

Probiotics Partly Suppress the Impact of Sugar Stress on the Oral Microbiota—A Randomized, Double-Blinded, Placebo-Controlled Trial
 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
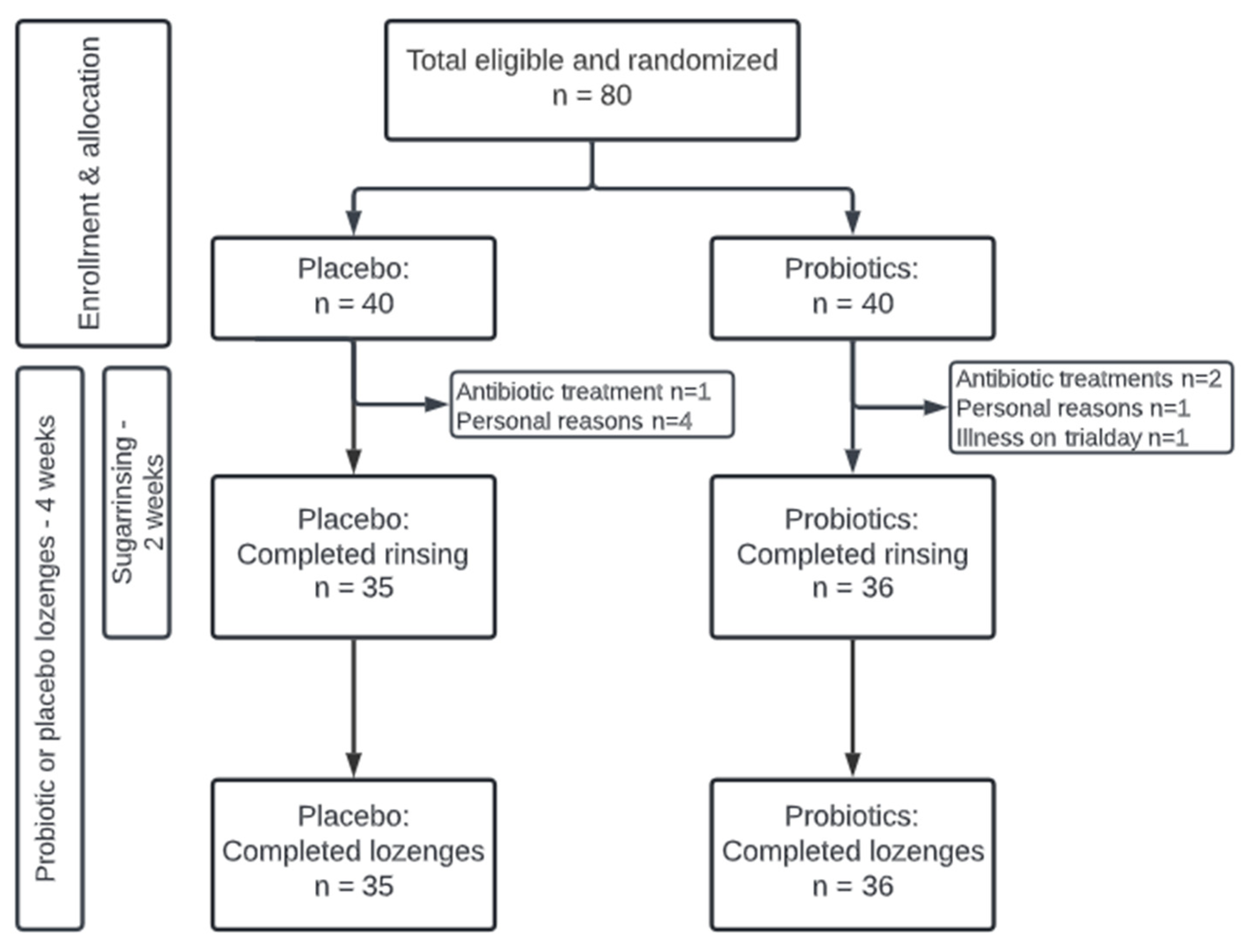
2.1. Study Design
2.2. Study Population
2.3. Clinical Examination
2.4. Collection of Samples
2.5. Probiotics and Placebo
2.6. Sucrose Solution
2.7. DNA Extraction, Library Preparation, and DNA Sequencing
2.8. Cytokine Analysis
2.9. Protein and Enzyme Analysis
2.10. Bioinformatic Processing and Statistics
3. Results
3.1. Background and Clinical Data
3.2. Sequencing Metadata
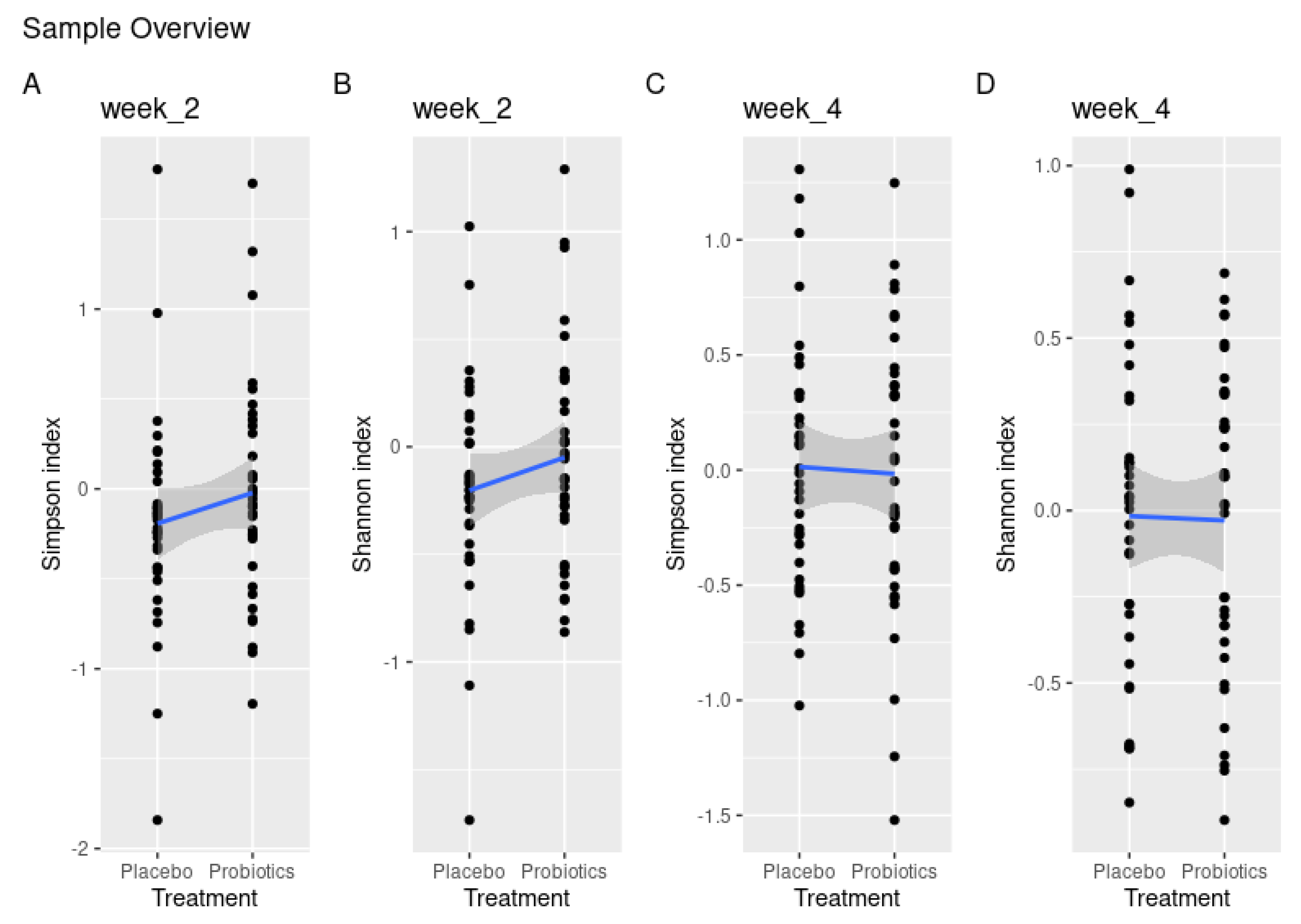
3.3. Probiotics Partly Protect against Sugar-Mediated Loss of α-Diversity
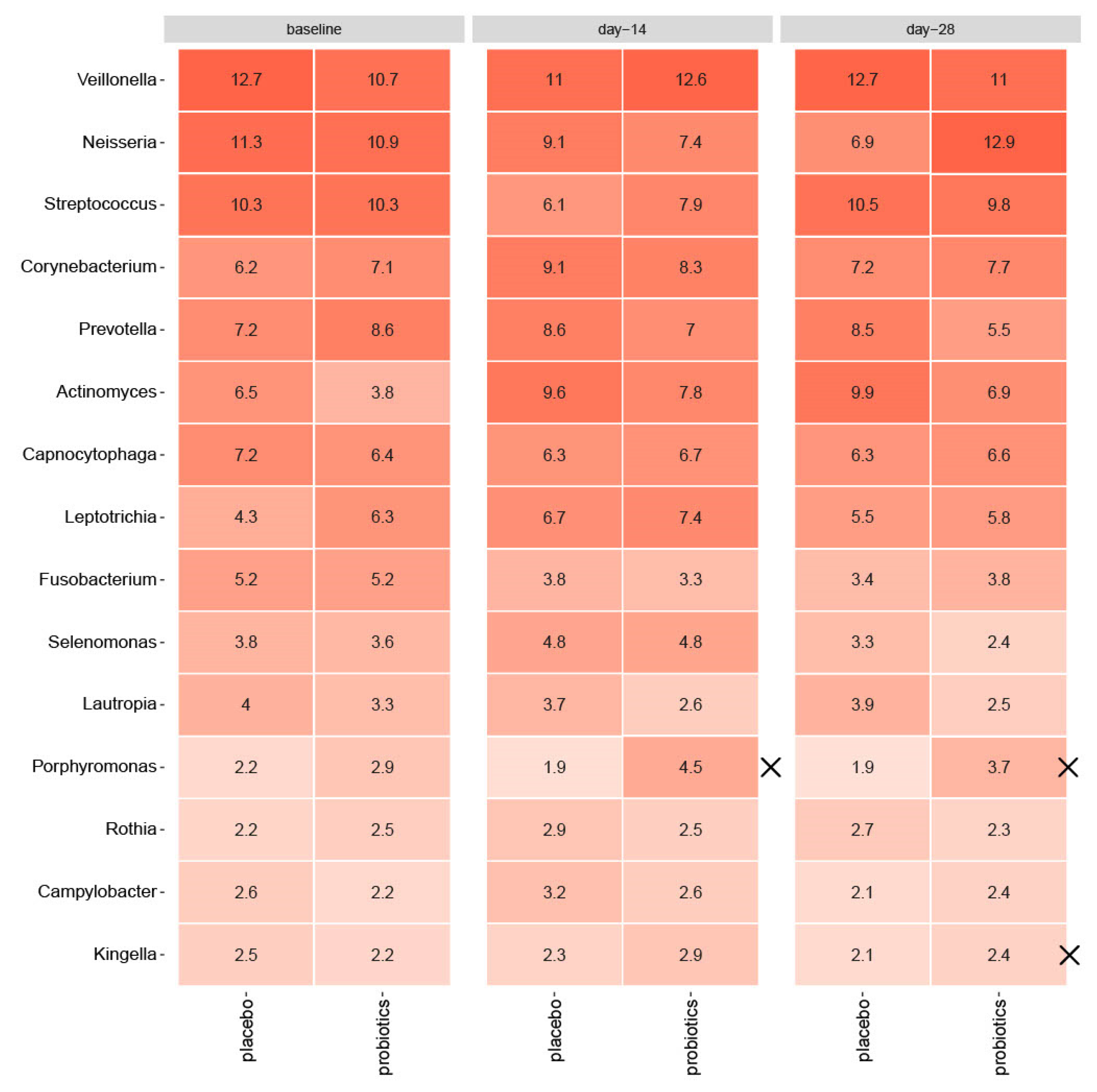
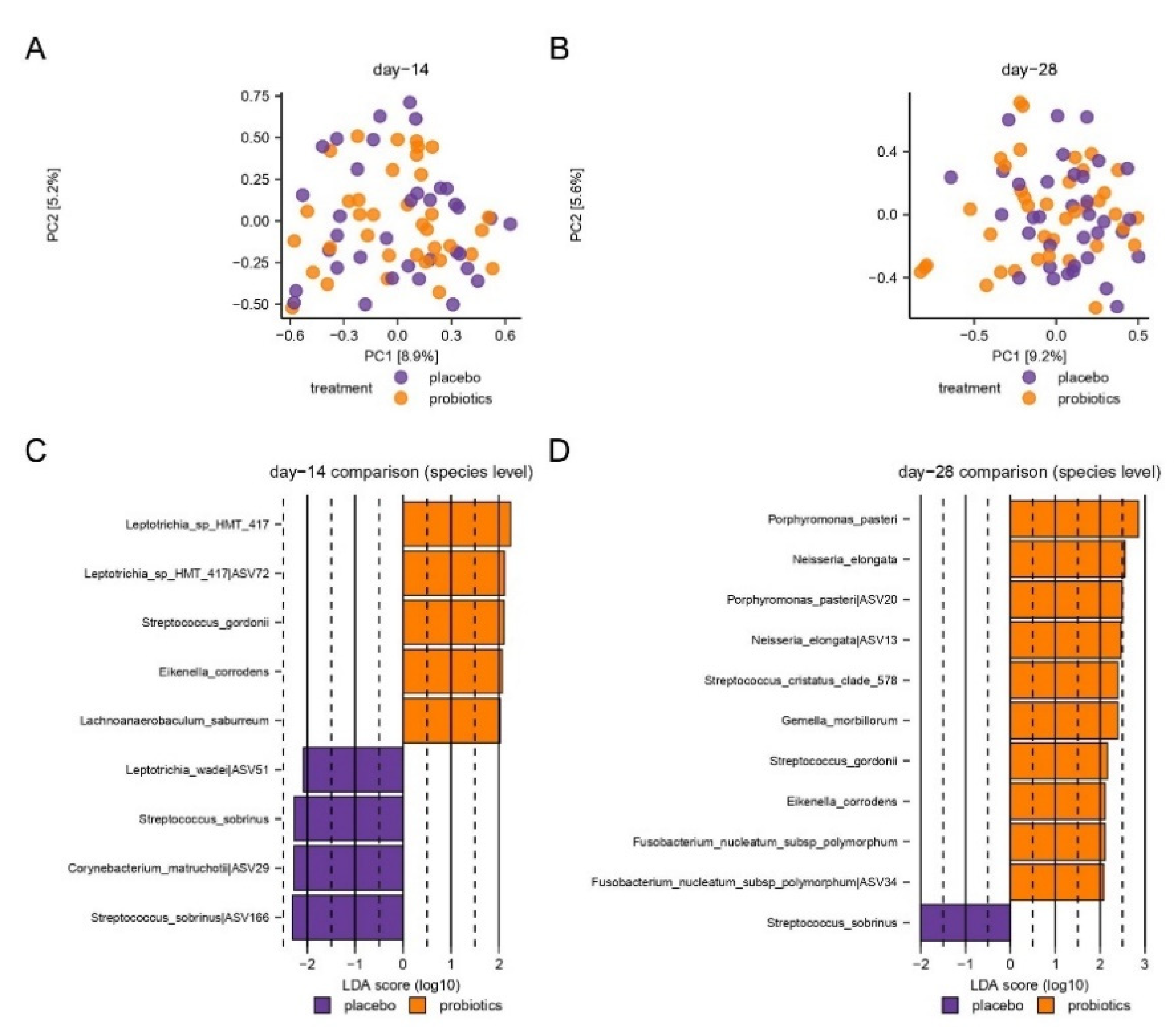
3.4. Compositional Changes of the Supragingival Microbiota
3.5. Salivary Levels of Pro-Inflammatory Cytokines and Proteases
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Escapa, I.F.; Chen, T.; Huang, Y.; Gajare, P.; Dewhirst, F.E.; Lemon, K.P. New Insights into Human Nostril Microbiome from the Expanded Human Oral Microbiome Database (eHOMD): A Resource for the Microbiome of the Human Aerodigestive Tract. mSystems 2018, 3, e00187-18. [Google Scholar] [CrossRef] [PubMed]
- Marsh, P.D.; Zaura, E. Dental biofilm: Ecological interactions in health and disease. J. Clin. Periodontol. 2017, 44 (Suppl. S18), S12–S22. [Google Scholar] [CrossRef] [PubMed]
- Guarner, F.; Schaafsma, G.J. Probiotics. Int. J. Food Microbiol. 1998, 39, 237–238. [Google Scholar] [CrossRef] [PubMed]
- Zaura, E.; Twetman, S. Critical Appraisal of Oral Pre- and Probiotics for Caries Prevention and Care. Caries Res. 2019, 53, 514–526. [Google Scholar] [CrossRef]
- Hadj-Hamou, R.; Senok, A.C.; Athanasiou, A.E.; Kaklamanos, E.G. Do probiotics promote oral health during orthodontic treatment with fixed appliances? A systematic review. BMC Oral Health 2020, 20, 126. [Google Scholar] [CrossRef]
- Meng, N.; Liu, Q.; Dong, Q.; Gu, J.; Yang, Y. Effects of probiotics on preventing caries in preschool children: A systematic review and meta-analysis. J. Clin. Pediatr. Dent. 2023, 47, 85–100. [Google Scholar] [CrossRef]
- Twetman, S.; Jørgensen, M.R. Can probiotic supplements prevent early childhood caries? A systematic review and meta-analysis. Benef. Microbes 2021, 12, 231–238. [Google Scholar] [CrossRef]
- Sheiham, A. Dietary effects on dental diseases. Public Health Nutr. 2001, 4, 569–591. [Google Scholar] [CrossRef]
- Diet, Nutrition and the Prevention of Chronic Diseases: Report of a Joint WHO/FAO Expert Consultation; WHO Technical Report Series, No. 916; World Health Organization: Geneva, Switzerland, 2003.
- Moynihan, P.J.; Kelly, S.A. Effect on caries of restricting sugars intake: Systematic review to inform WHO guidelines. J. Dent. Res. 2014, 93, 8–18. [Google Scholar] [CrossRef]
- Esberg, A.; Haworth, S.; Hasslöf, P.; Lif Holgerson, P.; Johansson, I. Oral Microbiota Profile Associates with Sugar Intake and Taste Preference Genes. Nutrients 2020, 12, 681. [Google Scholar] [CrossRef]
- Angarita-Díaz, M.D.P.; Fong, C.; Bedoya-Correa, C.M.; Cabrera-Arango, C.L. Does high sugar intake really alter the oral microbiota?: A systematic review. Clin. Exp. Dent. Res. 2022, 8, 1376–1390. [Google Scholar] [CrossRef] [PubMed]
- Marsh, P.D. Microbial ecology of dental plaque and its significance in health and disease. Adv. Dent. Res. 1994, 8, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Nyvad, B.; Takahashi, N. Integrated hypothesis of dental caries and periodontal diseases. J. Oral Microbiol. 2020, 12, 1710953. [Google Scholar] [CrossRef]
- Lundtorp-Olsen, C.; Enevold, C.; Juel Jensen, C.A.; Stofberg, S.N.; Twetman, S.; Belstrøm, D. Impact of Probiotics on the Salivary Microbiota and Salivary Levels of Inflammation-Related Proteins during Short-Term Sugar Stress: A Randomized Controlled Trial. Pathogens 2021, 10, 392. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Nan, F.; Liang, H.; Shu, P.; Fan, X.; Song, X.; Hou, Y.; Zhang, D. Excessive intake of sugar: An accomplice of inflammation. Front. Immunol. 2022, 13, 988481. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Fan, J.; Su, Q.; Yang, Z. Cytokines and Abnormal Glucose and Lipid Metabolism. Front. Endocrinol. 2019, 10, 703. [Google Scholar] [CrossRef] [PubMed]
- Lundtorp Olsen, C.; Markvart, M.; Vendius, V.F.D.; Damgaard, C.; Belstrøm, D. Short-term sugar stress induces compositional changes and loss of diversity of the supragingival microbiota. J. Oral Microbiol. 2023, 15, 2189770. [Google Scholar] [CrossRef]
- Lundtorp-Olsen, C.; Enevold, C.; Twetman, S.; Belstrøm, D. Probiotics Do Not Alter the Long-Term Stability of the Supragingival Microbiota in Healthy Subjects: A Randomized Controlled Trial. Pathogens 2021, 10, 391. [Google Scholar] [CrossRef]
- Lobene, R.R.; Soparkar, P.M.; Newman, M.B. Use of dental floss. Effect on plaque and gingivitis. Clin. Prev. Dent. 1982, 4, 5–8. [Google Scholar]
- Bardow, A.; Lykkeaa, J.; Qvist, V.; Ekstrand, K.; Twetman, S.; Fiehn, N.E. Saliva composition in three selected groups with normal stimulated salivary flow rates, but yet major differences in caries experience and dental erosion. Acta Odontol. Scand. 2014, 72, 466–473. [Google Scholar] [CrossRef]
- Belstrøm, D.; Sembler-Møller, M.L.; Grande, M.A.; Kirkby, N.; Cotton, S.L.; Paster, B.J.; Twetman, S.; Holmstrup, P. Impact of Oral Hygiene Discontinuation on Supragingival and Salivary Microbiomes. JDR Clin. Transl. Res. 2018, 3, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Keller, M.K.; Brandsborg, E.; Holmstrom, K.; Twetman, S. Effect of tablets containing probiotic candidate strains on gingival inflammation and composition of the salivary microbiome: A randomised controlled trial. Benef Microbes 2018, 9, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, M.; Demir, E.; Gürsoy, M.; Firatli, E.; Loimaranta, V.; Gürsoy, U.K. Salivary levels of BAFF, TWEAK, and soluble CD163 and salivary arginase activity before and after periodontal treatment. J. Periodontal Res. 2023, 58, 646–654. [Google Scholar] [CrossRef]
- Yilmaz, N.; Polat, R.; Gürsoy, M.; Kaman, W.; Gül Aydin, E.; Fteita, D.; Yilmaz, D.; Bikker, F.; Gürsoy, U.K. Salivary macrophage activation-related chemokines and mitogen-activated kinase kinase (MAPKK)-degrading proteolytic activity in type 1 diabetes mellitus. J. Periodontol. 2023, 94, 896–904. [Google Scholar] [CrossRef] [PubMed]
- Bikker, F.J.; Nascimento, G.G.; Nazmi, K.; Silbereisen, A.; Belibasakis, G.N.; Kaman, W.E.; Lopez, R.; Bostanci, N. Salivary Total Protease Activity Based on a Broad-Spectrum Fluorescence Resonance Energy Transfer Approach to Monitor Induction and Resolution of Gingival Inflammation. Mol. Diagn. Ther. 2019, 23, 667–676. [Google Scholar] [CrossRef]
- Prodan, A.; Brand, H.S.; Ligtenberg, A.J.; Imangaliyev, S.; Tsivtsivadze, E.; van der Weijden, F.; Crielaard, W.; Keijser, B.J.; Veerman, E.C. Interindividual variation, correlations, and sex-related differences in the salivary biochemistry of young healthy adults. Eur. J. Oral Sci. 2015, 123, 149–157. [Google Scholar] [CrossRef]
- Morquecho-Campos, P.; Bikker, F.J.; Nazmi, K.; de Graaf, K.; Laine, M.L.; Boesveldt, S. A stepwise approach investigating salivary responses upon multisensory food cues. Physiol. Behav. 2020, 226, 113116. [Google Scholar] [CrossRef]
- Janus, M.M.; Keijser, B.J.; Bikker, F.J.; Exterkate, R.A.; Crielaard, W.; Krom, B.P. In vitro phenotypic differentiation towards commensal and pathogenic oral biofilms. Biofouling 2015, 31, 503–510. [Google Scholar] [CrossRef]
- Hochberg, Y.; Benjamini, Y. More powerful procedures for multiple significance testing. Stat. Med. 1990, 9, 811–818. [Google Scholar] [CrossRef]
- Albertsen, M.; Karst, S.M.; Ziegler, A.S.; Kirkegaard, R.H.; Nielsen, P.H. Back to Basics--The Influence of DNA Extraction and Primer Choice on Phylogenetic Analysis of Activated Sludge Communities. PLoS ONE 2015, 10, e0132783. [Google Scholar] [CrossRef]
- Gross, E.L.; Beall, C.J.; Kutsch, S.R.; Firestone, N.D.; Leys, E.J.; Griffen, A.L. Beyond Streptococcus mutans: Dental caries onset linked to multiple species by 16S rRNA community analysis. PLoS ONE 2012, 7, e47722. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Ran, S.; Huang, Z.; Liang, J. Bacterial Diversity and Community Structure of Supragingival Plaques in Adults with Dental Health or Caries Revealed by 16S Pyrosequencing. Front. Microbiol. 2016, 7, 1145. [Google Scholar] [CrossRef] [PubMed]
- Wolff, D.; Frese, C.; Schoilew, K.; Dalpke, A.; Wolff, B.; Boutin, S. Amplicon-based microbiome study highlights the loss of diversity and the establishment of a set of species in patients with dentin caries. PLoS ONE 2019, 14, e0219714. [Google Scholar] [CrossRef]
- Belstrøm, D.; Holmstrup, P.; Fiehn, N.E.; Kirkby, N.; Kokaras, A.; Paster, B.J.; Bardow, A. Salivary microbiota in individuals with different levels of caries experience. J. Oral Microbiol. 2017, 9, 1270614. [Google Scholar] [CrossRef]
- Abranches, J.; Zeng, L.; Kajfasz, J.K.; Palmer, S.R.; Chakraborty, B.; Wen, Z.T.; Richards, V.P.; Brady, L.J.; Lemos, J.A. Biology of Oral Streptococci. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- Donati, C.; Zolfo, M.; Albanese, D.; Tin Truong, D.; Asnicar, F.; Iebba, V.; Cavalieri, D.; Jousson, O.; De Filippo, C.; Huttenhower, C.; et al. Uncovering oral Neisseria tropism and persistence using metagenomic sequencing. Nat. Microbiol. 2016, 1, 16070. [Google Scholar] [CrossRef]
- Korona-Glowniak, I.; Skawinska-Bednarczyk, A.; Wrobel, R.; Pietrak, J.; Tkacz-Ciebiera, I.; Maslanko-Switala, M.; Krawczyk, D.; Bakiera, A.; Borek, A.; Malm, A.; et al. Streptococcus sobrinus as a Predominant Oral Bacteria Related to the Occurrence of Dental Caries in Polish Children at 12 Years Old. Int. J. Environ. Res. Public Health 2022, 19, 15005. [Google Scholar] [CrossRef]
- Fragkou, S.; Balasouli, C.; Tsuzukibashi, O.; Argyropoulou, A.; Menexes, G.; Kotsanos, N.; Kalfas, S. Streptococcus mutans, Streptococcus sobrinus and Candida albicans in oral samples from caries-free and caries-active children. Eur. Arch. Paediatr. Dent. Off. J. Eur. Acad. Paediatr. Dent. 2016, 17, 367–375. [Google Scholar] [CrossRef]
- Jiang, Q.; Yu, M.; Min, Z.; Yi, A.; Chen, D.; Zhang, Q. AP-PCR detection of Streptococcus mutans and Streptococcus sobrinus in caries-free and caries-active subjects. Mol. Cell. Biochem. 2012, 365, 159–164. [Google Scholar] [CrossRef]
- Gao, X.; Jiang, S.; Koh, D.; Hsu, C.Y. Salivary biomarkers for dental caries. Periodontology 2000 2016, 70, 128–141. [Google Scholar] [CrossRef]
- Marchant, S.; Brailsford, S.R.; Twomey, A.C.; Roberts, G.J.; Beighton, D. The predominant microflora of nursing caries lesions. Caries Res. 2001, 35, 397–406. [Google Scholar] [CrossRef]
- Dong, Y.; Chen, Y.Y.; Snyder, J.A.; Burne, R.A. Isolation and molecular analysis of the gene cluster for the arginine deiminase system from Streptococcus gordonii DL1. Appl. Environ. Microbiol. 2002, 68, 5549–5553. [Google Scholar] [CrossRef]
- Lin, X.; Lamont, R.J.; Wu, J.; Xie, H. Role of differential expression of streptococcal arginine deiminase in inhibition of fimA expression in Porphyromonas gingivalis. J. Bacteriol. 2008, 190, 4367–4371. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kuramitsu, H.K.; Wang, B.Y. Virulence properties of cariogenic bacteria. BMC Oral Health 2006, 6 (Suppl. S1), S11. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.Y.; Kuramitsu, H.K. Interactions between oral bacteria: Inhibition of Streptococcus mutans bacteriocin production by Streptococcus gordonii. Appl. Environ. Microbiol. 2005, 71, 354–362. [Google Scholar] [CrossRef]
- Qudeimat, M.A.; Alyahya, A.; Karched, M.; Behbehani, J.; Salako, N.O. Dental plaque microbiota profiles of children with caries-free and caries-active dentition. J. Dent. 2021, 104, 103539. [Google Scholar] [CrossRef] [PubMed]
- Yasunaga, H.; Takeshita, T.; Shibata, Y.; Furuta, M.; Shimazaki, Y.; Akifusa, S.; Ninomiya, T.; Kiyohara, Y.; Takahashi, I.; Yamashita, Y. Exploration of bacterial species associated with the salivary microbiome of individuals with a low susceptibility to dental caries. Clin. Oral Investig. 2017, 21, 2399–2406. [Google Scholar] [CrossRef]
- Henskens, Y.M.; van der Velden, U.; Veerman, E.C.; Nieuw Amerongen, A.V. Protein, albumin and cystatin concentrations in saliva of healthy subjects and of patients with gingivitis or periodontitis. J. Periodontal Res. 1993, 28, 43–48. [Google Scholar] [CrossRef]
- Verhulst, M.J.L.; Teeuw, W.J.; Bizzarro, S.; Muris, J.; Su, N.; Nicu, E.A.; Nazmi, K.; Bikker, F.J.; Loos, B.G. A rapid, non-invasive tool for periodontitis screening in a medical care setting. BMC Oral Health 2019, 19, 87. [Google Scholar] [CrossRef]
- Katsiki, P.; Nazmi, K.; Loos, B.G.; Laine, M.L.; Schaap, K.; Hepdenizli, E.; Bikker, F.J.; Brand, H.S.; Veerman, E.C.I.; Nicu, E.A. Comparing periodontitis biomarkers in saliva, oral rinse and gingival crevicular fluid: A pilot study. J. Clin. Periodontol. 2021, 48, 1250–1259. [Google Scholar] [CrossRef]
- Gustafsson, B.E.; Quensel, C.E.; Lanke, L.S.; Lundqvist, C.; Grahnen, H.; Bonow, B.E.; Krasse, B. The Vipeholm dental caries study; the effect of different levels of carbohydrate intake on caries activity in 436 individuals observed for five years. Acta Odontol. Scand. 1954, 11, 232–264. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Placebo (n = 35) | Probiotic (n = 36) | |
|---|---|---|
| Sex (female/male) | 26/9 | 29/7 |
| Age (mean, range) | 23.4 (19–30) | 24 (20–30) |
| Dental students | 25/35 | 21/36 |
| Clinical examination | ||
| Baseline | ||
| Plaque mean (sd) | 1.72 (0.3) | 1.75 (0.3) |
| BOP% mean (sd) | 2.82 (2.8) | 3.10 (2.9) |
| Day 14 | ||
| Plaque mean (sd) | 1.57 (0.3) | 1.69 (0.3) |
| BOP% mean (sd) | 3.37 (2.9) | 4.18 (3.2) |
| Day 28 | ||
| Plaque mean (sd) | 1.55 (0.3) | 1.57 (0.3) |
| BOP% mean (sd) | 3.47 (2.7) | 3.94 (3.4) |
| IL-1β | IL-8 | MCP-1 | MIF | |
|---|---|---|---|---|
| Baseline | ||||
| Placebo mean (sd) | 1.06 (0.53) | 2.2 (0.43) | 1.41 (0.43) | 1.61 (0.75) |
| Probiotics mean (sd) | 1.05 (0.44) | 2.17 (0.37) | 1.51 (0.39) | 1.75 (0.6) |
| Day 14 | ||||
| Placebo mean (sd) | 1.23 (0.48) | 2.26 (0.41) | 1.48 (0.42) | 1.72 (0.77) |
| Probiotics mean (sd) | 1.11 (0.46) | 2.14 (0.42) | 1.54 (0.43) | 1.69 (0.65) |
| p-value (CI) | 0.10 (−0.26; 0.03) | 0.19 (−0.23; 0.05) | 0.79 (−0.16; 0.12) | 0.46 (−0.40; 0.18) |
| Day 28 | ||||
| Placebo mean (sd) | 1.31 (0.52) | 2.28 (0.45) | 1.50 (0.44) | 1.78 (0.77) |
| Probiotics mean (sd) | 1.17 (0.51) | 2.14 (0.44) | 1.49 (0.50) | 1.62 (0.97) |
| p-value (CI) | 0.11 (−0.29; 0.03) | 0.14 (−0.27; 0.04) | 0.30 (−0.25; 0.08) | 0.16 (−0.61; 0.10) |
| Amylase Activity (slope/min) | Total Protease Activity (dF/dT) | Chitinase Activity (slope/s) | Albumin (µg/mL) | |
|---|---|---|---|---|
| Baseline | ||||
| Placebo mean (sd) | 0.04 (0.02) | 3.12 (0.24) | 0.35 (0.19) | 1.77 (0.3) |
| Probiotics mean (sd) | 0.04 (0.02) | 3.13 (0.22) | 0.3 (0.18) | 1.74 (0.25) |
| Day 14 | ||||
| Placebo mean (sd) | 0.04 (0.02) | 3.16 (0.27) | 0.31 (0.17) | 1.65 (0.26) |
| Probiotics mean (sd) | 0.04 (0.02) | 3.11 (0.23) | 0.27 (0.16) | 1.61 (0.6) |
| p -value (CI) | 0.87 | 0.25 | 0.72 | 0.63 |
| (−0.005; 0.007) | (−0.16; 0.04) | (−0.03; 0.05) | (−0.13; 0.08) | |
| Day 28 | ||||
| Placebo mean (sd) | 0.04 (0.02) | 3.17 (0.26) | 0.31 (0.16) | 1.68 (0.32) |
| Probiotics mean (sd) | 0.04 (0.02) | 3.11 (0.20) | 0.25 (0.16) | 1.54 (0.25) |
| p -value (CI) | 0.32 | 0.18 | 0.57 | 0.02 |
| (−0.003; 0.009) | (−0.15; 0.03) | (−0.05; 0.03) | (−0.23; −0.02) * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lundtorp Olsen, C.; Massarenti, L.; Vendius, V.F.D.; Gürsoy, U.K.; Van Splunter, A.; Bikker, F.J.; Gürsoy, M.; Damgaard, C.; Markvart, M.; Belstrøm, D. Probiotics Partly Suppress the Impact of Sugar Stress on the Oral Microbiota—A Randomized, Double-Blinded, Placebo-Controlled Trial. Nutrients 2023, 15, 4810. https://doi.org/10.3390/nu15224810
Lundtorp Olsen C, Massarenti L, Vendius VFD, Gürsoy UK, Van Splunter A, Bikker FJ, Gürsoy M, Damgaard C, Markvart M, Belstrøm D. Probiotics Partly Suppress the Impact of Sugar Stress on the Oral Microbiota—A Randomized, Double-Blinded, Placebo-Controlled Trial. Nutrients. 2023; 15(22):4810. https://doi.org/10.3390/nu15224810
Chicago/Turabian StyleLundtorp Olsen, Christine, Laura Massarenti, Vincent Frederik Dahl Vendius, Ulvi Kahraman Gürsoy, Annina Van Splunter, Floris J. Bikker, Mervi Gürsoy, Christian Damgaard, Merete Markvart, and Daniel Belstrøm. 2023. "Probiotics Partly Suppress the Impact of Sugar Stress on the Oral Microbiota—A Randomized, Double-Blinded, Placebo-Controlled Trial" Nutrients 15, no. 22: 4810. https://doi.org/10.3390/nu15224810
APA StyleLundtorp Olsen, C., Massarenti, L., Vendius, V. F. D., Gürsoy, U. K., Van Splunter, A., Bikker, F. J., Gürsoy, M., Damgaard, C., Markvart, M., & Belstrøm, D. (2023). Probiotics Partly Suppress the Impact of Sugar Stress on the Oral Microbiota—A Randomized, Double-Blinded, Placebo-Controlled Trial. Nutrients, 15(22), 4810. https://doi.org/10.3390/nu15224810