
Phycobiliprotein Peptide Extracts from Arthrospira platensis Ameliorate Nonalcoholic Fatty Liver Disease by Modulating Hepatic Lipid Profile and Strengthening Fat Mobilization

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of PPE
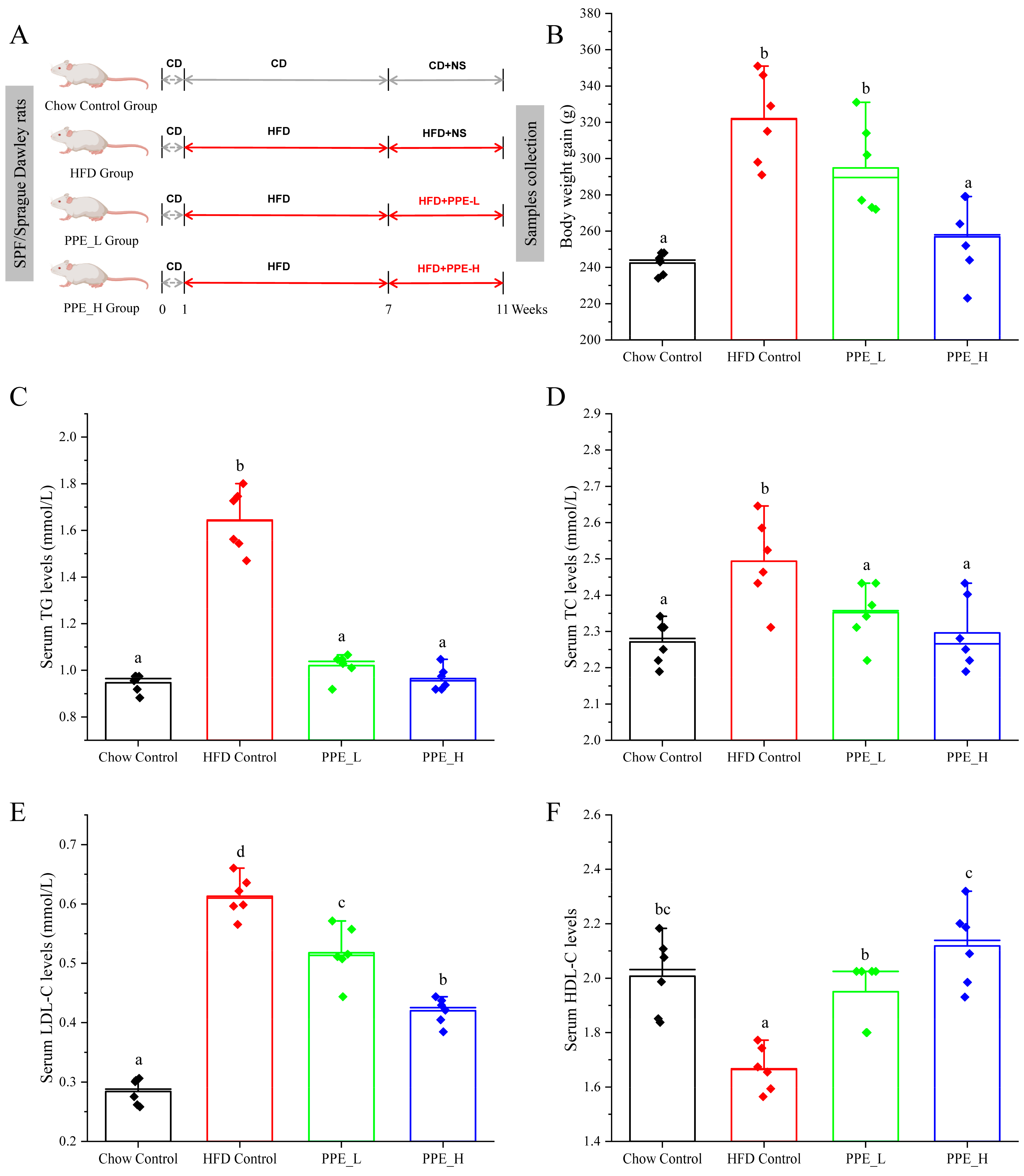
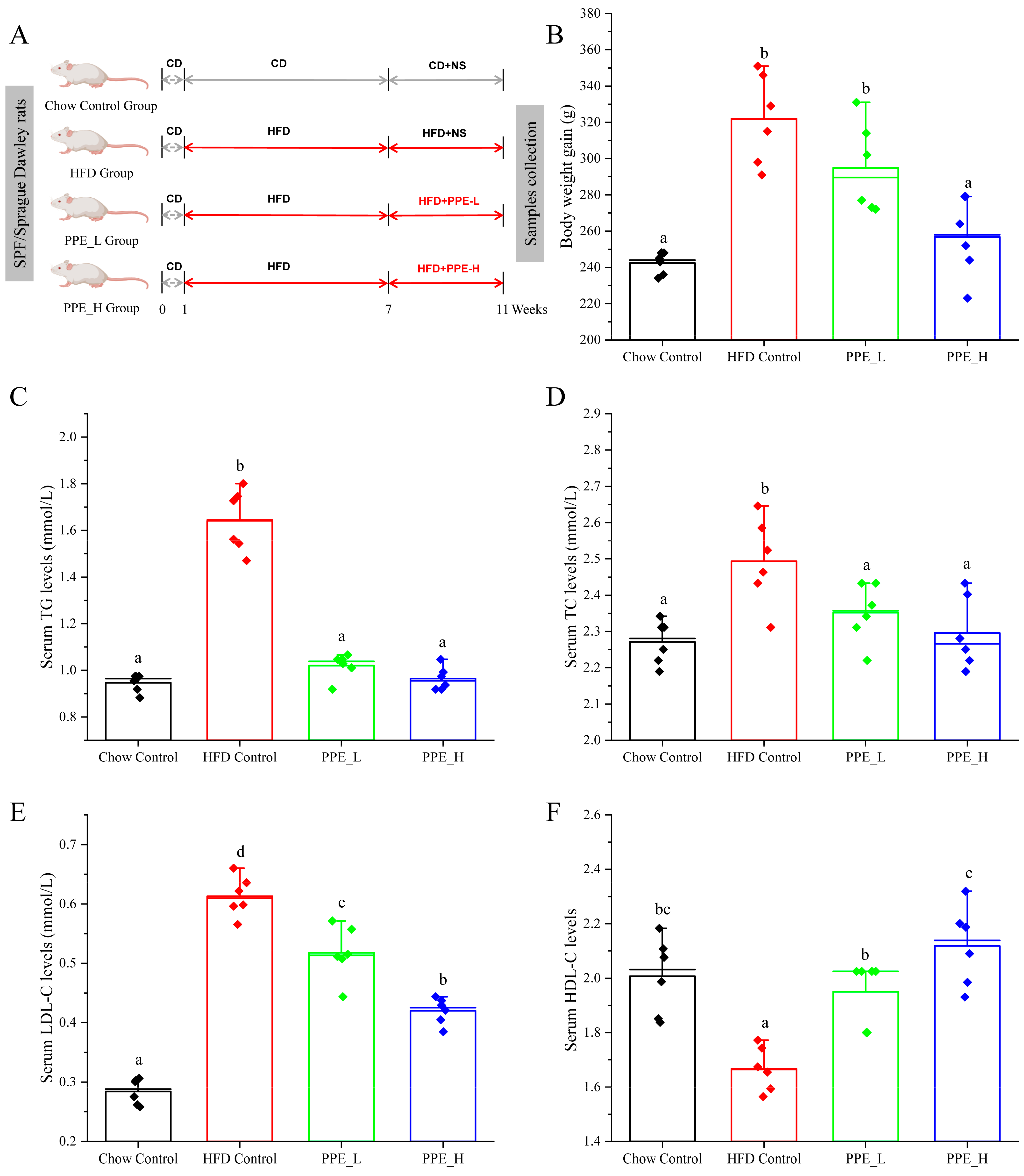
2.2. Animal Experiments and Sample Collection
2.3. Serum Biochemical Assessment
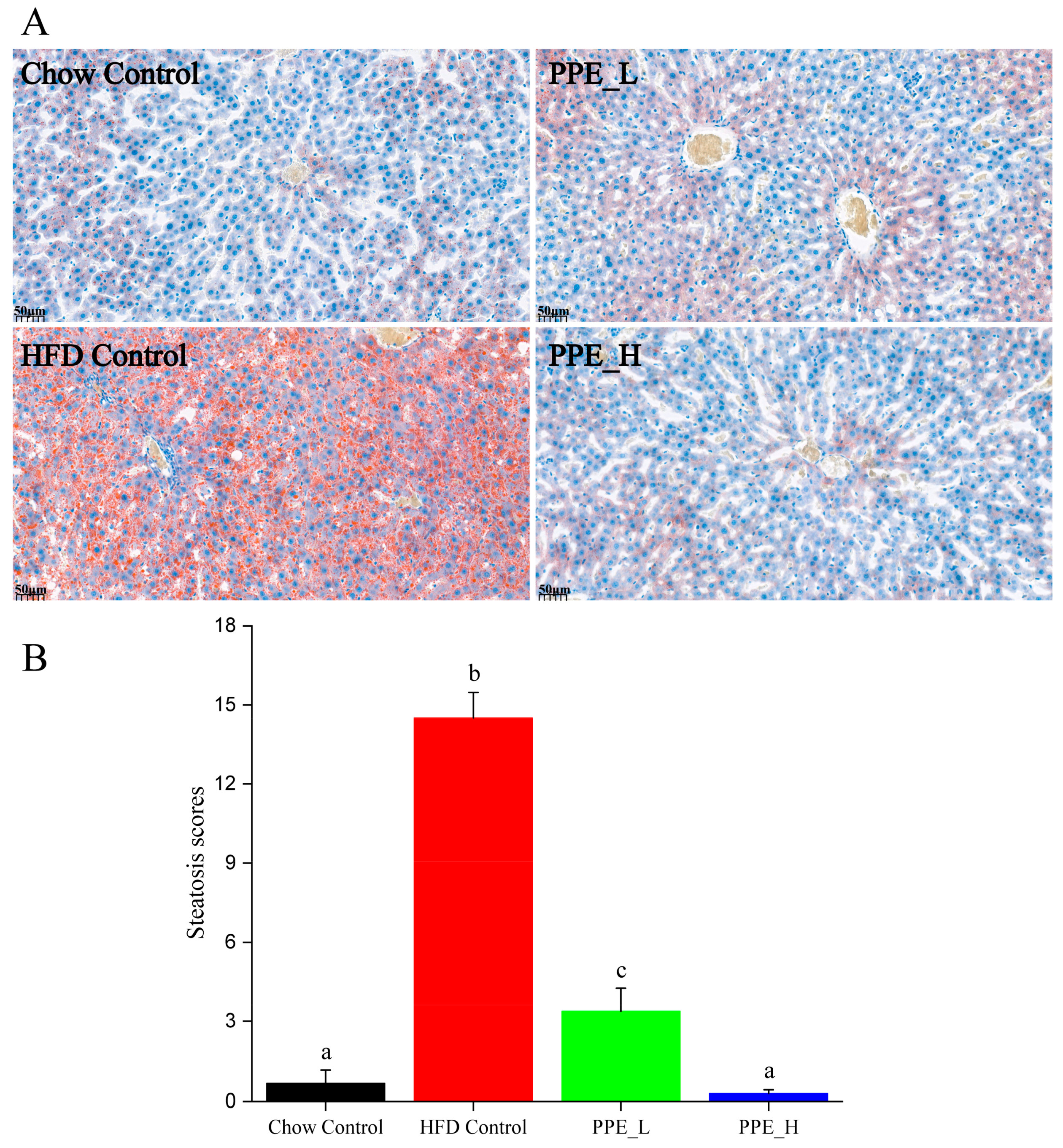
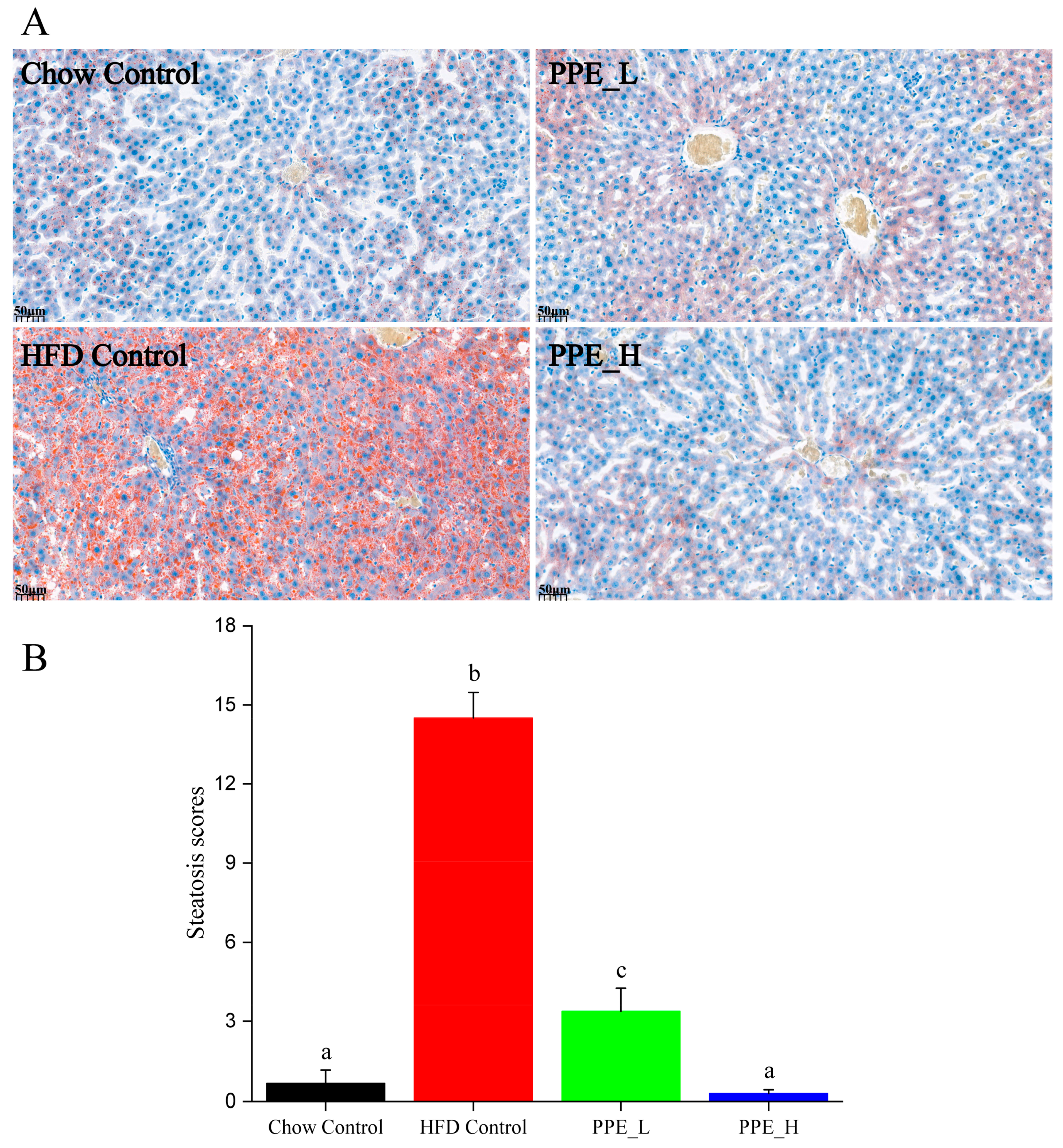
2.4. Quantification of Lipid Abundance in Liver Tissues by Oil Red O
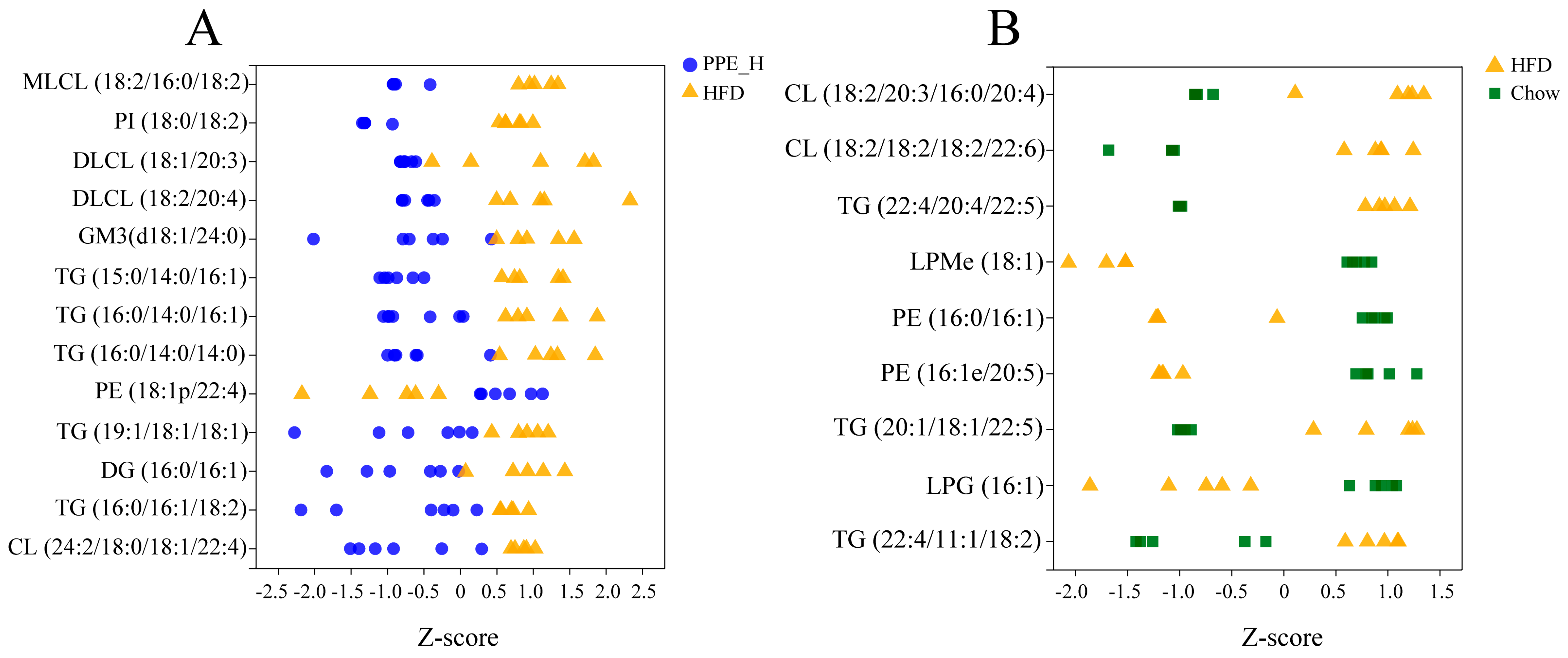
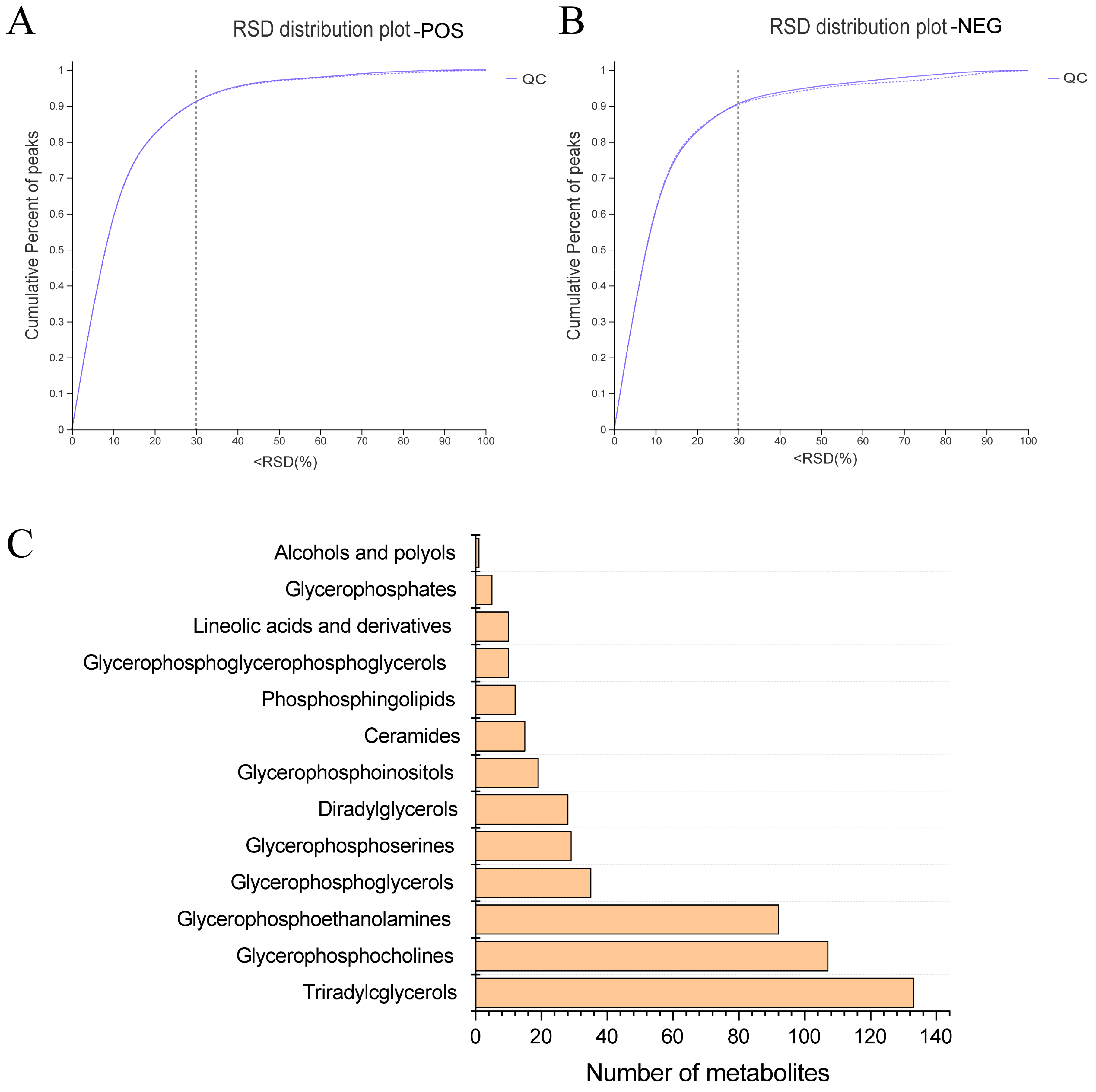
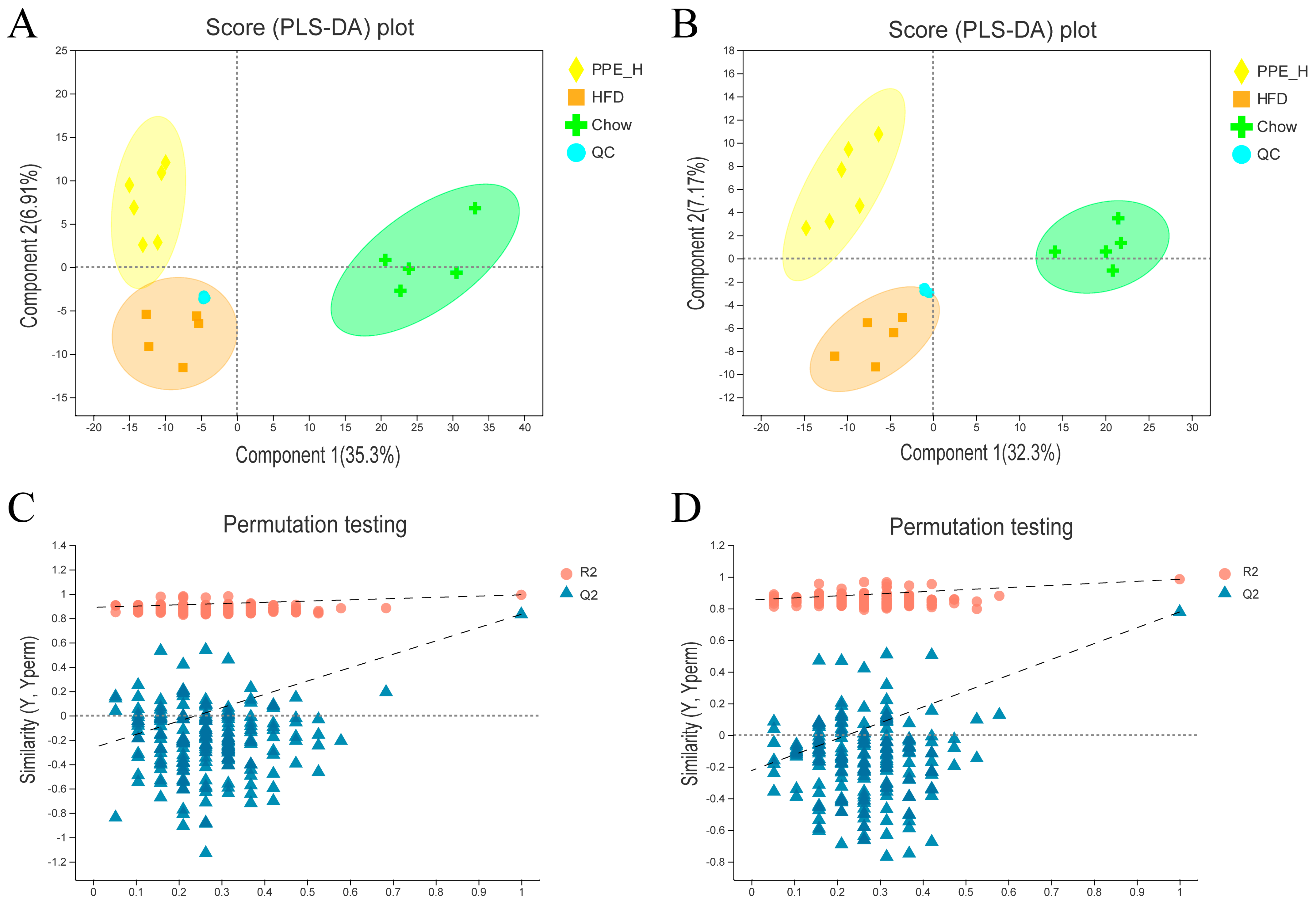
2.5. Lipidomic Analysis
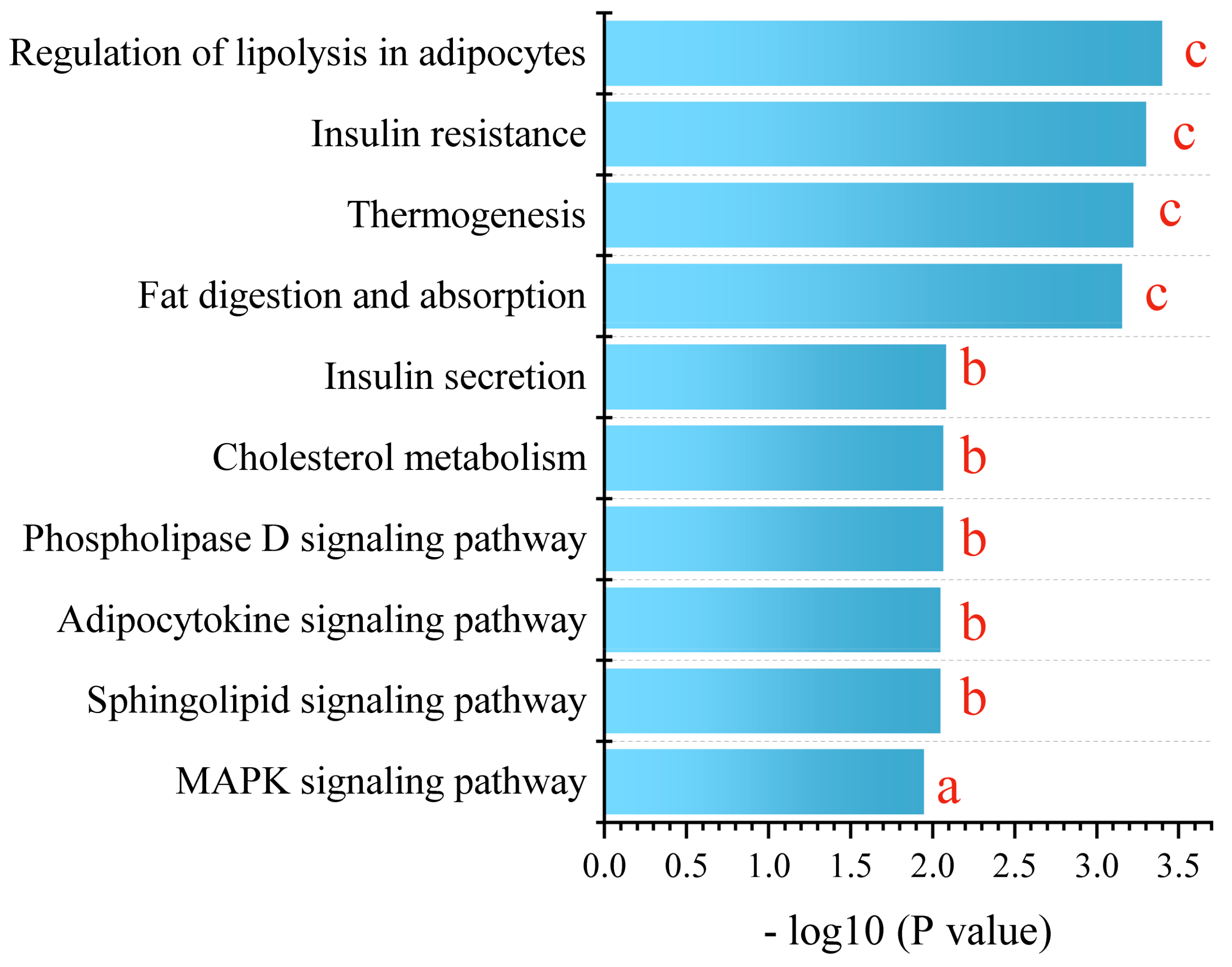
2.6. Gene Expression Analyses in White Adipose Tissue
2.7. Metabolite Profiling of Cecal Contents Using GC-MS
2.8. Statistical Analyses
3. Results
3.1. PPE Ameliorated HFD-Induced Hepatic Steatosis in Rats
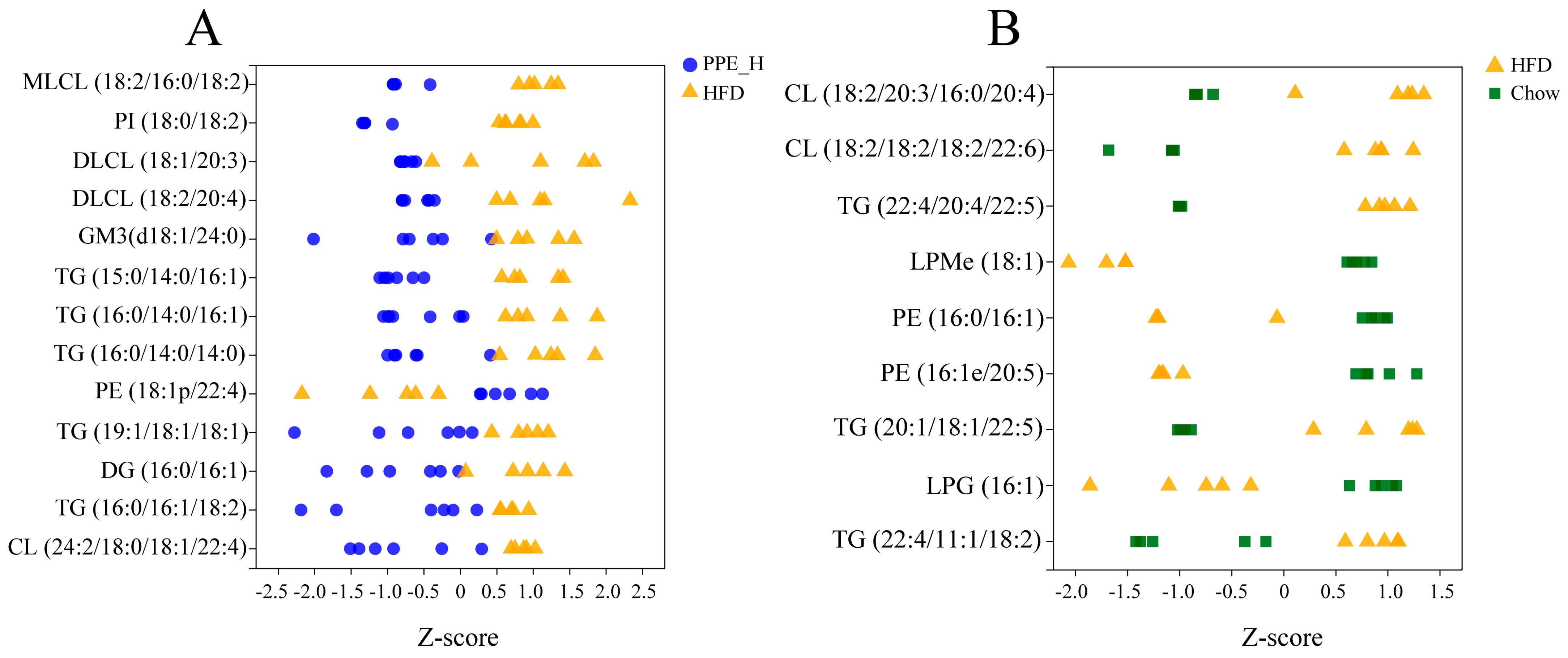
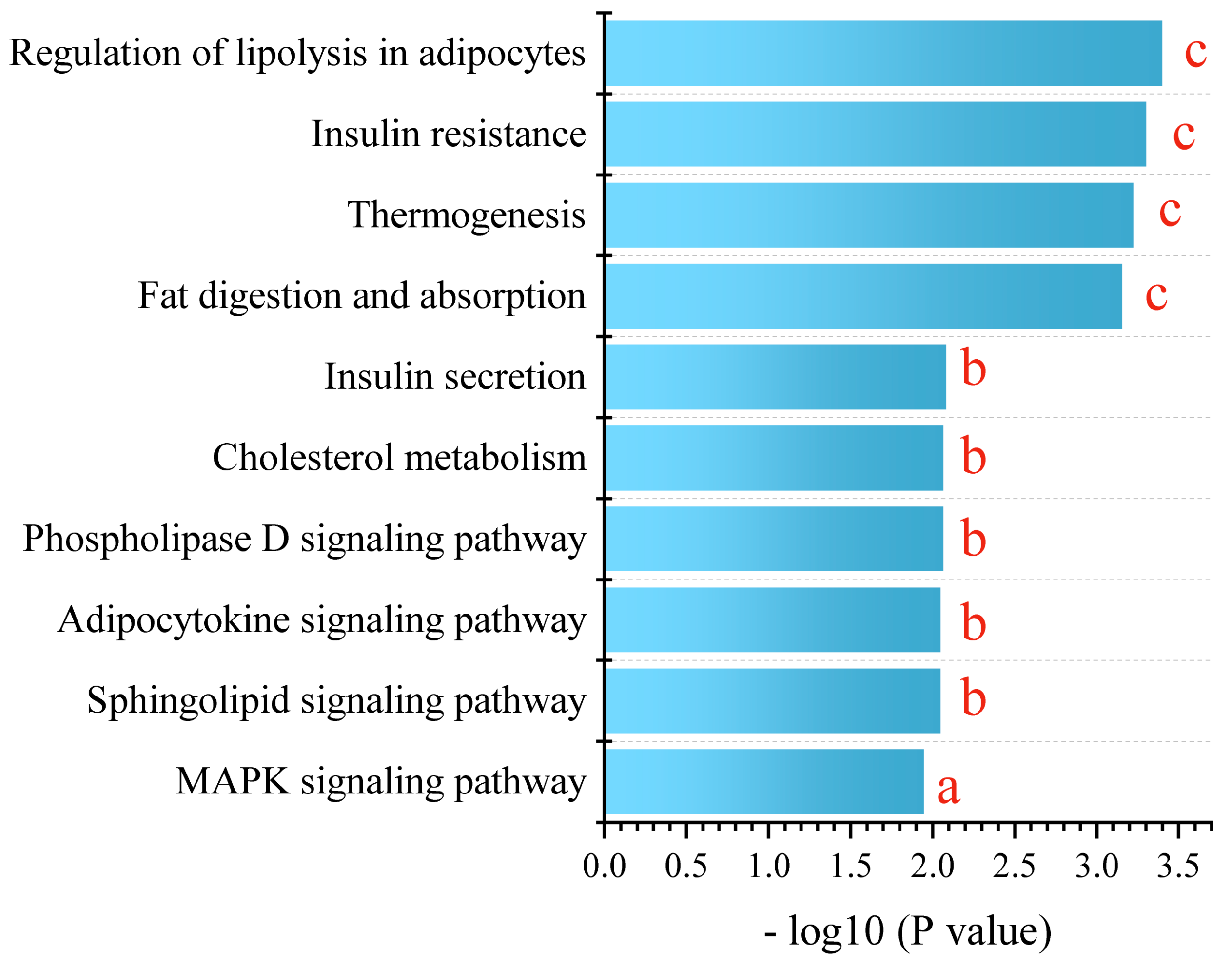
3.2. PPE Extensively Modulated Hepatic Lipid Profile
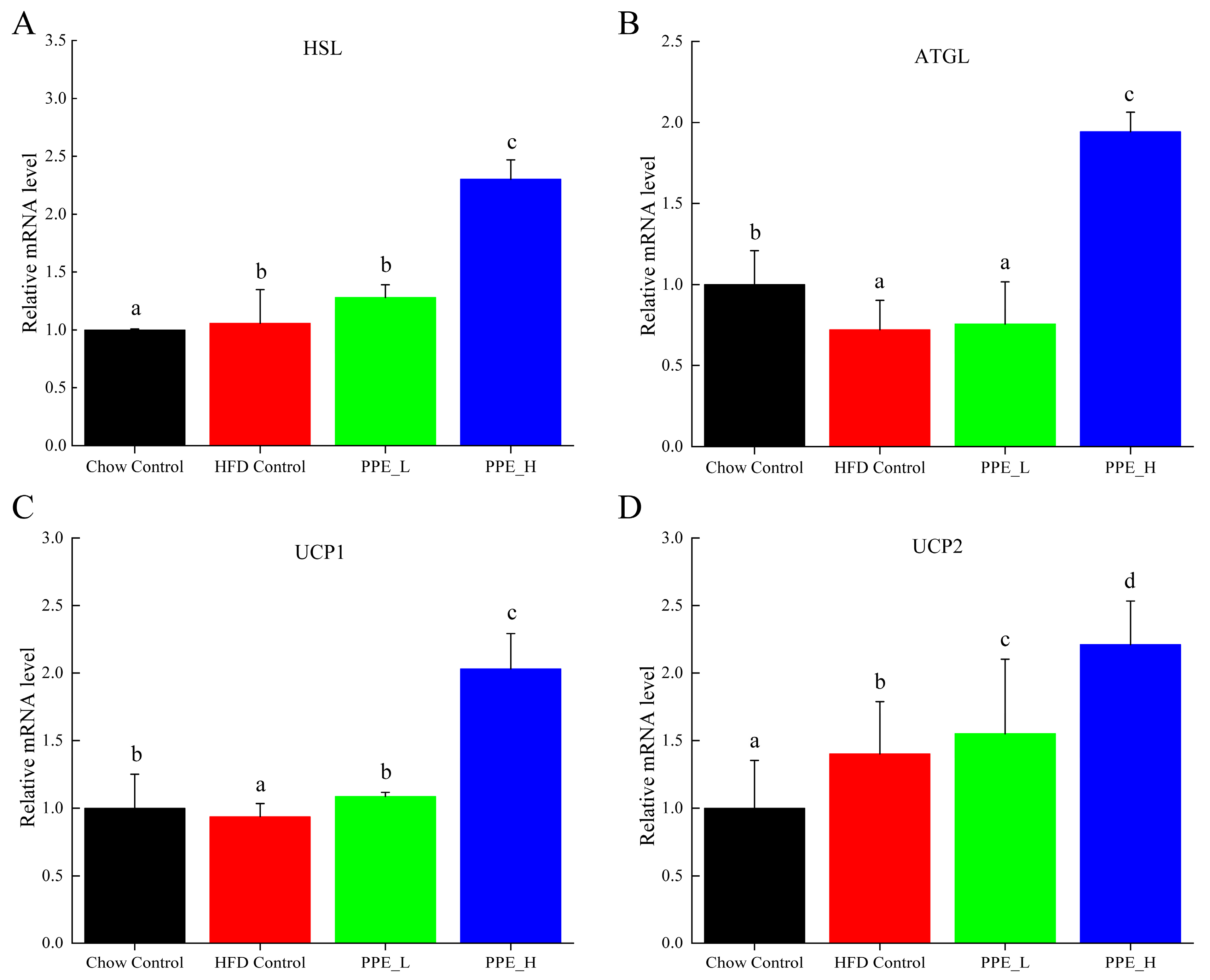
3.3. PPE Regulated Expression of the Lipolysis Gene in White Adipose Tissue and Strengthened Mobilization from Fat
3.4. PPE Affected Intestinal Metabolites: Long-Chain Fatty Acids and Short-Chain Fatty Acids
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Afonso, M.B.; Rodrigues, P.M.; Mateus-Pinheiro, M.; Simao, A.L.; Gaspar, M.M.; Majdi, A.; Arretxe, E.; Alonso, C.; Santos-Laso, A.; Jimenez-Aguero, R.; et al. RIPK3 acts as a lipid metabolism regulator contributing to inflammation and carcinogenesis in non-alcoholic fatty liver disease. Gut 2021, 70, 2359–2372. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Das, C.J.; Baruah, M.P. Imaging of nonalcoholic fatty liver disease: A road less travelled. Indian J. Endocrinol. Metab. 2013, 17, 990–995. [Google Scholar] [CrossRef] [PubMed]
- Bourebaba, L.; Łyczko, J.; Alicka, M.; Bourebaba, N.; Szumny, A.; Fal, A.M.; Marycz, K. Inhibition of protein-tyrosine phosphatase PTP1B and LMPTP promotes palmitate/oleate-challenged HepG2 cell survival by reducing lipoapoptosis, improving mitochondrial dynamics and mitigating oxidative and endoplasmic reticulum stress. J. Clin. Med. 2020, 9, 1294. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Tian, R.; She, Z.; Cai, J.; Li, H. Role of oxidative stress in the pathogenesis of nonalcoholic fatty liver disease. Free Radic. Biol. Med. 2020, 152, 116–141. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Zhang, W.; Zeng, L.Q.; Bai, H.; Li, J.; Zhou, J.; Zhou, G.Y.; Fang, C.W.; Wang, F.; Qin, X.J. Exercise and dietary intervention ameliorate high-fat diet-induced NAFLD and liver aging by inducing lipophagy. Redox Biol. 2020, 36, 101635. [Google Scholar] [CrossRef]
- Li, X.; Ge, J.; Li, Y.; Cai, Y.; Zheng, Q.; Huang, N.; Gu, Y.; Han, Q.; Li, Y.; Sun, R.; et al. Integrative lipidomic and transcriptomic study unravels the therapeutic effects of saikosaponins A and D on non-alcoholic fatty liver disease. Acta Pharm. Sin. B 2021, 11, 3527–3541. [Google Scholar] [CrossRef]
- Soto-Sierra, L.; Stoykova, P.; Nikolov, Z.L. Extraction and fractionation of microalgae-based protein products. Algal Res. 2018, 36, 175–192. [Google Scholar] [CrossRef]
- Lupatini, A.L.; Colla, L.M.; Canan, C.; Colla, E. Potential application of microalga Spirulina platensis as a protein source. J. Sci. Food Agric. 2017, 97, 724–732. [Google Scholar] [CrossRef]
- Li, Y.; Lammi, C.; Boschin, G.; Arnoldi, A.; Aiello, G. Recent Advances in Microalgae Peptides: Cardiovascular Health Benefits and Analysis. J. Agric. Food Chem. 2019, 67, 11825–11838. [Google Scholar] [CrossRef]
- Liu, J.; Bai, X.; Fu, P. In silico and in vitro assessment of bioactive peptides from Arthrospira platensis phycobiliproteins for DPP-IV inhibitory activity, ACE inhibitory activity, and antioxidant activity. J. Appl. Phycol. 2022, 34, 1497–1511. [Google Scholar] [CrossRef]
- Sun, M.; Ye, H. Natural foods for the treatment of nonalcoholic fatty liver disease. J. Med. Food 2023, 26, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Rangboo, V.; Noroozi, M.; Zavoshy, R.; Rezadoost, S.A.; Mohammadpoorasl, A. The effect of artichokeleaf extract on alanine aminotransferase and aspartate aminotransferase in the patients with nonalcoholic steatohepatitis. Int. J. Hepatol. 2016, 2016, 4030476. [Google Scholar] [CrossRef]
- Gao, L.-L.; Li, Y.-X.; Ma, J.-M.; Guo, Y.-Q.; Li, L.; Gao, Q.-H.; Fan, Y.-N.; Zhang, M.-W.; Tao, X.-J.; Yu, J.-Q. Effect of Lycium barbarum polysaccharide supplementation in non-alcoholic fatty liver disease patients: Study protocol for a randomized controlled trial. Trials 2021, 22, 566. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Guo, R.; Guo, W.; Hong, J.; Li, L.; Ni, L.; Sun, J.; Liu, B.; Rao, P.; Lv, X. Monascus yellow, red and orange pigments from red yeast rice ameliorate lipid metabolic disorders and gut microbiota dysbiosis in Wistar rats fed on a high-fat diet. Food Funct. 2019, 10, 1073–1084. [Google Scholar] [CrossRef]
- Sangouni, A.A.; Azar, M.R.M.H.; Alizadeh, M. Effect of garlic powder supplementation on hepatic steatosis, liver enzymes and lipid profile in patients with non-alcoholic fatty liver disease: A double-blind randomised controlled clinical trial. Br. J. Nutr. 2020, 124, 450–456. [Google Scholar] [CrossRef]
- Soleimani, D.; Paknahad, Z.; Rouhani, M.H. Therapeutic effects of garlic on hepatic steatosis in nonalcoholic fatty liver disease patients: A randomized clinical trial. Diabetes Metab. Syndr. Obes. Targets Ther. 2020, 13, 2389. [Google Scholar] [CrossRef] [PubMed]
- Fougerat, A.; Montagner, A.; Loiseau, N.; Guillou, H.; Wahli, W. Peroxisome proliferator-activated receptors and their novel ligands as candidates for the treatment of non-alcoholic fatty liver disease. Cells 2020, 9, 1638. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.Q.; Zeng, Y.; Xu, J.; Le Xu, X. Naringenin alleviates nonalcoholic steatohepatitis in middle-aged Apoe−/− mice: Role of SIRT1. Phytomedicine 2021, 81, 153412. [Google Scholar] [CrossRef]
- Dowman, J.K.; Tomlinson, J.; Newsome, P. Pathogenesis of non-alcoholic fatty liver disease. QJM Int. J. Med. 2010, 103, 71–83. [Google Scholar] [CrossRef]
- Bugianesi, E.; Moscatiello, S.; Ciaravella, M.; Marchesini, G. Insulin resistance in nonalcoholic fatty liver disease. Curr. Pharm. Des. 2010, 16, 1941–1951. [Google Scholar] [CrossRef]
- Foretz, M.; Guichard, C.; Ferré, P.; Foufelle, F. Sterol regulatory element binding protein-1c is a major mediator of insulin action on the hepatic expression of glucokinase and lipogenesis-related genes. Proc. Natl. Acad. Sci. USA 1999, 96, 12737–12742. [Google Scholar] [CrossRef] [PubMed]
- Araya, J.; Rodrigo, R.; Videla, L.A.; Thielemann, L.; Orellana, M.; Pettinelli, P.; Poniachik, J. Increase in long-chain polyunsaturated fatty acid n − 6/n − 3 ratio in relation to hepatic steatosis in patients with non-alcoholic fatty liver disease. Clin. Sci. 2004, 106, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Puri, P.; Baillie, R.A.; Wiest, M.M.; Mirshahi, F.; Choudhury, J.; Cheung, O.; Sargeant, C.; Contos, M.J.; Sanyal, A.J. A lipidomic analysis of nonalcoholic fatty liver disease. Hepatology 2007, 46, 1081–1090. [Google Scholar] [CrossRef] [PubMed]
- Peng, K.-Y.; Watt, M.J.; Rensen, S.; Greve, J.W.; Huynh, K.; Jayawardana, K.S.; Meikle, P.J.; Meex, R.C. Mitochondrial dysfunction-related lipid changes occur in nonalcoholic fatty liver disease progression. J. Lipid Res. 2018, 59, 1977–1986. [Google Scholar] [CrossRef] [PubMed]
- Chiappini, F.; Coilly, A.; Kadar, H.; Gual, P.; Tran, A.; Desterke, C.; Samuel, D.; Duclos-Vallée, J.-C.; Touboul, D.; Bertrand-Michel, J.; et al. Metabolism dysregulation induces a specific lipid signature of nonalcoholic steatohepatitis in patients. Sci. Rep. 2017, 7, 46658. [Google Scholar] [CrossRef]
- Vance, D.E.; Vance, J.E. Physiological consequences of disruption of mammalian phospholipid biosynthetic genes. J. Lipid Res. 2009, 50, S132–S137. [Google Scholar] [CrossRef]
- Sundler, R.; Akesson, B. Regulation of phospholipid biosynthesis in isolated rat hepatocytes. Effect of different substrates. J. Biol. Chem. 1975, 250, 3359–3367. [Google Scholar] [CrossRef]
- Sakane, S.; Hikita, H.; Shirai, K.; Myojin, Y.; Sasaki, Y.; Kudo, S.; Fukumoto, K.; Mizutani, N.; Tahata, Y.; Makino, Y. White adipose tissue autophagy and adipose-liver crosstalk exacerbate nonalcoholic fatty liver disease in mice. Cell. Mol. Gastroenterol. Hepatol. 2021, 12, 1683–1699. [Google Scholar] [CrossRef]
- Francisco, V.; Sanz, M.J.; Real, J.T.; Marques, P.; Capuozzo, M.; Ait Eldjoudi, D.; Gualillo, O. Adipokines in non-alcoholic fatty liver disease: Are we on the road toward new biomarkers and therapeutic targets? Biology 2022, 11, 1237. [Google Scholar] [CrossRef]
- Zhang, Q.S.; Tian, F.W.; Zhao, J.X.; Zhang, H.; Zhai, Q.X.; Chen, W. The influence of dietary patterns on gut microbiome and its consequences for nonalcoholic fatty liver disease. Trends Food Sci. Technol. 2020, 96, 135–144. [Google Scholar] [CrossRef]
- Ríos-Covián, D.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M.; De Los Reyes-gavilán, C.G.; Salazar, N. Intestinal short chain fatty acids and their link with diet and human health. Front. Microbiol. 2016, 7, 185. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Fan, C.; Li, P.; Lu, Y.; Chang, X.; Qi, K. Short chain fatty acids prevent high-fat-diet-induced obesity in mice by regulating G protein-coupled receptors and gut microbiota. Sci. Rep. 2016, 6, 37589. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Wang, F.; Yuan, J.; Li, J.; Jiang, D.; Zhang, J.; Li, H.; Wang, R.; Tang, J.; Huang, T.; et al. Effects of dietary fat on gut microbiota and faecal metabolites, and their relationship with cardiometabolic risk factors: A 6-month randomised controlled-feeding trial. Gut 2019, 68, 1417–1429. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NO. | Compounds Name | Structural Formula | RT (min) | Chow Control | HFD Control | PPE_L | PPE_H |
|---|---|---|---|---|---|---|---|
| 1 | 9,12-Octadecadienoic acid (Z,Z)- |  | 51.37 | (1.24 ± 0.07) × 107 a | (5.71 ± 0.16) × 107 d | (3.17 ± 0.17) × 107 c | (2.59 ± 0.16) × 107 b |
| 2 | Oleic Acid | 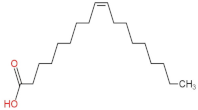 | 48.497 | (3.52 ± 0.24) × 107 a | (3.18 ± 0.61) × 108 d | (2.41 ± 0.30) × 108 c | (1.28 ± 0.10) × 108 b |
| 3 | Octadecanoic acid |  | 47.088 | (3.65 ± 0.17) × 107 a | (3.64 ± 0.31) × 108 c | (2.02 ± 0.48) × 108 b | (1.77 ± 0.22) × 108 b |
| 4 | Heptadecanoic acid |  | 42.723 | (1.62 ± 0.28) × 106 a | (8.51 ± 0.14) × 106 d | (7.16 ± 0.33) × 106 c | (5.08 ± 0.58) × 106 b |
| 5 | n-Hexadecanoic acid |  | 39.389 | (1.57 ± 0.27) × 108 a | (2.58 ± 0.29) × 109 d | (1.89 ± 0.16) × 109 c | (1.60 ± 0.19) × 109 b |
| 6 | Pentadecanoic acid |  | 36.699 | (3.98 ± 0.20) × 106 a | (1.16 ± 0.27) × 107 c | (7.67 ± 0.50) × 106 b | (5.80 ± 0.45) × 106 ab |
| 7 | Tetradecanoic acid |  | 34.56 | (2.72 ± 0.23) × 106 a | (3.33 ± 0.37) × 107 d | (2.04 ± 0.34) × 107 c | (1.54 ± 0.19) × 107 b |
| NO. | Compounds Name | Structural Formula | RT (min) | Chow Control | HFD Control | PPE_L | PPE_H |
|---|---|---|---|---|---|---|---|
| 1 | Pentanoic acid | 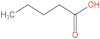 | 16.996 | (2.01 ± 0.55) × 106 a | (1.85 ± 0.47) × 106 a | (3.77 ± 0.66) × 106 b | (3.29 ± 0.65) × 106 b |
| 2 | Butanoic acid | 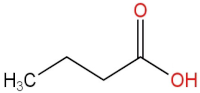 | 14.63 | (5.32 ± 0.40) × 106 a | (5.83 ± 0.67) × 106 a | (1.23 ± 0.17) × 107 c | (8.95 ± 0.74) × 106 b |
| 3 | Propanoic acid, 2-methyl- |  | 13.27 | (9.96 ± 0.67) × 105 a | (1.04 ± 0.10) × 106 a | (1.75 ± 0.50) × 106 b | (1.33 ± 0.21) × 106 a |
| 4 | Propanoic acid | 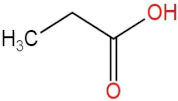 | 12.618 | (4.46 ± 0.40) × 106 a | (6.94 ± 0.35) × 106 b | (9.59 ± 0.70) × 106 c | (7.32 ± 0.70) × 106 b |
| 5 | Acetic acid |  | 10.66 | (4.38 ± 0.36) × 106 a | (5.79 ± 0.66) × 106 b | (8.68 ± 0.62) × 106 d | (6.60 ± 0.43) × 106 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Wu, H.; Zhang, Y.; Hu, C.; Zhen, D.; Fu, P.; He, Y. Phycobiliprotein Peptide Extracts from Arthrospira platensis Ameliorate Nonalcoholic Fatty Liver Disease by Modulating Hepatic Lipid Profile and Strengthening Fat Mobilization. Nutrients 2023, 15, 4573. https://doi.org/10.3390/nu15214573
Liu J, Wu H, Zhang Y, Hu C, Zhen D, Fu P, He Y. Phycobiliprotein Peptide Extracts from Arthrospira platensis Ameliorate Nonalcoholic Fatty Liver Disease by Modulating Hepatic Lipid Profile and Strengthening Fat Mobilization. Nutrients. 2023; 15(21):4573. https://doi.org/10.3390/nu15214573
Chicago/Turabian StyleLiu, Jing, Huan Wu, Yan Zhang, Changbao Hu, Dongyu Zhen, Pengcheng Fu, and Yanfu He. 2023. "Phycobiliprotein Peptide Extracts from Arthrospira platensis Ameliorate Nonalcoholic Fatty Liver Disease by Modulating Hepatic Lipid Profile and Strengthening Fat Mobilization" Nutrients 15, no. 21: 4573. https://doi.org/10.3390/nu15214573
APA StyleLiu, J., Wu, H., Zhang, Y., Hu, C., Zhen, D., Fu, P., & He, Y. (2023). Phycobiliprotein Peptide Extracts from Arthrospira platensis Ameliorate Nonalcoholic Fatty Liver Disease by Modulating Hepatic Lipid Profile and Strengthening Fat Mobilization. Nutrients, 15(21), 4573. https://doi.org/10.3390/nu15214573