Preventive Effect of Arctium lappa Polysaccharides on Acute Lung Injury through Anti-Inflammatory and Antioxidant Activities

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Extraction of ALP
2.3. The yield and Chemical Composition Analysis
2.4. FT-IR and UV–Vis Analysis
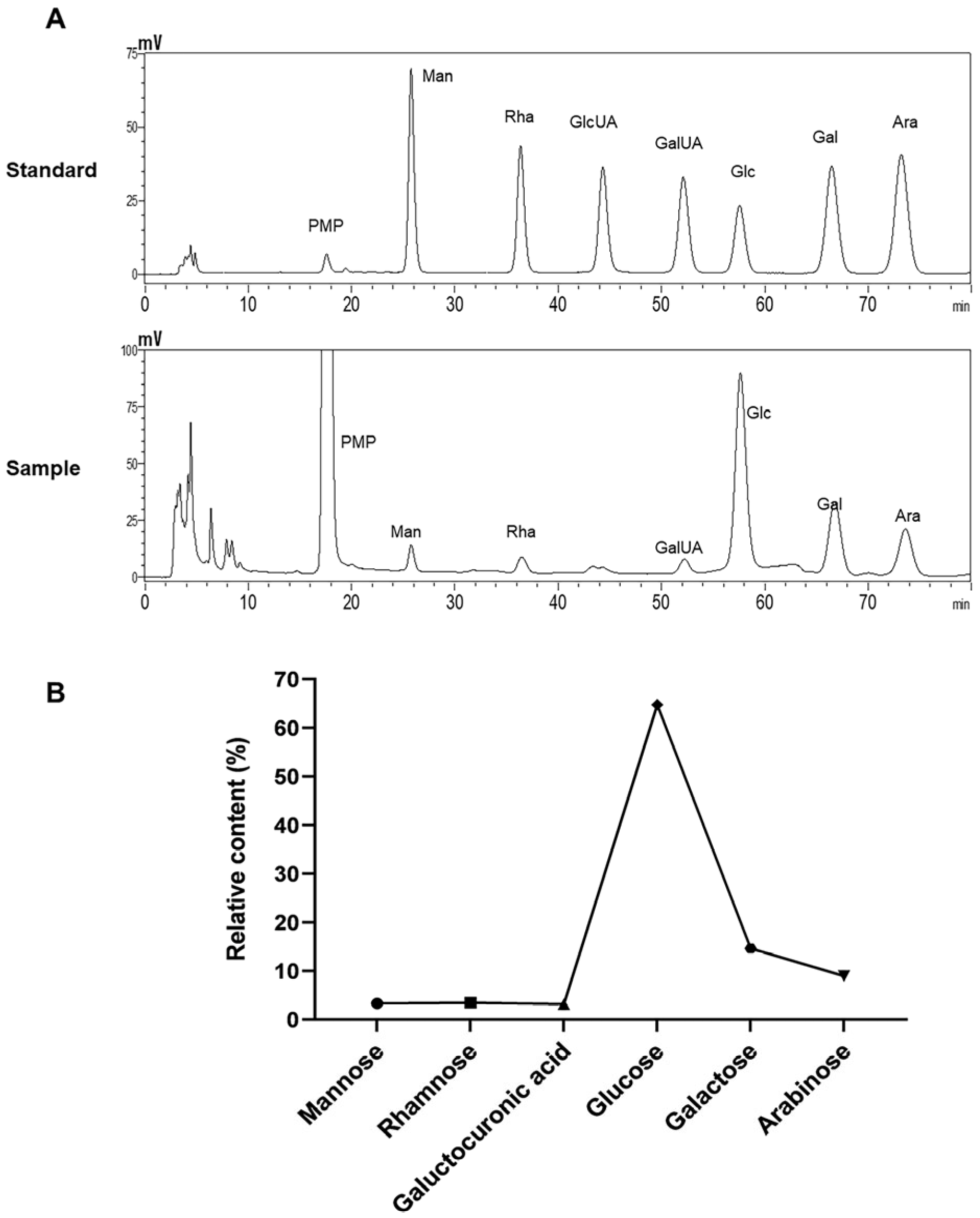
2.5. Molecular Weight and Monosaccharide Composition Analysis
2.6. Microstructural and Surface Morphological Analyses
2.7. Evaluation of Cell Viability
2.8. Animals and Treatments
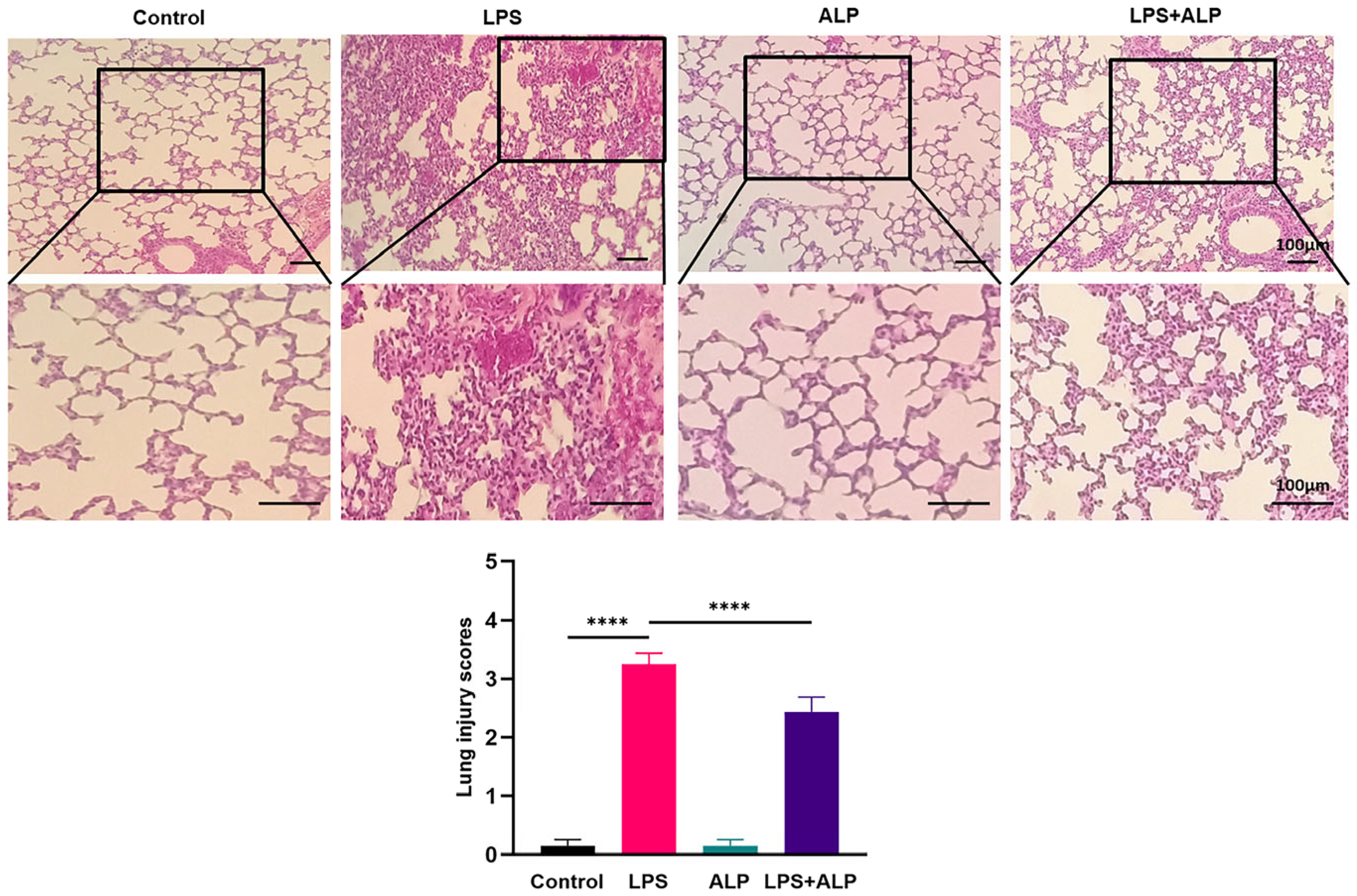
2.9. Histopathological Examination
2.10. The Determination of Oxidative Stress Indicators in Bronchoalveolar Lavage Fluid (BALF) and Serum
2.11. Evaluation of the Lung Wet/Dry Weight Ratio
2.12. Collection of BALF and Total Cell Count
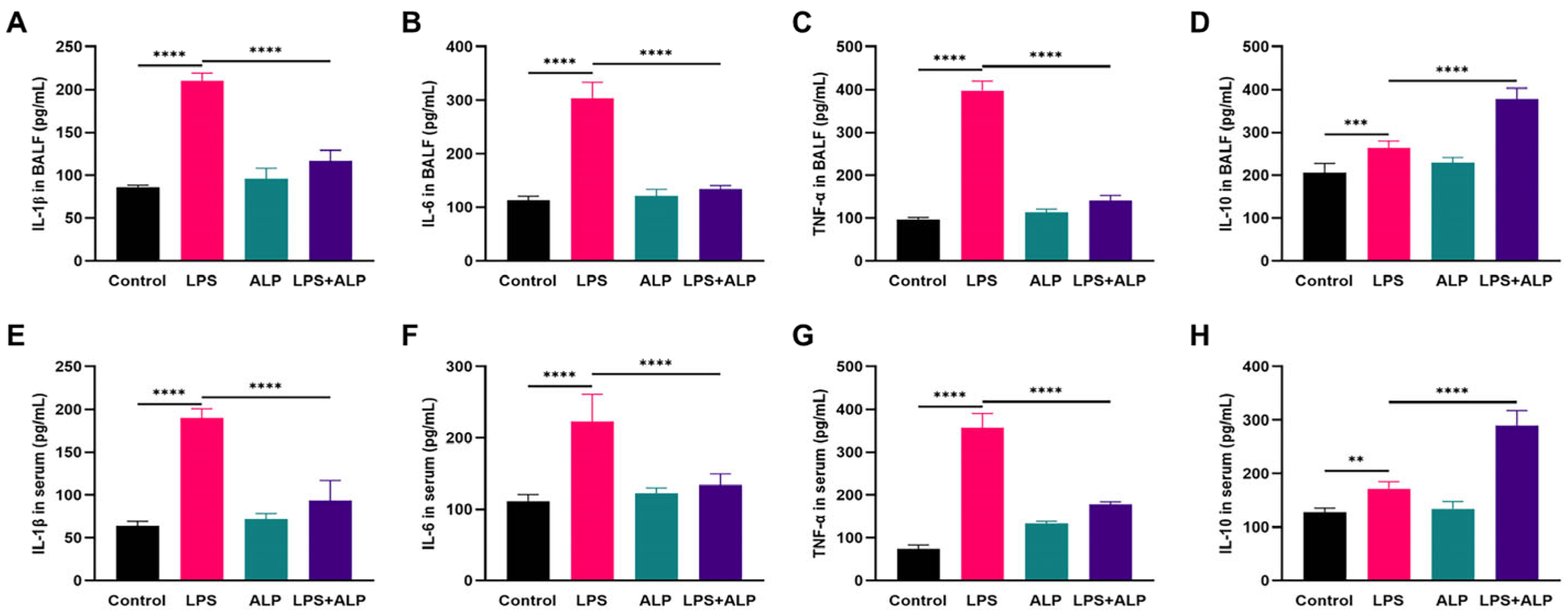
2.13. Measurement of Cytokine Levels in BALF and Serum
2.14. Immunohistochemical Examination
2.15. Statistical Analysis
3. Results
3.1. Chemical Composition and Structural Characterization Analysis of ALP
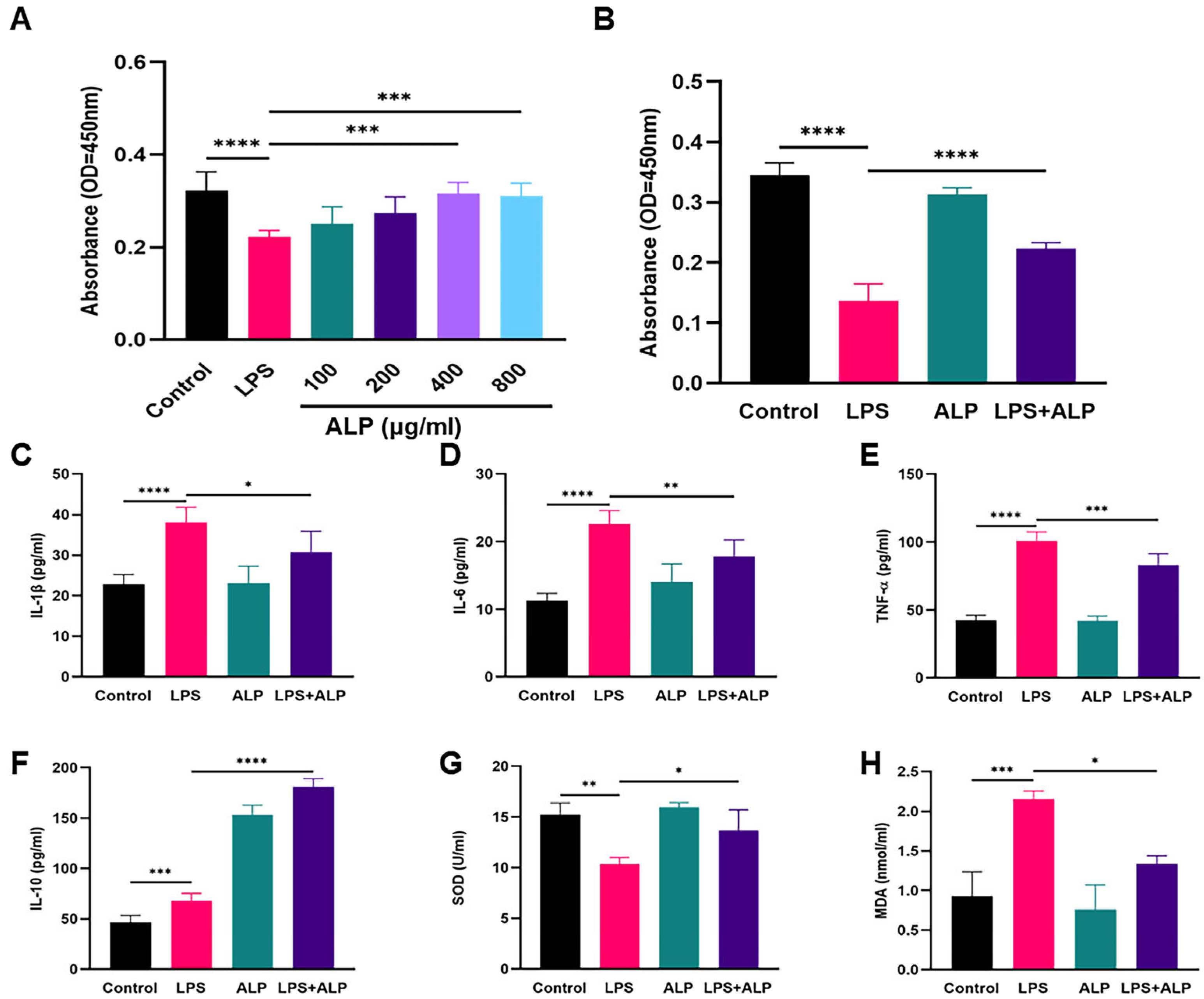
3.2. ALP Improved Cell Viability, Oxidative Stress, and Inflammation
3.3. ALP Improved the Pathological Morphology of LPS-Induced Lung Tissue in Mice
3.4. Effect of ALP on Inflammation in LPS-Induced ALI Mice
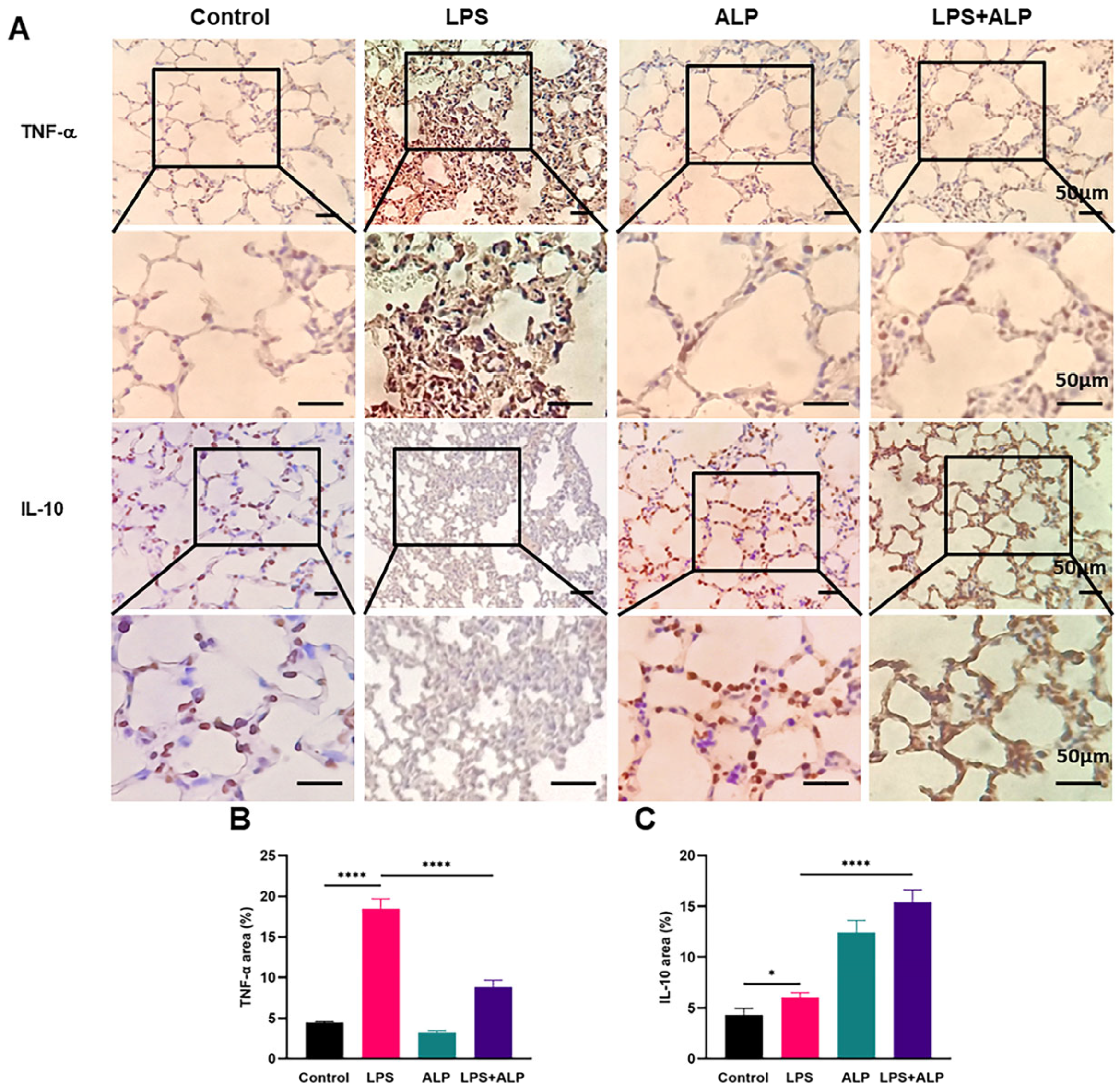
3.5. Effects of ALP on the Protein Expression of TNF-α and IL-10 in Mice
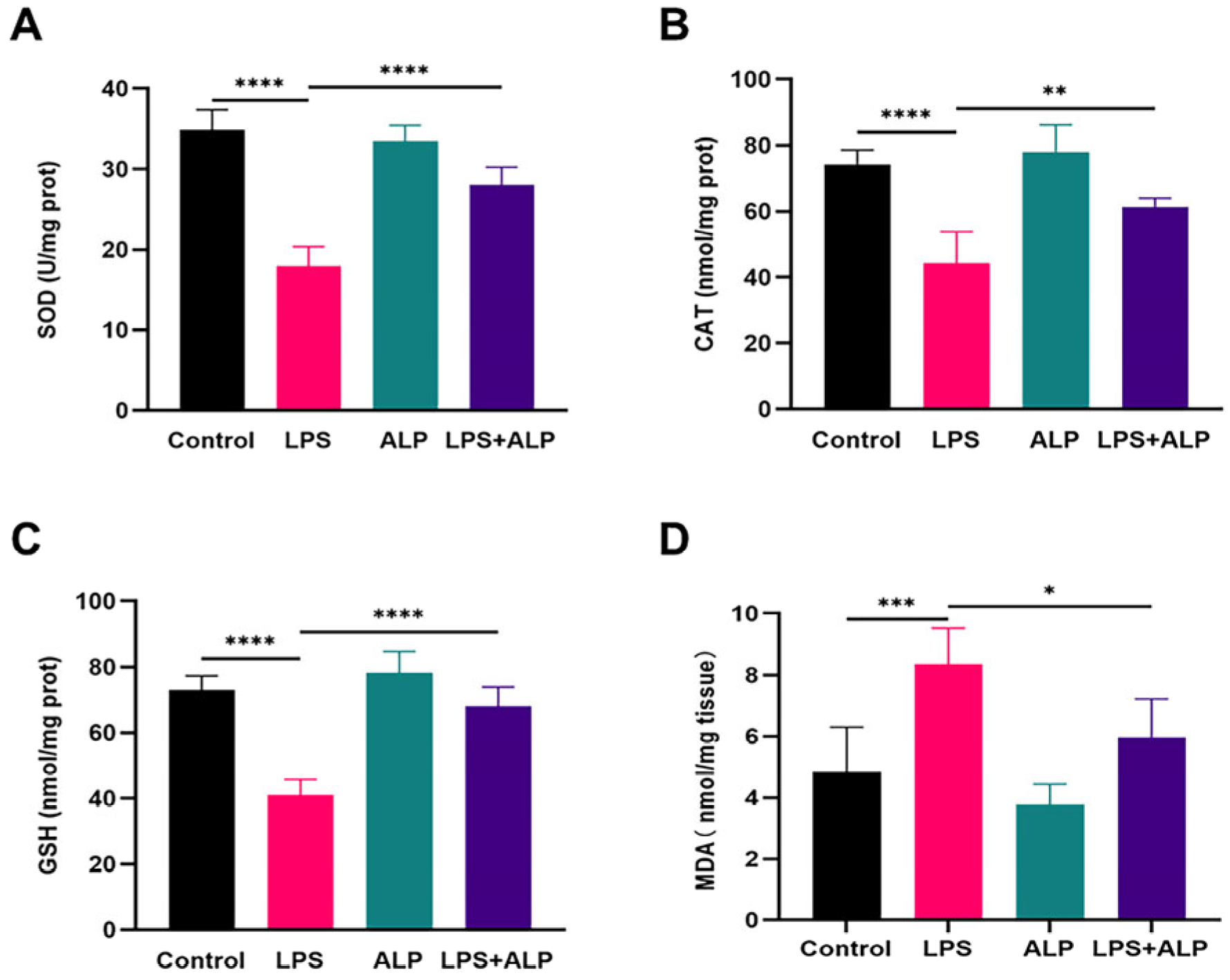
3.6. Effect of ALP on Levels of Oxidative Stress Biomarkers in LPS-Induced ALI Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yu, H.; Wu, W.; Lin, X.; Feng, Y. Polysaccharide-based nanomaterials for ocular drug delivery: A perspective. Front. Bioeng. Biotechnol. 2020, 8, 601246. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, J.H.; Wang, L.S.; Ding, J.; Zhao, M.W.; Liu, R. GlPP2C1 Silencing Increases the Content of Ganodermalingzhi Polysaccharide (GL-PS) and Enhances Slt2 Phosphorylation. J. Fungi 2022, 8, 949. [Google Scholar] [CrossRef]
- He, G.; Liu, Z.; Chen, H.; Wang, Y.; Huang, W.; Lu, X.; Tian, Y.; Liu, H. Effects of different boiling processes on chemical compositions of Lilii Bulbus soup. Front. Nutr. 2022, 9, 985105. [Google Scholar] [CrossRef]
- Wu, S.; Liu, H.; Li, S.; Sun, H.; He, X.; Huang, Y.; Long, H. Transcriptome analysis reveals possible immunomodulatory activity mechanism of Chlorella sp. exopolysaccharides on RAW264. 7 macrophages. Mar. Drugs 2021, 19, 217. [Google Scholar] [CrossRef]
- Wan, L.; Zhang, Q.; Luo, H.; Xu, Z.; Huang, S.; Yang, F.; Liu, Y.; Mahaman, Y.A.R.; Ke, D.; Wang, Q.; et al. Codonopsis pilosula polysaccharide attenuates Aβ toxicity and cognitive defects in APP/PS1 mice. Aging 2020, 12, 13422–13436. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Sheng, J.; He, X.; Sun, J.; Wei, Z.; Liu, G.; Li, C.; Lin, B.; Li, L. Novel Antioxidant and Hypoglycemic Water-Soluble Polysaccharides from Jasmine Tea. Foods 2021, 10, 2375. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Herrera-Balandrano, D.D.; Huang, W.; Chai, Z.; Beta, T.; Wang, J.; Feng, J.; Li, Y. Comparison of Nutritional and Nutraceutical Properties of Burdock Roots Cultivated in Fengxian and Peixian of China. Foods 2021, 10, 2095. [Google Scholar] [CrossRef]
- Tan, Y.J.; Ren, Y.S.; Gao, L.; Li, L.F.; Cui, L.J.; Li, B.; Li, X.; Yang, J.; Wang, M.Z.; Lv, Y.Y.; et al. 28-Day Oral Chronic Toxicity Study of Arctigenin in Rats. Front. Pharmacol. 2018, 9, 1077. [Google Scholar] [CrossRef] [PubMed]
- Yosri, N.; Alsharif, S.M.; Xiao, J.; Musharraf, S.G.; Zhao, C.; Saeed, A.; Gao, R.; Said, N.S.; Di Minno, A.; Daglia, M. Arctium lappa (Burdock): Insights from ethnopharmacology potential, chemical constituents, clinical studies, pharmacological utility and nanomedicine. Biomed. Pharmacother. 2023, 158, 114104. [Google Scholar] [CrossRef]
- Li, L.; Qiu, Z.; Dong, H.; Ma, C.; Qiao, Y.; Zheng, Z. Structural characterization and antioxidant activities of one neutral polysaccharide and three acid polysaccharides from the roots of Arctium lappa L.: A comparison. Int. J. Biol. Macromol. 2021, 182, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Wang, J.; Zhang, Z.; Xu, J.; Xie, Z.; Slavin, M.; Gao, X. In vitro and in vivo antioxidant activity of a fructan from the roots of Arctium lappa L. Int. J. Biol. Macromol. 2014, 65, 446–453. [Google Scholar] [CrossRef]
- Zhang, X.-J.; Liu, S.-F.; Lu, Y.; Wang, J.-Y.; Chen, K.-S. Immunomodulatory activity of a fructooligosaccharide isolated from burdock roots. RSC Adv. 2019, 9, 11092–11100. [Google Scholar] [CrossRef]
- Yu, M.-S.; Lai, S.-W.; Lin, K.-F.; Fang, J.-N.; Yuen, W.-H.; Chang, R.C.-C. Characterization of polysaccharides from the flowers of Nerium indicum and their neuroprotective effects. Int. J. Mol. Med. 2004, 14, 917–924. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Ma, Q.; Xu, X.; Feng, L.; Chen, Q.; Chen, Y.; Li, Z.; Liu, C.; Chen, K. Burdock fructooligosaccharide ameliorates the hypercholesterolemia and vascular inflammation in mice by regulating cholesterol homeostasis and anti-inflammatory properties. J. Funct. Foods 2023, 107, 105678. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, N.; Kan, J.; Zhang, X.; Wu, X.; Sun, R.; Tang, S.; Liu, J.; Qian, C.; Jin, C. Structural characterization of water-soluble polysaccharide from Arctium lappa and its effects on colitis mice. Carbohydr. Polym. 2019, 213, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Yin, H.; Xu, L.; Meng, X.; Kang, T. Chemical characterization of polysaccharides from Arctium lappa root and its hepatoprotective effects on mice. J. Funct. Foods 2023, 103, 105482. [Google Scholar] [CrossRef]
- Zhao, G.; Zhang, T.; Ma, X.; Jiang, K.; Wu, H.; Qiu, C.; Guo, M.; Deng, G. Oridonin attenuates the release of pro-inflammatory cytokines in lipopolysaccharide-induced RAW264.7 cells and acute lung injury. Oncotarget 2017, 8, 68153–68164. [Google Scholar] [CrossRef]
- Mohammed, A.; FK Alghetaa, H.; Miranda, K.; Wilson, K.; Singh, N.P.; Cai, G.; Putluri, N.; Nagarkatti, P.; Nagarkatti, M. Δ9-tetrahydrocannabinol prevents mortality from acute respiratory distress syndrome through the induction of apoptosis in immune cells, leading to cytokine storm suppression. Int. J. Mol. Sci. 2020, 21, 6244. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Li, X.; Li, T.; Nyunoya, T.; Chen, K.; Kitsios, G.D.; Nouraie, S.M.; Zhang, Y.; McVerry, B.J.; Lee, J.S. Endotoxin stabilizes protein arginine methyltransferase 4 (PRMT4) protein triggering death of lung epithelia. Cell Death Dis. 2021, 12, 828. [Google Scholar] [CrossRef]
- Son, E.; Cho, W.H.; Jang, J.H.; Kim, T.; Jeon, D.; Kim, Y.S.; Yeo, H.J. Neutrophil gelatinase-associated lipocalin as a prognostic biomarker of severe acute respiratory distress syndrome. Sci. Rep. 2022, 12, 7909. [Google Scholar] [CrossRef]
- Lei, J.; Wei, Y.; Song, P.; Li, Y.; Zhang, T.; Feng, Q.; Xu, G. Cordycepin inhibits LPS-induced acute lung injury by inhibiting inflammation and oxidative stress. Eur. J. Pharmacol. 2018, 818, 110–114. [Google Scholar] [CrossRef]
- Cheng, L.; Chai, Y.M.; Zhao, G.; Wen, G.; Han, P. Glucocorticoid receptorβ isoform exhibits a disproportionate increase over the α isoform in the lungs of a polytrauma rat model. Int. J. Clin. Exp. Pathol. 2018, 11, 3046–3051. [Google Scholar]
- Altintas, N.D.; Atilla, P.; Iskit, A.B.; Topeli, A. Long-term simvastatin attenuates lung injury and oxidative stress in murine acute lung injury models induced by oleic acid and endotoxin. Respir. Care 2011, 56, 1156–1163. [Google Scholar] [CrossRef]
- Li, Y.-H.; Xu, Y.; Wu, H.-M.; Yang, J.; Yang, L.-H.; Yue-Meng, W. Umbilical cord-derived mesenchymal stem cell transplantation in hepatitis B virus related acute-on-chronic liver failure treated with plasma exchange and entecavir: A 24-month prospective study. Stem Cell Rev. Rep. 2016, 12, 645–653. [Google Scholar] [CrossRef]
- Dai, J.; Wu, Y.; Chen, S.-w.; Zhu, S.; Yin, H.-p.; Wang, M.; Tang, J. Sugar compositional determination of polysaccharides from Dunaliella salina by modified RP-HPLC method of precolumn derivatization with 1-phenyl-3-methyl-5-pyrazolone. Carbohydr. Polym. 2010, 82, 629–635. [Google Scholar] [CrossRef]
- Wang, N.; Geng, C.; Sun, H.; Wang, X.; Li, F.; Liu, X. Hesperetin ameliorates lipopolysaccharide-induced acute lung injury in mice through regulating the TLR4–MyD88–NF-κB signaling pathway. Arch. Pharmacal Res. 2019, 42, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ye, Y.; Hu, X.; Wang, J. Structural characterization and anti-inflammatory activity of a polysaccharide from the lignified okra. Carbohydr. Polym. 2021, 265, 118081. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Qian, L.; Wang, B.; Zhang, Z.; Liu, H.; Zhang, Y.; Liu, J. Synergistic hypoglycemic effects of pumpkin polysaccharides and puerarin on type II diabetes mellitus mice. Molecules 2019, 24, 955. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Pan, X.; Ran, S.; Wang, K. Purification, structural elucidation and anti-inflammatory activity in vitro of polysaccharides from Smilax china L. Int. J. Biol. Macromol. 2019, 139, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, N.; Kan, J.; Sun, R.; Tang, S.; Wang, Z.; Chen, M.; Liu, J.; Jin, C. Anti-inflammatory activity of alkali-soluble polysaccharides from Arctium lappa L. and its effect on gut microbiota of mice with inflammation. Int. J. Biol. Macromol. 2020, 154, 773–787. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Park, E.-J.; Kim, J.-Y.; Choi, J.; Lee, H.-J. Anti-inflammatory effects of fermented lotus root and linoleic acid in lipopolysaccharide-induced RAW 264.7 cells. Life 2020, 10, 293. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Chen, F.; Cheng, N.T.; Tang, Z.; Wang, X.G.; Xu, M. MicroRNA-1224-5p Aggravates Sepsis-Related Acute Lung Injury in Mice. Oxidative Med. Cell. Longev. 2022, 2022, 9493710. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Lv, B.; Zheng, J.; Lu, C.; Liu, J.; Lei, Y.; Yang, M.; Wang, Y.; Li, Z.; Yang, Y. RBC-hitchhiking chitosan nanoparticles loading methylprednisolone for lung-targeting delivery. J. Control. Release 2022, 341, 702–715. [Google Scholar] [CrossRef] [PubMed]
- Cain, D.W.; Cidlowski, J.A. After 62 years of regulating immunity, dexamethasone meets COVID-19. Nat. Rev. Immunol. 2020, 20, 587–588. [Google Scholar] [CrossRef]
- Li, W.; Cai, Z.-N.; Mehmood, S.; Wang, Y.; Pan, W.-J.; Zhang, W.-N.; Lu, Y.-M.; Chen, Y. Polysaccharide FMP-1 from Morchella esculenta attenuates cellular oxidative damage in human alveolar epithelial A549 cells through PI3K/AKT/Nrf2/HO-1 pathway. Int. J. Biol. Macromol. 2018, 120, 865–875. [Google Scholar] [CrossRef]
- Gong, Z.; Zhang, S.; Gu, B.; Cao, J.; Mao, W.; Yao, Y.; Zhao, J.; Ren, P.; Zhang, K.; Liu, B. Codonopsis pilosula polysaccharides attenuate Escherichia coli-induced acute lung injury in mice. Food Funct. 2022, 13, 7999–8011. [Google Scholar] [CrossRef]
- Zhang, Y.; Cui, Y.; Feng, Y.; Jiao, F.; Jia, L. Lentinus edodes Polysaccharides Alleviate Acute Lung Injury by Inhibiting Oxidative Stress and Inflammation. Molecules 2022, 27, 7328. [Google Scholar] [CrossRef]
- Swinarew, A.S.; Stanula, A.J.; Gabor, J.; Raif, P.; Paluch, J.; Karpiński, J.; Kubik, K.; Okła, H.; Ostrowski, A.; Tkacz, E.; et al. The influence of chlorine in indoor swimming pools on the composition of breathing phase of professional swimmers. Respir. Res. 2020, 21, 88. [Google Scholar] [CrossRef]
- Wu, Z.; Chen, L.; Wang, Q.; Govindasamy, C.; Subramaniyan Sivakumar, A.; Chen, X. Betanin ameliorates Lipopolysaccharide-induced acute lung injury in mice via inhibition of inflammatory response and oxidative stress. Arab. J. Chem. 2023, 16, 104763. [Google Scholar] [CrossRef]
- Chen, C.; Wang, J.; Chen, J.; Zhou, L.; Wang, H.; Chen, J.; Xu, Z.; Zhu, S.; Liu, W.; Yu, R. Morusin alleviates mycoplasma pneumonia via the inhibition of Wnt/β-catenin and NF-κB signaling. Biosci. Rep. 2019, 39, BSR20190190. [Google Scholar] [CrossRef]
- Lai, W.-Y.; Wang, J.-W.; Huang, B.-T.; Lin, E.P.-Y.; Yang, P.-C. A novel TNF-α-targeting aptamer for TNF-α-mediated acute lung injury and acute liver failure. Theranostics 2019, 9, 1741. [Google Scholar] [CrossRef] [PubMed]
- Iyer, S.S.; Cheng, G. Role of interleukin 10 transcriptional regulation in inflammation and autoimmune disease. Crit. Rev. Immunol. 2012, 32, 23–63. [Google Scholar] [CrossRef] [PubMed]
- Sanjeewa, K.K.A.; Fernando, I.P.S.; Kim, E.-A.; Ahn, G.; Jee, Y.; Jeon, Y.-J. Anti-inflammatory activity of a sulfated polysaccharide isolated from an enzymatic digest of brown seaweed Sargassum horneri in RAW 264.7 cells. Nutr. Res. Pract. 2017, 11, 3–10. [Google Scholar] [CrossRef]
- Rogers, L.K.; Cismowski, M.J. Oxidative stress in the lung–the essential paradox. Curr. Opin. Toxicol. 2018, 7, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Han, S.Y.; Hwang, K.; Kim, D.; Kim, E.M.; Hossain, M.A.; Kim, J.H.; Cho, J.Y. Antioxidant and Cytoprotective Effects of (-)-Epigallocatechin-3-(3″-O-methyl) Gallate. Int. J. Mol. Sci. 2019, 20, 3993. [Google Scholar] [CrossRef]
- Arioz, B.I.; Tastan, B.; Tarakcioglu, E.; Tufekci, K.U.; Olcum, M.; Ersoy, N.; Bagriyanik, A.; Genc, K.; Genc, S. Melatonin Attenuates LPS-Induced Acute Depressive-Like Behaviors and Microglial NLRP3 Inflammasome Activation Through the SIRT1/Nrf2 Pathway. Front. Immunol. 2019, 10, 1511. [Google Scholar] [CrossRef] [PubMed]
- Xiang, L.; Zhang, Q.; Chi, C.; Wu, G.; Lin, Z.; Li, J.; Gu, Q.; Chen, G. Curcumin analog A13 alleviates oxidative stress by activating Nrf2/ARE pathway and ameliorates fibrosis in the myocardium of high-fat-diet and streptozotocin-induced diabetic rats. Diabetol. Metab. Syndr. 2020, 12, 1. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, J.; Li, B.; He, C.; Chen, Y.; Tian, S. Influence of Oxidative Stress on Biocontrol Activity of Cryptococcus laurentii against Blue Mold on Peach Fruit. Front. Microbiol. 2017, 8, 151. [Google Scholar] [CrossRef]
- Liang, Q.; Zhao, Q.; Hao, X.; Wang, J.; Ma, C.; Xi, X.; Kang, W. The Effect of Flammulina velutipes Polysaccharide on Immunization Analyzed by Intestinal Flora and Proteomics. Front. Nutr. 2022, 9, 841230. [Google Scholar] [CrossRef]
- Wall, S.B.; Wood, R.; Dunigan, K.; Li, Q.; Li, R.; Rogers, L.K.; Tipple, T.E. Thioredoxin reductase-1 inhibition augments endogenous glutathione-dependent antioxidant responses in experimental bronchopulmonary dysplasia. Oxidative Med. Cell. Longev. 2019, 2019, 7945983. [Google Scholar] [CrossRef]
- Zheng, J.; Zhang, X.; Herrera-Balandrano, D.D.; Wang, J.; Chai, Z.; Beta, T.; Huang, W.; Li, Y. Extraction Optimization of Arctium lappa L. Polysaccharides by Box-Behnken Response Surface Design and Their Antioxidant Capacity. Starch Stärke 2022, 74, 2100305. [Google Scholar] [CrossRef]
- Jiang, Y.-Y.; Yu, J.; Li, Y.-B.; Wang, L.; Hu, L.; Zhang, L.; Zhou, Y.-H. Extraction and antioxidant activities of polysaccharides from roots of Arctium lappa L. Int. J. Biol. Macromol. 2019, 123, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Weng, J.; Li, C.; Feng, Y.; Xie, M.; Wang, X.; Chang, Q.; Li, M.; Chung, K.F.; Adcock, I.M. Attenuation of PM2.5-induced alveolar epithelial cells and lung injury through regulation of mitochondrial fission and fusion. Part. Fibre Toxicol. 2023, 20, 28. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, N.; Wei, J.; Gong, X.; Tang, X.; Zhang, X.; Xiang, W.; Liu, S.; Luo, C.; Wang, X. Preventive Effect of Arctium lappa Polysaccharides on Acute Lung Injury through Anti-Inflammatory and Antioxidant Activities. Nutrients 2023, 15, 4946. https://doi.org/10.3390/nu15234946
Lu N, Wei J, Gong X, Tang X, Zhang X, Xiang W, Liu S, Luo C, Wang X. Preventive Effect of Arctium lappa Polysaccharides on Acute Lung Injury through Anti-Inflammatory and Antioxidant Activities. Nutrients. 2023; 15(23):4946. https://doi.org/10.3390/nu15234946
Chicago/Turabian StyleLu, Naiyan, Jiayi Wei, Xuelei Gong, Xue Tang, Xuan Zhang, Wen Xiang, Samuel Liu, Cherry Luo, and Xun Wang. 2023. "Preventive Effect of Arctium lappa Polysaccharides on Acute Lung Injury through Anti-Inflammatory and Antioxidant Activities" Nutrients 15, no. 23: 4946. https://doi.org/10.3390/nu15234946
APA StyleLu, N., Wei, J., Gong, X., Tang, X., Zhang, X., Xiang, W., Liu, S., Luo, C., & Wang, X. (2023). Preventive Effect of Arctium lappa Polysaccharides on Acute Lung Injury through Anti-Inflammatory and Antioxidant Activities. Nutrients, 15(23), 4946. https://doi.org/10.3390/nu15234946