
Microbiota, Diet and Acute Leukaemia: Tips and Tricks on Their Possible Connections



Abstract
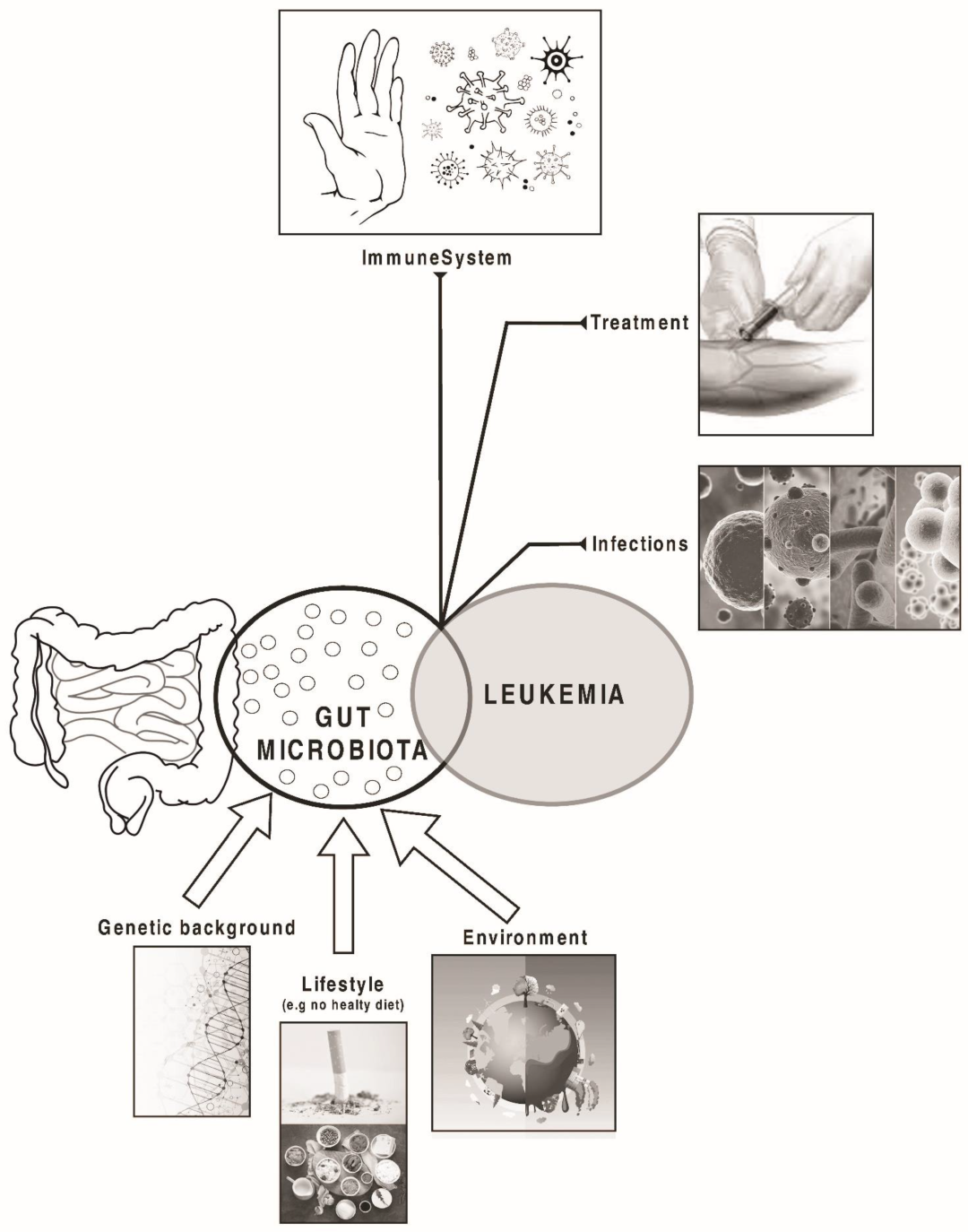
:1. Introduction
2. The Risk of Leukaemia and Diet: Possible Associations
3. Maternal Diet Quality and the Risk of Childhood Leukaemia: What Is the Possible Mechanism?
4. Microbiome and Acute Leukaemia: A Possible Relation?
5. Microbiota and Drug Pharmacokinetics in Leukaemia
6. Vitamin D and Acute Leukaemia
7. Possible Therapeutic Strategies for the Recovery of Microbiota in Leukaemia Patients
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wen, Y.; Jin, R.; Chen, H. Interactions Between Gut Microbiota and Acute Childhood Leukemia. Front. Microbiol. 2019, 10, 1300. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, Y.; Sakata-Haga, H.; Saikawa, Y.; Hatta, T. Influence of Immune System Abnormalities Caused by Maternal Immune Activation in the Postnatal Period. Cells 2023, 12, 741. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Liu, H.-X.; Yan, H.-Y.; Wu, D.-M.; Ping, J. Developmental origins of inflammatory and immune diseases. Mol. Hum. Reprod. 2016, 22, 858–865. [Google Scholar] [CrossRef]
- Belson, M.; Kingsley, B.; Holmes, A. Risk factors for acute leukemia in children: A review. Environ. Health Perspect. 2007, 115, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Deschler, B.; Lübbert, M. Acute myeloid leukaemia: Epidemiology and etiology. Cancer 2006, 107, 2099–2107. [Google Scholar] [CrossRef]
- Shu, X.O.; Potter, J.D.; Linet, M.S.; Severson, R.K.; Han, D.; Kersey, J.H.; Neglia, J.P.; Trigg, M.E.; Robison, L.L. Diagnostic X-rays and ultrasound exposure and risk of childhood acute lymphoblastic leukemia by immunophenotype. Cancer Epidemiol. Biomark. Prev. 2002, 11, 177–185. [Google Scholar]
- Allegra, A.; Tonacci, A.; Pioggia, G.; Musolino, C.; Gangemi, S. Anticancer Activity of Rosmarinus officinalis L.: Mechanisms of Action and Therapeutic Potentials. Nutrients 2020, 12, 1739. [Google Scholar] [CrossRef] [PubMed]
- Steliarova-Foucher, E.; Colombet, M.; Ries, L.A.G.; Moreno, F.; Dolya, A.; Bray, F.; Hesseling, P.; Shin, H.Y.; Stiller, C.A.; Bouzbid, S.; et al. International incidence of childhood cancer, 2001–2010: A population-based registry study. Lancet Oncol. 2017, 18, 719–731. [Google Scholar] [CrossRef] [PubMed]
- Little, M.P.; Wakeford, R.; Borrego, D.; French, B.; Zablotska, L.B.; Adams, M.J.; Allodji, R.; de Vathaire, F.; Lee, C.; Brenner, A.V.; et al. Leukaemia and myeloid malignancy among people exposed to low doses (<100 mSv) of ionising radiation during childhood: A pooled analysis of nine historical cohort studies. Lancet Haematol. 2018, 5, 346–358. [Google Scholar]
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment dominates over host genetics in shaping human gut microbiota. Nature 2018, 555, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, E.; Bargiela, R.; Diez, M.S.; Friedrichs, A.; Perez-Cobas, A.E.; Gosalbes, M.J.; Knecht, H.; Martinez-Martinez, M.; Seifert, J.; von Bergen, M.; et al. Functional consequences of microbial shifts in the human gastrointestinal tract linked to antibiotic treatment and obesity. Gut Microbes 2013, 4, 306–315. [Google Scholar] [CrossRef]
- Perez-Cobas, A.E.; Artacho, A.; Knecht, H.; Ferrus, M.L.; Friedrichs, A.; Ott, S.J.; Moya, A.; Latorre, A.; Gosalbes, M.J. Differential effects of antibiotic therapy on the structure and function of human gut microbiota. PLoS ONE 2013, 8, e80201. [Google Scholar] [CrossRef] [PubMed]
- Janakiraman, M.; Salei, N.; Krishnamoorthy, G. High salt diet does not impact the development of acute myeloid leukemia in mice. Cancer Immunol. Immunother. 2023, 72, 265–273. [Google Scholar] [CrossRef]
- Cao, Y.; Lu, J. Paternal Smoking Before Conception and During Pregnancy Is Associated with an Increased Risk of Childhood Acute Lymphoblastic Leukemia: A Systematic Review and Meta-Analysis of 17 Case-Control Studies. J. Pediatr. Hematol. Oncol. 2020, 42, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Karalexi, M.A.; Dessypris, N.; Thomopoulos, T.P.; Ntouvelis, E.; Kantzanou, M.; Diamantaras, A.A.; Moschovi, M.; Baka, M.; Hatzipantelis, E.; Kourti, M.; et al. Parental alcohol consumption and risk of leukemia in the offspring: A systematic review and meta-analysis. Eur. J. Cancer Prev. 2017, 26, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Metayer, C.; Milne, E.; Dockerty, J.D.; Clavel, J.; Pombo-de-Oliveira, M.S.; Wesseling, C.; Spector, L.G.; Schuz, J.; Petridou, E.; Ezzat, S.; et al. Maternal supplementation with folic acid and other vitamins and risk of leukemia in offspring: A Childhood Leukemia International Consortium study. Epidemiology 2014, 25, 811–822. [Google Scholar] [CrossRef] [PubMed]
- Jensen, C.D.; Block, G.; Buffler, P.; Ma, X.; Selvin, S.; Month, S. Maternal dietary risk factors in childhood acute lymphoblastic leukemia (United States). Cancer Causes Control 2004, 15, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Kwan, M.L.; Jensen, C.D.; Block, G.; Hudes, M.L.; Chu, L.W.; Buffer, P.A. Maternal diet and risk of childhood acute lymphoblastic leukemia. Public. Health Rep. 2009, 124, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Singer, A.W.; Carmichael, S.L.; Selvin, S.; Fu, C.; Block, G.; Metayer, C. Maternal diet quality before pregnancy and risk of childhood leukaemia. Br. J. Nutr. 2016, 116, 1469–1478. [Google Scholar] [CrossRef]
- Fenech, M. The role of folic acid and vitamin B12 in genomic stability of human cells. Mutat. Res. 2001, 475, 57–67. [Google Scholar] [CrossRef]
- Marques, A.H.; O’Connor, T.G.; Roth, R.; Susser, E.; Bjørke-Monsen, A.L. The influence of maternal prenatal and early childhood nutrition and maternal prenatal stress on offspring immune system development and neurodevelopmental disorders. Front. Neurosci. 2013, 7, 120. [Google Scholar] [CrossRef] [PubMed]
- Palmer, A.C. Nutritionally mediated programming of the developing immune system. Adv. Nutr. 2011, 2, 377–395. [Google Scholar] [CrossRef]
- Petridou, E.; Ntouvelis, E.; Dessypris, N.; Terzidis, A.; Trichopoulos, D.; The Childhood Hematology-Oncology Group. Maternal diet and acute lymphoblastic leukemia in young children. Cancer Epidemiol. Biomark. Prev. 2005, 14, 1935–1939. [Google Scholar] [CrossRef] [PubMed]
- Devine, S.M.; Larson, R.A. Acute leukemia in adults: Recent developments in diagnosis and treatment. CA Cancer J. Clin. 1994, 44, 326–352. [Google Scholar] [CrossRef]
- Bonaventure, A.; Rudant, J.; Goujon-Bellec, S.; Orsi, L.; Leverger, G.; Baruchel, A.; Bertrand, Y.; Nelken, B.; Pasquet, M.M.; Michel, G. Childhood acute leukemia, maternal beverage intake during pregnancy, and metabolic polymorphisms. Cancer Causes Control 2013, 24, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Lopez, J.; Iguacel, I.; Pisanu, S.; Almeida, C.C.B.; Steliarova-Foucher, E.; Sierens, C.; Gunter, M.J.; Ladas, E.J.; Barr, R.D.; Van Herck, K.; et al. Role of Maternal Diet in the Risk of Childhood Acute Leukemia: A Systematic Review and Meta-Analysis. Int. J. Environ. Res. Public Health 2023, 20, 5428. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, V.; Lagudu, P.B.B.; Gangopadhyay, D.; Vijaykumar, V.; Rajaraman, S.; PerumalKalaiyarasi, J.; Ganesan, P.; Ganesan, T.S. Neutropenic versus regular diet for acute leukaemia induction chemotherapy: Randomised controlled trial. BMJ Support. Palliat. Care 2022, 12, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Ayub, A.; Ahmad, Q.M.; Javed, T.; Hayat, M.Z.; Farooq, M.A.; Anwar, H.M.Z.; Khan, M.A. Evaluation of diet as a risk factor in the development of childhood leukaemia: A case control study. J. Pak. Med. Assoc. 2020, 70, 404–409. [Google Scholar] [CrossRef] [PubMed]
- Kincaid, J.W.R.; Weiss, G.; Hill-Baskin, A.E.; Schmidt, H.M.; Omoijuanfo, O.; Thompson, C.L.; Beck, C.R.; Berger, N.A. Obesity accelerates acute promyelocyticleukemia in mice and reduces sex differences in latency and penetrance. Obesity 2022, 30, 1420–1429. [Google Scholar] [CrossRef]
- Amitay, E.L.; Keinan-Boker, L. Breastfeeding and Childhood Leukemia Incidence: A Meta-analysis and Systematic Review. JAMA Pediatr. 2015, 169, e151025. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Chang, H.W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15, 73. [Google Scholar] [CrossRef] [PubMed]
- Ponzo, V.; Fedeli, D.; Gozzo, I.; Leone, F.; Lezzo, A.; Monzeglio, C.; Finocchiaro, C.; Ghigo, E.; Bo, S. Diet-GutMicrobiotaInteractions and GestationalDiabetesMellitus (GDM). Nutrients 2019, 11, 330. [Google Scholar] [CrossRef] [PubMed]
- Koren, O.; Goodrich, J.K.; Cullender, T.C.; Spor, A.; Laitinen, K.; Bäckhed, H.K.; Gonzalez, A.; Werner, J.J.; Angenent, L.T.; Knight, R.; et al. Host remodeling of the gut microbiome and metabolic changes during pregnancy. Cell 2012, 150, 470–480. [Google Scholar] [CrossRef]
- Ferretti, P.; Pasolli, E.; Tett, A.; Asnicar, F.; Gorfer, V.; Fedi, S.; Armanini, S.; Truong, D.T.; Manara, S.; Zolfo, M.; et al. Mother-to-infant microbial transmission from different body sites shapes the developing infant gut microbiome. Cell Host Microbe 2018, 24, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Peppas, I.; Ford, A.M.; Furness, C.L.; Greaves, M.F. Gut microbiome immaturity and childhood acute lymphoblastic leukaemia. Nat. Rev. Cancer 2023, 23, 565–576. [Google Scholar] [CrossRef]
- Locasale, J.W. Serine, glycine and one-carbon units: Cancer metabolism in full circle. Nat. Rev. Cancer 2013, 13, 572–583. [Google Scholar] [CrossRef]
- Cantarella, C.D.; Ragusa, D.; Giammanco, M.; Tosi, S. Folate deficiency as predisposing factor for childhood leukaemia: A review of the literature. Genes Nutr. 2017, 12, 14. [Google Scholar] [CrossRef]
- Brown, A.S.; Begg, M.D.; Gravenstein, S.; Schaefer, C.A.; Wyatt, R.J.; Bresnahan, M.; Babulas, V.P.; Susser, E.S. Serologic evidence of prenatal influenza in the etiology of schizophrenia. Arch. Gen. Psychiatry 2004, 61, 774–780. [Google Scholar] [CrossRef] [PubMed]
- Aberg, K.A.; Liu, Y.; Bukszár, J.; McClay, J.L.; Khachane, A.N.; Andreassen, O.A.; Blackwood, D.; Corvin, A.; Djurovic, S.; Gurling, H.; et al. A comprehensive family-based replication study of schizophrenia genes. JAMA Psychiatry 2013, 70, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Tangri, S.; Wegmann, T.G.; Lin, H.; Raghupathy, R. Maternal anti-placental reactivity in natural, immunologically-mediated fetalresorptions. J. Immunol. 1994, 152, 4903–4911. [Google Scholar] [CrossRef] [PubMed]
- Szekeres-Bartho, J. Immunological relationship between the mother and the fetus. Int. Rev. Immunol. 2002, 21, 471–495. [Google Scholar] [CrossRef] [PubMed]
- Breckler, L.A.; Hale, J.; Taylor, A.; Dunstan, J.A.; Thornton, C.A.; Prescott, S.L. Pregnancy IFN-gamma responses to foetal alloantigens are altered by maternal allergy and gravidity status. Allergy 2008, 63, 1473–1480. [Google Scholar] [CrossRef] [PubMed]
- O’Garra, A.; Vieira, P. Regulatory T cells and mechanisms of immune system control. Nat. Med. 2004, 10, 801–805. [Google Scholar] [CrossRef]
- Yasuda, K.; Takeuchi, Y.; Hirota, K. The pathogenicity of Th17 cells in autoimmune diseases. Semin. Immunopathol. 2019, 41, 283–297. [Google Scholar] [CrossRef] [PubMed]
- Imdad, A.; Bhutta, Z.A. Maternal nutrition and birth outcomes: Effect of balanced protein-energy supplementation. Paediatr. Perinat. Epidemiol. 2012, 26, 178–190. [Google Scholar] [CrossRef]
- Black, R.E.; Allen, L.H.; Bhutta, Z.A.; Caulfield, L.E.; de Onis, M.; Ezzati, M.; Mathers, C.; Rivera, J.; Maternal and Child Undernutrition Study Group. Maternal and child undernutrition: Global and regional exposures and health consequences. Lancet 2008, 371, 243–260. [Google Scholar] [CrossRef]
- Ricciardi, L.; Furci, F.; Gangemi, S. Nickel sensitization influence microbiota in allergic and non-allergic disorders: What’s up? J. Biol. Regul. Homeost. Agents. 2021, 35, 757–760. [Google Scholar] [PubMed]
- Ottman, N.; Smidt, H.; de Vos, W.M.; Belzer, C. The function of our microbiota: Who is out there and what do they do? Front. Cell Infect. Microbiol. 2012, 2, 104. [Google Scholar] [CrossRef] [PubMed]
- de Vos, W.M.; Tilg, H.; Van Hul, M.; Cani, P.D. Gut microbiome and health: Mechanistic insights. Gut 2022, 71, 1020–1032. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, J.; Wang, R. Gut microbiota modulates drug pharmacokinetics. Drug Metab. Rev. 2018, 50, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.Y.; Wu, C.Y.; Yu, J. The role of gut microbiota in cancer treatment: Friend or foe? Gut 2020, 69, 1867–1876. [Google Scholar] [CrossRef] [PubMed]
- Vernocchi, P.; Gili, T.; Conte, F.; Del Chierico, F.; Conta, G.; Miccheli, A.; Botticelli, A.; Pacci, P.; Caldarelli, G.; Noti, M.; et al. Network analysis of gut microbiome and metabolome to discover microbiota-linked biomarkers in patients affected by non-small cell lung cancer. Int. J. Mol. Sci. 2020, 21, 8730. [Google Scholar] [CrossRef] [PubMed]
- Guevara-Ramírez, P.; Cadena-Ullauri, S.; Paz-Cruz, E.; Tamayo-Trujillo, R.; Ruiz-Pozo, V.A.; Zambrano, A.K. Role of the gut microbiota in hematgueologic cancer. Front. Microbiol. 2023, 14, 1185787. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, I.I.; FrutosRde, L.; Manel, N.; Yoshinaga, K.; Rifkin, D.B.; Sartor, R.B.; Finlay, B.B.; Littman, D.R. Specific microbiota direct the differentiation of IL-17-producing T-helper cells in the mucosa of the small intestine. Cell Host Microbe 2008, 4, 337–349. [Google Scholar] [CrossRef] [PubMed]
- Gaboriau-Routhiau, V.; Rakotobe, S.; Lecuyer, E.; Mulder, I.; Lan, A.; Bridonneau, C.; Rochet, V.; Pisi, A.; De Paepe, M.; Brandi, G.; et al. The key role of segmented filamentous bacteria in the coordinated maturation of gut helper T cell responses. Immunity 2009, 31, 677–689. [Google Scholar] [CrossRef]
- Tsilimigras, M.C.; Fodor, A.; Jobin, C. Carcinogenesis and therapeutics: The microbiota perspective. Nat. Microbiol. 2017, 2, 17008. [Google Scholar] [CrossRef]
- Song, Y.; Gyarmati, P. Microbiota changes in a pediatric acute lymphocytic leukemia mouse model. Microbiology 2020, 9, 982. [Google Scholar] [CrossRef]
- Rajagopala, S.V.; Yooseph, S.; Harkins, D.M.; Moncera, K.J.; Zabokrtsky, K.B.; Torralba, M.G.; Tovchigrechko, A.; Highlander, S.K.; Pieper, R.; Sender, L.; et al. Gastrointestinal microbial populations can distinguish pediatric and adolescent acute lymphoblastic leukemia (ALL) at the time of disease diagnosis. BMC Genom. 2016, 17, 635. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhou, C.; Zhang, A. Gut microbiota in acute leukemia: Current evidence and future directions. Front. Microbiol. 2022, 13, 1045497. [Google Scholar] [CrossRef]
- Strick, R.; Strissel, P.L.; Borgers, S.; Smith, S.L.; Rowley, J.D. Dietary bioflavonoids induce cleavage in the MLL gene and may contribute to infant leukemia. Proc. Natl. Acad. Sci. USA 2000, 97, 4790–4795. [Google Scholar] [CrossRef]
- Vicente-Dueñas, C.; Janssen, S.; Oldenburg, M.; Auer, F.; González-Herrero, I.; Casado-García, A.; Isidro-Hernandez, M.; Raboso-Gallego, J.; Westhoff, P.; Pandyra, A.A.; et al. An intact gut microbiome protects genetically predisposed mice against leukemia. Blood 2020, 136, 2003–2017. [Google Scholar] [CrossRef] [PubMed]
- Sears, C.L.; Garrett, W.S. Microbes, Microbiota, and Colon Cancer. Cell Host Microbe 2014, 15, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Lavelle, A.; Sokol, H. Gut microbiota-derived metabolites as key actors in inflammatory bowel disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 223–237. [Google Scholar] [CrossRef]
- Matson, V.; Chervin, C.S.; Gajewski, T.F. Cancer and the microbiomeinfluence of the commensal microbiota on cancer, immune responses, and immunotherapy. Gastroenterology 2021, 160, 600–613. [Google Scholar] [CrossRef]
- Hakim, H.; Dallas, R.; Wolf, J.; Tang, L.; Schultz-Cherry, S.; Darling, V.; Johnson, C.; Karlsson, E.A.; Chang, T.C.; Jeha, S.; et al. Gut microbiome composition predicts infection risk during chemotherapy in children with acute lymphoblastic leukemia. Clin. Infect. Dis. 2018, 67, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Chua, L.L.; Rajasuriar, R.; Lim, Y.A.L.; Woo, Y.L.; Loke, P.; Ariffin, H. Temporal changes in gut microbiota profile in children with acute lymphoblastic leukemia prior to commencement-, during-, and post-cessation of chemotherapy. BMC Cancer 2020, 20, 151. [Google Scholar] [CrossRef]
- De Pietri, S.; Ingham, A.C.; Frandsen, T.L.; Rathe, M.; Krych, L.; Castro Mejia, J.L.; Nielsen, D.S.; Nersting, J.; Wehner, P.S.; Schmiegelow, K.; et al. Gastrointestinal toxicity during induction treatment for childhood acute lymphoblastic leukemia: The impact of the gut microbiota. Int. J. Cancer 2020, 147, 1953–1962. [Google Scholar] [CrossRef]
- Nearing, J.T.; Connors, J.; Whitehouse, S.; Van Limbergen, J.; Macdonald, T.; Kulkarni, K.; Langille, M.G.I. Infectious complications are associated with alterations in the gut microbiome in pediatric patients with acute lymphoblastic leukemia. Front. Cell. Infect. Microbiol. 2019, 9, 28. [Google Scholar] [CrossRef] [PubMed]
- Galloway-Pena, J.R.; Smith, D.P.; Sahasrabhojane, P.; Wadsworth, W.D.; Fellman, B.M.; Ajami, N.J.; Shpall, E.J.; Daver, N.; Guindani, M.; Petrosino, J.F.; et al. Characterization of oral and gut microbiome temporal variability in hospitalized cancer patients. Genome Med. 2017, 9, 21. [Google Scholar] [CrossRef] [PubMed]
- Lucafo, M.; Franzin, M.; Lagatolla, C.; Franca, R.; Bramuzzo, M.; Stocco, G.; Decorti, G. Emerging insights on the interaction between anticancer and immunosuppressant drugs and intestinal microbiota in pediatric patients. Clin. Transl. Sci. 2020, 13, 238–259. [Google Scholar] [CrossRef]
- Zhou, B.; Xia, X.; Wang, P.; Chen, S.; Yu, C.; Huang, R.; Zhang, R.; Wang, Y.; Lu, L.; Yuan, F.; et al. Induction and amelioration of methotrexate-induced gastrointestinal toxicity are related to immune response and gut microbiota. EBioMedicine 2018, 33, 122–133. [Google Scholar] [CrossRef] [PubMed]
- Viaud, S.; Saccheri, F.; Mignot, G.; Yamazaki, T.; Daillere, R.; Hannani, D.; Enot, D.P.; Pfirschke, C.; Engblom, C.; Pittet, M.J.; et al. The intestinal microbiota modulates the anticancer immune effects of cyclophosphamide. Science 2013, 342, 971–976. [Google Scholar] [CrossRef] [PubMed]
- Daillère, R.; Vétizou, M.; Waldschmitt, N.; Yamazaki, T.; Isnard, C.; Poirier-Colame, V.; Duong, C.P.M.; Flament, C.; Lepage, P.; Roberti, P.P.; et al. Enterococcus hirae and Barnesiellaintestinihominis facilitate cyclophosphamide-induced therapeutic immunomodulatory effects. Immunity 2016, 45, 931–943. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M.; Zimmermann-Kogadeeva, M.; Wegmann, R.; Goodman, A.L. Mapping human microbiome drug metabolism by gut bacteria and their genes. Nature 2019, 570, 462–467. [Google Scholar] [CrossRef]
- Smith, M.; Dai, A.; Ghilardi, G.; Amelsberg, K.V.; Devlin, S.M.; Pajarillo, R.; Slingerland, J.B.; Beghi, S.; Herrera, P.S.; Giardina, P.; et al. Gut microbiome correlates of response and toxicity following anti-CD19 CAR T cell therapy. Nat. Med. 2022, 28, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Medrano, M.; Carrillo-Cruz, E.; Montero, I.; Perez-Simon, J.A. Vitamin D: Effect on Haematopoiesis and Immune System and Clinical Applications. Int. J. Mol. Sci. 2018, 19, 2663. [Google Scholar] [CrossRef] [PubMed]
- Murdaca, G.; Gerosa, A.; Paladin, F.; Petrocchi, L.; Banchero, S.; Gangemi, S. Vitamin D and Microbiota: IsThere a Link with Allergies? Int. J. Mol. Sci. 2021, 22, 4288. [Google Scholar] [CrossRef]
- Fakhoury, H.M.A.; Kvietys, P.R.; AlKattan, W.; Anouti, F.A.; Elahi, M.A.; Karras, S.N.; Grant, W.B. Vitamin D and intestinal homeostasis: Barrier, microbiota, and immune modulation. J. Steroid Biochem. Mol. Biol. 2020, 200, 105663. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Abe, E.; Miyaura, C.; Shiina, Y.; Suda, T. 1α, 25-dihydroxyvitamin D3 induces differentiation of human promyelocyticleukemia cells (HL-60) into monocyte- macrophages, but not into granulocytes. Biochem. Biophys. Res. Commun. 1983, 117, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Muto, A.; Kizaki, M.; Yamato, K.; Kawai, Y.; Kamata-Matsushita, M.; Ueno, H.; Ohguchi, M.; Nishihara, T.; Koeffler, H.P.; Ikeda, Y. 1, 25-dihydroxyvitamin D3 induces differentiation of a retinoic acid–resistant acute promyelocyticleukemia cell line (UF-1) associated with expression of p21 WAF1/CIP1 and p27 KIP1. Blood 1999, 93, 2225–2233. [Google Scholar] [CrossRef]
- Abe, E.; Miyaura, C.; Sakagami, H.; Takeda, M.; Konno, K.; Yamazaki, T.; Yoshiki, S.; Suda, T. Differentiation of mouse myeloid leukemia cells induced by 1α, 25-dihydroxyvitamin D3. Proc. Natl. Acad. Sci. USA 1981, 78, 4990–4994. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Studzinski, G.P. Retinoblastoma protein and CCAAT/enhancer-binding protein β are required for 1, 25-dihydroxyvitamin D3-induced monocytic differentiation of HL60 cells. Cancer Res. 2004, 64, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Motomura, S.; Kanamori, H.; Maruta, A.; Kodama, F.; Ohkubo, T. The effect of 1-hydroxyvitamin D3 for prolongation of leukemic transformation-free survival in myelodysplastic syndromes. Am. J. Hematol. 1991, 38, 67–68. [Google Scholar] [CrossRef]
- Siitonen, T.; Timonen, T.; Juvonen, E.; Terävä, V.; Kutila, A.; Honkanen, T.; Mikkola, M.; Hallman, H.; Kauppila, M.; Nyländen, P.; et al. Valproic acid combined with 13-cis retinoic acid and 1, 25-dihydroxyvitamin D3 in the treatment of patients with myelodysplastic syndromes. Haematologica 2007, 92, 1119–1122. [Google Scholar] [CrossRef]
- Gemelli, C.; Orlandi, C.; Zanocco Marani, T.; Martello, A.; Vignudelli, T.; Ferrari, F.; Montanari, M.; Parenti, S.; Testa, A.; Grande, A.; et al. The vitamin D3/Hox-A10 pathway supports MafB function during the monocyte differentiation of human CD34+ hemopoietic progenitors. J. Immunol. 2008, 181, 5660–5672. [Google Scholar] [CrossRef]
- Callens, C.; Coulon, S.; Naudin, J.; Radford-Weiss, I.; Boissel, N.; Raffoux, E.; Wang, P.H.; Agarwal, S.; Tamouza, H.; Paubelle, E.; et al. Targeting iron homeostasis induces cellular differentiation and synergizes with differentiating agents in acute myeloid leukemia. J. Exp. Med. 2010, 207, 731–750. [Google Scholar] [CrossRef]
- Harrison, J.S.; Bershadskiy, A. Clinical experience using vitamin D and analogs in the treatment of myelodysplasia and acute myeloid leukemia: A review of the literature. Leuk. Res. Treat. 2012, 2012, 125814. [Google Scholar] [CrossRef] [PubMed]
- Reyna-Figueroa, J.; Barron-Calvillo, E.; Garcia-Parra, C.; Galindo-Delgado, P.; Contreras-Ochoa, C.; Lagunas-Martinez, A.; Campos-Romero, F.H.; Silva-Estrada, J.A.; Limon-Rojas, A.E. Probiotic supplementation decreases chemotherapy-induced gastrointestinal side effects in patients with acute leukemia. J. Pediatr. Hematol. Oncol. 2019, 41, 468–472. [Google Scholar] [CrossRef] [PubMed]
- Vindigni, S.M.; Surawicz, C.M. Fecalmicrobiota transplantation. Gastroenterol. Clin. N. Am. 2017, 46, 171–185. [Google Scholar] [CrossRef] [PubMed]
- Tkach, S.; Dorofeyev, A.; Kuzenko, I.; Boyko, N.; Falalyeyeva, T.; Boccuto, L.; Scarpellini, E.; Kobyliak, N.; Abenavoli, L. Current status and future therapeutic options for fecalmicrobiota. Medicina 2022, 58, 84. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.W.; Kuo, C.H.; Kuo, F.C.; Wang, Y.K.; Hsu, W.H.; Yu, F.J.; Hu, H.M.; Hsu, P.I.; Wang, J.Y.; Wu, D.C. Fecalmicrobiota transplantation: Review and update. J. Formos. Med. Assoc. 2019, 118, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Wang, P.; Yan, J.; Liu, G.; Zeng, B.; Hussain, T.; Peng, C.; Yin, J.; Li, T.; Wei, H.; et al. Melatonin alleviates weanling stress in mice: Involvement of intestinal microbiota. J. Pineal Res. 2018, 64, e12448. [Google Scholar] [CrossRef]
- Ishibashi, N.; Yamazaki, S. Probiotics and safety. Am. J. Clin. Nutr. 2001, 73, 46–70. [Google Scholar] [CrossRef] [PubMed]
- Allegra, A.; Musolino, C.; Tonacci, A.; Pioggia, G.; Gangemi, S. Interactions between the MicroRNAs and Microbiota in Cancer Development: Roles and Therapeutic Opportunities. Cancers 2020, 12, 805. [Google Scholar] [CrossRef] [PubMed]
- Furci, F.; Allegra, A.; Tonacci, A.; Isola, S.; Senna, G.; Pioggia, G.; Gangemi, S. Air Pollution and microRNAs: The Role of Association in Airway Inflammation. Life 2023, 13, 1375. [Google Scholar] [CrossRef]
- De Martinis, M.; Ginaldi, L.; Allegra, A.; Sirufo, M.M.; Pioggia, G.; Tonacci, A.; Gangemi, S. The Osteoporosis/MicrobiotaLinkage: The Role of miRNA. Int. J. Mol. Sci. 2020, 21, 8887. [Google Scholar] [CrossRef]
- Hermetet, F.; Mshaik, R.; Simonet, J.; Callier, P.; Delva, L.; Quéré, R. High-fat diet intensifies MLL-AF9-induced acute myeloid leukaemia through activation of the FLT3 signalling in mouse primitive hematopoietic cells. Sci. Rep. 2020, 10, 16187. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Shen, N.; Pang, J.X.; Zhang, Y.W.; Rao, E.Y.; Bode, A.M.; Al-Kali, A.; Zhang, D.E.; Litzow, M.R.; Li, B.; et al. Fatty acid-binding protein FABP4 mechanistically links obesity with aggressive AML by enhancing aberrant DNA methylation in AML cells. Leukemia 2017, 31, 1434–1442. [Google Scholar] [CrossRef] [PubMed]
- Yamamura, Y.; Oum, R.; Gbito, K.Y.; Garcia-Manero, G.; Strom, S.S. Dietary intake of vegetables, fruits, and meats/beans as potential risk factors of acute myeloid leukemia: A Texas case-control study. Nutr. Cancer. 2013, 65, 1132–1140. [Google Scholar] [CrossRef]
- Lanoue, L.; Green, K.K.; Kwik-Uribe, C.; Keen, C.L. Dietary factors and the risk for acute infant leukemia: Evaluating the effects of cocoa-derived flavanols on DNA topoisomerase activity. ExpBiol. Med. 2010, 235, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Park, Y.; Mayne, S.T.; Wang, R.; Sinha, R.; Hollenbeck, A.R.; Schatzkin, A.; Cross, A.J. Diet, lifestyle, and acute myeloid leukemia in the NIH-AARP cohort. Am. J. Epidemiol. 2010, 171, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.A. Maternal diet and infant leukaemia: A role for DNA topoisomerase II inhibitors? Int. J. Cancer Suppl. 1998, 11, 26–28. [Google Scholar] [CrossRef]
- Li, Y.; Moysich, K.B.; Baer, M.R.; Weiss, J.R.; Brasure, J.; Graham, S.; McCann, S.E. Intake of selected food groups and beverages and adult acute myeloid leukaemia. Leuk. Res. 2006, 30, 1507–1515. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.A.; Kasum, C.M.; Davies, S.M.; Jacobs, D.R.; Folsom, A.R.; Potter, J.D. Diet and risk of leukaemia in the Iowa Women’s Health Study. Cancer Epidemiol. Biomark. Prev. 2002, 11, 777–781. [Google Scholar]
- Kwiatkowski, A. Dietary and other environmental risk factors in acute leukaemias: A case-control study of 119 patients. Eur. J. Cancer Prev. 1993, 2, 139–146. [Google Scholar] [CrossRef]
- Greaves, M.; Cazzaniga, V.; Ford, A. Can. we prevent childhood Leukaemia? Leukemia 2021, 35, 1258–1264. [Google Scholar] [CrossRef]
- Moore, R.E.; Townsend, S.D. Temporal development of the infant gut microbiome. Open Biol. 2019, 9, 190128. [Google Scholar] [CrossRef]
- Tamburini, S.; Shen, N.; Wu, H.C.; Clemente, J.C. The microbiome in early life: Implications for health outcomes. Nat. Med. 2016, 22, 713–722. [Google Scholar] [CrossRef]
- Hooper, L.V.; Littman, D.R.; Macpherson, A.J. Interactions between the microbiota and the immune system. Science 2012, 336, 1268–1273. [Google Scholar] [CrossRef]
- Levy, M.; Kolodziejczyk, A.A.; Thaiss, C.A.; Elinav, E. Dysbiosis and the immune system. Nat. Rev. Immunol. 2017, 17, 219–232. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Authors | Paper | Main Results |
|---|---|---|
| [13] | High salt diet does not impact the development of acute myeloid leukaemia in mice. | A high-salt diet induces a significant change in microbiota composition, TH17 responses, and NK cells, not inducing a significant role in tumour development. |
| [19] | Maternal diet quality before pregnancy and risk of childhood leukaemia. | Higher maternal diet quality score was associated with a reduced risk of acute myeloid leukaemia. |
| [26] | Role of Maternal Diet in the Risk of Childhood Acute Leukemia: A Systematic Review and Meta-Analysis. | Maternal consumption of fruit was inversely associated with acute lymphoblastic leukaemia, while maternal coffee intake was associated with an increased risk of acute lymphoblastic leukaemia. |
| [27] | Neutropenic versus regular diet for acute leukaemia induction chemotherapy: randomised controlled trial. | A neutropenic diet did not play a preventive role in infections, reducing mortality or changing stool microbiota composition in patients with acute leukaemia. |
| [28] | Evaluation of diet as a risk factor in the development of childhood leukaemia: a case-control study. | Higher consumption of caffeinated drinks and junk food was reported in children of either gender aged 2–12 years with acute lymphocytic or acute myelocytic leukaemia. |
| [29] | Obesity accelerates acute promyelocytic leukaemia in mice and reduces sex differences in latency and penetrance. | In vivo, obesity and obesogenic diet play a key role in promoting the development of acute promyelocytic leukaemia. |
| [61] | Dietary bioflavonoids induce cleavage in the MLL gene and may contribute to infant leukaemia. | Maternal ingestion of bioflavonoids may induce MLL breaks and may induce translocations in utero, causing infant and childhood leukaemia. |
| [98] | High-fat diet intensifies MLL-AF9-induced acute myeloid leukaemia through activation of the FLT3 signalling in mouse primitive hematopoietic cells. | Using an MLL-AF9 knock-in mouse model, consumption of a high-fat diet accelerates the risk of developing acute myeloid leukaemia, with increased clusterization of FLT3 within lipid rafts on cell surface of primitive hematopoietic cells and consequent overactivation of JAK/STAT pathway. |
| [99] | Fatty acid-binding protein FABP4 mechanistically links obesity with aggressive AML by enhancing aberrant DNA methylation in AML cells. | Leukaemia burden was much higher in high-fat diet-induced obese mice, in which increased levels of FABP4 and interleukin (IL)-6 in sera were described. |
| [100] | Dietary intake of vegetables, fruits, and meats/beans as potential risk factors of acute myeloid leukaemia: a Texas case-control study. | A decreased risk of acute myeloid leukaemia was reported among patients who consumed higher quantities of dark green vegetables, seafood, and nuts/seeds. The risk was instead significantly increased among greatest consumers of red meat. |
| [101] | Dietary factors and the risk for acute infant leukaemia: evaluating the effects of cocoa-derived flavanols on DNA topoisomerase activity. | In vitro data reported that cocoa-derived flavanols have limited effects on topo II activity and cellular proliferation in cancer cell lines, with a leukemogenic potential at physiological concentrations. |
| [102] | Diet, lifestyle, and acute myeloid leukaemia in the NIH-AARP cohort. | Increased risk of acute myeloid leukaemia was related to a higher meat intake; patients who did not drink coffee appeared to have a higher risk of acute myeloid leukaemia than those who drank various quantities of coffee; the intake of fruit and vegetables was not related to acute myeloid leukaemia. |
| [103] | Maternal diet and infant leukaemia: a role for DNA topoisomerase II inhibitors? | There was no positive correlation with increasing maternal assumption of DNA topo 2 inhibitor-containing foods (specific fruits and vegetables, and in soy, coffee, wine, tea and cocoa) neither for the overall group nor for infants in the acute lymphoblastic leukaemia stratum. |
| [104] | Intake of selected food groups and beverages and adult acute myeloid leukaemia. | The risk of AML was negatively related to the assumption of milk and tea among women and positively associated with the intake of beer, wine, and beef. |
| [105] | Diet and risk of leukaemia in the Iowa Women’s Health Study. | Risk of adult leukaemia may be decreased by increased vegetable consumption. |
| [106] | Dietary and other environmental risk factors in acute leukaemias: a case-control study of 119 patients. | Low consumption of raw vegetables, frequent drinking of milk, frequent consumption of poultry, and drinking of soft water were related to an increased risk of acute leukaemia. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Furci, F.; Cicero, N.; Allegra, A.; Gangemi, S. Microbiota, Diet and Acute Leukaemia: Tips and Tricks on Their Possible Connections. Nutrients 2023, 15, 4253. https://doi.org/10.3390/nu15194253
Furci F, Cicero N, Allegra A, Gangemi S. Microbiota, Diet and Acute Leukaemia: Tips and Tricks on Their Possible Connections. Nutrients. 2023; 15(19):4253. https://doi.org/10.3390/nu15194253
Chicago/Turabian StyleFurci, Fabiana, Nicola Cicero, Alessandro Allegra, and Sebastiano Gangemi. 2023. "Microbiota, Diet and Acute Leukaemia: Tips and Tricks on Their Possible Connections" Nutrients 15, no. 19: 4253. https://doi.org/10.3390/nu15194253
APA StyleFurci, F., Cicero, N., Allegra, A., & Gangemi, S. (2023). Microbiota, Diet and Acute Leukaemia: Tips and Tricks on Their Possible Connections. Nutrients, 15(19), 4253. https://doi.org/10.3390/nu15194253