Association of Plasma Vitamins and Carotenoids, DNA Methylation of LCAT, and Risk of Age-Related Macular Degeneration

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Measurement of Circulating Vitamins and Carotenoids
2.3. DNA Methylation Detection of LCAT
2.4. Assessment of Covariates
2.5. Statistical Analysis
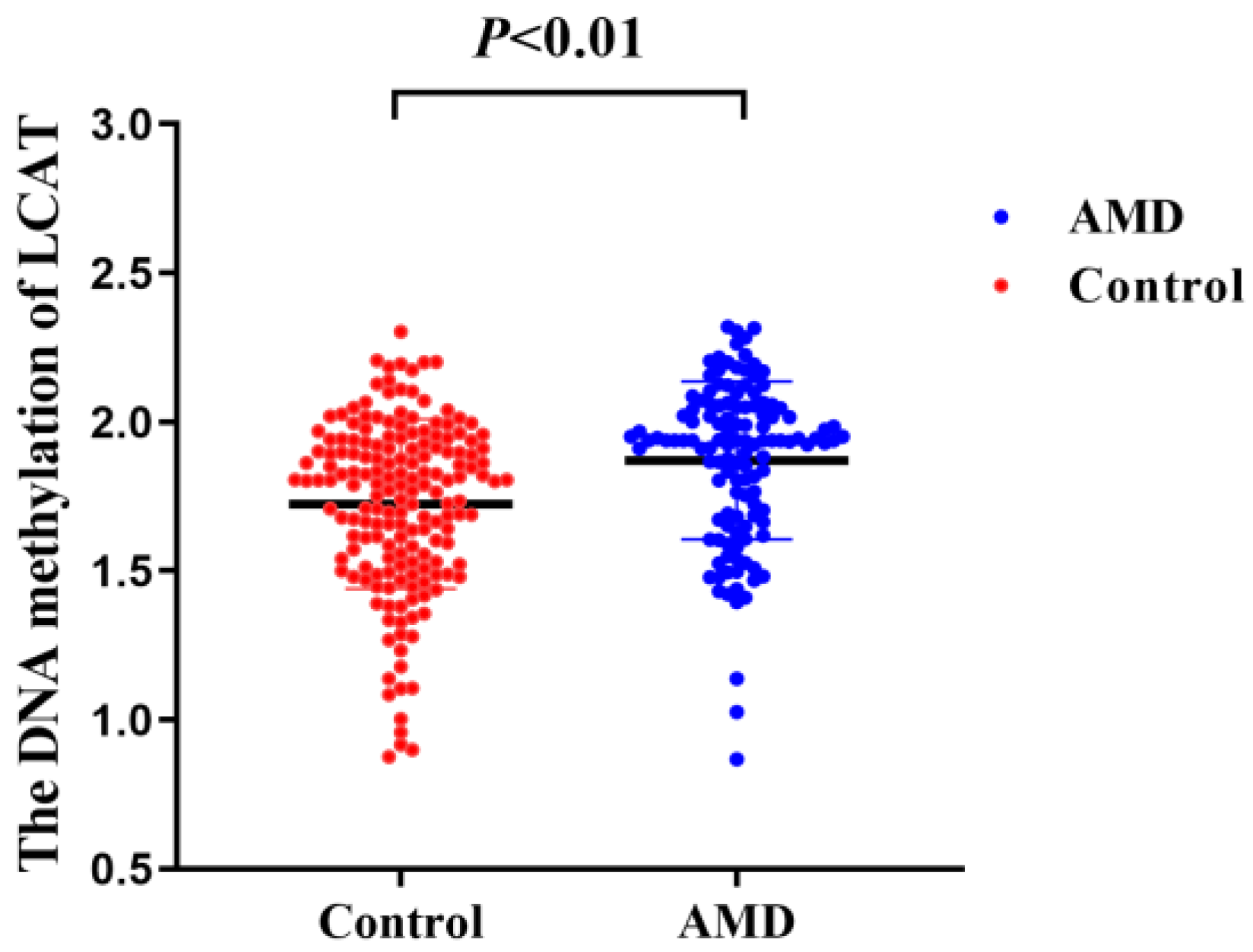
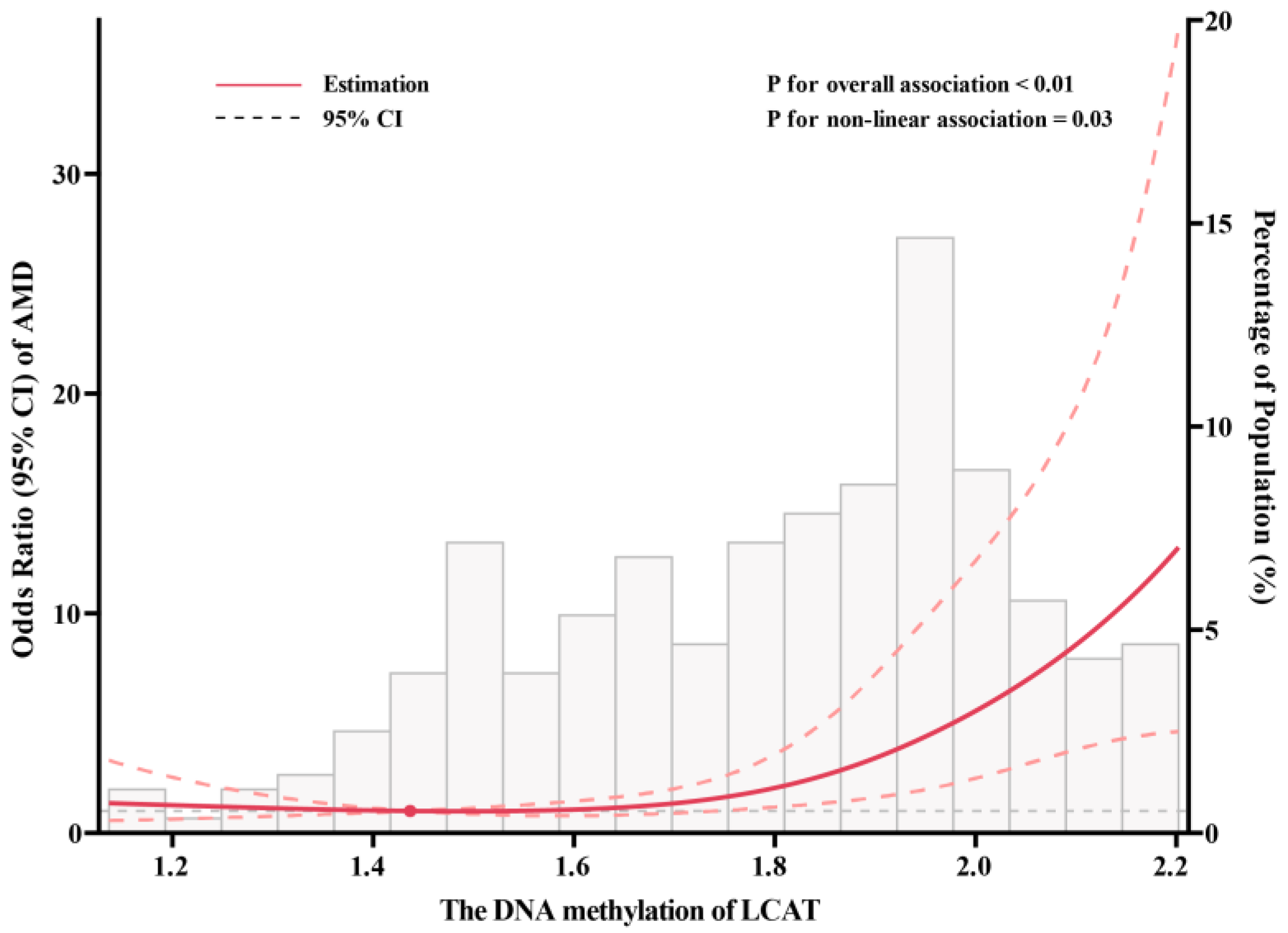
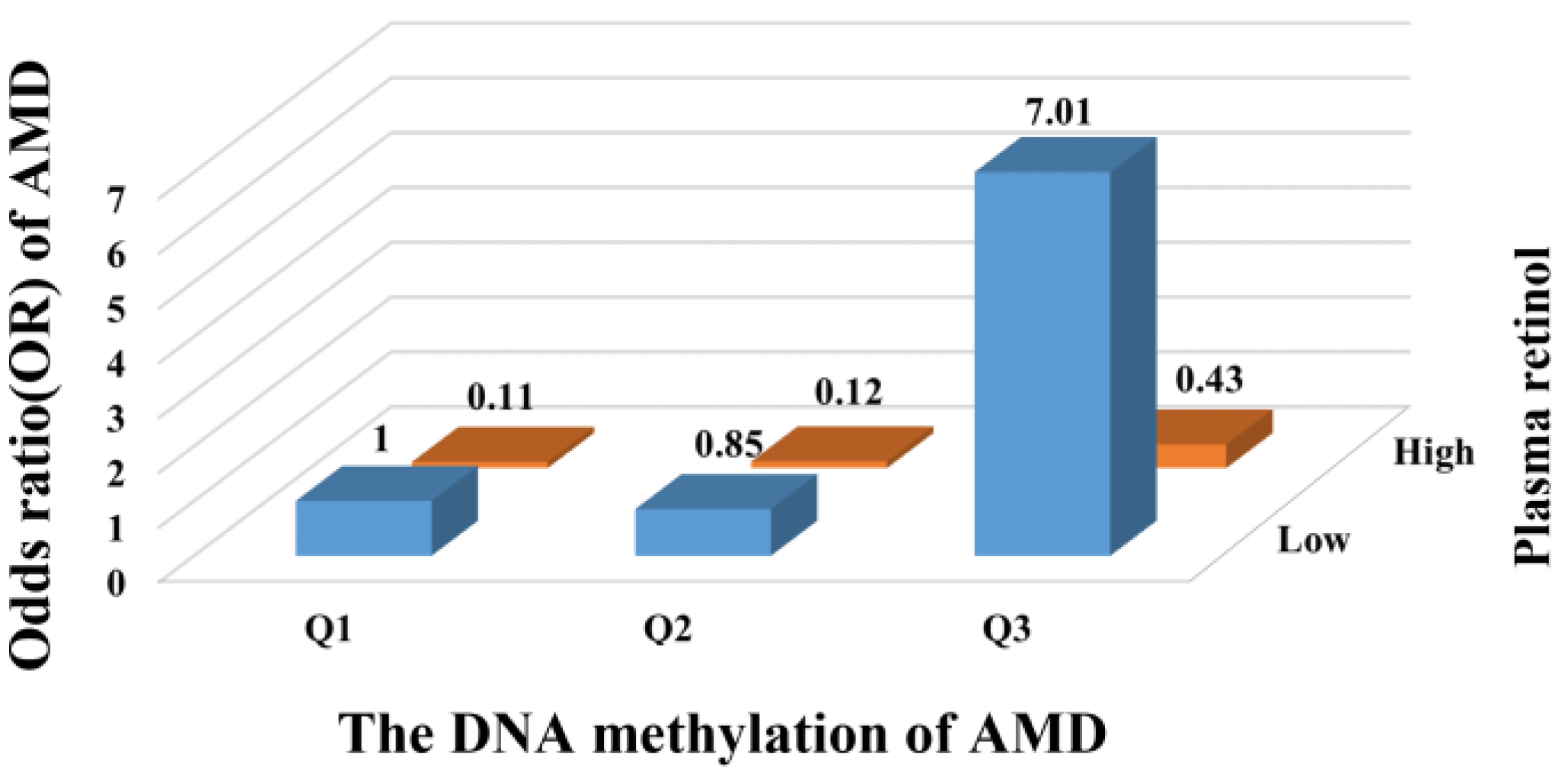
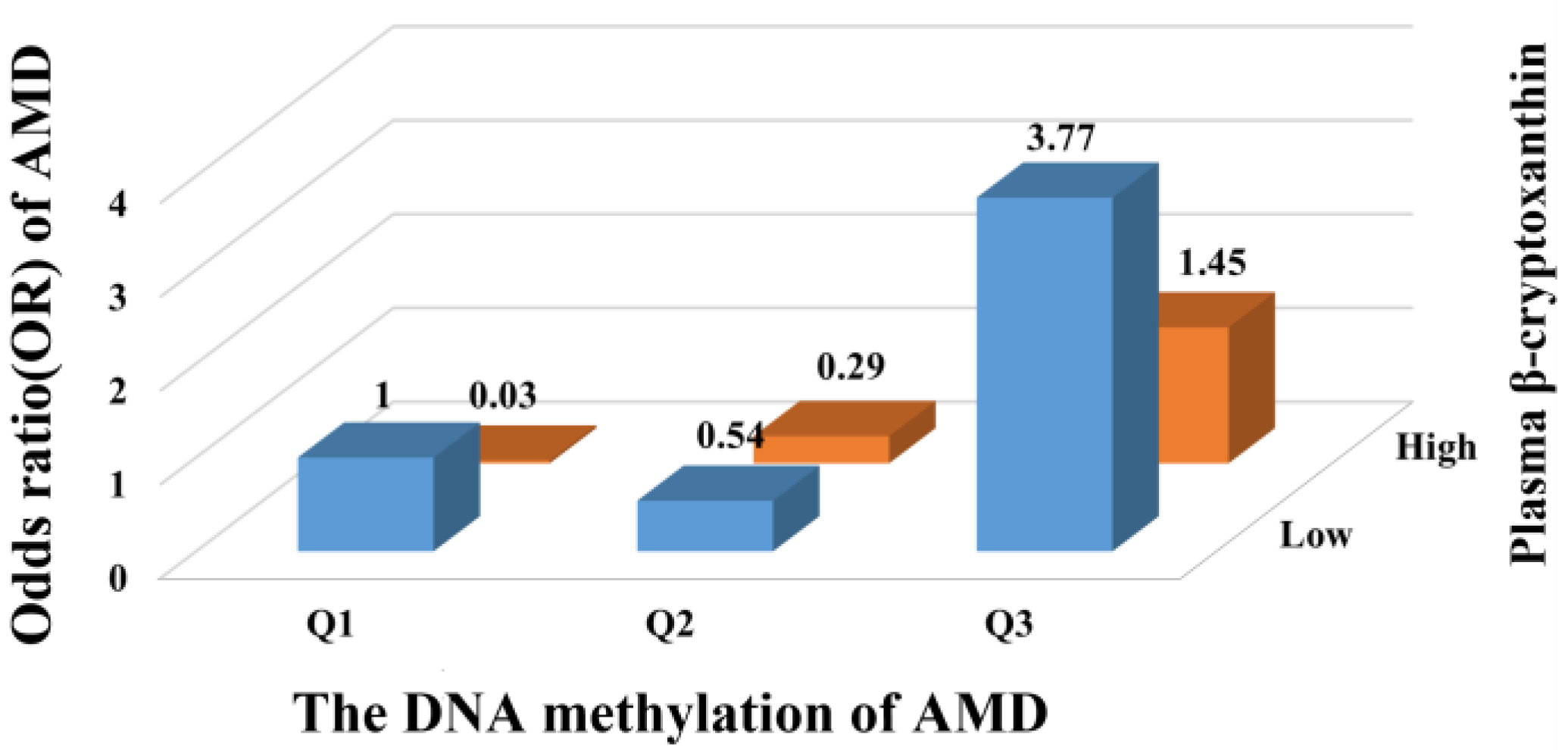
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wong, W.L.; Su, X.; Li, X.; Cheung, C.M.; Klein, R.; Cheng, C.Y.; Wong, T.Y. Global prevalence of age-related macular degeneration and disease burden projection for 2020 and 2040: A systematic review and meta-analysis. Lancet Glob. Health 2014, 2, e106–e116. [Google Scholar] [CrossRef] [PubMed]
- Fleckenstein, M.; Keenan, T.; Guymer, R.H.; Chakravarthy, U.; Schmitz-Valckenberg, S.; Klaver, C.C.; Wong, W.T.; Chew, E.Y. Age-related macular degeneration. Nat. Rev. Dis. Prim. 2021, 7, 31. [Google Scholar] [CrossRef] [PubMed]
- Holleboom, A.G.; Daniil, G.; Fu, X.; Zhang, R.; Hovingh, G.K.; Schimmel, A.W.; Kastelein, J.J.; Stroes, E.S.; Witztum, J.L.; Hutten, B.A.; et al. Lipid oxidation in carriers of lecithin:cholesterol acyltransferase gene mutations. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 3066–3075. [Google Scholar] [CrossRef]
- Goyal, J.; Wang, K.; Liu, M.; Subbaiah, P.V. Novel function of lecithin-cholesterol acyltransferase: Hydrolysis of oxidized polar phospholipids generated during lipoprotein oxidation. J. Biol. Chem. 1997, 272, 16231–16239. [Google Scholar] [CrossRef]
- Yang, K.; Wang, J.; Xiang, H.; Ding, P.; Wu, T.; Ji, G. LCAT-targeted therapies: Progress, failures and future. Biomed. Pharmacother. 2022, 147, 112677. [Google Scholar] [CrossRef]
- Bonaca, M.P.; George, R.T.; Morrow, D.A.; Bergmark, B.A.; Park, J.G.; Abuhatzira, L.; Vavere, A.L.; Karathanasis, S.K.; Jin, C.; She, D.; et al. Recombinant human lecithin-cholesterol acyltransferase in patients with atherosclerosis: Phase 2a primary results and phase 2b design. Eur. Heart J.-Cardiovasc. Pharmacother. 2022, 8, 243–252. [Google Scholar] [CrossRef]
- Sasaki, M.; Delawary, M.; Sakurai, H.; Kobayashi, H.; Nakao, N.; Tsuru, H.; Fukushima, Y.; Honzumi, S.; Moriyama, S.; Wada, N.; et al. Novel LCAT (Lecithin:Cholesterol Acyltransferase) Activator DS-8190a Prevents the Progression of Plaque Accumulation in Atherosclerosis Models. Arterioscler. Thromb. Vasc. Biol. 2021, 41, 360–376. [Google Scholar] [CrossRef]
- Ossoli, A.; Simonelli, S.; Varrenti, M.; Morici, N.; Oliva, F.; Stucchi, M.; Gomaraschi, M.; Strazzella, A.; Arnaboldi, L.; Thomas, M.J.; et al. Recombinant LCAT (Lecithin:Cholesterol Acyltransferase) Rescues Defective HDL (High-Density Lipoprotein)-Mediated Endothelial Protection in Acute Coronary Syndrome. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 915–924. [Google Scholar] [CrossRef]
- He, D.; Liao, S.; Cai, L.; Huang, W.; Xie, X.; You, M. Integrated analysis of methylation-driven genes and pretreatment prognostic factors in patients with hepatocellular carcinoma. BMC Cancer 2021, 21, 599. [Google Scholar] [CrossRef]
- Seale, K.; Horvath, S.; Teschendorff, A.; Eynon, N.; Voisin, S. Making sense of the ageing methylome. Nat. Rev. Genet. 2022, 23, 585–605. [Google Scholar] [CrossRef]
- Stahl, W.; Sies, H. Antioxidant activity of carotenoids. Mol. Asp. Med. 2003, 24, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.R.; Lawrenson, J.G. Antioxidant vitamin and mineral supplements for slowing the progression of age-related macular degeneration. Cochrane Database Syst. Rev. 2012, 11, CD000254. [Google Scholar] [PubMed]
- Beetch, M.; Harandi-Zadeh, S.; Shen, K.; Lubecka, K.; Kitts, D.D.; O’Hagan, H.M.; Stefanska, B. Dietary antioxidants remodel DNA methylation patterns in chronic disease. Br. J. Pharmacol. 2020, 177, 1382–1408. [Google Scholar] [CrossRef]
- Fujii, R.; Yamada, H.; Munetsuna, E.; Yamazaki, M.; Ando, Y.; Mizuno, G.; Tsuboi, Y.; Ohashi, K.; Ishikawa, H.; Hagiwara, C.; et al. Associations between dietary vitamin intake, ABCA1 gene promoter DNA methylation, and lipid profiles in a Japanese population. Am. J. Clin. Nutr. 2019, 110, 1213–1219. [Google Scholar] [CrossRef] [PubMed]
- Fujii, R.; Yamada, H.; Munetsuna, E.; Yamazaki, M.; Mizuno, G.; Tsuboi, Y.; Ohashi, K.; Ishikawa, H.; Ando, Y.; Hagiwara, C.; et al. Dietary vegetable intake is inversely associated with ATP-binding cassette protein A1 (ABCA1) DNA methylation levels among Japanese women. Nutrition 2019, 65, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Fan, Y.; Li, J.; Wang, J.; Kong, L.; Wang, L.; Li, Z.; Ma, M.; Shi, X.; Liu, S.; et al. The Associations of Plasma Carotenoids and Vitamins with Risk of Age-Related Macular Degeneration: Results from a Matched Case-Control Study in China and Meta-Analysis. Front. Nutr. 2022, 9, 745390. [Google Scholar] [CrossRef] [PubMed]
- Bird, A.C.; Bressler, N.M.; Bressler, S.B.; Chisholm, I.H.; Coscas, G.; Davis, M.D.; de Jong, P.T.; Klaver, C.C.; Klein, B.E.; Klein, R.; et al. An international classification and grading system for age-related maculopathy and age-related macular degeneration. The International ARM Epidemiological Study Group. Surv. Ophthalmol. 1995, 39, 367–374. [Google Scholar] [CrossRef]
- Li, L.C.; Dahiya, R. MethPrimer: Designing primers for methylation PCRs. Bioinformatics 2002, 18, 1427–1431. [Google Scholar] [CrossRef]
- Klein, R.; Lee, K.E.; Tsai, M.Y.; Cruickshanks, K.J.; Gangnon, R.E.; Klein, B. Oxidized Low-density Lipoprotein and the Incidence of Age-related Macular Degeneration. Ophthalmology 2019, 126, 752–758. [Google Scholar] [CrossRef]
- Lin, J.B.; Sene, A.; Santeford, A.; Fujiwara, H.; Sidhu, R.; Ligon, M.M.; Shankar, V.A.; Ban, N.; Mysorekar, I.U.; Ory, D.S.; et al. Oxysterol Signatures Distinguish Age-Related Macular Degeneration from Physiologic Aging. Ebiomedicine 2018, 32, 9–20. [Google Scholar] [CrossRef]
- Sreekumar, P.G.; Su, F.; Spee, C.; Araujo, E.; Nusinowitz, S.; Reddy, S.T.; Kannan, R. Oxidative Stress and Lipid Accumulation Augments Cell Death in LDLR-Deficient RPE Cells and Ldlr−/− Mice. Cells 2022, 12, 43. [Google Scholar] [CrossRef] [PubMed]
- Hovingh, G.K.; Hutten, B.A.; Holleboom, A.G.; Petersen, W.; Rol, P.; Stalenhoef, A.; Zwinderman, A.H.; de Groot, E.; Kastelein, J.J.; Kuivenhoven, J.A. Compromised LCAT function is associated with increased atherosclerosis. Circulation 2005, 112, 879–884. [Google Scholar] [CrossRef] [PubMed]
- Duivenvoorden, R.; Holleboom, A.G.; van den Bogaard, B.; Nederveen, A.J.; de Groot, E.; Hutten, B.A.; Schimmel, A.W.; Hovingh, G.K.; Kastelein, J.J.; Kuivenhoven, J.A.; et al. Carriers of lecithin cholesterol acyltransferase gene mutations have accelerated atherogenesis as assessed by carotid 3.0-T magnetic resonance imaging. J. Am. Coll. Cardiol. 2011, 58, 2481–2487. [Google Scholar] [CrossRef]
- Sene, A.; Apte, R.S. Eyeballing cholesterol efflux and macrophage function in disease pathogenesis. Trends Endocrinol. Metab. 2014, 25, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Pikuleva, I.A.; Curcio, C.A. Cholesterol in the retina: The best is yet to come. Prog. Retin. Eye Res. 2014, 41, 64–89. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Zhang, S.; Fang, L.; Huang, J.; Wang, Q.; Wang, C.; Chen, M. In utero exposure to phenanthrene induces hepatic steatosis in F1 adult female mice. Chemosphere 2020, 258, 127360. [Google Scholar] [CrossRef]
- Long, J.; Chen, P.; Lin, J.; Bai, Y.; Yang, X.; Bian, J.; Lin, Y.; Wang, D.; Yang, X.; Zheng, Y.; et al. DNA methylation-driven genes for constructing diagnostic, prognostic, and recurrence models for hepatocellular carcinoma. Theranostics 2019, 9, 7251–7267. [Google Scholar] [CrossRef]
- Tsai, Y.T.; Li, Y.; Ryu, J.; Su, P.Y.; Cheng, C.H.; Wu, W.H.; Li, Y.S.; Quinn, P.; Leong, K.W.; Tsang, S.H. Impaired cholesterol efflux in retinal pigment epithelium of individuals with juvenile macular degeneration. Am. J. Hum. Genet. 2021, 108, 903–918. [Google Scholar] [CrossRef]
- Storti, F.; Klee, K.; Todorova, V.; Steiner, R.; Othman, A.; van der Velde-Visser, S.; Samardzija, M.; Meneau, I.; Barben, M.; Karademir, D.; et al. Impaired ABCA1/ABCG1-mediated lipid efflux in the mouse retinal pigment epithelium (RPE) leads to retinal degeneration. eLife 2019, 8, e45100. [Google Scholar] [CrossRef]
- Duka, A.; Fotakis, P.; Georgiadou, D.; Kateifides, A.; Tzavlaki, K.; von Eckardstein, L.; Stratikos, E.; Kardassis, D.; Zannis, V.I. ApoA-IV promotes the biogenesis of apoA-IV-containing HDL particles with the participation of ABCA1 and LCAT. J. Lipid Res. 2013, 54, 107–115. [Google Scholar] [CrossRef]
- Czarnecka, H.; Yokoyama, S. Regulation of cellular cholesterol efflux by lecithin:cholesterol acyltransferase reaction through nonspecific lipid exchange. J. Biol. Chem. 1996, 271, 2023–2028. [Google Scholar] [CrossRef]
- Mertens, A.; Verhamme, P.; Bielicki, J.K.; Phillips, M.C.; Quarck, R.; Verreth, W.; Stengel, D.; Ninio, E.; Navab, M.; Mackness, B.; et al. Increased low-density lipoprotein oxidation and impaired high-density lipoprotein antioxidant defense are associated with increased macrophage homing and atherosclerosis in dyslipidemic obese mice: LCAT gene transfer decreases atherosclerosis. Circulation 2003, 107, 1640–1646. [Google Scholar] [CrossRef] [PubMed]
- Vohl, M.C.; Neville, T.A.; Kumarathasan, R.; Braschi, S.; Sparks, D.L. A novel lecithin-cholesterol acyltransferase antioxidant activity prevents the formation of oxidized lipids during lipoprotein oxidation. Biochemistry 1999, 38, 5976–5981. [Google Scholar] [CrossRef] [PubMed]
- Hoppe, K.L.; Francone, O.L. Binding and functional effects of transcription factors Sp1 and Sp3 on the proximal human lecithin:cholesterol acyltransferase promoter. J. Lipid Res. 1998, 39, 969–977. [Google Scholar] [CrossRef] [PubMed]
- Kriaa, A.; Jablaoui, A.; Rhimi, S.; Soussou, S.; Mkaouar, H.; Mariaule, V.; Gruba, N.; Gargouri, A.; Maguin, E.; Lesner, A.; et al. SP-1, a Serine Protease from the Gut Microbiota, Influences Colitis and Drives Intestinal Dysbiosis in Mice. Cells 2021, 10, 2658. [Google Scholar] [CrossRef]
- Song, L.; Wang, X.; Qu, X.; Lv, C. Transcription Factor Specificity Protein 1 Regulates Inflammation and Fibrin Deposition in Nasal Polyps via the Regulation of microRNA-125b and the Wnt/β-catenin Signaling Pathway. Inflammation 2022, 45, 1118–1132. [Google Scholar] [CrossRef]
- Vallée, A. Curcumin and Wnt/β-catenin signaling in exudative age-related macular degeneration (Review). Int. J. Mol. Med. 2022, 49, 79. [Google Scholar] [CrossRef]
- McEneny, J.; Henry, S.L.; Woodside, J.; Moir, S.; Rudd, A.; Vaughan, N.; Thies, F. Lycopene-rich diets modulate HDL functionality and associated inflammatory markers without affecting lipoprotein size and distribution in moderately overweight, disease-free, middle-aged adults: A randomized controlled trial. Front. Nutr. 2022, 9, 954593. [Google Scholar] [CrossRef]
- Daniels, J.A.; Mulligan, C.; McCance, D.; Woodside, J.V.; Patterson, C.; Young, I.S.; McEneny, J. A randomised controlled trial of increasing fruit and vegetable intake and how this influences the carotenoid concentration and activities of PON-1 and LCAT in HDL from subjects with type 2 diabetes. Cardiovasc. Diabetol. 2014, 13, 16. [Google Scholar] [CrossRef]
- Matsumoto, A.; Mizukami, H.; Mizuno, S.; Umegaki, K.; Nishikawa, J.; Shudo, K.; Kagechika, H.; Inoue, M. beta-Cryptoxanthin, a novel natural RAR ligand, induces ATP-binding cassette transporters in macrophages. Biochem. Pharmacol. 2007, 74, 256–264. [Google Scholar] [CrossRef]
- Lehmann, J.M.; Jong, L.; Fanjul, A.; Cameron, J.F.; Lu, X.P.; Haefner, P.; Dawson, M.I.; Pfahl, M. Retinoids selective for retinoid X receptor response pathways. Science 1992, 258, 1944–1946. [Google Scholar] [CrossRef]
- Duester, G. Towards a Better Vision of Retinoic Acid Signaling during Eye Development. Cells 2022, 11, 322. [Google Scholar] [CrossRef]
- Janssen, J.J.; Kuhlmann, E.D.; van Vugt, A.H.; Winkens, H.J.; Janssen, B.P.; Deutman, A.F.; Driessen, C.A. Retinoic acid receptors and retinoid X receptors in the mature retina: Subtype determination and cellular distribution. Curr. Eye Res. 1999, 19, 338–347. [Google Scholar] [CrossRef]
- Subramanyam, D.; Belair, C.D.; Barry-Holson, K.Q.; Lin, H.; Kogan, S.C.; Passegué, E.; Blelloch, R. PML-RARα and Dnmt3a1 cooperate in vivo to promote acute promyelocytic leukemia. Cancer Res. 2010, 70, 8792–8801. [Google Scholar] [CrossRef] [PubMed]
- O’Hagan, H.M.; Wang, W.; Sen, S.; Destefano, S.C.; Lee, S.S.; Zhang, Y.W.; Clements, E.G.; Cai, Y.; Van Neste, L.; Easwaran, H.; et al. Oxidative damage targets complexes containing DNA methyltransferases, SIRT1, and polycomb members to promoter CpG Islands. Cancer Cell 2011, 20, 606–619. [Google Scholar] [CrossRef] [PubMed]
- Vaccaro, J.A.; Qasem, A.; Naser, S.A. Folate and Vitamin B(12) Deficiency Exacerbate Inflammation during Mycobacterium avium paratuberculosis (MAP) Infection. Nutrients 2023, 15, 261. [Google Scholar] [CrossRef]
- Kumar, A.; Verma, A.; Chaurasia, R.N. Vitamin D and inflammatory cytokines association in mild cognitive impaired subjects. Neurosci. Lett. 2023, 795, 137044. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.C.; Voors, A.A.; van Veldhuisen, D.J.; van der Veer, E.; Belonje, A.M.; Szymanski, M.K.; Silljé, H.H.; van Gilst, W.H.; Jaarsma, T.; de Boer, R.A. Vitamin D status and outcomes in heart failure patients. Eur. J. Heart Fail. 2011, 13, 619–625. [Google Scholar] [CrossRef]
- Heloterä, H.; Kaarniranta, K. A Linkage between Angiogenesis and Inflammation in Neovascular Age-Related Macular Degeneration. Cells 2022, 11, 3453. [Google Scholar] [CrossRef]
- Ding, N.; Maiuri, A.R.; O’Hagan, H.M. The emerging role of epigenetic modifiers in repair of DNA damage associated with chronic inflammatory diseases. Mutat. Res. Mol. Mech. Mutagen. 2019, 780, 69–81. [Google Scholar] [CrossRef]
- Simonelli, S.; Ossoli, A.; Banfi, C.; Pavanello, C.; Calabresi, L.; Gianazza, E. A proteomic approach to identify novel disease biomarkers in LCAT deficiency. J. Proteom. 2019, 198, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Koukos, G.; Chroni, A.; Duka, A.; Kardassis, D.; Zannis, V.I. Naturally occurring and bioengineered apoA-I mutations that inhibit the conversion of discoidal to spherical HDL: The abnormal HDL phenotypes can be corrected by treatment with LCAT. Biochem. J. 2007, 406, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, A.W.; Januar, V.; Sexton-Oates, A.; Joo, J.E.; Franchina, M.; Wang, J.J.; Liang, H.; Craig, J.E.; Saffery, R. DNA methylation landscape of ocular tissue relative to matched peripheral blood. Sci. Rep. 2017, 7, 46330. [Google Scholar] [CrossRef]
- Oliver, V.F.; Jaffe, A.E.; Song, J.; Wang, G.; Zhang, P.; Branham, K.E.; Swaroop, A.; Eberhart, C.G.; Zack, D.J.; Qian, J.; et al. Differential DNA methylation identified in the blood and retina of AMD patients. Epigenetics 2015, 10, 698–707. [Google Scholar] [CrossRef]
- Saptarshi, N.; Green, D.; Cree, A.; Lotery, A.; Paraoan, L.; Porter, L.F. Epigenetic Age Acceleration Is Not Associated with Age-Related Macular Degeneration. Int. J. Mol. Sci. 2021, 22, 13457. [Google Scholar] [CrossRef] [PubMed]
- Moore, L.D.; Le, T.; Fan, G. DNA methylation and its basic function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Liu, Y.; Zhao, S.; Zheng, Z.; Shen, C.; An, L.; Yuan, Y. Large-scale analysis reveals a novel risk score to predict overall survival in hepatocellular carcinoma. Cancer Manag. Res. 2018, 10, 6079–6096. [Google Scholar] [CrossRef]
- Ouyang, G.; Yi, B.; Pan, G.; Chen, X. A robust twelve-gene signature for prognosis prediction of hepatocellular carcinoma. Cancer Cell Int. 2020, 20, 207. [Google Scholar] [CrossRef]
- Newman, A.M.; Gallo, N.B.; Hancox, L.S.; Miller, N.J.; Radeke, C.M.; Maloney, M.A.; Cooper, J.B.; Hageman, G.S.; Anderson, D.H.; Johnson, L.V.; et al. Systems-level analysis of age-related macular degeneration reveals global biomarkers and phenotype-specific functional networks. Genome Med. 2012, 4, 16. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Controls (N = 174) | AMD Patients (N = 126) | p |
|---|---|---|---|
| Age, years | 65.45 ± 7.60 | 66.99 ± 7.37 | 0.08 |
| Male, % | 37.93 | 40.47 | 0.66 |
| Cigarette smoking, % | 10.91 | 29.37 | <0.01 |
| Alcohol consumption, % | 7.47 | 15.08 | 0.04 |
| Education Level, % | <0.01 | ||
| Less than college | 36.21 | 61.90 | |
| College | 63.79 | 38.10 | |
| Physical activity, % | <0.01 | ||
| >7 h/w | 31.03 | 13.50 | |
| 1–7 h/w | 50.57 | 35.71 | |
| Rarely/never | 18.39 | 50.79 | |
| Multivitamin use, % | 21.26 | 27.78 | 0.19 |
| BMI, kg/m2 | 23.42 ± 1.43 | 23.69 ± 1.29 | 0.10 |
| History of diabetes, % | 10.92 | 21.43 | 0.01 |
| History of coronary heart disease, % | 6.90 | 15.87 | 0.01 |
| Family history of diabetes, % | 20.11 | 19.05 | 0.82 |
| Family history of coronary heart disease, % | 27.59 | 20.63 | 0.17 |
| DNA Methylation | Odds Ratios (95% CIs) by Tertiles of LCAT DNA Methylation | LCAT Increase, per 1 SD | Ptrend | ||
|---|---|---|---|---|---|
| T1 | T2 | T3 | |||
| DNA methylation level, median | 1.48 | 1.79 | 1.99 | ||
| No. of cases/controls | 26/58 | 24/58 | 76/58 | ||
| Model 1 | 1 | 0.93 (0.48, 1.81) | 2.88 (1.61, 5.13) | 1.75 (1.34, 2.29) | <0.01 |
| Model 2 | 1 | 0.87 (0.39, 1.96) | 5.09 (2.46, 10.53) | 2.18 (1.56, 3.06) | <0.01 |
| Model 3 | 1 | 0.91 (0.40, 2.05) | 5.37 (2.56, 11.28) | 2.23 (1.58, 3.13) | <0.01 |
| Characteristic | Model 1 | Model 2 | Model 3 | |||
|---|---|---|---|---|---|---|
| OR (95% CI) | p | OR (95% CI) | p | OR (95% CI) | p | |
| LCAT DNA methylation increase per 1 SD | ||||||
| <1.85 | 1.14 (0.83, 1.59) | 0.42 | 1.05 (0.74, 1.52) | 0.80 | 1.13 (0.78, 1.67) | 0.54 |
| ≥1.85 | 1.19 (0.81, 1.79) | 0.39 | 1.47 (1.05, 2.12) | 0.03 | 1.68 (1.21, 2.41) | <0.01 |
| Vitamins and Carotenoids | β | 95% CI | SE | p |
|---|---|---|---|---|
| Retinol (µmol/L) | −0.05 | (−0.08, −0.01) | 0.02 | <0.01 |
| α-tocopherol (µmol/L) | −0.24 | (−0.49, 0.01) | 0.13 | 0.06 |
| Lutein (µmol/L) | 0.04 | (−0.09, 0.17) | 0.07 | 0.53 |
| β-cryptoxanthin (µmol/L) | −0.25 | (−0.42, −0.08) | 0.09 | <0.01 |
| β-carotene (µmol/L) | −0.25 | (−0.78, 0.26) | 0.27 | 0.35 |
| Lycopene (µmol/L) | 0.13 | (−0.05, 0.32) | 0.09 | 0.16 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Li, Y.; Hou, Y.; Fan, Y.; Jiang, H.; Li, B.; Zhu, H.; Liu, Y.; Zhang, L.; Zhang, J.; et al. Association of Plasma Vitamins and Carotenoids, DNA Methylation of LCAT, and Risk of Age-Related Macular Degeneration. Nutrients 2023, 15, 2985. https://doi.org/10.3390/nu15132985
Li Z, Li Y, Hou Y, Fan Y, Jiang H, Li B, Zhu H, Liu Y, Zhang L, Zhang J, et al. Association of Plasma Vitamins and Carotenoids, DNA Methylation of LCAT, and Risk of Age-Related Macular Degeneration. Nutrients. 2023; 15(13):2985. https://doi.org/10.3390/nu15132985
Chicago/Turabian StyleLi, Zhaofang, Yajing Li, Yijing Hou, Yahui Fan, Hong Jiang, Baoyu Li, Hailu Zhu, Yaning Liu, Lei Zhang, Jie Zhang, and et al. 2023. "Association of Plasma Vitamins and Carotenoids, DNA Methylation of LCAT, and Risk of Age-Related Macular Degeneration" Nutrients 15, no. 13: 2985. https://doi.org/10.3390/nu15132985
APA StyleLi, Z., Li, Y., Hou, Y., Fan, Y., Jiang, H., Li, B., Zhu, H., Liu, Y., Zhang, L., Zhang, J., Wu, M., Ma, T., Zhao, T., & Ma, L. (2023). Association of Plasma Vitamins and Carotenoids, DNA Methylation of LCAT, and Risk of Age-Related Macular Degeneration. Nutrients, 15(13), 2985. https://doi.org/10.3390/nu15132985