
Folate and Vitamin B12 Deficiency Exacerbate Inflammation during Mycobacterium avium paratuberculosis (MAP) Infection


Abstract
1. Introduction
2. Materials and Methods
2.1. Measurement of Plasma Folate and Vitamin B12
2.2. Culture and Infection of Monocyte-Derived Macrophages
2.3. Measurement of IL-1β and TNF-α Expression in Cultured THP-1 Macrophages
2.4. Quantification of THP-1 Macrophage Viability and Apoptosis during MAP Infection
2.5. Culture and Treatment of Caco-2 Monolayers with Infected THP-1 Supernatant
2.6. Quantification of Cytotoxicity and Oxidative Stress in Caco-2 Monolayers
3. Statistical Analysis
4. Results
4.1. Folate and Vitamin B12 Are Reduced in MAP-Positive CD Patients
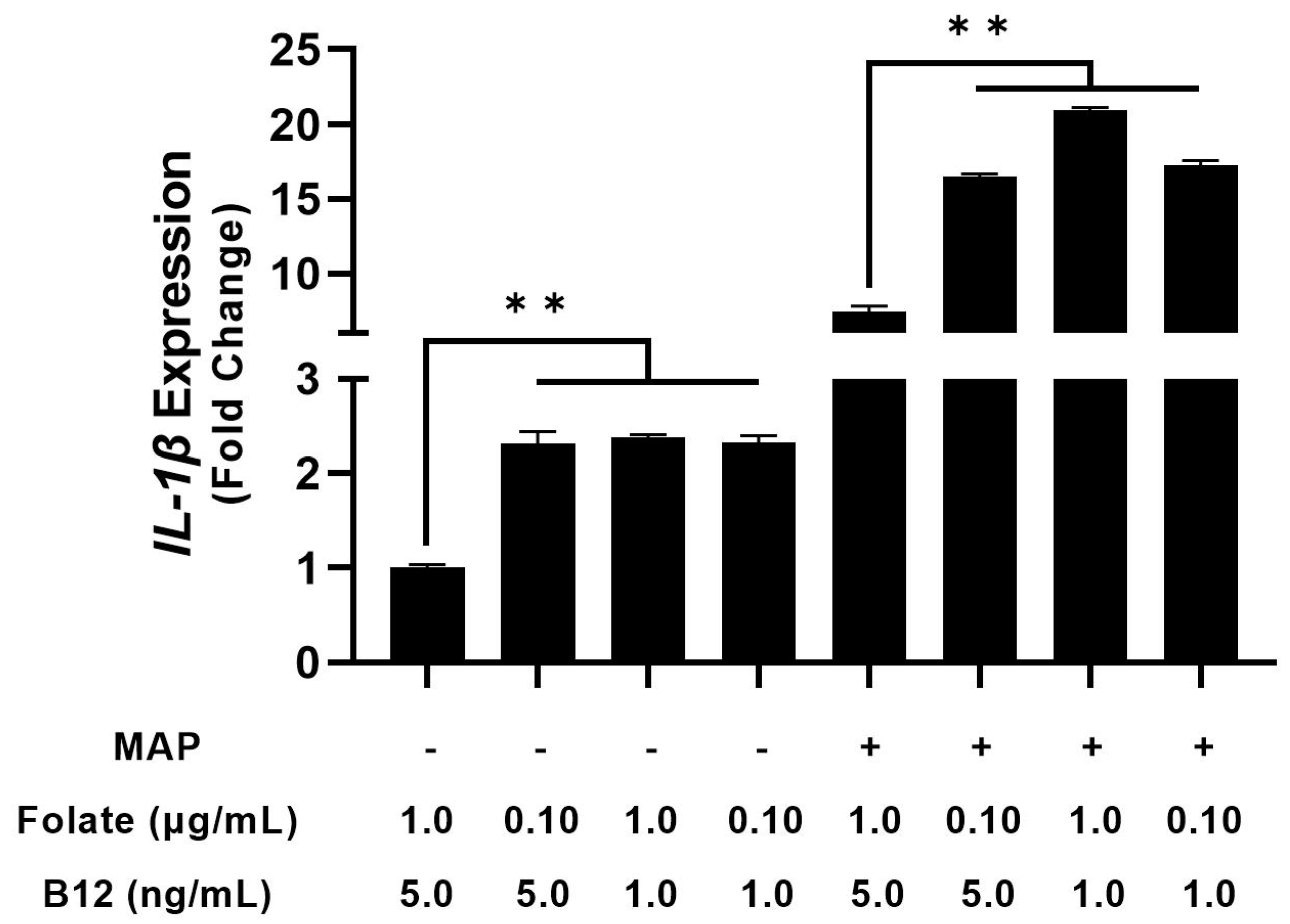
4.2. THP-1 Macrophages Cultured in Folate and B12 Supplemented and Deficient Media Show Altered Cytokine Expression
4.3. MAP Infection Increases Apoptosis during Folate and B12 Supplementation and Decreases Apoptosis during Depletion
4.4. Folate and Vitamin B12 Deficiency Individually Exacerbate Cytotoxicity in Caco-2 Monolayers
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ebara, S. Nutritional role of folate. Congenit. Anom. 2017, 57, 138–141. [Google Scholar] [CrossRef] [PubMed]
- Allen, L.H.; Miller, J.W.; de Groot, L.; Rosenberg, I.H.; Smith, A.D.; Refsum, H.; Raiten, D.J. Biomarkers of Nutrition for Development (BOND): Vitamin B-12 Review. J. Nutr. 2018, 148, 1995s–2027s. [Google Scholar] [CrossRef] [PubMed]
- Mosley, B.S.; Cleves, M.A.; Siega-Riz, A.M.; Shaw, G.M.; Canfield, M.A.; Waller, D.K.; Werler, M.M.; Hobbs, C.A.; for the National Birth Defects Prevention Study. Neural tube defects and maternal folate intake among pregnancies conceived after folic acid fortification in the United States. Am. J. Epidemiol. 2009, 169, 9–17. [Google Scholar] [CrossRef]
- Konings, E.J.M.; Roomans, H.H.S.; Dorant, E.; Goldbohm, R.A.; Saris, W.H.M.; van den Brandt, P.A. Folate intake of the Dutch population according to newly established liquid chromatography data for foods. Am. J. Clin. Nutr. 2001, 73, 765–776. [Google Scholar] [CrossRef]
- Rossi, M.; Amaretti, A.; Raimondi, S. Folate production by probiotic bacteria. Nutrients 2011, 3, 118–134. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, D.K.; Chatterjea, J.B. Serum vitamin B12 in vegetarians. Br. Med. J. 1960, 2, 992–994. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, W.; Schorr, H.; Obeid, R.; Geisel, J. Vitamin B-12 status, particularly holotranscobalamin II and methylmalonic acid concentrations, and hyperhomocysteinemia in vegetarians. Am. J. Clin. Nutr. 2003, 78, 131–136. [Google Scholar] [CrossRef]
- Wilson, R.S.; Mertes, M.P. Chemical model for thymidylate synthetase catalysis. J. Am. Chem. Soc. 1972, 94, 7182–7183. [Google Scholar] [CrossRef]
- Himes, R.H.; Rabinowitz, J.C. Formyltetrahydrofolate synthetase. II. Characteristics of the enzyme and the enzymic reaction. J. Biol. Chem. 1962, 237, 2903–2914. [Google Scholar] [CrossRef]
- Wasserman, G.F.; Mueller, W.T.; Benkovic, S.J.; Liao, W.S.L.; Taylor, J. Evidence that the folate-requiring enzymes of de novo purine biosynthesis are encoded by individual mRNAs. Biochemistry 1984, 23, 6704–6710. [Google Scholar] [CrossRef]
- Mahgoub, A.E.; Deliwala, S.S.; Bachuwa, G. Important reminder: Nitrous oxide (N2O) as a cause of functional vitamin B12 deficiency. BMJ Case Rep. 2021, 14, e241414. [Google Scholar] [CrossRef] [PubMed]
- Moreira, E.S.; Brasch, N.E.; Yun, J. Vitamin B12 protects against superoxide-induced cell injury in human aortic endothelial cells. Free Radic. Biol. Med. 2011, 51, 876–883. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, G.; Velez, L.I. Role of Hydroxocobalamin in Acute Cyanide Poisoning. Ann. Pharmacother. 2008, 42, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Wheatley, C. A scarlet pimpernel for the resolution of inflammation? The role of supra-therapeutic doses of cobalamin, in the treatment of systemic inflammatory response syndrome (SIRS), sepsis, severe sepsis, and septic or traumatic shock. Med. Hypotheses 2006, 67, 124–142. [Google Scholar] [CrossRef] [PubMed]
- Krebs, H.A.; Hems, R.; Tyler, B. The regulation of folate and methionine metabolism. Biochem. J. 1976, 158, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Shou, L.; Pan, F.; Chin, S.-f. Pancreatic Hormones and Hepatic Methionine Adenosyltransferase in the Rat. Proc. Soc. Exp. Biol. Med. 1969, 131, 1012–1018. [Google Scholar] [CrossRef] [PubMed]
- Soda, K. Polyamine Metabolism and Gene Methylation in Conjunction with One-Carbon Metabolism. Int. J. Mol. Sci. 2018, 19, 3106. [Google Scholar] [CrossRef]
- Bressenot, A.; Pooya, S.; Bossenmeyer-Pourie, C.; Gauchotte, G.; Germain, A.; Chevaux, J.-B.; Coste, F.; Vignaud, J.-M.; Guéant, J.-L.; Peyrin-Biroulet, L. Methyl donor deficiency affects small-intestinal differentiation and barrier function in rats. Br. J. Nutr. 2013, 109, 667–677. [Google Scholar] [CrossRef]
- Kunisawa, J.; Hashimoto, E.; Ishikawa, I.; Kiyono, H. A pivotal role of vitamin B9 in the maintenance of regulatory T cells in vitro and in vivo. PLoS ONE 2012, 7, e32094. [Google Scholar] [CrossRef]
- Lee, Y.-J.; Wang, M.-Y.; Lin, M.-C.; Lin, P.-T. Associations between Vitamin B-12 Status and Oxidative Stress and Inflammation in Diabetic Vegetarians and Omnivores. Nutrients 2016, 8, 118. [Google Scholar] [CrossRef]
- Ao, M.; Tsuji, H.; Shide, K.; Kosaka, Y.; Noda, A.; Inagaki, N.; Nakase, H.; Tanaka, K. High prevalence of vitamin B-12 insufficiency in patients with Crohn’s disease. Asia Pac. J. Clin. Nutr. 2017, 26, 1076–1081. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Yang, W.; Wu, C.; Sun, L.; Lin, D.; Lin, X.; Jiang, L.; Ding, R.; Jiang, Y. Association of ulcerative colitis with transcobalamin II gene polymorphisms and serum homocysteine, vitamin B12, and folate levels in Chinese patients. Immunogenetics 2017, 69, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Harb, Z.; Deckert, V.; Bressenot, A.M.; Christov, C.; Guéant-Rodriguez, R.-M.; Raso, J.; Alberto, J.M.; de Barros, J.-P.P.; Umoret, R.; Peyrin-Biroulet, L.; et al. The deficit in folate and vitamin B12 triggers liver macrovesicular steatosis and inflammation in rats with dextran sodium sulfate-induced colitis. J. Nutr. Biochem. 2020, 84, 108415. [Google Scholar] [CrossRef] [PubMed]
- Holland, N.; Harmatz, P.; Golden, D.; Hubbard, A.; Wu, Y.-Y.; Bae, J.; Chen, C.; Huen, K.; Heyman, M.B. Cytogenetic Damage in Blood Lymphocytes and Exfoliated Epithelial Cells of Children with Inflammatory Bowel Disease. Pediatr. Res. 2007, 61, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Kolb, A.F.; Petrie, L. Folate deficiency enhances the inflammatory response of macrophages. Mol. Immunol. 2013, 54, 164–172. [Google Scholar] [CrossRef]
- Madanchi, M.; Fagagnini, S.; Fournier, N.; Biedermann, L.; Zeitz, J.; Battegay, E.; Zimmerli, L.; Vavricka, S.R.; Rogler, G.; Scharl, M.; et al. The Relevance of Vitamin and Iron Deficiency in Patients with Inflammatory Bowel Diseases in Patients of the Swiss IBD Cohort. Inflamm. Bowel Dis. 2018, 24, 1768–1779. [Google Scholar] [CrossRef]
- Scalabrino, G. Vitamin-regulated cytokines and growth factors in the CNS and elsewhere. J. Neurochem. 2009, 111, 1309–1326. [Google Scholar] [CrossRef]
- Arumugam, M.K.; Paal, M.C.; Donohue, T.M., Jr.; Ganesan, M.; Osna, N.A.; Kharbanda, K.K. Beneficial Effects of Betaine: A Comprehensive Review. Biology 2021, 10, 456. [Google Scholar] [CrossRef]
- Leng, Y.-P.; Ma, Y.-S.; Li, X.-G.; Chen, R.-F.; Zeng, P.-Y.; Li, X.-H.; Qiu, C.-F.; Li, Y.-P.; Zhang, Z.; Chen, A.F. L-Homocysteine-induced cathepsin V mediates the vascular endothelial inflammation in hyperhomocysteinaemia. Br. J. Pharmacol. 2018, 175, 1157–1172. [Google Scholar] [CrossRef]
- Samblas, M.; Martínez, J.A.; Milagro, F. Folic Acid Improves the Inflammatory Response in LPS-Activated THP-1 Macrophages. Mediat. Inflamm. 2018, 2018, 1312626. [Google Scholar] [CrossRef]
- Ehmedah, A.; Nedeljkovic, P.; Dacic, S.; Repac, J.; Draskovic-Pavlovic, B.; Vučević, D.; Pekovic, S.; Nedeljkovic, B.B. Effect of Vitamin B Complex Treatment on Macrophages to Schwann Cells Association during Neuroinflammation after Peripheral Nerve Injury. Molecules 2020, 25, 5426. [Google Scholar] [CrossRef] [PubMed]
- Hoffbrand, A.V.; Stewart, J.S.; Booth, C.C.; Mollin, D.L. Folate deficiency in Crohn’s disease: Incidence, pathogenesis, and treatment. Br. Med. J. 1968, 2, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Torres, J.; Mehandru, S.; Colombel, J.-F.; Peyrin-Biroulet, L. Crohn’s disease. Lancet 2017, 389, 1741–1755. [Google Scholar] [CrossRef]
- Bermejo, F.; Algaba, A.; Guerra, I.; Chaparro, M.; De-La-Poza, G.; Valer, P.; Piqueras, B.; Bermejo, A.; García-Alonso, J.; Pérez, M.-J.; et al. Should we monitor vitamin B12 and folate levels in Crohn’s disease patients? Scand. J. Gastroenterol. 2013, 48, 1272–1277. [Google Scholar] [CrossRef] [PubMed]
- Weisshof, R.; Chermesh, I. Micronutrient deficiencies in inflammatory bowel disease. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 576–581. [Google Scholar] [CrossRef] [PubMed]
- Gimier, E.; Chervy, M.; Agus, A.; Sivignon, A.; Billard, E.; Privat, M.; Viala, S.; Minet-Quinard, R.; Buisson, A.; Vazeille, E.; et al. Methyl-donor supplementation prevents intestinal colonization by Adherent-Invasive E. coli in a mouse model of Crohn’s disease. Sci. Rep. 2020, 10, 12922. [Google Scholar] [CrossRef] [PubMed]
- Rathnaiah, G.; Zinniel, D.K.; Bannantine, J.P.; Stabel, J.R.; Gröhn, Y.T.; Collins, M.T.; Barletta, R.G. Pathogenesis, Molecular Genetics, and Genomics of Mycobacterium avium subsp. paratuberculosis, the Etiologic Agent of Johne’s Disease. Front. Vet. Sci. 2017, 4, 187. [Google Scholar] [CrossRef]
- Naser, S.A.; Ghobrial, G.; Romero, C.; Valentine, J.F. Culture of Mycobacterium avium subspecies paratuberculosis from the blood of patients with Crohn’s disease. Lancet 2004, 364, 1039–1044. [Google Scholar] [CrossRef]
- Naser, S.A.; Sagramsingh, S.R.; Naser, A.S.; Thanigachalam, S. Mycobacterium avium subspecies paratuberculosis causes Crohn’s disease in some inflammatory bowel disease patients. World. J. Gastroenterol. 2014, 20, 7403–7415. [Google Scholar] [CrossRef]
- Veauthier, B.; Hornecker, J.R. Crohn’s Disease: Diagnosis and Management. Am. Fam. Physician 2018, 98, 661–669. [Google Scholar]
- Qasem, A.; Naser, A.E.; Naser, S.A. The alternate effects of anti-TNFα therapeutics and their role in mycobacterial granulomatous infection in Crohn’s disease. Expert Rev. Anti-Infect. Ther. 2017, 15, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Qasem, A.; Naser, S.A. TNFα inhibitors exacerbate Mycobacterium paratuberculosis infection in tissue culture: A rationale for poor response of patients with Crohn’s disease to current approved therapy. BMJ Open Gastroenterol. 2018, 5, e000216. [Google Scholar] [CrossRef] [PubMed]
- Qasem, A.; Ramesh, S.; Naser, S.A. Genetic polymorphisms in tumour necrosis factor receptors (TNFRSF1A/1B) illustrate differential treatment response to TNFα inhibitors in patients with Crohn’s disease. BMJ Open Gastroenterol. 2019, 6, e000246. [Google Scholar] [CrossRef] [PubMed]
- Abendaño, N.; Juste, R.A.; Alonso-Hearn, M. Anti-inflammatory and antiapoptotic responses to infection: A common denominator of human and bovine macrophages infected with Mycobacterium avium subsp. paratuberculosis. BioMed. Res. Int. 2013, 2013, 908348. [Google Scholar] [CrossRef]
- Cox, S.R.; Clarke, H.; O’Keeffe, M.; Dubois, P.; Irving, P.M.; Lindsay, J.O.; Whelan, K. Nutrient, Fibre, and FODMAP Intakes and Food-related Quality of Life in Patients with Inflammatory Bowel Disease, and Their Relationship with Gastrointestinal Symptoms of Differing Aetiologies. J. Crohn’s Colitis 2021, 15, 2041–2053. [Google Scholar] [CrossRef]
- Chen, M.; Peyrin-Biroulet, L.; George, A.; Coste, F.; Bressenot, A.; Bossenmeyer-Pourie, C.; Alberto, J.-M.; Xia, B.; Namour, B.; Guéant, J.-L. Methyl deficient diet aggravates experimental colitis in rats. J. Cell. Mol. Med. 2011, 15, 2486–2497. [Google Scholar] [CrossRef]
- Levit, R.; de Giori, G.S.; de Moreno de LeBlanc, A.; LeBlanc, J.G. Folate-producing lactic acid bacteria reduce inflammation in mice with induced intestinal mucositis. J. Appl. Microbiol. 2018, 125, 1494–1501. [Google Scholar] [CrossRef]
- Nelson, J.D.; Haltalin, K.C. Effect of neonatal folic acid deprivation on later growth and susceptibility to Shigella infection in the guinea pig. Am. J. Clin. Nutr. 1972, 25, 992–996. [Google Scholar] [CrossRef]
- Kolho, K.-L.; Pessia, A.; Jaakkola, T.; de Vos, W.M.; Velagapudi, V. Faecal and Serum Metabolomics in Paediatric Inflammatory Bowel Disease. J. Crohn’s Colitis 2017, 11, 321–334. [Google Scholar] [CrossRef]
- Piovani, D.; Danese, S.; Peyrin-Biroulet, L.; Nikolopoulos, G.K.; Lytras, T.; Bonovas, S. Environmental Risk Factors for Inflammatory Bowel Diseases: An Umbrella Review of Meta-analyses. Gastroenterology 2019, 157, 647–659. [Google Scholar] [CrossRef]
- Bain, C.C.; Mowat, A.M. Macrophages in intestinal homeostasis and inflammation. Immunol. Rev. 2014, 260, 102–117. [Google Scholar] [CrossRef] [PubMed]
- Arsenault, R.J.; Maattanen, P.; Daigle, J.; Potter, A.; Griebel, P.; Napper, S. From mouth to macrophage: Mechanisms of innate immune subversion by Mycobacterium avium subsp. paratuberculosis. Vet. Res. 2014, 45, 54. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Culture Condition | Folate (μg/mL) | B12 (ng/mL) |
|---|---|---|
| Control RPMI 1640 | 1.0 | 5.0 |
| Folate-High | 10.0 | 5.0 |
| B12-High | 1.0 | 50.0 |
| Folate + B12-High | 10.0 | 50.0 |
| Folate-Low | 0.10 | 5.0 |
| B12-Low | 1.0 | 1.0 |
| Folate + B12-Low | 0.10 | 1.0 |
| Infection and Treatment | NADP+/(NADPH + NADP+) × 100 ± SD |
|---|---|
| Control (no infection) | 6.23 ± 0.396 |
| MAP infection (1 × 107 CFU/mL) | 6.89 ± 0.761 |
| Folate-low + MAP infection | 7.74 ± 0.750 |
| B12-low + MAP infection | 8.24 ± 0.657 |
| Folate/B12-low + MAP infection | 7.07 ± 0.801 |
| Folate-high + MAP infection | 7.63 ± 1.04 |
| B12-high + MAP infection | 6.59 ± 1.00 |
| Folate/B12-high + MAP | 6.80 ± 0.716 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vaccaro, J.A.; Qasem, A.; Naser, S.A. Folate and Vitamin B12 Deficiency Exacerbate Inflammation during Mycobacterium avium paratuberculosis (MAP) Infection. Nutrients 2023, 15, 261. https://doi.org/10.3390/nu15020261
Vaccaro JA, Qasem A, Naser SA. Folate and Vitamin B12 Deficiency Exacerbate Inflammation during Mycobacterium avium paratuberculosis (MAP) Infection. Nutrients. 2023; 15(2):261. https://doi.org/10.3390/nu15020261
Chicago/Turabian StyleVaccaro, Joseph A., Ahmad Qasem, and Saleh A. Naser. 2023. "Folate and Vitamin B12 Deficiency Exacerbate Inflammation during Mycobacterium avium paratuberculosis (MAP) Infection" Nutrients 15, no. 2: 261. https://doi.org/10.3390/nu15020261
APA StyleVaccaro, J. A., Qasem, A., & Naser, S. A. (2023). Folate and Vitamin B12 Deficiency Exacerbate Inflammation during Mycobacterium avium paratuberculosis (MAP) Infection. Nutrients, 15(2), 261. https://doi.org/10.3390/nu15020261