | 33.68 ± 6.06 | 18.0 | −15.8 |
| Samples | Relevant Label Information | YMW_CytB-F/R (Cq ± SD) a | Estimated T. molitor Content (Mean ± SD) b | |
|---|---|---|---|---|
| mg/kg | % | |||
| Protein bars | ||||
| 1 | Contains dried yellow flour larvae (T. molitor larvae) | 22.89 ± 0.08 | 2482 ± 160 | 0.248 ± 0.016 |
| 2 | Contains dried yellow flour larvae (T. molitor larvae) | 21.76 ± 0.11 | 5231 ± 197 | 0.523 ± 0.020 |
| 3 | Contains dried yellow flour larvae (T. molitor larvae) | 23.40 ± 0.12 | 1769 ± 104 | 0.177 ± 0.010 |
| 4 | Contains dried yellow flour larvae (T. molitor larvae) | 19.45 ± 0.07 | 23,925 ± 1449 | 2.39 ± 0.14 |
| Chocolates | ||||
| 5 | Contains dehydrated T. molitor larvae (0.2%) | 26.54 ± 0.85 | 2270 ± 110 | 0.227 ± 0.011 |
| 6 | Contains dehydrated T. molitor larvae (0.2%) | 22.82 ± 0.44 | 2426 ± 510 | 0.243 ± 0.051 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villa, C.; Moura, M.B.M.V.; Teixeira, C.S.S.; Costa, J.; Mafra, I. Monitoring Yellow Mealworm (Tenebrio molitor) as a Potential Novel Allergenic Food: Effect of Food Processing and Matrix. Nutrients 2023, 15, 482. https://doi.org/10.3390/nu15030482
Villa C, Moura MBMV, Teixeira CSS, Costa J, Mafra I. Monitoring Yellow Mealworm (Tenebrio molitor) as a Potential Novel Allergenic Food: Effect of Food Processing and Matrix. Nutrients. 2023; 15(3):482. https://doi.org/10.3390/nu15030482
Chicago/Turabian StyleVilla, Caterina, Mónica B. M. V. Moura, Carla S. S. Teixeira, Joana Costa, and Isabel Mafra. 2023. "Monitoring Yellow Mealworm (Tenebrio molitor) as a Potential Novel Allergenic Food: Effect of Food Processing and Matrix" Nutrients 15, no. 3: 482. https://doi.org/10.3390/nu15030482
APA StyleVilla, C., Moura, M. B. M. V., Teixeira, C. S. S., Costa, J., & Mafra, I. (2023). Monitoring Yellow Mealworm (Tenebrio molitor) as a Potential Novel Allergenic Food: Effect of Food Processing and Matrix. Nutrients, 15(3), 482. https://doi.org/10.3390/nu15030482