
Dietary Polyphenols—Natural Bioactive Compounds with Potential for Preventing and Treating Some Allergic Conditions

Abstract
:1. Introduction
2. Characterization of Polyphenols
3. Evidence from Epidemiological Studies
4. Potential Mechanisms of Action in the Prevention or Treatment of Allergic Diseases
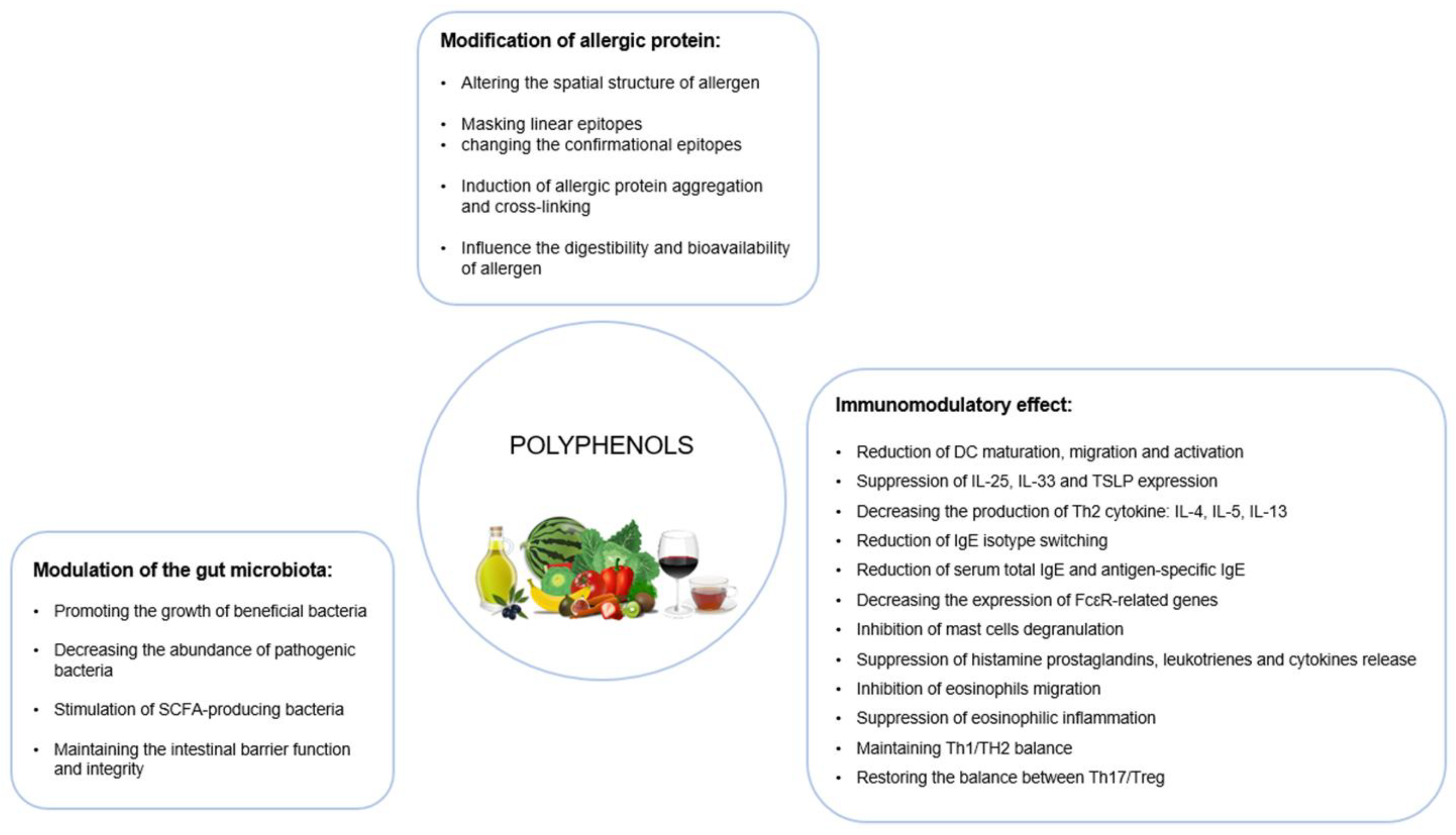
4.1. Modification of Allergic Protein
4.2. Immunomodulatory Effects
4.2.1. Sensitization Phase
4.2.2. Effector Phase
4.3. Modulation of the Gut Microflora
5. Polyphenols in the Prevention and Treatment of Food Allergy
6. Polyphenols in the Prevention and Treatment of Respiratory Allergy
6.1. Allergic Rhinitis
6.2. Asthma
7. Limitations and Future Challenges
8. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Christiansen, E.S.; Kjaer, H.F.; Eller, E.; Bindslev-Jensen, C.; Høst, A.; Mortz, C.G.; Halken, S. The Prevalence of Atopic Diseases and the Patterns of Sensitization in Adolescence. Pediatr. Allergy Immunol. 2016, 27, 847–853. [Google Scholar] [CrossRef] [PubMed]
- Asher, M.I.; García-Marcos, L.; Pearce, N.E.; Strachan, D.P. Trends in worldwide asthma prevalence. Eur. Respir. J. 2020, 56, 2002094. [Google Scholar] [CrossRef] [PubMed]
- Bousquet, J.; Anto, J.M.; Bachert, C.; Baiardini, I.; Bosnic-Anticevich, S.; Canonica, C.W.; Melén, E.; Palomares, O.; Scadding, G.K.; Togias, A.; et al. Allergic rhinitis. Nat. Rev. Dis. Primers 2020, 6, 95. [Google Scholar] [CrossRef] [PubMed]
- Weidinger, S.; Novak, N. Atopic dermatitis. Lancet 2016, 387, 1109–1122. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.B.; Mims, J.W.; Clinger, J.D. The Burden of Asthma and Allergic Rhinitis: Epidemiology and Health Care Costs. Otolaryngol. Clin. N. Am. 2023, in press. [Google Scholar] [CrossRef]
- Wise, S.K.; Damask, C.; Roland, L.T.; Ebert, C.; Levy, J.M.; Lin, S.; Luong, A.; Rodriguez, K.; Sedaghat, A.R.; Toskala, E.; et al. International consensus statement on allergy and rhinology: Allergic rhinitis–2023. Int. Forum Allergy Rhinol. 2023, 13, 293–859. [Google Scholar] [CrossRef]
- Yasaratne, D.; Idrose, N.S.; Dharmage, S.C. Asthma in developing countries in the Asia-Pacific Region (APR). Respirology 2023, 28, 992–1004. [Google Scholar] [CrossRef]
- Warren, C.; Nimmagadda, S.R.; Gupta, R.; Levin, M. The epidemiology of food allergy in adults. Ann. Allergy Asthma Immunol. 2023, 130, 276–287. [Google Scholar] [CrossRef]
- De, A.; Karekar, S.; Adhav, C. Current Burden of Atopic Dermatitis in India: A Systematic Literature Review. Indian J. Dermatol. 2023, 68, 487. [Google Scholar] [CrossRef]
- Gonzalez-Uribe, V.; Vidaurri-de la Cruz, H.; Gomez-Nuñez, A.; Leyva-Calderon, J.A.; Mojica-Gonzalez, Z.S. Comorbidities & burden of disease in atopic dermatitis. Asian Pac. J. Allergy Immunol. 2023, 41, 97–105. [Google Scholar] [CrossRef]
- Mahesh, P.A.; Kaleem Ullah, M.; Parthasarathi, A. Allergic sensitization to foods in India and other Low-Middle-income countries. Clin. Exp. Allergy 2023, 53, 739–750. [Google Scholar] [CrossRef] [PubMed]
- Spolidoro, G.C.I.; Amera, Y.T.; Ali, M.M.; Nyassi, S.; Lisik, D.; Ioannidou, A.; Rovner, G.; Khaleva, E.; Venter, C.; van Ree, R.; et al. Frequency of food allergy in Europe: An updated systematic review and meta-analysis. Allergy 2023, 78, 351–368. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, N.G.; Miligkos, M.; Xepapadaki, P. A Current Perspective of Allergic Asthma: From Mechanisms to Management. Handb. Exp. Pharmacol. 2022, 268, 69–93. [Google Scholar] [CrossRef]
- Julia, V.; Macia, L.; Dombrowicz, D. The impact of diet on asthma and allergic diseases. Nat. Rev. Immunol. 2015, 15, 308–322. [Google Scholar] [CrossRef]
- Lin, Y.P.; Kao, Y.C.; Pan, W.H.; Yang, Y.H.; Chen, Y.C.; Lee, Y.L. Associations between respiratory diseases and dietary patterns derived by factors analysis and reduced rank regression. Ann. Nutr. Metab. 2016, 68, 306–314. [Google Scholar] [CrossRef]
- Zhang, P. The Role of Diet and Nutrition in Allergic Diseases. Nutrients 2023, 15, 3683. [Google Scholar] [CrossRef]
- Rennie, G.H.; Zhao, J.; Camus-Ela, M.; Shi, J.; Jiang, L.; Zhang, L.; Wang, J.; Raghavan, V. Influence of Lifestyle and Dietary Habits on the Prevalence of Food Allergies: A Scoping Review. Foods 2023, 12, 3290. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yin, P.; Yu, L.; Tian, F.; Chen, W.; Zhai, Q. Effects of Early Diet on the Prevalence of Allergic Disease in Children: A Systematic Review and Meta-Analysis. Adv. Nutr. 2023, 100128. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhang, C.; Zhang, Y.; Liu, Y.; Wang, J.; Gao, Z.; Sun, J.; Li, Q.; Sun, J.; Cui, X.; et al. Early-life risk factors for food allergy: Dietary and environmental factors revisited. Compr. Rev. Food Sci. Food Saf. 2023, 22, 4355–4377. [Google Scholar] [CrossRef]
- Burney, P.G. The Causes of Asthma–Does Salt Potentiate Bronchial Activity? Discussion Paper. J. R. Soc. Med. 1987, 80, 364–367. [Google Scholar] [CrossRef]
- Garcia-Larsen, V.; Del Giacco, S.R.; Moreira, A.; Bonini, M.; Charles, D.; Reeves, T.; Carlsen, K.-H.; Haahtela, T.; Bonini, S.; Fonseca, J.; et al. Asthma and Dietary Intake: An Overview of Systematic Reviews. Allergy 2016, 71, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Brustad, N.; Bønnelykke, K.; Chawes, B. Dietary prevention strategies for childhood asthma. Pediatr. Allergy Immunol. 2023, 34, e13984. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; He, M.; Yu, Q.; Xiao, F.; Zhang, Y.; Liang, C. The Effects of a Healthy Diet on Asthma and Wheezing in Children and Adolescents: A Systematic Review and Meta-Analysis. J. Asthma Allergy 2023, 16, 1007–1024. [Google Scholar] [CrossRef] [PubMed]
- Panagiotou, E.; Andreou, E.; Nicolaou, S.A. The Effect of Dietary Components of the Mediterranean Diet on Food Allergies: A Systematic Review. Nutrients 2023, 15, 3295. [Google Scholar] [CrossRef] [PubMed]
- Venter, C. Immunonutrition: Diet Diversity, Gut Microbiome and Prevention of Allergic Diseases. Allergy Asthma Immunol. Res. 2023, 15, 545–561. [Google Scholar] [CrossRef] [PubMed]
- Vlieg-Boerstra, B.; Groetch, M.; Vassilopoulou, E.; Meyer, R.; Laitinen, K.; Swain, A.; Durban, R.; Benjamin, O.; Bottse, R.; Grimshaw, K.; et al. The immune-supportive diet in allergy management: A narrative review and proposal. Allergy 2023, 78, 1441–1458. [Google Scholar] [CrossRef]
- Phillips, C.M.; Chen, L.W.; Heude, B.; Bernard, J.Y.; Harvey, N.C.; Duijts, L.; Mensink-Bout, S.M.; Polanska, K.; Mancano, G.; Suderman, M.; et al. Dietary inflammatory index and non-communicable disease risk: A narrative review. Nutrients 2019, 11, 1873. [Google Scholar] [CrossRef]
- Rana, A.; Samtiya, M.; Dhewa, T.; Mishra, V.; Aluko, R.E. Health benefits of polyphenols: A concise review. J. Food Biochem. 2022, 46, e14264. [Google Scholar] [CrossRef]
- Eseberri, I.; Trepiana, J.; Léniz, A.; Gómez-García, I.; Carr-Ugarte, H.; González, M.; Portillo, M.P. Variability in the Beneficial Effects of Phenolic Compounds: A Review. Nutrients 2022, 14, 1925. [Google Scholar] [CrossRef]
- Rakha, A.; Umar, N.; Rabail, R.; Butt, M.S.; Kieliszek, M.; Hassoun, A.; Aadil, R.M. Anti-inflammatory and anti-allergic potential of dietary flavonoids: A review. Biomed. Pharmacother. 2022, 156, 113945. [Google Scholar] [CrossRef]
- Wu, T.; Li, Z.; Wu, Y.; Yang, X.; Li, L.; Chen, S.; Qi, B.; Wang, Y.; Li, C.; Zhao, Y. Exploring plant polyphenols as anti-allergic functional products to manage the growing incidence of food allergy. Front. Nutr. 2023, 10, 1102225. [Google Scholar] [CrossRef] [PubMed]
- Zeng, B.; Jiang, T.; Xiong, W.; Che, H.; Sun, S. Protective properties of polyphenols in food allergy: A review. Allergy 2023, 78, 1654–1656. [Google Scholar] [CrossRef] [PubMed]
- Kumari, M.; Siddiqui, M.A.; Gupta, A. Recent Advancement and Novel Application of Natural Polyphenols for the Treatment of Allergy Asthma: From Phytochemistry to Biological Implications. Crit. Rev. Immunol. 2023, 43, 29–41. [Google Scholar] [CrossRef]
- Belščak-Cvitanović, A.; Durgo, K.; Hudek, A.; Bačun-Družina, V.; Komes, D. Overview of polyphenols and their properties. Polyphenols 2018, 3–44. [Google Scholar] [CrossRef]
- de Araújo, F.F.; de Paulo Farias, D.; Neri-Numa, I.A.; Pastore, G.M. Polyphenols and their applications: An approach in food chemistry and innovation potential. Food Chem. 2020, 338, 127535. [Google Scholar] [CrossRef] [PubMed]
- Vuolo, M.M.; Lima, V.S.; Maróstica Junior, M.R. Phenolic Compounds: Structure, Classification, and Antioxidant Power; Elsevier Inc.: Amsterdam, The Netherlands, 2018. [Google Scholar] [CrossRef]
- Li, X.; Wang, T.; Zhou, B.; Gao, W.; Cao, J.; Huang, L. Chemical composition and antioxidant and anti-inflammatory potential of peels and flesh from 10 different pear varieties (Pyrus spp.). Food Chem. 2014, 152, 531–538. [Google Scholar] [CrossRef]
- Calderón-Oliver, M.; Ponce-Alquicira, E. Fruits: A Source of Polyphenols and Health Benefits; Academic Press: Cambridge, MA, USA, 2018. [Google Scholar] [CrossRef]
- Karak, P. Biological activities of flavonoids: An overview. Int. J. Pharm. Sci. Res. 2019, 10, 1567–1574. [Google Scholar] [CrossRef]
- Barreca, D.; Trombetta, D.; Smeriglio, A.; Mandalari, G.; Romeo, O.; Felice, M.R.; Gattuso, G.; Nabavi, S.M. Food flavonols: Nutraceuticals with complex health benefits and functionalities. Trends Food Sci. Technol. 2021, 117, 194–204. [Google Scholar] [CrossRef]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.E.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (poly)phenolics in human health: Structures, bioavailability, and evidence of protective effects against chronic diseases. Antioxid. Redox Signal. 2013, 18, 1818–1892. [Google Scholar] [CrossRef]
- Durazzo, A.; Lucarini, M.; Souto, E.B.; Cicala, C.; Caiazzo, E.; Izzo, A.A.; Novellino, E.; Santini, A. Polyphenols: A concise overview on the chemistry, occurrence, and human health. Phytother. Res. 2019, 33, 2221–2243. [Google Scholar] [CrossRef]
- Lucci, P.; Saurina, J.; Núñez, O. Trends in LC-MS and LC-HRMS analysis and characterization of polyphenols in food. TrAC–Trends Anal. Chem. 2017, 88, 1–24. [Google Scholar] [CrossRef]
- Mrduljaš, N.; Kreši´c, G.; Biluši´c, T. Polyphenols: Food sources and health benefits. In Functional Food-Improve Health through Adequate Food; IntechOpen: London, UK, 2017; pp. 23–41. [Google Scholar] [CrossRef]
- Suganthy, N.; Devi, K.P.; Nabavi, S.F.; Braidy, N.; Nabavi, S.M. Bioactive Effects of Quercetin in the Central Nervous System: Focusing on the Mechanisms of Actions. Biomed. Pharmacother. 2016, 84, 892–908. [Google Scholar] [CrossRef] [PubMed]
- Russo, M.; Bonaccorsi, I.; Inferrera, V.; Dugo, P.; Mondello, L. Underestimated sources of flavonoids, limonoids and dietary fiber: Availability in orange’s by-products. J. Funct. Foods 2015, 12, 150–157. [Google Scholar] [CrossRef]
- Popa, D.S.; Rusu, M.E. Isoflavones: Vegetable sources, biological activity, and analytical methods for their assessment. In Superfood and Functional Food—The Development of Superfoods and Their Roles as Medicine; IntechOpen: London, UK, 2017; pp. 133–153. [Google Scholar] [CrossRef]
- Kelebek, H.; Selli, S.; Kadiroglu, P.; Kola, O.; Kesen, S.; Uçar, B.; Çetiner, B. Bioactive compounds and antioxidant potential in tomato pastes as affected by hot and cold break process. Food Chem. 2017, 220, 31–41. [Google Scholar] [CrossRef]
- Bubalo, M.C.; Curko, N.; Tomašević, M.; Ganić, K.K.; Redovniković, I.R. Green extraction of grape skin phenolics by using deep eutectic solvents. Food Chem. 2016, 200, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Godos, J.; Marventano, S.; Mistretta, A.; Galvano, F.; Grosso, G. Dietary sources of polyphenols in the Mediterranean healthy eating, aging and lifestyle (MEAL) study cohort. Int. J. Food Sci. Nutr. 2017, 68, 750–756. [Google Scholar] [CrossRef]
- Grimshaw, K.E.; Maskell, J.; Oliver, E.M.; Morris, R.C.; Foote, K.D.; Mills, E.C.; Margetts, B.M.; Roberts, G. Diet and food allergy development during infancy: Birth cohort study findings using prospective food diary data. J. Allergy Clin. Immunol. 2014, 133, 511–519. [Google Scholar] [CrossRef]
- Papadopoulou, A.; Panagiotakos, D.; Hatziagorou, E.; Antonogeorgos, G.; Matziou, V.; Tsanakas, J.; Gratziou, C.; Tsabouri, S.; Priftis, K. Antioxidant foods consumption and childhood asthma and other allergic diseases: The Greek cohorts of the ISAAC II survey. Allergol. Immunopathol. 2015, 43, 353–360. [Google Scholar] [CrossRef]
- Nurmatov, U.; Devereux, G.; Sheikh, A. Nutrients and foods for the primary prevention of asthma and allergy: Systematic review and meta-analysis. J. Allergy Clin. Immunol. 2011, 127, 724–733.e30. [Google Scholar] [CrossRef]
- Rosenlund, H.; Kull, I.; Pershagen, G.; Wolk, A.; Wickman, M.; Bergstrom, A. Fruit and vegetable consumption in relation to allergy: Disease-related modification of consumption? J. Allergy Clin. Immunol. 2011, 127, 1219–1225. [Google Scholar] [CrossRef]
- Oh, H.Y.; Lee, S.; Yoon, J.; Cho, H.; Kim, Y.; Suh, D.I.; Yang, S.; Kwon, J.; Jang, G.C.; Sun, Y.H.; et al. Vegetable dietary pattern may protect mild and persistent allergic rhinitis phenotype depending on genetic risk in school children. Pediatr. Allergy Immunol. 2020, 31, 920–929. [Google Scholar] [CrossRef] [PubMed]
- Mattioli, V.; Zanolin, M.E.; Cazzoletti, L.; Bono, R.; Cerveri, I.; Ferrari, M.; Pirina, P.; Garcia-Larsen, V. Dietary flavonoids and respiratory diseases: A population-based multi-case-control study in Italian adults. Public Health Nutr. 2020, 23, 2548–2556. [Google Scholar] [CrossRef] [PubMed]
- Kusunoki, T.; Takeuchi, J.; Morimoto, T.; Sakuma, M.; Yasumi, T.; Nishikomori, R.; Higashi, A.; Heike, T. Fruit intake reduces the onset of respiratory allergic symptoms in schoolchildren. Pediatr. Allergy Immunol. 2017, 28, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Cepeda, A.M.; Del Giacco, S.R.; Villalba, S.; Tapias, E.; Jaller, R.; Segura, A.M.; Reyes, G.; Potts, J.; Garcia-Larsen, V. A traditional diet is associated with a reduced risk of eczema and wheeze in Colombian children. Nutrients 2015, 7, 5098–5110. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Larsen, V.; Thawer, N.; Charles, D.; Cassidy, A.; van Zele, T.; Thilsing, T.; Ahlström, M.; Haahtela, T.; Keil, T.; Matricardi, P.; et al. Dietary intake of flavonoids and ventilatory function in european adults: A GA(2)LEN Study. Nutrients 2015, 10, 95. [Google Scholar] [CrossRef] [PubMed]
- Woods, R.K.; Walters, E.H.; Raven, J.M.; Wolfe, R.; Ireland, P.D.; Thien, F.C.; Abramson, M.J. Food and nutrient intakes and asthma risk in young adults. Am. J. Clin. Nutr. 2003, 78, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Knekt, P.; Kumpulainen, J.; Järvinen, R.; Rissanen, H.; Heliövaara, M.; Reunanen, A.; Hakulinen, T.; Aromaa, A. Flavonoid intake and risk of chronic diseases. Am. J. Clin. Nutr. 2002, 76, 560–568. [Google Scholar] [CrossRef]
- Shaheen, S.O.; Sterne, J.A.; Thompson, R.L.; Songhurst, C.E.; Margetts, B.M.; Burney, P.G. Dietary antioxidants and asthma in adults: Population-based case–control study. Am. J. Respir. Crit. Care Med. 2001, 164, 1823–1828. [Google Scholar] [CrossRef]
- Willers, S.; Devereux, G.; Craig, L.; McNeill, G.; Wijga, A.; El-Magd, W.A.; Turner, S.; Helms, P.; Seaton, A. Maternal food consumption during pregnancy and asthma, respiratory and atopic symptoms in 5-year-old children. Thorax 2007, 62, 773–779. [Google Scholar] [CrossRef]
- van Brakel, L.; Mensink, R.P.; Wesseling, G.; Plat, J. Nutritional Interventions to Improve Asthma-Related Outcomes through Immunomodulation: A Systematic Review. Nutrients 2020, 12, 3839. [Google Scholar] [CrossRef]
- Pounis, G.; Arcari, A.; Costanzo, S.; Di Castelnuovo, A.; Bonaccio, M.; Persichillo, M.; Donati, M.B.; de Gaetano, G.; Iacoviello, L. Favorable association of polyphenol-rich diets with lung function: Cross-sectional findings from the Moli-sani study. Respir. Med. 2018, 136, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Vassilopoulou, E.; Guibas, G.V.; Papadopoulos, N.G. Mediterranean-type diets as a protective factor for asthma and atopy. Nutrients 2022, 14, 1825. [Google Scholar] [CrossRef] [PubMed]
- Koumpagioti, D.; Boutopoulou, B.; Moriki, D.; Priftis, K.N.; Douros, K. Does Adherence to the Mediterranean Diet Have a Protective Effect against Asthma and Allergies in Children? A Systematic Review. Nutrients 2022, 14, 1618. [Google Scholar] [CrossRef]
- Mazzocchi, A.; Leone, L.; Agostoni, C.; Pali-Schöll, I. The Secrets of the Mediterranean Diet. Does [Only] Olive Oil Matter? Nutrients 2019, 11, 22941. [Google Scholar] [CrossRef] [PubMed]
- Cazzoletti, L.; Zanolin, M.E.; Spelta, F.; Bono, R.; Chamitava, L.; Cerveri, I.; Garcia-Larsen, V.; Grosso, A.; Mattioli, V.; Pirina, P.; et al. Dietary fats, olive oil and respiratory diseases in Italian adults: A population-based study. Clin. Exp. Allergy 2019, 49, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Ghozal, M.; Kadawathagedara, M.; Delvert, R.; Adel-Patient, K.; Tafflet, M.; Annesi-Maesano, I.; Crépet, A.; Sirot, V.; Charles, M.A.; Heude, B.; et al. Prenatal dietary exposure to chemicals and allergy or respiratory diseases in children in the EDEN mother-child cohort. Environ. Int. 2023, 180, 108195. [Google Scholar] [CrossRef] [PubMed]
- Bessa, C.; Francisco, T.; Dias, R.; Mateus, N.; de Freitas, V.; Pérez-Gregorio, R. Use of Polyphenols as Modulators of Food Allergies. From Chemistry to Biological Implications. Front. Sustain. Food Syst. 2021, 5, 623611. [Google Scholar] [CrossRef]
- Yan, X.; Zeng, Z.; McClements, D.J.; Gong, X.; Yu, P.; Xia, J.; Gong, D. A review of the structure, function, and application of plant-based protein-phenolic conjugates and complexes. Compr. Rev. Food Sci. Food Saf. 2023, 22, 1312–1336. [Google Scholar] [CrossRef]
- Pan, T.; Wu, Y.; He, S.; Wu, Z.; Jin, R. Food allergenic protein conjugation with plant polyphenols for allergenicity reduction. Curr. Opin. Food Sci. 2022, 43, 36–42. [Google Scholar] [CrossRef]
- Zhang, Q.; Cheng, Z.; Wang, Y.; Fu, L. Dietary protein-phenolic interactions: Characterization, biochemical-physiological consequences, and potential food applications. Crit. Rev. Food Sci. Nutr. 2021, 61, 3589–3615. [Google Scholar] [CrossRef]
- Zhou, E.; Li, Q.; Zhu, D.; Chen, G.; Wu, L. Characterization of Physicochemical and Immunogenic Properties of Allergenic Proteins Altered by Food Processing: A Review. Food Sci. Hum. Wellness 2023, 13. [Google Scholar] [CrossRef]
- Liu, X.; Song, Q.; Li, X.; Chen, Y.; Liu, C.; Zhu, X.; Huang, J. Effects of different dietary polyphenols on conformational changes and functional properties of protein–polyphenol covalent complexes. Food Chem. 2021, 361, 130071. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.T.; Han, Y.N.; Wang, Y.; Zhang, Y.H.; Yin, Y.Q.; Liu, B.H.; Zhang, H.L.; Zhao, X.H. Effect of ferulic acid covalent conjugation on the functional properties and antigenicity of β-lactoglobulin. Food Chem. 2023, 406, 135095. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, Y.; Tu, Z.; Chen, W.; Yuan, T. Bovine β-Lactoglobulin Covalent Modification by Flavonoids: Effect on the Allergenicity and Human Intestinal Microbiota. J. Agric. Food Chem. 2021, 69, 6820–6828. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Xia, Y.; Chen, L.; Zhao, Y.; Wang, R.; Liang, G. Insight into covalent conjugates of β-lactoglobulin with rutin: Characterizing allergenicity, digestibility, and antioxidant properties in vitro. Food Res. Int. 2023, 173, 113401. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, K.; Mao, Y.Y.; Shao, Y.H.; Liu, J.; Tu, Z.C. A comparative study on the allergenic potential of β-lactoglobulin conjugated to glucose, caffeic acid and caffeoyl glucopyranose. Food Funct. 2023, 14, 4354–4367. [Google Scholar] [CrossRef]
- Xu, J.; Hao, M.; Sun, Q.; Tang, L. Comparative studies of interaction of β-lactoglobulin with three polyphenols. Int. J. Biol. Macromol. 2019, 136, 804–812. [Google Scholar] [CrossRef]
- Wu, X.; Lu, Y.; Xu, H.; Lin, D.; He, Z.; Wu, H.; Wang, Z. Reducing the allergenic capacity of β-lactoglobulin by covalent conjugation with dietary polyphenols. Food Chem. 2018, 256, 427–434. [Google Scholar] [CrossRef]
- Pu, P.; Zheng, X.; Jiao, L.; Chen, L.; Yang, H.; Zhang, Y.; Liang, G. Six flavonoids inhibit the antigenicity of β-lactoglobulin by noncovalent interactions: A spectroscopic and molecular docking study. Food Chem. 2021, 339, 128106. [Google Scholar] [CrossRef]
- Zhang, T.; Hu, Z.; Cheng, Y.; Xu, H.; Velickovic, T.C.; He, K.; Sun, F.; He, Z.; Liu, Z.; Wu, X. Changes in allergenicity of ovalbumin in vitro and in vivo on conjugation with quercetin. J. Agric. Food Chem. 2020, 68, 4027–4035. [Google Scholar] [CrossRef]
- Zhou, E.; Xue, X.; Xu, H.; Zhao, L.; Wu, L.; Li, Q. Effects of covalent conjugation with quercetin and its glycosides on the structure and allergenicity of Bra c p from bee pollen. Food Chem. 2023, 406, 135075. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, Z.; Li, Z.; Li, X.M.; Wang, H.; Lin, H. Insight into the conformational and allergenicity alterations of shrimp tropomyosin induced by Sargassum fusiforme polyphenol. Food Res. Int. 2023, 165, 112521. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.; Qu, X.; Yang, N.; Liu, Z.; Wu, X. Changes in structure and allergenicity of shrimp tropomyosin by dietary polyphenols treatment. Food Res. Int. 2021, 140, 109997. [Google Scholar] [CrossRef]
- Xu, X.; Cao, Q.; Yuan, J.; Tong, P.; Li, X.; Yang, A.; Wu, Z.; Liu, X.; Chen, H.; Gao, J. Shedding light on the interaction of ovalbumin and resveratrol: Structure, digestibility, transport, and allergenicity assessment of OVA-RES complexes. J. Sci. Food Agric. 2023. [Google Scholar] [CrossRef]
- Mao, T.; Wescombe, P.; Mohan, M.S. Predominance of non-covalent interactions of polyphenols with milk proteins and their health promoting properties. Crit. Rev. Food Sci. Nutr. 2023, 1–23. [Google Scholar] [CrossRef]
- He, W.; Zhang, T.; Velickovic, T.C.; Li, S.; Lyu, Y.; Wang, L.; Wu, X. Covalent conjugation with (-)-epigallo-catechin 3-gallate and chlorogenic acid changes allergenicity and functional properties of Ara h1 from peanut. Food Chem. 2020, 331, 127355. [Google Scholar] [CrossRef]
- He, W.; He, K.; Liu, X.; Ye, L.; Lin, X.; Ma, L.; Yang, P.; Wu, X. Modulating the allergenicity and functional properties of peanut protein by covalent conjugation with polyphenols. Food Chem. 2023, 415, 135733. [Google Scholar] [CrossRef] [PubMed]
- Bansode, R.R.; Randolph, P.D.; Plundrich, N.J.; Lila, M.A.; Williams, L.L. Peanut protein-polyphenol aggregate complexation suppresses allergic sensitization to peanut by reducing peanut-specific IgE in C3H/HeJ mice. Food Chem. 2019, 299, 125025. [Google Scholar] [CrossRef]
- Sun, S.; Jiang, T.; Gu, Y.; Yao, L.; Du, H.; Luo, J.; Che, H. Contribution of five major apple polyphenols in reducing peanut protein sensitization and alleviating allergencitiy of peanut by changing allergen structure. Food Res. Int. 2023, 164, 112297. [Google Scholar] [CrossRef]
- Zhang, K.; Wen, Q.; Li, T.; Zhang, Y.; Huang, J.; Huang, Q.; Gao, L. Effect of covalent conjugation with chlorogenic acid and luteolin on allergenicity and functional properties of wheat gliadin. J. Cereal Sci. 2022, 106, 103484. [Google Scholar] [CrossRef]
- Yang, R.; Shao, H.; Yan, Y.; Wu, Y.; Meng, X.; Yang, A.; Wu, Z.; Gao, J.; Li, X.; Chen, H. Changes in structural and functional properties of whey protein cross-linked by polyphenol oxidase. Food Res. Int. 2023, 164, 112377. [Google Scholar] [CrossRef] [PubMed]
- Pi, X.; Liu, J.; Sun, Y.; Ban, Q.; Cheng, J.; Guo, M. Protein modification, IgE binding capacity, and functional properties of soybean protein upon conjugation with polyphenols. Food Chem. 2023, 405, 134820. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Ye, L.; He, K.; Zhang, T.; Sun, F.; Mei, T.; Wu, X.L. A new method to reduce allergenicity by improving the functional properties of soybean 7S protein through covalent modification with polyphenols. Food Chem. 2022, 373, 131589. [Google Scholar] [CrossRef]
- Zhou, S.D.; Lin, Y.F.; Xu, X.; Meng, L.; Dong, M.S. Effect of non-covalent and covalent complexation of (-)-epigallocatechin gallate with soybean protein isolate on protein structure and in vitro digestion characteristics. Food Chem. 2020, 309, 125718. [Google Scholar] [CrossRef]
- Li, L.; Chai, W.; Ma, L.; Zhang, T.; Chen, J.; Zhang, J.; Wu, X. Covalent polyphenol with soybean 11S protein to develop hypoallergenic conjugates for potential immunotherapy. J. Funct. Foods 2023, 104, 105518. [Google Scholar] [CrossRef]
- Shakoor, H.; Feehan, J.; Apostolopoulos, V.; Platat, C.; Al Dhaheri, A.S.; Ali, H.I.; Ismail, L.C.; Bosevski, M.; Stojanovska, L. Immunomodulatory Effects of Dietary Polyphenols. Nutrients 2021, 13, 728. [Google Scholar] [CrossRef] [PubMed]
- Haftcheshmeh, S.M.; Mirhafez, S.R.; Abedi, M.; Heydarlou, H.; Shakeri, A.; Mohammadi, A.; Sahebkar, A. Therapeutic potency of curcumin for allergic diseases: A focus on immunomodulatory actions. Biomed. Pharmacother. 2022, 154, 113646. [Google Scholar] [CrossRef]
- Mlcek, J.; Jurikova, T.; Skrovankova, S.; Sochor, J. Quercetin and its anti-allergic immune response. Molecules 2016, 21, 623. [Google Scholar] [CrossRef]
- del Cornò, M.; Scazzocchio, B.; Masella, R.; Gessani, S. Regulation of Dendritic Cell Function by Dietary Polyphenols. Crit. Rev. Food Sci. Nutr. 2016, 56, 737–747. [Google Scholar] [CrossRef]
- Magrone, T.; Magrone, M.; Russo, M.A.; Jirillo, E. Recent Advances on the Anti-Inflammatory and Antioxidant Properties of Red Grape Polyphenols: In Vitro and In Vivo Studies. Antioxidants 2019, 9, 35. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhao, Y.; Han, Y.; Yang, B.; Lin, H.; Li, Z. The natural substances with anti-allergic properties in food allergy. Trends Food Sci. Technol. 2022, 128, 53–67. [Google Scholar] [CrossRef]
- Shaik, Y.; Caraffa, A.; Ronconi, G.; Lessiani, G.; Conti, P. Impact of polyphenols on mast cells with special emphasis on the effect of quercetin and luteolin. Cent.-Eur. J. Immunol. 2018, 43, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Mwakalukwa, R.; Ashour, A.; Amen, Y.; Niwa, Y.; Shimizu, K. Anti-allergic activity of polyphenolic compounds isolated from olive mill wastes. J. Funct. Foods 2019, 58, 207–217. [Google Scholar] [CrossRef]
- Humeniuk, P.; Dubiela, P.; Hoffmann-Sommergruber, K. Dendritic cells and their role in allergy: Uptake, proteolytic processing and presentation of allergens. Int. J. Mol. Sci. 2017, 18, 1491. [Google Scholar] [CrossRef] [PubMed]
- Svajger, U.; Obermajer, N.; Jeras, M. Dendritic cells treated with resveratrol during differentiation from monocytes gain substantial tolerogenic properties upon activation. Immunology 2010, 129, 525–535. [Google Scholar] [CrossRef]
- Buttari, B.; Profumo, E.; Facchiano, F.; Ozturk, E.I.; Segoni, L.; Saso, L.; Riganò, R. Resveratrol prevents dendritic cell maturation in response to advanced glycation end products. Oxidative Med. Cell. Longev. 2013, 2013, 574029. [Google Scholar] [CrossRef]
- Huang, R.Y.; Yu, Y.L.; Cheng, W.C.; OuYang, C.N.; Fu, E.; Chu, C.L. Immunosuppressive effect of quercetin on dendritic cell activation and function. J. Immunol. 2010, 184, 6815–6821. [Google Scholar] [CrossRef]
- Yoneyama, S.; Kawai, K.; Tsuno, N.H.; Okaji, Y.; Asakage, M.; Tsuchiya, T.; Yamada, J.; Sunami, E.; Osada, T.; Kitayama, J.; et al. Epigallocatechin gallate affects human dendritic cell differentiation and maturation. J. Allergy Clin. Immunol. 2008, 121, 209–214. [Google Scholar] [CrossRef]
- Yoon, M.S.; Lee, J.S.; Choi, B.M.; Jeong, Y.I.; Lee, C.M.; Park, J.H.; Moon, Y.; Sung, S.C.; Lee, S.K.; Chang, Y.H.; et al. Apigenin inhibits immunostimulatory function of dendritic cells: Implication of immunotherapeutic adjuvant. Mol. Pharmacol. 2006, 70, 1033–1044. [Google Scholar] [CrossRef]
- Liu, S.H.; Lin, C.H.; Hung, S.K.; Chou, J.H.; Chi, C.W.; Fu, S.L. Fisetin inhibits lipopolysaccharide-induced macrophage activation and dendritic cell maturation. J. Agric. Food Chem. 2010, 58, 10831–10839. [Google Scholar] [CrossRef]
- Dai, J.; Patel, J.D.; Mumper, R.J. Characterization of blackberry extract and its antiproliferative and anti-inflammatory properties. J. Med. Food 2007, 10, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.K.; Ecklund, D.; Liu, M.; Datta, S.K. Apigenin, a non-mutagenic dietary flavonoid, suppresses lupus by inhibiting autoantigen presentation for expansion of autoreactive Th1 and Th17 cells. Arthritis Res. Ther. 2009, 11, R59. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Jeong, G.S. Therapeutic effect of kaempferol on AD by attenuation of T cell activity via interaction with multidrug associated protein. Br. J. Pharmacol. 2021, 178, 1772–1788. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Kim, J.; Choi, H.G.; Kim, E.K.; Jun, C.D. Licoricidin Abrogates T-Cell Activation by Modulating PTPN1 Activity and Attenuates Atopic Dermatitis In Vivo. J. Investig. Dermatol. 2021, 141, 2490–2498.e6. [Google Scholar] [CrossRef] [PubMed]
- Pasha, M.A.; Patel, G.; Hopp, R.; Yang, Q. Role of innate lymphoid cells in allergic diseases. Allergy Asthma Proc. 2019, 40, 138–145. [Google Scholar] [CrossRef]
- Hammad, H.; Lambrecht, B.N. Barrier Epithelial Cells and the Control of Type 2 Immunity. Immunity 2015, 43, 29–40. [Google Scholar] [CrossRef]
- Zheng, H.; Zhang, Y.; Pan, J.; Liu, N.; Qin, Y.; Qiu, L.; Liu, M.; Wang, T. The Role of Type 2 Innate Lymphoid Cells in Allergic Diseases. Front. Immunol. 2021, 12, 586078. [Google Scholar] [CrossRef]
- Beken, B.; Serttas, R.; Yazicioglu, M.; Turkekul, K.; Erdogan, S. Quercetin Improves Inflammation, Oxidative Stress, and Impaired Wound Healing in Atopic Dermatitis Model of Human Keratinocytes. Pediatr. Allergy Immunol. Pulmonol. 2020, 33, 69–79. [Google Scholar] [CrossRef]
- Wang, L.; Xian, Y.F.; Loo, S.K.F.; Ip, S.P.; Yang, W.; Chan, W.Y.; Lin, Z.X.; Wu, J.C.Y. Baicalin ameliorates 2,4-dinitrochlorobenzene-induced atopic dermatitis-like skin lesions in mice through modulating skin barrier function, gut microbiota and JAK/STAT pathway. Bioorg. Chem. 2022, 119, 105538. [Google Scholar] [CrossRef]
- Sharma, S.; Sethi, G.S.; Naura, A.S. Curcumin Ameliorates Ovalbumin-Induced Atopic Dermatitis and Blocks the Progression of Atopic March in Mice. Inflammation 2020, 43, 358–369. [Google Scholar] [CrossRef]
- Moon, P.D.; Han, N.R.; Lee, J.S.; Jee, H.W.; Kim, J.H.; Kim, H.M.; Jeong, H.J. Effects of Resveratrol on Thymic Stromal Lymphopoietin Expression in Mast Cells. Medicina 2020, 57, 21. [Google Scholar] [CrossRef] [PubMed]
- Moon, P.D.; Choi, I.H.; Kim, H.M. Berberine inhibits the production of thymic stromal lymphopoietin by the blockade of caspase-1/NF-κB pathway in mast cells. Int. Immunopharmacol. 2011, 11, 1954–1959. [Google Scholar] [CrossRef]
- Caglayan Sozmen, S.; Karaman, M.; Cilaker Micili, S.; Isik, S.; Bagriyanik, A.; Arikan Ayyildiz, Z.; Uzuner, N.; Anal, O.; Karaman, O. Effects of Quercetin Treatment on Epithelium-derived Cytokines and Epithelial Cell Apoptosis in Allergic Airway Inflammation Mice Model. Iran. J. Allergy Asthma Immunol. 2016, 15, 487–497. [Google Scholar] [PubMed]
- Jin, J.; Fan, Y.J.; Nguyen, T.V.; Yu, Z.N.; Song, C.H.; Lee, S.-Y.; Shin, H.S.; Chai, O.H. Fallopia japonica Root Extract Ameliorates Ovalbumin-Induced Airway Inflammation in a CARAS Mouse Model by Modulating the IL-33/TSLP/NF-B Signaling Pathway. Int. J. Mol. Sci. 2023, 24, 12514. [Google Scholar] [CrossRef] [PubMed]
- Satitsuksanoa, P.; Daanje, M.; Akdis, M.; Boyd, S.D.; van de Veen, W. Biology and dynamics of B cells in the context of IgE-mediated food allergy. Allergy 2021, 76, 1707–1717. [Google Scholar] [CrossRef]
- Singh, A.; Holvoet, S.; Mercenier, A. Dietary polyphenols in the prevention and treatment of allergic diseases. Clin. Exp. Allergy 2011, 41, 1346–1359. [Google Scholar] [CrossRef]
- Acar, M.; Muluk, N.B.; Yigitaslan, S.; Cengiz, B.P.; Shojaolsadati, P.; Karimkhani, H.; Ada, S.; Berkoz, M.; Cingi, C. Can curcumin modulate allergic rhinitis in rats? J. Laryngol. Otol. 2016, 130, 1103–1109. [Google Scholar] [CrossRef]
- Shahgordi, S.; Sankian, M.; Yazdani, Y.; Mashayekhi, K.; Hasan Ayati, S.; Sadeghi, M.; Saeidi, M.; Hashemi, M. Immune responses modulation by curcumin and allergen encapsulated into PLGA nanoparticles in mice model of rhinitis allergic through sublingual immunotherapy. Int. Immunopharmacol. 2020, 84, 106525. [Google Scholar] [CrossRef]
- Mine, Y.; Majumder, K.; Jin, Y.; Zeng, Y. Chinese sweet tea (Rubus suavissimus) polyphenols attenuate the allergic responses in a Balb/c mouse model of egg allergy. J. Funct. Foods 2020, 67, 103827. [Google Scholar] [CrossRef]
- Zhou, Z.; Shi, T.; Hou, J.; Li, M. Ferulic acid alleviates atopic dermatitis-like symptoms in mice via its potent anti-inflammatory effect. Immunopharmacol. Immunotoxicol. 2020, 42, 156–164. [Google Scholar] [CrossRef]
- Xing, Y.; Tan, C.; Luo, Y.; Liu, W. Effect of quercetin on rhinitis via inflammatory pathway. Pak. J. Zool. 2021, 53, 619–626. [Google Scholar] [CrossRef]
- Liang, Z.; Wu, L.; Deng, X.; Liang, Q.; Xu, Y.; Deng, R.; Lv, L.; Ji, M.; Hao, Z.; He, J. The antioxidant rosmarinic acid ameliorates oxidative lung damage in experimental allergic asthma via modulation of NADPH oxidases and antioxidant enzymes. Inflammation 2020, 43, 1902–1912. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.F.; Liu, Q.M.; Liu, B.; Shu, Z.D.; Han, J.; Liu, H.; Liu, G.M. Dihydromyricetin inhibited ovalbumin-induced mice allergic responses by suppressing the activation of mast cells. Food Funct. 2019, 10, 7131–7141. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, Y.; Katsuzaki, H.; Imai, K.; Amano, H. The anti-allergic and anti-inflammatory effects of phlorotannins from the edible brown algae, Ecklonia sp. and Eisenia sp. Nat. Prod. Commun. 2021, 16, 1934578X211060924. [Google Scholar] [CrossRef]
- Kim, M.J.; Je, I.G.; Song, J.; Fei, X.; Lee, S.; Yang, H.; Kang, W.; Jang, Y.H.; Seo, S.Y.; Kim, S.H. SG-SP1 Suppresses Mast Cell-Mediated Allergic Inflammation via Inhibition of FcεRI Signaling. Front. Immunol. 2020, 11, 50. [Google Scholar] [CrossRef]
- Li, Q.-S.; Wang, Y.-Q.; Liang, Y.-R.; Lu, J.-L. The anti-allergic potential of tea: A review of its components, mechanisms and risks. Food Funct. 2021, 12, 57–69. [Google Scholar] [CrossRef]
- Kong, Z.L.; Sudirman, S.; Lin, H.J.; Chen, W.N. In vitro anti-inflammatory effects of curcumin on mast cell-mediated allergic responses via inhibiting FcεRI protein expression and protein kinase C delta translocation. Cytotechnology 2020, 72, 81–95. [Google Scholar] [CrossRef]
- Alam, S.B.; Wagner, A.; Willows, S.; Kulka, M. Quercetin and Resveratrol Differentially Decrease Expression of the High-Affinity IgE Receptor (FcεRI) by Human and Mouse Mast Cells. Molecules 2020, 27, 6704. [Google Scholar] [CrossRef]
- Nagata, K.; Araumi, S.; Ando, D.; Ito, N.; Ando, M.; Ikeda, Y.; Takahashi, M.; Noguchi, S.; Yasuda, Y.; Nakano, N.; et al. Kaempferol Suppresses the Activation of Mast Cells by Modulating the Expression of FcεRI and SHIP1. Int. J. Mol. Sci. 2023, 24, 5997. [Google Scholar] [CrossRef]
- Civelek, M.; Bilotta, S.; Lorentz, A. Resveratrol Attenuates Mast Cell Mediated Allergic Reactions: Potential for Use as a Nutraceutical in Allergic Diseases? Mol. Nutr. Food Res. 2020, 66, e2200170. [Google Scholar] [CrossRef]
- Kandhare, A.D.; Aswar, U.M.; Mohan, V.; Thakurdesai, P.A. Ameliorative effects of type-A procyanidins polyphenols from cinnamon bark in compound 48/80-induced mast cell degranulation. Anat. Cell Biol. 2017, 50, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Vo, T.S.; Le, T.T.; Kim, S.Y.; Ngo, D.H. The role of myricetin from Rhodomyrtus tomentosa (Aiton) Hassk fruits on downregulation of FcεRI-mediated mast cell activation. J. Food Biochem. 2020, 44, e13143. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Zorig, A.; Sato, N.; Yanagihara, A.; Kanazawa, T.; Takasugi, M.; Arai, H. Effect of Polyphenols in Sea Buckthorn Berry on Chemical Mediator Release from Mast Cells. Prev. Nutr. Food Sci. 2023, 28, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Park, J. Anti-Anaphylactic Activity of Isoquercitrin (Quercetin-3-O-β-d-Glucose) in the Cardiovascular System of Animals. Biomedicines 2020, 8, 139. [Google Scholar] [CrossRef] [PubMed]
- Yousef, M.; Crozier, R.W.E.; Hicks, N.J.; Watson, C.J.F.; Boyd, T.; Tsiani, E.; MacNeil, A.J. Attenuation of allergen-mediated mast cell activation by rosemary extract (Rosmarinus officinalis L.). J. Leukoc. Biol. 2020, 107, 843–857. [Google Scholar] [CrossRef]
- Zeng, J.; Hao, J.; Yang, Z.; Ma, C.; Gao, L.; Chen, Y.; Li, G.; Li, J. Anti-Allergic Effect of Dietary Polyphenols Curcumin and Epigallocatechin Gallate via Anti-Degranulation in IgE/Antigen-Stimulated Mast Cell Model: A Lipidomics Perspective. Metabolites 2023, 13, 628. [Google Scholar] [CrossRef]
- Barbosa, M.; Lopes, G.; Andrade, P.B.; Valentão, P. Bioprospecting of brown seaweeds for biotechnological applications: Phlorotannin actions in inflammation and allergy network. Trends Food Sci. Technol. 2019, 86, 153–171. [Google Scholar] [CrossRef]
- Hwang, D.; Kang, M.; Kang, C.; Kim, G. Kaempferol-3-O-β-rutinoside suppresses the inflammatory responses in lipopolysaccharide-stimulated RAW264.7 cells via the NF-κB and MAPK pathways. Int. J. Mol. Med. 2019, 44, 2321–2328. [Google Scholar] [CrossRef]
- Cao, J.; Wang, Y.; Hu, S.; Ding, Y.; Jia, Q.; Zhu, J.; An, H. Kaempferol ameliorates secretagogue-induced pseudo-allergic reactions via inhibiting intracellular calcium fluctuation. J. Pharm. Pharmacol. 2020, 72, 1221–1231. [Google Scholar] [CrossRef]
- Crozier, R.W.E.; Yousef, M.; Coish, J.M.; Fajardo, V.A.; Tsiani, E.; MacNeil, A.J. Carnosic acid inhibits secretion of allergic inflammatory mediators in IgE-activated mast cells via direct regulation of Syk activation. J. Biol. Chem. 2023, 299, 102867. [Google Scholar] [CrossRef]
- Yong, P.Y.A.; Yip, A.J.W.; Islam, F.; Hong, H.J.; Teh, Y.E.; Tham, C.L.; Tan, J.W. The anti-allergic potential of stingless bee honey from different botanical sources via modulation of mast cell degranulation. BMC Complement. Med. Ther. 2023, 23, 307. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J. Pathophysiology of allergic inflammation. Immunol. Rev. 2011, 242, 31–50. [Google Scholar] [CrossRef] [PubMed]
- Shahid, H.; Shahzad, M.; Shabbir, A.; Saghir, G. Immunomodulatory and Anti-Inflammatory Potential of Curcumin for the Treatment of Allergic Asthma: Effects on Expression Levels of Pro-inflammatory Cytokines and Aquaporins. Inflammation 2019, 42, 2037–2047. [Google Scholar] [CrossRef]
- Chauhan, P.S.; Jaiswal, A.; Subhashini; Singh, R. Combination Therapy with Curcumin Alone Plus Piperine Ameliorates Ovalbumin-Induced Chronic Asthma in Mice. Inflammation 2018, 41, 1922–1933. [Google Scholar] [CrossRef]
- Molitorisova, M.; Sutovska, M.; Kazimierova, I.; Barborikova, J.; Joskova, M.; Novakova, E.; Franova, S. The anti-asthmatic potential of flavonol kaempferol in an experimental model of allergic airway inflammation. Eur. J. Pharmacol. 2021, 891, 173698. [Google Scholar] [CrossRef] [PubMed]
- Shakeri, F.; Eftekhar, N.; Roshan, N.M.; Rezaee, R.; Moghimi, A.; Boskabady, M. Rosmarinic acid affects immunological and inflammatory mediator levels and restores lung pathological features in asthmatic rats. Allergol. Immunopathol. 2019, 47, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Xu, O.; Wang, J.; Shan, C.; Ren, X. Luteolin ameliorates inflammation and Th1/Th2 imbalance via regulating the TLR4/NF-kB pathway in allergic rhinitis rats. Immunopharmacol. Immunotoxicol. 2021, 43, 319–327. [Google Scholar] [CrossRef]
- Feng, H.; Xiong, X.; Xu, Q.; Zhang, Z.; Feng, J.; Wu, Y. Study on the immunomodulatory effect of quercetin nanoparticles loaded with chitosan on a mouse model of ovalbumin-induced food allergy. Nanosci. Nanotechnol. Lett. 2020, 12, 915–920. [Google Scholar] [CrossRef]
- Hwang, K.-A.; Hwang, Y.-J.; Song, J. Anti-allergic effect of aster yomena on ovalbumin-sensitized mouse and RHL-2H3 Cells via Th1/Th2 cytokine balance. J. Funct. Foods 2018, 44, 1–8. [Google Scholar] [CrossRef]
- Ke, X.; Chen, Z.; Wang, X.; Kang, H.; Hong, S. Quercetin improves the imbalance of Th1/Th2 cells and Treg/Th17 cells to attenuate allergic rhinitis. Autoimmunity 2023, 56, 2189133. [Google Scholar] [CrossRef]
- Shi, Y.N.; Su, J.Z.; Wang, J.; Geng, J.Q. Myricetin alleviates ovalbumin-induced allergic rhinitis in mice by regulating Th1/Th2 balance. Asian Pac. J. Trop. Biomed. 2023, 13, 306–314. [Google Scholar]
- Tortola, L.; Jacobs, A.; Pohlmeier, L.; Obermair, F.J.; Ampenberger, F.; Bodenmiller, B.; Kopf, M. High-dimensional T helper cell profiling reveals a broad diversity of stably committed effector states and uncovers interlineage Relationships. Immunity 2020, 53, 597. [Google Scholar] [CrossRef] [PubMed]
- Boonpiyathad, T.; Sözener, Z.C.; Akdis, M.; Akdis, C.A. The role of Treg cell subsets in allergic disease. Asian Pac. J. Allergy Immunol. 2020, 30, 139–149. [Google Scholar] [CrossRef]
- Li, J.; Lin, X.; Liu, X.; Ma, Z.; Li, Y. Baicalin regulates Treg/Th17 cell imbalance by inhibiting autophagy in allergic rhinitis. Mol. Immunol. 2020, 125, 162–171. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, L.; Wang, S.; Wang, Y.; Du, Y.; Fan, Y. Luteolin restored Treg/Th17 balance to ameliorate allergic rhinitis in a mouse model. Immunopharmacol. Immunotoxicol. 2023, 45, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhu, L.; Fukuda, K.; Ouyang, S.; Chen, X.; Wang, C.; Zhang, C.J.; Martin, B.; Gu, C.; Qin, L.; et al. The flavonoid cyanidin blocks binding of the cytokine interleukin-17A to the IL-17RA subunit to alleviate inflammation in vivo. Sci. Signal. 2017, 10, eaaf8823. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.F.; Chen, Y.Q.; Li, Q.; Ye, X.Y.; Zuo, X.; Shi, Y.L.; Guo, X.Y.; Xu, L.; Sun, L.; Li, C.W.; et al. Supplementation with Tetrahydrocurcumin Enhances the Therapeutic Effects of Dexamethasone in a Murine Model of Allergic Asthma. Int. Arch. Allergy Immunol. 2020, 181, 822–830. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Ma, Z.; Fu, Q.; Ma, S. Curcumin attenuates allergic airway inflammation by regulation of CD4+CD25+ regulatory T cells (Tregs)/Th17 balance in ovalbumin-sensitized mice. Fitoterapia 2013, 87, 57–64. [Google Scholar] [CrossRef]
- Chen, B.L.; Chen, Y.Q.; Ma, B.H.; Yu, S.F.; Li, L.Y.; Zeng, Q.X.; Zhou, Y.T.; Wu, Y.F.; Liu, W.L.; Wan, J.B.; et al. Tetrahydrocurcumin, a major metabolite of curcumin, ameliorates allergic airway inflammation by attenuating Th2 response and suppressing the IL-4Rα-Jak1-STAT6 and Jagged1/Jagged2-Notch1/Notch2 pathways in asthmatic mice. Clin. Exp. Allergy 2018, 48, 1494–1508. [Google Scholar] [CrossRef]
- Stiemsma, L.T.; Michels, K.B. The Role of the Microbiome in the Developmental Origins of Health and Disease. Pediatrics 2018, 141, e20172437. [Google Scholar] [CrossRef]
- Hufnagl, K.; Pali-Schöll, I.; Roth-Walter, F.; Jensen-Jarolim, E. Dysbiosis of the gut and lung microbiome has a role in asthma. Semin. Immunopathol. 2020, 42, 75–93. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.H.; Song, Y.; Wu, W.; Yu, K.; Zhang, G. The gut microbiota, environmental factors, and links to the development of food allergy. Clin. Mol. Allergy 2020, 18, 5. [Google Scholar] [CrossRef] [PubMed]
- Han, P.; Gu, J.Q.; Li, L.S.; Wang, X.Y.; Wang, H.T.; Wang, Y.; Chang, C.; Sun, J.L. The Association between Intestinal Bacteria and Allergic Diseases-Cause or Consequence? Front. Cell. Infect. Microbiol. 2021, 11, 650893. [Google Scholar] [CrossRef] [PubMed]
- Pantazi, A.C.; Mihai, C.M.; Balasa, A.L.; Chisnoiu, T.; Lupu, A.; Frecus, C.E.; Mihai, L.; Ungureanu, A.; Kassim, M.A.K.; Andrusca, A.; et al. Relationship between Gut Microbiota and Allergies in Children: A Literature Review. Nutrients 2023, 15, 2529. [Google Scholar] [CrossRef]
- Hoskinson, C.; Dai, D.L.Y.; Del Bel, K.L.; Becker, A.B.; Moraes, T.J.; Mandhane, P.J.; Finlay, B.B.; Simons, E.; Kozyrskyj, A.L.; Azad, M.B.; et al. Delayed gut microbiota maturation in the first year of life is a hallmark of pediatric allergic disease. Nat. Commun. 2023, 14, 4785. [Google Scholar] [CrossRef]
- Cheng, H.; Zhang, D.; Wu, J.; Liu, J.; Zhou, Y.; Tan, Y.; Feng, W.; Peng, C. Interactions between gut microbiota and polyphenols: A mechanistic and metabolomic review. Phytomedicine 2023, 119, 154979. [Google Scholar] [CrossRef]
- Alves-Santos, A.M.; Sugizaki, C.S.A.; Lima, G.C.; Naves, M.M.V. Prebiotic effect of dietary polyphenols: A systematic review. J. Funct. Foods 2020, 74, 104169. [Google Scholar] [CrossRef]
- Pei, R.; Liu, X.; Bolling, B. Flavonoids and gut health. Curr. Opin. Biotechnol. 2020, 61, 153–159. [Google Scholar] [CrossRef]
- Ulusoy, H.G.; Sanlier, N. A minireview of quercetin: From its metabolism to possible mechanisms of its biological activities. Crit. Rev. Food Sci. Nutr. 2020, 60, 3290–3303. [Google Scholar] [CrossRef]
- Peterson, C.T.; Vaughn, A.R.; Sharma, V.; Chopra, D.; Mills, P.J.; Peterson, S.N.; Sivamani, R.K. Effects of turmeric and curcumin dietary supplementation on human gut microbiota: A double-blind, randomized, placebo-controlled pilot study. J. Evid.-Based Integr. Med. 2018, 23, 2515690X18790725. [Google Scholar] [CrossRef]
- Chen, F.; Wen, Q.; Jiang, J.; Li, H.L.; Tan, Y.F.; Li, Y.H.; Zeng, N.K. Could the gut microbiota reconcile the oral bioavailability conundrum of traditional herbs? J. Ethnopharmacol. 2016, 179, 253–264. [Google Scholar] [CrossRef] [PubMed]
- Gwiazdowska, D.; Juś, K.; Jasnowska-Małecka, J.; Kluczyńska, K. The impact of polyphenols on Bifidobacterium growth. Acta Biochim. Pol. 2015, 62, 895–901. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.; Bian, X.; Yao, Z.; Wang, Y.; Gao, W.; Guo, C. Quercetin improves gut dysbiosis in antibiotic-treated mice. Food Funct. 2020, 11, 8003–8013. [Google Scholar] [CrossRef] [PubMed]
- Oteiza, P.I.; Fraga, C.G.; Mills, D.A.; Taft, D.H. Flavonoids and the gastrointestinal tract: Local and systemic effects. Mol. Asp. Med. 2018, 61, 41–49. [Google Scholar] [CrossRef]
- Yang, T.; Li, C.; Xue, W.; Huang, L.; Wang, Z. Natural immunomodulating substances used for alleviating food allergy. Crit. Rev. Food Sci. Nutr. 2023, 63, 2407–2425. [Google Scholar] [CrossRef]
- Masumoto, S.; Terao, A.; Yamamoto, Y.; Mukai, T.; Miura, T.; Shoji, T. Non-absorbable apple procyanidins prevent obesity associated with gut microbial and metabolomic changes. Sci. Rep. 2016, 6, 31208. [Google Scholar] [CrossRef]
- Wang, J.; Tang, L.; Zhou, H.; Zhou, J.; Glenn, T.C.; Shen, C.L.; Wang, J.S. Long-term treatment with green tea polyphenols modifies the gut microbiome of female sprague-dawley rats. J. Nutr. Biochem. 2018, 56, 55–64. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, Y.; Zhu, J.; Zhang, M.; Ho, C.T.; Huang, Q.; Cao, J. Metagenomics analysis of gut microbiota in a high fat diet–induced obesity mouse model fed with (−)-epigallocatechin 3-O-(3-O-Methyl) gallate (EGCG3″Me). Mol. Nutr. Food Res. 2018, 62, 1800274. [Google Scholar] [CrossRef]
- Sáyago-Ayerdi, S.G.; Zamora-Gasga, V.M.; Venema, K. Prebiotic effect of predigested mango peel on gut microbiota assessed in a dynamic in vitro model of the human colon (TIM-2). Food Res. Int. 2019, 118, 89–95. [Google Scholar] [CrossRef]
- Smeriglio, A.; Barreca, D.; Bellocco, E.; Trombetta, D. Proanthocyanidins and hydrolysable tannins: Occurrence, dietary intake and pharmacological effects. Br. J. Pharmacol. 2017, 174, 1244–1262. [Google Scholar] [CrossRef]
- Marchesi, J.R.; Adams, D.H.; Fava, F.; Hermes, G.D.; Hirschfield, G.M.; Hold, G.; Hart, A. The gut microbiota and host health: A new clinical frontier. Gut 2016, 65, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhong, J.; Meng, X.; Gao, J.; Li, H.; Sun, J.; Chen, H. The gut microbiome-immune axis as a target for nutrition-mediated modulation of food allergy. Trends Food Sci. Technol. 2021, 114, 116–132. [Google Scholar] [CrossRef]
- Liang, J.; Zheng, B.; Zhang, Y.; Zeng, H. Food allergy and gut microbiota. Trends Food Sci. Technol. 2023, 140, 104141. [Google Scholar] [CrossRef]
- Cheng, Y.; Liu, X.; Chen, F.; Rolnik, B.M.; Chleilat, F.; Ling, Z.; Zhou, X. The Roles and Mechanisms of Gut Microbiota in Food Allergy. Adv. Gut Microbiome Res. 2023, 2023, 9575410. [Google Scholar] [CrossRef]
- Shim, J.A.; Ryu, J.H.; Jo, Y.; Hong, C. The role of gut microbiota in T cell immunity and immune mediated disorders. Int. J. Biol. Sci. 2023, 19, 1178. [Google Scholar] [CrossRef]
- Peng, Y.; Yan, Y.; Wan, P.; Dong, W.; Huang, K.; Ran, L.; Cao, Y. Effects of long-term intake of anthocyanins from Lycium ruthenicum Murray on the organism health and gut microbiota in vivo. Food Res. Int. 2020, 130, 108952. [Google Scholar] [CrossRef]
- Li, J.; Zou, C.; Liu, Y. Amelioration of Ovalbumin-Induced Food Allergy in Mice by Targeted Rectal and Colonic Delivery of Cyanidin-3-O-Glucoside. Foods 2022, 11, 1542. [Google Scholar] [CrossRef]
- Zhu, L.; Xu, L.Z.; Zhao, S.; Shen, Z.F.; Shen, H.; Zhan, L.B. Protective effect of baicalin on the regulation of Treg/Th17 balance, gut microbiota and short-chain fatty acids in rats with ulcerative colitis. Appl. Microbiol. Biotechnol. 2020, 104, 5449–5460. [Google Scholar] [CrossRef]
- Hu, R.; Wu, S.; Li, B.; Tan, J.; Yan, J.; Wang, Y.; He, J. Dietary ferulic acid and vanillic acid on inflammation, gut barrier function and growth performance in lipopolysaccharide-challenged piglets. Anim. Nutr. 2022, 8, 144–152. [Google Scholar] [CrossRef]
- Camps-Bossacoma, M.; Pérez-Cano, F.J.; Franch, À.; Castell, M. Gut Microbiota in a Rat Oral Sensitization Model: Effect of a Cocoa-Enriched Diet. Oxidative Med. Cell. Longev. 2017, 2017, 7417505. [Google Scholar] [CrossRef]
- Dang, A.T.; Marsland, B.J. Microbes, metabolites, and the gut–lung axis. Mucosal Immunol. 2019, 12, 843–850. [Google Scholar] [CrossRef]
- Espírito Santo, C.; Caseiro, C.; Martins, M.J.; Monteiro, R.; Brandão, I. Gut Microbiota, in the Halfway between Nutrition and Lung Function. Nutrients 2021, 13, 1716. [Google Scholar] [CrossRef] [PubMed]
- Bingula, R.; Filaire, M.; Radosevic-Robin, N.; Bey, M.; Berthon, J.Y.; Bernalier-Donadille, A.; Vasson, M.P.; Filaire, E. Desired Turbulence? Gut-Lung Axis, Immunity, and Lung Cancer. J. Oncol. 2017, 2017, 5035371. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Li, S.; Wang, N.; Tan, H.-Y.; Zhang, Z.; Feng, Y. The Cross-Talk between Gut Microbiota and Lungs in Common Lung Diseases. Front. Microbiol. 2020, 11, 301. [Google Scholar] [CrossRef] [PubMed]
- Enaud, R.; Prevel, R.; Ciarlo, E.; Beaufils, F.; Wieërs, G.; Guery, B.; Delhaes, L. The gut-lung axis in health and respiratory diseases: A place for inter-organ and inter-kingdom crosstalks. Front. Cell. Infect. Microbiol. 2020, 10, 9. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Liu, C.; Zhang, K.; Xue, W. The role of gut microbiota and its metabolites short-chain fatty acids in food allergy. Food Sci. Hum. Wellness 2023, 12, 702–710. [Google Scholar] [CrossRef]
- Akagawa, S.; Kaneko, K. Gut microbiota and allergic diseases in children. Allergol. Int. 2022, 71, 301–309. [Google Scholar] [CrossRef]
- Kawabata, K.; Yoshioka, Y.; Terao, J. Role of intestinal microbiota in the bioavailability and physiological functions of dietary polyphenols. Molecules 2019, 24, 370. [Google Scholar] [CrossRef]
- Luo, Q.; Cheng, D.; Huang, C.; Li, Y.; Lao, C.; Xia, Y.; Chen, Z. Improvement of colonic immune function with soy isoflavones in high-fat diet-induced obese rats. Molecules 2019, 24, 1139. [Google Scholar] [CrossRef]
- Ma, H.; Zhang, B.; Hu, Y.; Wang, J.; Liu, J.; Qin, R.; Wang, S. Correlation analysis of intestinal redox state with the gut microbiota reveals the positive intervention of tea polyphenols on hyperlipidemia in high fat diet fed mice. J. Agric. Food Chem. 2019, 67, 7325–7335. [Google Scholar] [CrossRef]
- Wu, Z.; Huang, S.; Li, T.; Li, N.; Han, D.; Zhang, B.; Xu, Z.Z.; Zhang, S.; Pang, J.; Wang, S.; et al. Gut microbiota from green tea polyphenol-dosed mice improves intestinal epithelial homeostasis and ameliorates experimental colitis. Microbiome 2021, 9, 184. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Ma, N.; Song, P.; He, T.; Levesque, C.; Bai, Y.; Zhang, A.; Ma, X. Grape Seed Proanthocyanidin Affects Lipid Metabolism via Changing Gut Microflora and Enhancing Propionate Production in Weaned Pigs. J. Nutr. 2019, 149, 1523–1532. [Google Scholar] [CrossRef] [PubMed]
- Alharris, E.; Mohammed, A.; Alghetaa, H.; Zhou, J.; Nagarkatti, M.; Nagarkatti, P. The Ability of Resveratrol to Attenuate Ovalbumin-Mediated Allergic Asthma Is Associated with Changes in Microbiota Involving the Gut-Lung Axis, Enhanced Barrier Function and Decreased Inflammation in the Lungs. Front. Immunol. 2022, 13, 805770. [Google Scholar] [CrossRef]
- Zhang, Q.; Yu, X.; Tian, L.; Cong, Y.; Li, L. Therapeutic effects of epigallocatechin and epigallocatechin gallate on the allergic reaction of αs1-casein sensitized mice. Food Sci. Hum. Wellness 2023, 12, 882–888. [Google Scholar] [CrossRef]
- Shin, H.S.; See, H.J.; Jung, S.Y.; Choi, D.W.; Kwon, D.A.; Bae, M.J.; Sung, K.S.; Shon, D.H. Turmeric (Curcuma longa) attenuates food allergy symptoms by regulating type 1/type 2 helper T cells (Th1/Th2) balance in a mouse model of food allergy. J. Ethnopharmacol. 2015, 175, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.F.; Liu, Q.M.; Gao, Y.Y.; Liu, B.; Liu, H.; Cao, M.J.; Liu, G.M. Attenuation of allergic responses following treatment with resveratrol in anaphylactic models and IgE-mediated mast cells. Food Funct. 2019, 10, 2030–2039. [Google Scholar] [CrossRef]
- Singh, A.; Demont, A.; Actis-Goretta, L.; Holvoet, S.; Lévêques, A.; Lepage, M.; Nutten, S.; Mercenier, A. Identification of epicatechin as one of the key bioactive constituents of polyphenol-enriched extracts that demonstrate an anti-allergic effect in a murine model of food allergy. Br. J. Nutr. 2014, 112, 358–368. [Google Scholar] [CrossRef]
- Wang, C.C.; Lin, Y.R.; Liao, M.H.; Jan, T.R. Oral supplementation with areca-derived polyphenols attenuates food allergic responses in ovalbumin-sensitized mice. BMC Complement. Altern. Med. 2013, 13, 154. [Google Scholar] [CrossRef]
- Abril-Gil, M.; Pérez-Cano, F.J.; Franch, À.; Castell, M. Effect of a cocoa-enriched diet on immune response and anaphylaxis in a food allergy model in Brown Norway rats. J. Nutr. Biochem. 2016, 27, 317–326. [Google Scholar] [CrossRef]
- Ding, S.; Jiang, H.; Fang, J. Regulation of Immune Function by Polyphenols. J. Immunol. Res. 2018, 2018, 1264074. [Google Scholar] [CrossRef]
- Bae, M.-J.; Shin, H.S.; See, H.-J.; Jung, S.Y.; Kwon, D.-A.; Shon, D.-H. Baicalein induces CD4+Foxp3+ T cells and enhances intestinal barrier function in a mouse model of food allergy. Sci. Rep. 2016, 6, 32225. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Li, J.; Guo, Y.; Ma, L.; Liu, Y.; Kuang, H.; Wang, Y. Dietary olive oil enhances the oral tolerance of the food allergen ovalbumin in mice by regulating intestinal microecological homeostasis. J. Food Biochem. 2022, 46, e14297. [Google Scholar] [CrossRef] [PubMed]
- Fujiki, T.; Shinozaki, R.; Udono, M.; Katakura, Y. Identification and Functional Evaluation of Polyphenols That Induce Regulatory T Cells. Nutrients 2022, 14, 2862. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M. Multifunctions of dietary polyphenols in the regulation of intestinal inflammation. J. Food Drug Anal. 2017, 25, 93–99. [Google Scholar] [CrossRef]
- Peron, G.; Hidalgo-Liberona, N.; González-Domínguez, R.; Garcia-Aloy, M.; Guglielmetti, S.; Bernardi, S.; Andrés-Lacueva, C. Exploring the molecular pathways behind the effects of nutrients and dietary polyphenols on gut microbiota and intestinal permeability: A perspective on the potential of metabolomics and future clinical applications. J. Agric. Food Chem. 2019, 68, 1780–1789. [Google Scholar] [CrossRef]
- Yang, G.; Bibi, S.; Du, M.; Suzuki, T.; Zhu, M.J. Regulation of the intestinal tight junction by natural polyphenols: A mechanistic perspective. Crit. Rev. Food Sci. Nutr. 2017, 57, 3830–3839. [Google Scholar] [CrossRef]
- Bernardi, S.; Del Bo’, C.; Marino, M.; Gargari, G.; Cherubini, A.; Andrés-Lacueva, C.; Riso, P. Polyphenols and intestinal permeability: Rationale and future perspectives. J. Agric. Food Chem. 2019, 68, 1816–1829. [Google Scholar] [CrossRef]
- Marino, M.; Venturi, S.; Rendine, M.; Porrini, M.; Gardana, C.; Klimis-Zacas, D.; Riso, P. Wild blueberry (V. angustifolium) improves TNFα-induced cell barrier permeability through claudin-1 and oxidative stress modulation in Caco-2 cells. Food Funct. 2023, 14, 7387–7399. [Google Scholar] [CrossRef]
- Ishimoto, K.; Konishi, Y.; Otani, S.; Maeda, S.; Ago, Y.; Hino, N.; Nakagawa, S. Suppressive effect of black tea polyphenol theaflavins in a mouse model of ovalbumin-induced food allergy. J. Nat. Med. 2023, 77, 604–609. [Google Scholar] [CrossRef]
- Yun, Z.; Zou, Z.; Sun, S.; Che, H. Chlorogenic acid improves food allergy through the AMPK/ACC/CPT-1 pathway. J. Food Biochem. 2022, 46, e14505. [Google Scholar] [CrossRef]
- Ma, Y.; Liu, M.; Li, D.; Li, J.; Guo, Z.; Liu, Y.; Liu, Y. Olive oil ameliorates allergic response in murine ovalbumin-induced food allergy by promoting intestinal mucosal immunity. Food Sci. Hum. Wellness 2023, 12, 801–808. [Google Scholar] [CrossRef]
- Dufourny, S.; Antoine, N.; Pitchugina, E.; Delcenserie, V.; Godbout, S.; Douny, C.; Scippo, M.-L.; Froidmont, E.; Rondia, P.; Wavreille, J.; et al. Apple Pomace and Performance, Intestinal Morphology and Microbiota of Weaned Piglets—A Weaning Strategy for Gut Health? Microorganisms 2021, 9, 572. [Google Scholar] [CrossRef] [PubMed]
- Jafarinia, M.; Sadat Hosseini, M.; Kasiri, N.; Fazel, N.; Fathi, F.; Ganjalikhani Hakemi, M.; Eskandari, N. Quercetin with the potential effect on allergic diseases. Allergy Asthma Clin. Immunol. 2022, 16, 36. [Google Scholar] [CrossRef] [PubMed]
- Sagit, M.; Polat, H.; Gurgen, S.G.; Berk, E.; Guler, S.; Yasar, M. Effectiveness of quercetin in an experimental rat model of allergic rhinitis. Eur. Arch. Oto-Rhino-Laryngol. 2017, 274, 3087–3095. [Google Scholar] [CrossRef] [PubMed]
- Kashiwabara, M.; Asano, K.; Mizuyoshi, T.; Kobayashi, H. Suppression of neuropeptide production by quercetin in allergic rhinitis model rats. BMC Complement. Altern. Med. 2016, 16, 132. [Google Scholar] [CrossRef]
- Edo, Y.; Otaki, A.; Asano, K. Quercetin enhances the thioredoxin production of nasal epithelial cells in vitro and in vivo. Medicines 2018, 5, 124. [Google Scholar] [CrossRef]
- Ebihara, N.; Takahashi, K.; Takemura, H.; Akanuma, Y.; Asano, K.; Sunagawa, M. Suppressive effect of quercetin on nitric oxide production from nasal epithelial cells in vitro. Evid.-Based Complement. Altern. Med. 2018, 2018, 6097625. [Google Scholar] [CrossRef]
- Otaki, A.; Furuta, A.; Asano, K. Quercetin-Induced Enhancement of Nasal Epithelial Cells’ Ability to Produce Clara Cell 10-kD Protein In Vitro and In Vivo. Medicines 2023, 10, 28. [Google Scholar] [CrossRef]
- Seo, M.Y.; Kim, K.R.; Lee, J.J.; Ryu, G.; Lee, S.H.; Hong, S.D.; Kim, H.Y. Therapeutic effect of topical administration of red onion extract in a murine model of allergic rhinitis. Sci. Rep. 2019, 9, 2883. [Google Scholar] [CrossRef]
- Arpornchayanon, W.; Klinprung, S.; Chansakaow, S.; Hanprasertpong, N.; Chaiyasate, S.; Tokuda, M.; Tamura, H. Antiallergic activities of shallot (Allium ascalonicum L.) and its therapeutic effects in allergic rhinitis. Asian Pac. J. Allergy Immunol. 2022, 40, 393–400. [Google Scholar] [CrossRef]
- Almeida Brasiel, P.G.; Guimarães, F.V.; Rodrigues, P.M.; Bou-Habib, D.C.; Carvalho, V.F. Therapeutic Efficacy of Flavonoids in Allergies: A Systematic Review of Randomized Controlled Trials. J. Immunol. Res. 2022, 2022, 8191253. [Google Scholar] [CrossRef]
- Kim, H.J.; Woo, J.; Nam, Y.R.; Seo, Y.; Namkung, W.; Nam, J.H.; Kim, W.K. Luteolin reduces fluid hypersecretion by inhibiting TMEM16A in interleukin-4 treated Calu-3 airway epithelial cells. Korean J. Physiol. Pharmacol. 2020, 24, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Liang, K.-L.; Yu, S.-J.; Huang, W.-C.; Yen, H.-R. Luteolin Attenuates Allergic Nasal Inflammation via Inhibition of Interleukin-4 in an Allergic Rhinitis Mouse Model and Peripheral Blood from Human Subjects with Allergic Rhinitis. Front. Pharmacol. 2020, 11, 291. [Google Scholar] [CrossRef] [PubMed]
- Şahin, A.; Sakat, M.S.; Kılıç, K.; Aktan, B.; Yildirim, S.; Kandemir, F.M.; Kucukler, S. The protective effect of naringenin against ovalbumin-induced allergic rhinitis in rats. Eur. Arch. Oto-Rhino-Laryngol. 2021, 278, 4839–4846. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Chen, G.; Shu, S.; Xu, Y.; Ma, X. Metabolomics analysis of baicalin on ovalbumin-sensitized allergic rhinitis rats. R. Soc. Open Sci. 2019, 6, 181081. [Google Scholar] [CrossRef]
- Pellow, J.; Nolte, A.; Temane, A.; Solomon, E.M. Health supplements for allergic rhinitis: A mixed-methods systematic review. Complement. Ther. Med. 2020, 51, 102425. [Google Scholar] [CrossRef]
- Tanaka, T.; Iuchi, A.; Harada, H.; Hashimoto, S. Potential beneficial effects of wine flavonoids on allergic diseases. Diseases 2019, 7, 8. [Google Scholar] [CrossRef]
- Sugiura, Y.; Usui, M.; Miyata, M. The soothing effect of phlorotannins on cedar pollinosis in Cry j 1-stimulated mice. Biosci. Biotechnol. Biochem. 2023, 87, 649–652. [Google Scholar] [CrossRef]
- Yoshimura, M.; Enomoto, T.; Dake, Y.; Okuno, Y.; Ikeda, H.; Cheng, L.; Obata, A. An evaluation of the clinical efficacy of tomato extract for perennial allergic rhinitis. Allergol. Int. 2007, 56, 225–230. [Google Scholar] [CrossRef]
- Bakhshaee, M.; Jabbari, F.; Hoseini, S.; Farid, R.; Sadeghian, M.H.; Rajati, M.; Zamani, M.A. Effect of silymarin in the treatment of allergic rhinitis. Otolaryngol. Head Neck Surg. 2011, 145, 904–909. [Google Scholar] [CrossRef]
- Wilson, D.; Evans, M.; Guthrie, N.; Sharma, P.; Baisley, J.; Schonlau, F.; Burki, C. A randomized, double-blind, placebo-controlled exploratory study to evaluate the potential of pycnogenol® for improving allergic rhinitis symptoms. Phytother. Res. 2010, 24, 1115–1119. [Google Scholar] [CrossRef] [PubMed]
- Marseglia, G.; Licari, A.; Leonardi, S.; Papale, M.; Zicari, A.M.; Schiavi, L.; Italian Study Group on Pediatric Allergic Rhinoconjunctivitis; Cardinale, F.; Cherubini, S.; Giordano, P.; et al. A polycentric, randomized, parallel-group, study on Lertal®, a multicomponent nutraceutical, as preventive treatment in children with allergic rhinoconjunctivitis: Phase II. Ital. J. Pediatr. 2019, 45, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Wruss, J.; Lanzerstorfer, P.; Huemer, S.; Himmelsbach, M.; Mangge, H.; Höglinger, O.; Weghuber, J. Differences in pharmacokinetics of apple polyphenols after standardized oral consumption of unprocessed apple juice. Nutr. J. 2015, 14, 32. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, B.; Luo, Y.; Zhang, Q.; Bian, Y.; Wang, R. Resveratrol-mediated SIRT1 activation attenuates ovalbumin-induced allergic rhinitis in mice. Mol. Immunol. 2020, 122, 156–162. [Google Scholar] [CrossRef]
- Zhang, W.; Tang, R.; Ba, G.; Li, M.; Lin, H. Anti-allergic and anti-inflammatory effects of resveratrol via inhibiting TXNIP-oxidative stress pathway in a mouse model of allergic rhinitis. World Allergy Organ. J. 2020, 13, 100473. [Google Scholar] [CrossRef]
- Miraglia Del Giudice, M.; Maiello, N.; Capristo, C.; Alterio, E.; Capasso, M.; Perrone, L.; Ciprandi, G. Resveratrol plus carboxymethyl-β-glucan reduces nasal symptoms in children with pollen-induced allergic rhinitis. Curr. Med. Res. Opin. 2014, 30, 1931–1935. [Google Scholar] [CrossRef]
- Lv, C.; Zhang, Y.; Shen, L. Preliminary clinical effect evaluation of resveratrol in adults with allergic rhinitis. Int. Arch. Allergy Immunol. 2018, 175, 231–236. [Google Scholar] [CrossRef]
- Fu, M.; Fu, S.; Ni, S.; Wang, D.; Hong, T. Inhibitory effects of bisdemethoxycurcumin on mast cell-mediated allergic diseases. Int. Immunopharmacol. 2018, 65, 182–189. [Google Scholar] [CrossRef]
- Wu, S.; Xiao, D. Effect of curcumin on nasal symptoms and airflow in patients with perennial allergic rhinitis. Ann. Allergy Asthma Immunol. 2016, 117, 697–702. [Google Scholar] [CrossRef]
- Ceylan, E.; Turgut Cosan, D.; Bayar Muluk, N.; Cingi, C. Investigation of the effect of the curcumin component as an alternative to the local treatment of nasal diseases. Eur. Rev. Med. Pharmacol. Sci. 2023, 27, 39–43. [Google Scholar] [CrossRef]
- Leis, K.; Gałązka, P.; Kazik, J.; Jamrożek, T.; Bereźnicka, W.; Czajkowski, R. Resveratrol in the treatment of asthma based on an animal model. Adv. Dermatol. Allergol. 2022, 39, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Ni, Z.H.; Tang, J.H.; Chen, G.; Lai, Y.M.; Chen, Q.G.; Li, Z.; Wang, X.B. Resveratrol inhibits mucus overproduction and MUC5AC expression in a murine model of asthma. Mol. Med. Rep. 2016, 13, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Li, X.N.; Ma, L.Y.; Ji, H.; Qin, Y.H.; Jin, S.S.; Xu, L.X. Resveratrol protects against oxidative stress by activating the Keap-1/Nrf2 antioxidant defense system in obese-asthmatic rats. Exp. Ther. Med. 2018, 16, 4339–4348. [Google Scholar] [CrossRef]
- Lee, H.Y.; Kim, I.K.; Yoon, H.K.; Kwon, S.S.; Rhee, C.K.; Lee, S.Y. Inhibitory effects of resveratrol on airway remodeling by transforming growth factor-β/Smad signaling pathway in chronic asthma model. Allergy Asthma Immunol. Res. 2017, 9, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Andre, D.M.; Calixto, M.C.; Sollon, C.; Alexandre, E.C.; Leiria, L.O.; Tobar, N.; Antunes, E. Therapy with resveratrol attenuates obesity-associated allergic airway inflammation in mice. Int. Immunopharmacol. 2016, 38, 298–305. [Google Scholar] [CrossRef]
- Zhang, Y.U.N.; Guo, L.; Law, B.Y.K.; Liang, X.; Ma, N.; Xu, G.; Wang, X. Resveratrol decreases cell apoptosis through inhibiting DNA damage in bronchial epithelial cells. Int. J. Mol. Med. 2020, 45, 1673–1684. [Google Scholar] [CrossRef]
- Lelli, D.; Sahebkar, A.; Johnston, T.P.; Pedone, C. Curcumin use in pulmonary diseases: State of the art and future perspectives. Pharmacol. Res. 2017, 115, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Chong, L.; Zhang, W.; Nie, Y.; Yu, G.; Liu, L.; Lin, L.; Li, C. Protective effect of curcumin on acute airway inflammation of allergic asthma in mice through Notch1–GATA3 signaling pathway. Inflammation 2014, 37, 1476–1485. [Google Scholar] [CrossRef]
- He, Y.; Yue, Y.; Zheng, X.; Zhang, K.; Chen, S.; Du, Z. Curcumin, inflammation, and chronic diseases: How are they linked? Molecules 2015, 20, 9183–9213. [Google Scholar] [CrossRef]
- Abidi, A.; Gupta, S.; Agarwal, M.; Bhalla, H.L.; Saluja, M. Evaluation of efficacy of curcumin as an add-on therapy in patients of bronchial asthma. J. Clin. Diagn. Res. JCDR 2014, 8, HC19. [Google Scholar] [CrossRef]
- Manarin, G.; Anderson, D.; e Silva, J.M.; da Silva Coppede, J.; Roxo-Junior, P.; Pereira, A.M.S.; Carmona, F. Curcuma longa L. ameliorates asthma control in children and adolescents: A randomized, double-blind, controlled trial. J. Ethnopharmacol. 2019, 238, 111882. [Google Scholar] [CrossRef] [PubMed]
- Tenero, L.; Piazza, M.; Zanoni, L.; Bodini, A.; Peroni, D.; Piacentini, G.L. Antioxidant supplementation and exhaled nitric oxide in children with asthma. Allergy Asthma Proc. 2016, 37, 13. [Google Scholar] [CrossRef] [PubMed]
- Quan, M.; Alismail, A.; Daher, N.; Cleland, D.; Chavan, S.; Tan, L.D. Randomized, placebo controlled, double blinded pilot superiority phase 2 trial to evaluate the effect of curcumin in moderate to severe asthmatics. BMC Pulm. Med. 2021, 21, 268. [Google Scholar] [CrossRef] [PubMed]
- Jang, T.Y.; Jung, A.Y.; Kyung, T.S.; Kim, D.Y.; Hwang, J.H.; Kim, Y.H. Anti-allergic effect of luteolin in mice with allergic asthma and rhinitis. Cent. Eur. J. Immunol. 2017, 42, 24–29. [Google Scholar] [CrossRef]
- Shin, N.R.; Ryu, H.W.; Ko, J.W.; Park, S.H.; Yuk, H.J.; Kim, H.J.; Shin, I.S. Artemisia argyi attenuates airway inflammation in ovalbumin-induced asthmatic animals. J. Ethnopharmacol. 2017, 209, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Shan, L.; Kang, X.; Liu, F.; Cai, X.; Han, X.; Shang, Y. Epigallocatechin gallate improves airway inflammation through TGF-β1 signaling pathway in asthmatic mice. Mol. Med. Rep. 2018, 18, 2088–2096. [Google Scholar] [CrossRef]
- Li, Q.; Wu, Y.; Guo, X.; Wang, R.; Mao, Y.; Zhao, Y.; Yang, Y. Protocatechuic acid supplement alleviates allergic airway inflammation by inhibiting the IL-4Rα–STAT6 and Jagged 1/Jagged2–Notch1/Notch2 pathways in allergic asthmatic mice. Inflamm. Res. 2020, 69, 1027–1037. [Google Scholar] [CrossRef]
- Wang, S.; Wuniqiemu, T.; Tang, W.; Teng, F.; Bian, Q.; Yi, L.; Dong, J. Luteolin inhibits autophagy in allergic asthma by activating PI3K/Akt/mTOR signaling and inhibiting Beclin-1-PI3KC3 complex. Int. Immunopharmacol. 2021, 94, 107460. [Google Scholar] [CrossRef]
- Hosawi, S. Current Update on Role of Hesperidin in Inflammatory Lung Diseases: Chemistry, Pharmacology, and Drug Delivery Approaches. Life 2023, 13, 937. [Google Scholar] [CrossRef]
- Muhammad, H.; Salahuddin, Z.; Akhtar, T.; Aftab, U.; Rafi, A.; Hussain, S.; Shahzad, M. Immunomodulatory effect of glabridin in ovalbumin induced allergic asthma and its comparison with methylprednisolone in a preclinical rodent model. J. Cell. Biochem. 2023, 124, 1503–1515. [Google Scholar] [CrossRef]
- Yang, N.; Shang, Y.X. Epigallocatechin gallate ameliorates airway inflammation by regulating Treg/Th17 imbalance in an asthmatic mouse model. Int. Immunopharmacol. 2019, 72, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Li, X. Epigallocatechin gallate relieves asthmatic symptoms in mice by suppressing HIF-1α/VEGFA-mediated M2 skewing of macrophages. Biochem. Pharmacol. 2022, 202, 115112. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.H.; Cho, I.H.; Shin, D.; Han, S.Y.; Park, S.H.; Kang, Y.H. Inhibition of airway epithelial-to-mesenchymal transition and fibrosis by kaempferol in endotoxin-induced epithelial cells and ovalbumin-sensitized mice. Lab. Investig. 2014, 94, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Liu, R.; Shang, Y.; Qin, Q.; Ge, Y. In vitro and in vivo relaxation and anti-inflammation of natural flavonoids from Elaeagnus pungens leaf via L-type calcium channel and targeting MAPK signal pathway. J. Ethnopharmacol. 2021, 275, 114071. [Google Scholar] [CrossRef] [PubMed]
- Belal, S.A.; Choe, H.S.; Shin, D.K.; Shim, K.S. Effect of kaempferol on cyclooxygenase 2 (Cox2) and cytosolic phospholipase A2 (cPLA2) protein expression in BALB/c mice. Iran. J. Allergy Asthma Immunol. 2018, 17, 428–435. [Google Scholar] [CrossRef]
- Kianmehr, M.; Khazdair, M.R. Possible therapeutic effects of Crocus sativus stigma and its petal flavonoid, kaempferol, on respiratory disorders. Pharm. Biol. 2020, 58, 1140–1149. [Google Scholar] [CrossRef]
- Mokra, D.; Adamcakova, J.; Mokry, J. Green Tea Polyphenol (-)-Epigallocatechin-3-Gallate (EGCG): A Time for a New Player in the Treatment of Respiratory Diseases? Antioxidants 2022, 11, 1566. [Google Scholar] [CrossRef]
- Zhu, S.; Wang, H.; Zhang, J.; Yu, C.; Liu, C.; Sun, H.; Wu, Y.; Wang, Y.; Lin, X. Antiasthmatic activity of quercetin glycosides in neonatal asthmatic rats. 3 Biotech 2019, 9, 189. [Google Scholar] [CrossRef]
- Belcaro, G.; Luzzi, R.; Cesinaro Di Rocco, P.; Cesarone, M.R.; Dugall, M.; Feragalli, B.; Errichi, B.M.; Ippolito, E.; Grossi, M.G.; Hosoi, M.; et al. Pycnogenol® improvements in asthma management. Panminerva Medica 2011, 53 (Suppl. 1), 57–64. [Google Scholar]
- Robertson, N.U.; Schoonees, A.; Brand, A.; Visser, J. Pine bark (Pinus spp.) extract for treating chronic disorders. Cochrane Database Syst. Rev. 2020, 9, CD008294. [Google Scholar] [CrossRef]
- Shaw, O.M.; Hurst, R.D.; Cooney, J.; Sawyer, G.M.; Dinnan, H.; Martell, S. Boysenberry and apple juice concentrate reduced acute lung inflammation and increased M2 macrophage-associated cytokines in an acute mouse model of allergic airways disease. Food Sci. Nutr. 2021, 9, 1491–1503. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.Y.; Kim, J.W.; Kim, J.H.; Jeong, J.S.; Lim, J.O.; Ko, J.W.; Kim, T.W. Inner shell of the chestnut (Castanea crenatta) suppresses inflammatory responses in ovalbumin-induced allergic asthma mouse model. Nutrients 2022, 14, 2067. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Han, L.; Lv, R.; Ling, L. Magnolol exerts anti-asthmatic effects by regulating Janus kinase-signal transduction and activation of transcription and Notch signaling pathways and modulating Th1/Th2/Th17 cytokines in ovalbumin-sensitized asthmatic mice. Korean J. Physiol. Pharmacol. 2019, 23, 251–261. [Google Scholar] [CrossRef]
- Wang, J.; Xian, M.; Cao, H.; Wu, L.; Zhou, L.; Ma, Y.; Xiao, X. Prophylactic and therapeutic potential of magnolol-loaded PLGA-PEG nanoparticles in a chronic murine model of allergic asthma. Front. Bioeng. Biotechnol. 2023, 11, 1182080. [Google Scholar] [CrossRef]
- Yu, L.; Li, J. Punicalagin attenuated allergic airway inflammation via regulating IL4/IL-4Rα/STAT6 and Notch-GATA3 pathways. Acta Pharm. 2022, 72, 561–573. [Google Scholar] [CrossRef]
- Danesi, F.; Ferguson, L.R. Could pomegranate juice help in the control of inflammatory diseases? Nutrients 2017, 9, 958. [Google Scholar] [CrossRef]
- Hosseini, S.A.; Shateri, Z.; Abolnezhadian, F.; Maraghi, E.; Shoushtari, M.H.; Zilaee, M. Does pomegranate extract supplementation improve the clinical symptoms of patients with allergic asthma? A double-blind, randomized, placebo-controlled trial. Front. Pharmacol. 2023, 14, 1109966. [Google Scholar] [CrossRef]
- Lai, Y.T.; He, D.; Li, B.; Lund, M.; Xing, Y.F.; Wang, Y.; Cao, X. Engineering polyphenols with biological functions via polyphenol-protein interactions as additives for functional foods. Trends Food Sci. Technol. 2021, 110, 470–482. [Google Scholar] [CrossRef]
- Chen, L.; Cao, H.; Xiao, J. Polyphenols: Absorption, bioavailability, and metabolomics. In Polyphenols: Properties, Recovery, and Applications; Woodhead Publishing: Cambridge, UK, 2018; pp. 45–67. [Google Scholar] [CrossRef]
- Williamson, G.; Kay, C.D.; Crozier, A. The bioavailability, transport, and bioactivity of dietary flavonoids: A review from a historical perspective. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1054–1112. [Google Scholar] [CrossRef]
- Gorantla, S.; Wadhwa, G.; Jain, S.; Sankar, S.; Nuwal, K.; Mahmood, A.; Singhvi, G. Recent advances in nanocarriers for nutrient delivery. Drug Deliv. Transl. Res. 2021, 12, 2359–2384. [Google Scholar] [CrossRef]
- Grgić, J.; Šelo, G.; Planinić, M.; Tišma, M.; Bucić-Kojić, A. Role of the encapsulation in bioavailability of phenolic compounds. Antioxidants 2020, 9, 923. [Google Scholar] [CrossRef] [PubMed]
- Hadidi, M.; Jafarzadeh, S.; Forough, M.; Garavand, F.; Alizadeh, S.; Salehabadi, A.; Jafari, S.M. Plant protein-based food packaging films; recent advances in fabrication, characterization, and applications. Trends Food Sci. Technol. 2020, 120, 154–173. [Google Scholar] [CrossRef]
- Liu, Q.; Lin, S.; Sun, N. How does food matrix components affect food allergies, food allergens and the detection of food allergens? A systematic review. Trends Food Sci. Technol. 2020, 127, 280–290. [Google Scholar] [CrossRef]
- Iglesias-Aguirre, C.E.; Cortés-Martín, A.; Ávila-Gálvez, M.Á.; Giménez-Bastida, J.A.; Selma, M.V.; González-Sarrías, A.; Espín, J.C. Main drivers of (poly) phenol effects on human health: Metabolite production and/or gut microbiota-associated metabotypes? Food Funct. 2021, 12, 10324–10355. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Polyphenols Classes | Type | Dietary Source | Reference |
|---|---|---|---|
| Flavonoids | |||
| Flavonols | Quercetin Kaempferol Myricetin | apples, cherries, berries, apricots, cranberries, grapes, mango peel, onions, kale, tomatoes, broccoli, fennel, capers, okra, rocket, tea, red wine, beer, cocoa, bee pollen | [30,40,43,45] |
| Flavones | Luteolin Apigenin Baicalein | lemon, tangerine, and orange peel and pulp parsley, green pepper, celery, artichoke, lettuce watermelon, melon, cantaloupe, apples green and black tea | [39,45,46] |
| Isoflavones | Genistein Daidzein | legumes such as soybean green peas and black beans | [30,42,47] |
| Flavanones | Hesperidin Naringenin | citrus peel, tomato peels, seeds, spices aromatic plants: mint, chamomile | [38,48] |
| Flavanols | Catechin Epicatechin Gallocatechin Epigallocatechin | peels of grapes peels of apples grapes, seeds, roasted peanuts, almonds, pistachios green tea leaves chocolate, red wine, rosemary | [30,38,41,44] |
| Anthocyanidins | Cyanidin | grape skin, wine lees grape pomace blueberries, banana, strawberries, cherries, pears, cranberries, plums, beans, red cabbage | [38,39,49] |
| Non-flavonoids | |||
| Phenolic acids | Gallic acid Ferulic acid Caffeic acid Curcumin | onions, black radishes, red fruits, citrus peels grapes (seed and skin) potato peel tea and fruit tea, coffee | [34,43] |
| Stilbenes | Resveratrol | grape by-products, red and white wine, berry fruits, strawberries | [44,50] |
| Lignans | grains and cereals: oat, wheat, rye, barley strawberries, apricots cabbage, broccoli, garlic olive | [38,50] |
| Polyphenol | Polyphenol Dose | Study Type | Results/Observations | Side Effects | Ref. |
|---|---|---|---|---|---|
| Epigallocatechin, Epigallocatechin gallate | 50 mg/day extracted from tea | In vivo BALB/c mice model of αs1-casein milk protein allergy | Significantly reduced levels of mast cell protease, histamine, specific IgE antibodies, and Th2 cytokines Reduced degree of pathological changes in the intestine | No data available | [218] |
| Ellagitannins Gallic acid | 0.1%; 0.5%; 1.0% tea leaf extract: gallic acid 1659.0 mg/100 g dry weight ellagic acid 4622.7 mg/100 g dry weight | In vivo BALB/c mice model of egg allergy | Reduction in symptoms such as scratching, lethargy, and gastrointestinal signs Modulating the Th1/Th2 balance Increased percentage of the Treg subtype Enhancing intestinal IgA secretions | No data available | [133] |
| Curcumin | 3 mg, 30 mg/kg Curcuma longa extract | In vivo BALB/c mice food allergy model | Reduction in food allergy symptoms such as decreased rectal temperature and anaphylactic response Inhibited IgE, reduced Th2-related cytokines, and enhanced Th1-related cytokine Maintaining Th1/Th2 balance | No data available | [219] |
| Resveratrol | 2.5–40 μg/mL 5, 10, 20 mg Abies georgei extract | In vitro RBL-2H3 cells In vivo BALB/c mice | Reduced mast cell degranulation and release of β-hexosaminidase and histamine Suppression in the development of diarrhea upregulates the rectal temperature Decreased serum level of specific IgE, mouse mast cell protease-1, and histamine | No cytotoxic effect | [220] |
| Catechins | 0.05% 0.1% areca nut extract via drinking water | In vivo BALB/c mice | Attenuated OVA-induced allergic responses, including diarrhea Reduced infiltration and degranulation of mast cells in the duodenum Suppressed specific IgE production and Th2 immune response | No data available | [222] |
| Flavonoids | 100 g/kg cocoa beans powder | In vivo Brown Norway rats | Suppressed synthesis of specific IgE Suppressed Th2-related cytokines released from mesenteric lymph node and spleen cells | No data available | [223] |
| Baicalin | 50, 100, 200 μ mol/L 20 mg/kg Scutellaria baicalensis extract | In vitro Caco-2 cells In vivo BALB/c mice food allergy model | Reduction in food allergy symptoms, serum IgE, and effector Th2 cells Up-regulation of Treg Enhancement of intestinal barrier function through the regulation of tight junctions | No data available | [224] |
| Anthocyanidins | 1 and 5 mg/mL wild blueberry extract | In vitro Caco-2 cells | Enhancement of intestinal barrier function and integrity of the intestinal cell monolayer Reduced intestinal permeability, increased TEER, upregulation of claudin-1 | No data available | [232] |
| Theaflavins | 0.02–0.20% black tea theaflavin mixture via food powder | In vivo BALB/c mice | Reduction in food allergy symptoms: the severity of diarrhea Alleviating oxidative stress | No data available | [233] |
| Chlorogenic acid | 50, 200 mg/kg pure isolated polyphenol | In vivo BALB/c mice model of shrimp allergy | Reduction in food allergy symptoms Decreased IgE level Regulation of AMPK/ACC/CPT-1 signaling pathway | No data available | [234] |
| Ferulic acid Caffeic acid Apigenin Luteolin | 1–3 g/kg/day olive oil | In vivo BALB/c mice | Repaired ileum villi, and upregulated tight junction protein expression Increased levels of Treg-related cytokines (IL-10) in lamina propria Decreased levels of Th2 cell-associated cytokines in lamina propria Reduced Burkholderiaceae and increased Clostridiaceae in the intestinal microflora | No data available | [226] |
| 600 mg/kg/day olive oil | In vivo BALB/c mice | Reduction in food allergy symptoms decreased the IgE level, increased expression of intestinal tight junction proteins (Claudin-1, Occludin), increased levels of mucin 2 and β-defensin | No data available | [235] |
| Polyphenol | Polyphenols Dose | Study Type | Results/Observations | Side Effects | Ref. |
|---|---|---|---|---|---|
| Quercetin | 1, 10 and 50 mg/kg, p.o. pure isolated polyphenol | In vivo BALB/c mice | Decreased sneezing, nasal rubbing, and nasal redness frequency Decreased level of NO, decreased IgE and Th2-cytokine production | No data available | [135] |
| 20, 35, or 50 mg/kg/day pure isolated polyphenol | In vivo BALB/c mice | Reduced rubbing and sneezing Reduced IgE, histamine in serum Decreased number of inflammation cells and goblet cells in tissues Inhibited Th1/Th2 imbalance and Treg/Th17 imbalance | No data available | [164] | |
| 80 mg/kg pure isolated polyphenol | In vivo Sprague–Dawley rats | Decreased secretion, sneezing, and itching Decreased IgE and Th2-cytokine production Decreased eosinophil count in the mucosa of the nasal turbinate | No data available | [238] | |
| 25 mg/kg pure isolated polyphenol | In vivo Sprague–Dawley rats | Inhibited nasal rubbing movements and sneezing | No data available | [239] | |
| 20 mg/kg pure isolated polyphenol | In vitro HNEpC In vivo BALB/c mice | Inhibited nasal symptoms and increased TRX levels in nasal lavage fluids | No data available | [240] | |
| 100.0 pM, 1.0 nM, 10.0 nM, 100.0 nM pure isolated polyphenol | In vitro HNEpC | Reduced NO production Downregulated Th2 cytokine responses | No data available | [241] | |
| 20, 40 μL red onion extract | In vivo BALB/c mice | Reduced allergic rhinitis symptom Decreased levels of IL-4, IL-5, IL-10, IL-13 Reduced eosinophil infiltration of nasal turbinate | No data available | [243] | |
| 3 g/day shallot oral supplement | Clinical study | Improved symptoms such as sneezing, rhinorrhea, itchy nose, and eyes | No side effects, well-tolerated | [244] | |
| 100 mg/day pure isolated polyphenol | Clinical study | Improved nasal and ocular symptoms Prevention of the development of symptoms | No side effects | [245] | |
| Luteolin | 10, 30 mg/kg pure isolated polyphenol | In vitro PBMC In vivo BALB/c mice | Decreased allergic symptoms and serum HDM-specific IgE Inhibition of IL-4 production | No side effects | [247] |
| 10 mg/kg pure isolated polyphenol | In vivo BALB/c mice | Decreased nasal sneezing frequency, nasal mucosa thickness, and levels of specific-IgE and IL-17A, increased IL-10 and Foxp3 expression, suppressed Treg/Th 17 imbalance | No side effects | [169] | |
| Myricetin | 50, 100, 200 mg/kg pure isolated polyphenol | In vivo BALB/c mice | Protected against histamine challenge, decreased serum level of total and specific-IgE Inhibition of mast cell degranulation, regulation of Th1/Th2 balance | No data available | [165] |
| Naringenin | 100 mg/kg pure isolated polyphenol | In vivo Sprague–Dawley rats | Decreased level of serum total IgE, IL4 and IL5 Reduced desquamation, erosion, and eosinophilic cell infiltration in nasal mucosa | No data available | [248] |
| 360 mg per day of tomato extract | Clinical study | Significantly decreased sneezing score, rhinorrhea, and nasal obstruction Improved patients’ quality of life | No side effects | [253] | |
| Baicalin | 100 μg/mL pure isolated polyphenol | In vitro PBMC In vivo BALB/c mice | Restored Th17/Treg cell balance Reduced infiltration of inflammatory cells of the nasal lavage fluid, improved nasal mucosal thickness and mucus secretion | No side effects mild laxative effect | [249] |
| Silymarin (silibinin, silydianine, silychristine) | 140 mg 3 times daily mixture extracted from milk thistle Silybum marianum | Clinical study | Significant improvement in clinical symptom severity | No side effects | [254] |
| Pycnogenol (procyanidins, catechins) | 50 mg French maritime pine bark extract | Clinical study | Reduced symptoms of allergic rhinitis in patients allergic to birch pollen | No side effects | [255] |
| Lertal (mixture of quercetin, rosmarinic, luteolin, apigenin) | 150 mg quercetin, 80 mg Perilla frutescens extract | Clinical study | Reduction in allergic rhinitis symptoms and the need to use symptomatic medications | No data available | [256] |
| Procyanidins Catechin Epicatechin | 50, 200, 500 mg/day apple polyphenols extract | Clinical study | Reduced nasal symptoms including sneezing, rhinorrhea, and swelling of the nasal turbinates | No data available | [250] |
| Resveratrol | 5, 30, 50 mg/kg pure isolated polyphenol | In vivo BALB/c mice | Decreased levels of histamine, specific-IgE, IL-4, IL-5, IL-13, IL-17, and inflammatory cell numbers (leucocytes, eosinophils, lymphocytes, and neutrophils) | No data available | [258,259] |
| 100 μL Polygonum cuspidatum extract | Clinical study | Significant reduction in nasal symptoms: itching, sneezing, rhinorrhea, and obstruction as well as the need to use antihistamine | No data available | [260] | |
| 100 μL Polygonum cuspidatum extract | Clinical study | Significant reduction in nasal symptoms decreased IgE, IL-4, and eosinophil levels in the blood, improved the patient’s quality of life | No data available | [261] | |
| Curcumin | 500 mg/d pure isolated polyphenol | Clinical study | Significant reduction in nasal symptoms (sneezing, itching, rhinorrhea), and increase the nasal airflow, suppression of IL-4, IL-8, IL-10 | No side effects | [263] |
| Polyphenol | Polyphenols Dose | Study Type | Results/Observations | Side Effects | Ref. |
|---|---|---|---|---|---|
| Resveratrol | 30 mg/day pure isolated polyphenol | In vivo BALB/c mice | Inhibited OVA-induced airway inflammation and mucus production | No data available | [266] |
| 10, 50 mg/kg/day pure isolated polyphenol | In vivo BALB/c mice | Reduction in inflammation, inhibition of respiratory tract remodelling progression Reduced collagen production Decreased IL-4, IL-5, IL-13,TGF-β1 and eosinophil level | No data available | [268] | |
| 100 mg/kg pure isolated polyphenol | In vivo C57/Bl16 mice | Reduced inflammation and eosinophil infiltration | No data available | [269] | |
| 100 mg/kg pure isolated polyphenol | In vivo C57BL/6J mice | Prevention of oxidative DNA damage and apoptosis in bronchial epithelial cells exposed to HDM allergen | No data available | [270] | |
| Curcumin | 10, 20 mg/kg pure isolated polyphenol | In vivo BALB/c mice | Significant decrease in airway inflammation and oxidative stress Treg cell stimulation | No data available | [158] |
| 120 mg/kg pure isolated polyphenol | In vivo BALB/c mice | Reduced cytokine production (IL-4, IL-5, IL-13) Suppression in tissue eosinophilia and mucus hyperproduction | No data available | [173] | |
| 200 mg/kg pure isolated polyphenol | In vivo BALB/c mice | Reduced total cell influx and number of lymphocytes, eosinophils, and neutrophils in BALF Reduction in airway inflammation | No data available | [272] | |
| 800 mg pure isolated polyphenol | In vivo BALB/c mice | Alleviation of lung inflammation Significantly reduced number of eosinophils and the hyperproduction of goblet cells Decreased Th2-related cytokines IL-4, IL-5, and IL-13 and Th17 cytokine IL-17A production | No data available | [171] | |
| 1000 mg/per day of pure isolated polyphenol | Clinical study adults | Significant improvement in the mean FEV1 values | No side effects | [274] | |
| 30 mg/kg/day roots of Curcuma longa | Clinical study children | Improved disease control: less frequent nighttime awakenings, less frequent use of short-acting β-adrenergic agonists | No side effects | [275] | |
| Luteolin | 0.1 mg/kg pure isolated polyphenol | In vivo BALB/c mice | Significant decrease in IL-4, IL-5, and IL-13 in their lung homogenate and in inflammatory cell infiltration in lung tissue | No data available | [278] |
| 50, 100 mg/kg Artemisia argyi extract | In vivo BALB/c mice | Reduced inflammatory cell counts, Th2 cytokines, airway hyperresponsiveness and mucus hypersecretion | No data available | [279] | |
| Glabridin | 40 mg/kg Glycyrrhiza glabra (licorice) roots extract | In vivo Wistar rats | Decreased serum IgE levels and the expression of TNF-α, IL-4, IL-5 Decreased inflammatory cells in the blood and BALF | No cytotoxic effect | [292] |
| Epigallocatechin gallate | 20 mg/kg green tea extract | In vivo BALB/c mice | Reduced asthmatic symptoms, lung inflammatory cell infiltration, level of inflammatory factors, and increased the percentage of Treg | No data available | [280] |
| 5, 50 mg/kg green tea | In vivo BALB/c mice | Decreased airway hyperresponsiveness, tissue injury, airway inflammation, eosinophil infiltrations Reduced specific IgE in the serum and BALF Upregulated amount of Treg cells and expression of Foxp3 mRNA in the lung tissue | No data available | [285,286] | |
| Kaempferol | 1–20 μM 10–20 mg/kg E. pungens leaf extract | In vitro BEAS-2B cells In vivo BALB/c mice | Improvement in symptoms of asthma Suppressed collagen deposition, epithelial excrescency, goblet hyperplasia, and fibrotic airway remodeling Decreased eosinophils and leukocyte numbers in blood and BLAF | No cytotoxic effect | [282,283,284,285,286,287,288,289] |
| 50, 250, 500 μg/mL 1, 10 mg/kg, p.o. Crocus sativus extract | In vitro PBMC In vivo BALB/c mice | Reduced nitric oxide level and inflammatory cytokines in the lung tissue Inhibited activation of NF-κB and STAT-1 in macrophages Reduced percentage of neutrophils and eosinophils in bronchoalveolar lavage fluid | No data available | [290] | |
| Quercetin | 25, 50 mg/kg pure isolated polyphenol | Neonatal asthmatic rats | Reduced total number of leukocytes, eosinophils, level of TNF-α, IL-6, nitric oxide synthesis and apoptosis, regulation of the Th2/Th1 imbalance | No data available | [292] |
| Pycnogenol (procyanidins, catechins) | 100 mg/day French maritime pine bark extract | Clinical study adults | Improved disease control: less frequent nighttime awakenings, decreased number of days with PEF < 80% and days with asthma score > 1, less frequent use of salbutamol and additional asthma medication Improvement in the severity of chest symptoms, wheezing, dyspnea, and daytime symptoms | No side effects, well-tolerated | [293] |
| 1 mg/kg/day French maritime pine bark extract | Clinical study children | Decrease in symptom scores, increase in lung function FEV1, PEF | No side effects, well-tolerated | [294] | |
| Anthocyanins, ellagitannins, chlorogenic acids | 0.2 mg/kg human equivalent dose BerriQi® Boysenberry and apple juice concentrate | In vivo BALB/c mice | Significantly decreased OVA-induced infiltrating eosinophils, neutrophils, and T cells in the lung, and mucous production | No data available | [295] |
| Gallic acid, ellagic acid | 100, 300 mg/kg | In vivo BALB/c mice | Reduced inflammatory cytokines, IgE, and number of inflammatory cells Reduction in inflammatory cell migration and mucus secretion in lung tissue | No data available | [296] |
| Magnolol | 12.5, 25, 50 mg/kg Magnolia officinalis extract | In vivo BALB/c mice | Reduction in allergic inflammation, decreased levels of Th2 and Th17 cytokines Suppression of allergen-induced airway hyperactivity, airway eosinophilic inflammation, airway collagen deposition, and airway mucus hypersecretion | No data available | [297,298] |
| Punicalagin | 12.5, 25, 50 mg/kg pomegranate extract | In vivo BALB/c mice | Decreased inflammatory cell infiltration into BALF Reduced levels of Th2-derived cytokines and specific IgE levels Regulation of IL-4/STAT6 and Notch/GATA3 signalling pathways | No data available | [299] |
| Ellagic acid | 500 mg/day pomegranate extract | Clinical study | Improved clinical symptoms of asthma like daily breath shortness, nocturnal breath shortness, and limitation of asthma-related activity Reduction in eosinophil, basophil, and neutrophil counts | No data available | [301] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dębińska, A.; Sozańska, B. Dietary Polyphenols—Natural Bioactive Compounds with Potential for Preventing and Treating Some Allergic Conditions. Nutrients 2023, 15, 4823. https://doi.org/10.3390/nu15224823
Dębińska A, Sozańska B. Dietary Polyphenols—Natural Bioactive Compounds with Potential for Preventing and Treating Some Allergic Conditions. Nutrients. 2023; 15(22):4823. https://doi.org/10.3390/nu15224823
Chicago/Turabian StyleDębińska, Anna, and Barbara Sozańska. 2023. "Dietary Polyphenols—Natural Bioactive Compounds with Potential for Preventing and Treating Some Allergic Conditions" Nutrients 15, no. 22: 4823. https://doi.org/10.3390/nu15224823
APA StyleDębińska, A., & Sozańska, B. (2023). Dietary Polyphenols—Natural Bioactive Compounds with Potential for Preventing and Treating Some Allergic Conditions. Nutrients, 15(22), 4823. https://doi.org/10.3390/nu15224823