
Featured Prebiotic Agent: The Roles and Mechanisms of Direct and Indirect Prebiotic Activities of Lactoferrin and Its Application in Disease Control



Abstract
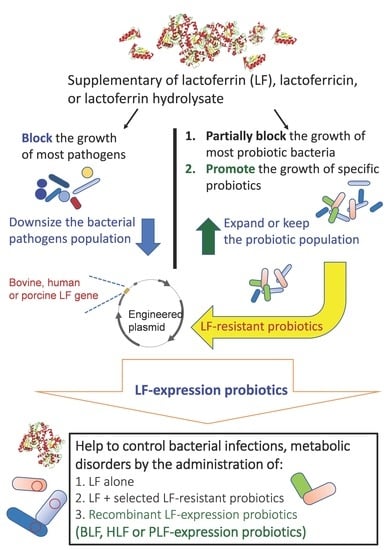
1. Introduction
2. Contrasting Antibacterial Effects of Lactoferrins on Pathogens and Probiotics
3. The Potential Prebiotic Activity of Lactoferrin In Vitro
4. The Direct and Indirect Prebiotic Roles of Lactoferrin In Vivo: Benefits of LF Supplementation on Gut Microbiota or Disease Control
5. The Mechanism of LF Becoming a Prebiotic Agent
6. Combining LF with Probiotics to Fight Bacterial Infections or Provide Beneficial Characteristics in Disease Control
7. Safety Issue
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sorensen, M.; Sorensen, S. The proteins in whey. Compte rendu des Travaux du Laboratoire de Carlsberg. Ser. Chim. 1940, 23, 55–99. [Google Scholar]
- Groves, M.L. The isolation of a red protein from Milk2. J. Am. Chem. Soc. 1960, 82, 3345–3350. [Google Scholar] [CrossRef]
- Levay, P.F.; Viljoen, M. Lactoferrin: A general review. Haematologica 1995, 80, 252–267. [Google Scholar]
- Iyer, S.; Lonnerdal, B. Lactoferrin, lactoferrin receptors and iron metabolism. Eur. J. Clin. Nutr. 1993, 47, 232–241. [Google Scholar] [PubMed]
- Gruden, Š.; Poklar Ulrih, N. Diverse mechanisms of antimicrobial activities of lactoferrins, lactoferricins, and other lactoferrin-derived peptides. Int. J. Mol. Sci. 2021, 22, 11264. [Google Scholar] [CrossRef] [PubMed]
- Frioni, A.; Conte, M.P.; Cutone, A.; Longhi, C.; Musci, G.; di Patti, M.C.; Natalizi, T.; Marazzato, M.; Lepanto, M.S.; Puddu, P.; et al. Lactoferrin differently modulates the inflammatory response in epithelial models mimicking human inflammatory and infectious diseases. Biometals 2014, 27, 843–856. [Google Scholar] [CrossRef] [PubMed]
- Legrand, D. Overview of lactoferrin as a natural immune modulator. J. Pediatr. 2016, 173, S10–S15. [Google Scholar] [CrossRef]
- Valenti, P.; Antonini, G. Lactoferrin: An important host defence against microbial and viral attack. Cell. Mol. Life Sci. 2005, 62, 2576–2587. [Google Scholar] [CrossRef]
- Sabra, S.; Agwa, M.M. Lactoferrin, a unique molecule with diverse therapeutical and nanotechnological applications. Int. J. Biol. Macromol. 2020, 164, 1046–1060. [Google Scholar] [CrossRef]
- Superti, F. Lactoferrin from bovine milk: A protective companion for life. Nutrients 2020, 12, 2562. [Google Scholar] [CrossRef]
- Sinopoli, A.; Isonne, C.; Santoro, M.M.; Baccolini, V. The effects of orally administered lactoferrin in the prevention and management of viral infections: A systematic review. Rev. Med. Virol. 2022, 32, e2261. [Google Scholar] [CrossRef] [PubMed]
- Sienkiewicz, M.; Jaśkiewicz, A.; Tarasiuk, A.; Fichna, J. Lactoferrin: An overview of its main functions, immunomodulatory and antimicrobial role, and clinical significance. Crit. Rev. Food Sci. Nutr. 2022, 62, 6016–6033. [Google Scholar] [CrossRef] [PubMed]
- Embleton, N.D.; Berrington, J.E.; McGuire, W.; Stewart, C.J.; Cummings, S.P. Lactoferrin: Antimicrobial activity and therapeutic potential. Semin. Fetal Neonatal Med. 2013, 18, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Bruni, N.; Capucchio, M.T.; Biasibetti, E.; Pessione, E.; Cirrincione, S.; Giraudo, L.; Corona, A.; Dosio, F. Antimicrobial activity of lactoferrin-related peptides and applications in human and veterinary medicine. Molecules 2016, 21, 752. [Google Scholar] [CrossRef]
- Dierick, M.; Vanrompay, D.; Devriendt, B.; Cox, E. Lactoferrin, a versatile natural antimicrobial glycoprotein that modulates the host’s innate immunity. Biochem. Cell Biol. 2021, 99, 61–65. [Google Scholar] [CrossRef]
- Brock, J. Lactoferrin in human milk: Its role in iron absorption and protection against enteric infection in the newborn infant. Arch. Dis. Child. 1980, 55, 417. [Google Scholar] [CrossRef]
- Bullen, J.; Rogers, H.J.; Leigh, L. Iron-binding proteins in milk and resistance to Escherichia coli infection in infants. Br. Med. J. 1972, 1, 69–75. [Google Scholar] [CrossRef]
- Tomita, M.; Bellamy, W.; Takase, M.; Yamauchi, K.; Wakabayashi, H.; Kawase, K. Potent antibacterial peptides generated by pepsin digestion of bovine lactoferrin. J. Dairy Sci. 1991, 74, 4137–4142. [Google Scholar] [CrossRef]
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef]
- Davani-Davari, D.; Negahdaripour, M.; Karimzadeh, I.; Seifan, M.; Mohkam, M.; Masoumi, S.J.; Berenjian, A.; Ghasemi, Y. Prebiotics: Definition, Types, Sources, Mechanisms, and Clinical Applications. Foods 2019, 8, 92. [Google Scholar] [CrossRef] [PubMed]
- Neri-Numa, I.A.; Pastore, G.M. Novel insights into prebiotic properties on human health: A review. Food Res. Int. 2020, 131, 108973. [Google Scholar] [CrossRef] [PubMed]
- Peng, M.; Tabashsum, Z.; Anderson, M.; Truong, A.; Houser, A.K.; Padilla, J.; Akmel, A.; Bhatti, J.; Rahaman, S.O.; Biswas, D. Effectiveness of probiotics, prebiotics, and prebiotic-like components in common functional foods. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1908–1933. [Google Scholar] [CrossRef] [PubMed]
- Fuller, R. Probiotics in man and animals. J. Appl. Bacteriol. 1989, 66, 365–378. [Google Scholar]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Wang, G.; Chen, Y.; Xia, Y.; Song, X.; Ai, L. Characteristics of probiotic preparations and their applications. Foods 2022, 11, 2472. [Google Scholar] [CrossRef]
- Bhogoju, S.; Nahashon, S. Recent advances in probiotic application in animal health and nutrition: A review. Agriculture 2022, 12, 304. [Google Scholar] [CrossRef]
- Yan, F.; Polk, D.B. Probiotics and probiotic-derived functional factors—Mechanistic insights into applications for intestinal homeostasis. Front. Immunol. 2020, 11, 1428. [Google Scholar] [CrossRef]
- Rahman, M.M.; Kim, W.-S.; Ito, T.; Kumura, H.; Shimazaki, K.-I. Growth promotion and cell binding ability of bovine lactoferrin to Bifidobacterium longum. Anaerobe 2009, 15, 133–137. [Google Scholar] [CrossRef]
- Kim, W.S.; Ohashi, M.; Tanaka, T.; Kumura, H.; Kim, G.Y.; Kwon, I.K.; Goh, J.S.; Shimazaki, K. Growth-promoting effects of lactoferrin on L. acidophilus and Bifidobacterium spp. Biometals 2004, 17, 279–283. [Google Scholar] [CrossRef]
- Petschow, B.W.; Talbott, R.D.; Batema, R.P. Ability of lactoferrin to promote the growth of Bifidobacterium spp. in vitro is independent of receptor binding capacity and iron saturation level. J. Med. Microbiol. 1999, 48, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.-W.; Jheng, T.; Shyu, C.-L.; Mao, F. Antimicrobial potential for the combination of bovine lactoferrin or its hydrolysate with lactoferrin-resistant probiotics against foodborne pathogens. J. Dairy Sci. 2013, 96, 1438–1446. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.-W.; Liu, Z.-S.; Kuo, T.-C.; Hsieh, M.-C.; Li, Z.-W. Prebiotic effects of bovine lactoferrin on specific probiotic bacteria. Biometals 2017, 30, 237–248. [Google Scholar] [CrossRef]
- Liu, Z.-S.; Lin, C.-F.; Chen, P.-W. Transcriptome analysis of Lactobacillus rhamnosus GG strain treated with prebiotic-bovine lactoferrin under a cold environment. J. Food Drug Anal. 2021, 29, 402. [Google Scholar] [CrossRef] [PubMed]
- Vega-Bautista, A.; de la Garza, M.; Carrero, J.C.; Campos-Rodríguez, R.; Godínez-Victoria, M.; Drago-Serrano, M.E. The impact of Lactoferrin on the growth of intestinal inhabitant bacteria. Int. J. Mol. Sci. 2019, 20, 4707. [Google Scholar] [CrossRef]
- Artym, J.; Zimecki, M. Antimicrobial and prebiotic activity of lactoferrin in the female reproductive tract: A comprehensive review. Biomedicines 2021, 9, 1940. [Google Scholar] [CrossRef] [PubMed]
- Niaz, B.; Saeed, F.; Ahmed, A.; Imran, M.; Maan, A.A.; Khan, M.K.I.; Tufail, T.; Anjum, F.M.; Hussain, S.; Suleria, H.A.R. Lactoferrin (LF): A natural antimicrobial protein. Int. J. Food Prop. 2019, 22, 1626–1641. [Google Scholar] [CrossRef]
- Shini, V.S.; Udayarajan, C.T.; Nisha, P. A comprehensive review on lactoferrin: A natural multifunctional glycoprotein. Food Funct. 2022, 13, 11954–11972. [Google Scholar] [CrossRef]
- Reiter, B.; Perraudin, J.-P. The antibacterial activity of lactoferrin and neonatal E. coli infections: A selective and critical review. Adv. Lact. Res. 1998, 443, 175–188. [Google Scholar]
- Adlerova, L.; Bartoskova, A.; Faldyna, M. Lactoferrin: A review. Vet. Med. 2008, 53, 457–468. [Google Scholar] [CrossRef]
- Gu, Q.; Yin, Y.; Yan, X.; Liu, X.; Liu, F.; McClements, D.J. Encapsulation of multiple probiotics, synbiotics, or nutrabiotics for improved health effects: A review. Adv. Colloid Interface Sci. 2022, 309, 102781. [Google Scholar] [CrossRef]
- García-Montoya, I.A.; Cendón, T.S.; Arévalo-Gallegos, S.; Rascón-Cruz, Q. Lactoferrin a multiple bioactive protein: An overview. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2012, 1820, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Jenssen, H.; Hancock, R.E. Antimicrobial properties of lactoferrin. Biochimie 2009, 91, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Kim, W. Transferrin family proteins bind to Bifidobacterium spp. Milchwissenschaft 2004, 59, 491–494. [Google Scholar]
- Kim, W.-S.; Rahman, M.M.; Kumura, H.; Shimazaki, K.-I. Comparison of growth promoting effects on Bifidobacterium spp. by bovine lactoferrin hydrolysates. Biosci. Microflora 2005, 24, 119–123. [Google Scholar] [CrossRef]
- Chen, P.-W.; Ku, Y.-W.; Chu, F.-Y. Influence of bovine lactoferrin on the growth of selected probiotic bacteria under aerobic conditions. Biometals 2014, 27, 905–914. [Google Scholar] [CrossRef]
- Oda, H.; Wakabayashi, H.; Yamauchi, K.; Abe, F. Lactoferrin and bifidobacteria. Biometals 2014, 27, 915–922. [Google Scholar] [CrossRef]
- Inay, O.M.; da Silva, A.S.; Honjoya, E.R.; Sugimoto, H.H.; de Souza, C.H.B.; de Santana, E.H.W.; de Rezende Costa, M.; Aragon-Alegro, L.C. Action of lactoferrin on the multiplication of Lactobacillus casei in vitro and in Minas fresh cheese. Semin. Ciências Agrárias 2012, 33, 3153–3162. [Google Scholar] [CrossRef]
- Duran, A. The effect of bovine lactoferrin on the microbiological properties of raw milk. Gıda 2021, 46, 681–691. [Google Scholar] [CrossRef]
- Wu, J.; Zang, M.; Wang, S.; Qiao, X.; Zhao, B.; Bai, J.; Zhao, Y.; Shi, Y. Lactoferricin, an antimicrobial motif derived from lactoferrin with food preservation potential. Crit. Rev. Food Sci. Nutr. 2023, 1–13. [Google Scholar] [CrossRef]
- Hassan, M.A.; Abedelmaksoud, T.G.; El-Maksoud, A.; Ahmed, A. Effects of Lactoferrin Supplemented with Fermented Milk on Obesity-Associated Pancreatic Damage in Rats. Life 2022, 12, 2019. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.K.; Chierici, R.; Sawatzki, G.; Hill, M.J.; Volpato, S.; Vigi, V. Supplementation of an adapted formula with bovine lactoferrin: 1. Effect on the infant faecal flora. Acta Paediatr. 1992, 81, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Mastromarino, P.; Capobianco, D.; Campagna, G.; Laforgia, N.; Drimaco, P.; Dileone, A.; Baldassarre, M.E. Correlation between lactoferrin and beneficial microbiota in breast milk and infant’s feces. Biometals 2014, 27, 1077–1086. [Google Scholar] [CrossRef] [PubMed]
- Dix, C.; Wright, O. Bioavailability of a novel form of microencapsulated bovine lactoferrin and its effect on inflammatory markers and the gut microbiome: A pilot study. Nutrients 2018, 10, 1115. [Google Scholar] [CrossRef] [PubMed]
- Sherman, M.P.; Sherman, J.; Arcinue, R.; Niklas, V. Randomized control trial of human recombinant lactoferrin: A substudy reveals effects on the fecal microbiome of very low birth weight infants. J. Pediatr. 2016, 173, S37–S42. [Google Scholar] [CrossRef]
- Haiwen, Z.; Rui, H.; Bingxi, Z.; Qingfeng, G.; Jifeng, Z.; Xuemei, W.; Beibei, W. Oral administration of bovine lactoferrin-derived lactoferricin (Lfcin) B could attenuate enterohemorrhagic Escherichia coli O157: H7 induced intestinal disease through improving intestinal barrier function and microbiota. J. Agric. Food Chem. 2019, 67, 3932–3945. [Google Scholar] [CrossRef]
- Zhou, H.-H.; Wang, G.; Luo, L.; Ding, W.; Xu, J.-Y.; Yu, Z.; Qin, L.-Q.; Wan, Z. Dietary lactoferrin has differential effects on gut microbiota in young versus middle-aged APPswe/PS1dE9 transgenic mice but no effects on cognitive function. Food Nutr. Res. 2021, 65, 5496. [Google Scholar] [CrossRef]
- Bellés, A.; Aguirre-Ramírez, D.; Abad, I.; Parras-Moltó, M.; Sánchez, L.; Grasa, L. Lactoferrin modulates gut microbiota and Toll-like receptors (TLRs) in mice with dysbiosis induced by antibiotics. Food Funct. 2022, 13, 5854–5869. [Google Scholar] [CrossRef]
- Sun, J.; Ren, F.; Xiong, L.; Zhao, L.; Guo, H. Bovine lactoferrin suppresses high-fat diet induced obesity and modulates gut microbiota in C57BL/6J mice. J. Funct. Foods 2016, 22, 189–200. [Google Scholar] [CrossRef]
- Pammi, M.; Abrams, S.A. Oral lactoferrin for the prevention of sepsis and necrotizing enterocolitis in preterm infants. Cochrane Database Syst. Rev. 2015, CD007137. [Google Scholar] [CrossRef]
- Ganji, N.; Li, B.; Lee, C.; Pierro, A. Necrotizing enterocolitis: Recent advances in treatment with translational potential. Pediatr. Surg. Int. 2023, 39, 205. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhu, H.; Li, B.; Robinson, S.C.; Lee, C.; O’Connell, J.S.; Bindi, E.; Zheng, S.; Sherman, P.M.; Pierro, A. Lactoferrin reduces necrotizing enterocolitis severity by upregulating intestinal epithelial proliferation. Eur. J. Pediatr. Surg. 2020, 30, 090–095. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Francis, J.; Doster, R.S.; Haley, K.P.; Craft, K.M.; Moore, R.E.; Chambers, S.A.; Aronoff, D.M.; Osteen, K.; Damo, S.M. Lactoferrin: A critical mediator of both host immune response and antimicrobial activity in response to streptococcal infections. ACS Infect. Dis. 2020, 6, 1615–1623. [Google Scholar] [CrossRef]
- Allison, L.M.; Walker, L.; Sanders, B.J.; Yang, Z.; Eckert, G.; Gregory, R.L. Effect of human milk and its components on Streptococcus mutans biofilm formation. J. Clin. Pediatr. Dent. 2015, 39, 255–261. [Google Scholar] [CrossRef]
- Avery, T.M.; Boone, R.L.; Lu, J.; Spicer, S.K.; Guevara, M.A.; Moore, R.E.; Chambers, S.A.; Manning, S.D.; Dent, L.; Marshall, D.; et al. Analysis of Antimicrobial and Antibiofilm Activity of Human Milk Lactoferrin Compared to Bovine Lactoferrin against Multidrug Resistant and Susceptible Acinetobacter baumannii Clinical Isolates. ACS Infect. Dis. 2021, 7, 2116–2126. [Google Scholar] [CrossRef]
- Talbert, J.A.; Lu, J.; Spicer, S.K.; Moore, R.E.; Townsend, S.D.; Gaddy, J.A. Ameliorating adverse perinatal outcomes with Lactoferrin: An intriguing chemotherapeutic intervention. Bioorg. Med. Chem. 2022, 74, 117037. [Google Scholar] [CrossRef]
- Otsuki, K.; Imai, N. Effects of lactoferrin in 6 patients with refractory bacterial vaginosis. Biochem. Cell Biol. 2017, 95, 31–33. [Google Scholar] [CrossRef]
- Mayeur, S.; Spahis, S.; Pouliot, Y.; Levy, E. Lactoferrin, a Pleiotropic Protein in Health and Disease. Antioxid. Redox Signal. 2016, 24, 813–836. [Google Scholar] [CrossRef]
- Brimelow, R.E.; West, N.P.; Williams, L.T.; Cripps, A.W.; Cox, A.J. A role for whey-derived lactoferrin and immunoglobulins in the attenuation of obesity-related inflammation and disease. Crit. Rev. Food. Sci. Nutr. 2017, 57, 1593–1602. [Google Scholar] [CrossRef]
- Artym, J. [A remedy against obesity? The role of lactoferrin in the metabolism of glucose and lipids]. Postepy Hig. Med. Dosw. 2012, 66, 937–953. [Google Scholar] [CrossRef]
- Li, Y.-C.; Hsieh, C.-C. Lactoferrin dampens high-fructose corn syrup-induced hepatic manifestations of the metabolic syndrome in a murine model. PLoS ONE 2014, 9, e97341. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.-X.; He, Q.; Zhou, Y.; Xu, J.-Y.; Zhang, Z.; Chen, C.-L.; Wu, Y.-H.; Chen, Y.; Qin, L.-Q.; Li, Y.-H. Protective effect and mechanism of lactoferrin combined with hypoxia against high-fat diet induced obesity and non-alcoholic fatty liver disease in mice. Int. J. Biol. Macromol. 2023, 227, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Jamka, M.; Kaczmarek, N.; Mądry, E.; Krzyżanowska-Jankowska, P.; Bajerska, J.; Kręgielska-Narożna, M.; Bogdański, P.; Walkowiak, J. Metabolic Health in Obese Subjects—Is There a Link to Lactoferrin and Lactoferrin Receptor-Related Gene Polymorphisms? Nutrients 2020, 12, 2843. [Google Scholar] [CrossRef] [PubMed]
- Sato, B.; Kanai, S.; Sakaguchi, D.; Yajima, K.; Matsumoto, Y.; Morohoshi, K.; Kagaya, S.; Izumo, N.; Ichinose, M.; Kang, W. Suppressive Role of Lactoferrin in Overweight-Related Female Fertility Problems. Nutrients 2022, 14, 938. [Google Scholar] [CrossRef] [PubMed]
- Pilvi, T.K.; Harala, S.; Korpela, R.; Mervaala, E.M. Effects of high-calcium diets with different whey proteins on weight loss and weight regain in high-fat-fed C57BL/6J mice. Br. J. Nutr. 2009, 102, 337–341. [Google Scholar] [CrossRef]
- Shi, J.; Finckenberg, P.; Martonen, E.; Ahlroos-Lehmus, A.; Pilvi, T.K.; Korpela, R.; Mervaala, E.M. Metabolic effects of lactoferrin during energy restriction and weight regain in diet-induced obese mice. J. Funct. Foods 2012, 4, 66–78. [Google Scholar] [CrossRef]
- Takeuchi, T.; Shimizu, H.; Ando, K.; Harada, E. Bovine lactoferrin reduces plasma triacylglycerol and NEFA accompanied by decreased hepatic cholesterol and triacylglycerol contents in rodents. Br. J. Nutr. 2004, 91, 533–538. [Google Scholar] [CrossRef]
- McManus, B.; Korpela, R.; O’Connor, P.; Schellekens, H.; Cryan, J.F.; Cotter, P.D.; Nilaweera, K.N. Compared to casein, bovine lactoferrin reduces plasma leptin and corticosterone and affects hypothalamic gene expression without altering weight gain or fat mass in high fat diet fed C57/BL6J mice. Nutr. Metab. 2015, 12, 53. [Google Scholar] [CrossRef]
- Gomaa, E.Z. Human gut microbiota/microbiome in health and diseases: A review. Antonie Leeuwenhoek 2020, 113, 2019–2040. [Google Scholar] [CrossRef]
- Gagliardi, A.; Totino, V.; Cacciotti, F.; Iebba, V.; Neroni, B.; Bonfiglio, G.; Trancassini, M.; Passariello, C.; Pantanella, F.; Schippa, S. Rebuilding the Gut Microbiota Ecosystem. Int. J. Environ. Res. Public Health 2018, 15, 1679. [Google Scholar] [CrossRef]
- Cohen, M.S.; Britigan, B.E.; French, M.; Bean, K. Preliminary observations on lactoferrin secretion in human vaginal mucus: Variation during the menstrual cycle, evidence of hormonal regulation, and implications for infection with Neisseria gonorrhoeae. Am. J. Obstet. Gynecol. 1987, 157, 1122–1125. [Google Scholar] [CrossRef] [PubMed]
- Novak, R.M.; Donoval, B.A.; Graham, P.J.; Boksa, L.A.; Spear, G.; Hershow, R.C.; Chen, H.Y.; Landay, A. Cervicovaginal levels of lactoferrin, secretory leukocyte protease inhibitor, and RANTES and the effects of coexisting vaginoses in human immunodeficiency virus (HIV)-seronegative women with a high risk of heterosexual acquisition of HIV infection. Clin. Vaccine Immunol. 2007, 14, 1102–1107. [Google Scholar] [CrossRef] [PubMed]
- Asakuma, S.; Hatakeyama, E.; Urashima, T.; Yoshida, E.; Katayama, T.; Yamamoto, K.; Kumagai, H.; Ashida, H.; Hirose, J.; Kitaoka, M. Physiology of consumption of human milk oligosaccharides by infant gut-associated bifidobacteria. J. Biol. Chem. 2011, 286, 34583–34592. [Google Scholar] [CrossRef]
- Rahman, M.M.; Kim, W.-S.; Kumura, H.; Shimazaki, K.-I. In vitro effects of bovine lactoferrin on autoaggregation ability and surface hydrophobicity of bifidobacteria. Anaerobe 2008, 14, 73–77. [Google Scholar] [CrossRef]
- Garrido, D.; Nwosu, C.; Ruiz-Moyano, S.; Aldredge, D.; German, J.B.; Lebrilla, C.B.; Mills, D.A. Endo-β-N-acetylglucosaminidases from infant gut-associated bifidobacteria release complex N-glycans from human milk glycoproteins. Mol. Cell. Proteom. 2012, 11, 775–785. [Google Scholar] [CrossRef] [PubMed]
- Karav, S.; Le Parc, A.; Leite Nobrega de Moura Bell, J.M.; Frese, S.A.; Kirmiz, N.; Block, D.E.; Barile, D.; Mills, D.A. Oligosaccharides released from milk glycoproteins are selective growth substrates for infant-associated bifidobacteria. Appl. Environ. Microbiol. 2016, 82, 3622–3630. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Kim, W.-S.; Ito, T.; Kumura, H.; Shimazaki, K. Examination of bovine lactoferrin binding to bifidobacteria. Appl. Biochem. Microbiol. 2008, 44, 478–481. [Google Scholar] [CrossRef]
- Kim, W.-S.; Tanaka, T.; Kumura, H.; Shimazaki, K.-I. Lactoferrin-binding proteins in Bifidobacterium bifidum. Biochem. Cell Biol. 2002, 80, 91–94. [Google Scholar] [CrossRef]
- Schryvers, A.B.; Morris, L.J. Identification and characterization of the human lactoferrin-binding protein from Neisseria meningitidis. Infect. Immun. 1988, 56, 1144–1149. [Google Scholar] [CrossRef]
- Hammerschmidt, S.; Bethe, G.; Remane, H.P.; Chhatwal, G.S. Identification of pneumococcal surface protein A as a lactoferrin-binding protein of Streptococcus pneumoniae. Infect. Immun. 1999, 67, 1683–1687. [Google Scholar] [CrossRef]
- Dhaenens, L.; Szczebara, F.; Husson, M. Identification, characterization, and immunogenicity of the lactoferrin-binding protein from Helicobacter pylori. Infect. Immun. 1997, 65, 514–518. [Google Scholar] [CrossRef] [PubMed]
- Schryvers, A. Identification of the transferrin-and lactoferrin-binding proteins in Haemophilus influenzae. J. Med. Microbiol. 1989, 29, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Moshynskyy, I.; Jiang, M.; Fontaine, M.C.; Perez-Casal, J.; Babiuk, L.A.; Potter, A.A. Characterization of a bovine lactoferrin binding protein of Streptococcus uberis. Microb. Pathog. 2003, 35, 203–215. [Google Scholar] [CrossRef]
- Naidu, A.; Andersson, M.; Forsgren, A. Identification of a human lactoferrin-binding protein in Staphylococcus aureus. J. Med. Microbiol. 1992, 36, 177–183. [Google Scholar] [CrossRef]
- Seifert, A.; Freilich, S.; Kashi, Y.; Livney, Y.D. Protein-oligosaccharide conjugates as novel prebiotics. Polym. Adv. Technol. 2019, 30, 2577–2585. [Google Scholar] [CrossRef]
- Peled, S.; Livney, Y.D. Oligosaccharide-lactoferrin shell-crosslinked particles for selective targeting of proteins to probiotic bacteria in the colon. Food Hydrocoll. 2021, 120, 106973. [Google Scholar] [CrossRef]
- Chen, P.W.; Jheng, T.T.; Shyu, C.L.; Mao, F.C. Synergistic antibacterial efficacies of the combination of bovine lactoferrin or its hydrolysate with probiotic secretion in curbing the growth of meticillin-resistant Staphylococcus aureus. J. Med. Microbiol. 2013, 62, 1845–1851. [Google Scholar] [CrossRef]
- De Bortoli, N.; Leonardi, G.; Ciancia, E.; Merlo, A.; Bellini, M.; Costa, F.; Mumolo, M.G.; Ricchiuti, A.; Cristiani, F.; Santi, S. Helicobacter pylori Eradication: A Randomized Prospective Study of Triple Therapy: Versus: Triple Therapy Plus Lactoferrin and Probiotics. Off. J. Am. Coll. Gastroenterol. ACG 2007, 102, 951–956. [Google Scholar] [CrossRef]
- Tolone, S.; Pellino, V.; Vitaliti, G.; Lanzafame, A.; Tolone, C. Evaluation of Helicobacter Pylori eradication in pediatric patients by triple therapy plus lactoferrin and probiotics compared to triple therapy alone. Ital. J. Pediatr. 2012, 38, 63. [Google Scholar] [CrossRef]
- Al-Alaiyan, S.; Abdulaziz, N.; Alkohlani, A.; Almairi, S.O.; Al Hazzani, F.; Binmanee, A.; Alfattani, A.; Almairi, S.O.A.M. Effects of probiotics and lactoferrin on necrotizing enterocolitis in preterm infants. Cureus 2021, 13, e18256. [Google Scholar] [CrossRef]
- Manzoni, P.; Sánchez, R.G.; Meyer, M.; Stolfi, I.; Pugni, L.; Messner, H.; Cattani, S.; Betta, P.M.; Memo, L.; Decembrino, L. Exposure to gastric acid inhibitors increases the risk of infection in preterm very low birth weight infants but concomitant administration of lactoferrin counteracts this effect. J. Pediatr. 2018, 193, 62–67.e1. [Google Scholar] [CrossRef]
- Yu, H.; Yu, J.; Zhu, Y.; Li, Y.; Tang, L. Comparison of Improved Effect of Antibacterial and Antiviral Activity of Four Probiotic Lactobacillus Expressing Porcine Lactoferrin in Mice. Pak. Vet. J. 2015, 35, 274–278. [Google Scholar]
- Chen, H.L.; Lai, Y.W.; Chen, C.S.; Chu, T.W.; Lin, W.; Yen, C.C.; Lin, M.F.; Tu, M.Y.; Chen, C.M. Probiotic Lactobacillus casei expressing human lactoferrin elevates antibacterial activity in the gastrointestinal tract. Biometals 2010, 23, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.; Liu, S.; Wang, H.; Su, H.; Liu, Z. Enhanced antifungal activity of bovine lactoferrin-producing probiotic Lactobacillus casei in the murine model of vulvovaginal candidiasis. BMC Microbiol. 2019, 19, 7. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zong, X.; Han, B.; Li, Y.; Tang, L. Lactobacillus pentosus expressing porcine lactoferrin elevates antibacterial activity and improves the efficacy of vaccination against Aujeszky’s disease. Acta Vet. Hung. 2016, 64, 289–300. [Google Scholar] [CrossRef]
- Liu, Z.-S.; Lin, C.-F.; Lee, C.-P.; Hsieh, M.-C.; Lu, H.-F.; Chen, Y.-F.; Ku, Y.-W.; Chen, P.-W. A Single Plasmid of Nisin-Controlled Bovine and Human Lactoferrin Expressing Elevated Antibacterial Activity of Lactoferrin-Resistant Probiotic Strains. Antibiotics 2021, 10, 120. [Google Scholar] [CrossRef]
- Generally Recognized as Safe (GRAS). Notification 000423 for Cow’s Milk-Derived Lactoferrin as a Component of Cow’s Milk-Based Infant Formulas, Cow’s Milk Products, and Chewing Gum; Generally Recognized as Safe (GRAS), 2011. Available online: https://www.fda.gov/downloads/Food/IngredientsPackagingLabeling/GRAS/NoticeInventory/ucm303340.pdf (accessed on 9 May 2023).
- Tetens, I. EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA): Scientific Opinion on Bovine Lactoferrin; EFSA-Q-2010-01269; European Food Safety Authority: Parma, Italy, 2012. [Google Scholar]
- van der Velden, W.J.; van Iersel, T.M.; Blijlevens, N.; Donnelly, J.P. Safety and tolerability of the antimicrobial peptide human lactoferrin 1-11 (hLF1-11). BMC Med. 2009, 7, 44. [Google Scholar] [CrossRef]
- Chan, H.; Chan, G.; Santos, J.; Dee, K.; Co, J.K. A randomized, double-blind, placebo-controlled trial to determine the efficacy and safety of lactoferrin with vitamin E and zinc as an oral therapy for mild to moderate acne vulgaris. Int. J. Dermatol. 2017, 56, 686–690. [Google Scholar] [CrossRef]
- Taylor, S.; Brock, J.; Kruger, C.; Berner, T.; Murphy, M. Safety determination for the use of bovine milk-derived lactoferrin as a component of an antimicrobial beef carcass spray. Regul. Toxicol. Pharmacol. 2004, 39, 12–24. [Google Scholar] [CrossRef]
- Chioukh, F.; Hmida, H.B.; Ameur, K.B.; Toumi, A.; Monastiri, K. Saccharomyces cerevisiae fungemia in a premature neonate treated receiving probiotics. Med. Mal. Infect. 2013, 43, 359–360. [Google Scholar] [CrossRef]
- Dani, C.; Coviello, C.; Corsini, I.; Arena, F.; Antonelli, A.; Rossolini, G.M. Lactobacillus sepsis and probiotic therapy in newborns: Two new cases and literature review. Am. J. Perinatol. Rep. 2015, 6, e25–e29. [Google Scholar]
- Patel, R.M.; Underwood, M.A. Probiotics and necrotizing enterocolitis. Semin. Pediatr. Surg. 2018, 27, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Vallejo-Cordoba, B.; Castro-López, C.; García, H.S.; González-Córdova, A.F.; Hernández-Mendoza, A. Postbiotics and paraprobiotics: A review of current evidence and emerging trends. Adv. Food Nutr. Res. 2020, 94, 1–34. [Google Scholar] [PubMed]
- Teame, T.; Wang, A.; Xie, M.; Zhang, Z.; Yang, Y.; Ding, Q.; Gao, C.; Olsen, R.E.; Ran, C.; Zhou, Z. Paraprobiotics and postbiotics of probiotic Lactobacilli, their positive effects on the host and action mechanisms: A review. Front. Nutr. 2020, 7, 570344. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Probiotic | Form of Lactoferrin | Culture Condition | Dose and Key Features | References |
|---|---|---|---|---|
| Bifidobacterium breve BCRC 12584 | Apo-BLF | 22 °C, anaerobic | Dose–response growth (>1 mg/mL) | [33] |
| Bifidobacterium longum ATCC 15707 | Bulk BLF | 37 °C, anaerobic | A slight increase in growth response (2 and 4 mg/mL) | [29] |
| Bulk BLF | 37 °C, anaerobic | Growth was promoted (1 mg/mL) | [44] | |
| Bifidobacterium longum ATCC 15708 | Bulk BLF | 37 °C, anaerobic | Good increase in growth response (2 and 4 mg/mL) | [29] |
| Bifidobacterium longum kd-5-6 | Bulk BLF | 37 °C, anaerobic | Good increase in growth response (2 and 4 mg/mL) | [29] |
| Loigolactobacillus coryniformis subsp. coryniformis ATCC 25602 | Apo-BLF | 22 °C, anaerobic | Dose–response growth (>1 mg/mL) | [33] |
| Lactobacillus delbrueckii BCRC 140 | Apo-BLF | 22 °C, anaerobic | Dose–response growth (>1 mg/mL) | [33] |
| Lactobacillus acidophilus BCRC 14065 | Apo-BLF | 22 °C, anaerobic | Dose–response growth (>1 mg/mL) | [33] |
| Bifidobacterium angulatum ATCC 27535 | Apo-BLF | 22 °C, anaerobic | Dose–response growth (>1 mg/mL) | [33] |
| Bifidobacterium catenulatum ATCC 27539 | Apo-BLF | 22 °C, anaerobic | Dose–response growth (>1 mg/mL) | [33] |
| Lactiplantibacillus paraplantarum ATCC 70021 | Apo-BLF | 22 °C, anaerobic | Dose–response growth (>1 mg/mL) | [33] |
| Pediococcus pentosaceus ATCC 8081 | Apo-BLF | 22 °C, anaerobic | Growth enhancement (>1 mg/mL) | [33] |
| Lacticaseibacillus rhamnosus ATCC 53103 | Apo-BLF | 22 °C, anaerobic | Growth enhancement (>1 mg/mL) | [33] |
| Lacticaseibacillus paracasei BCRC 17483 | Apo-BLF | 22 °C, anaerobic | Growth enhancement (>1 mg/mL) | [33] |
| Lactobacillus acidophilus ATCC 4356 | Apo-BLF | 37 °C, anaerobic | Growth inhibition 8 to 16 mg/mL | [32] |
| BLF hydrolysate | 37 °C, anaerobic | Growth inhibition 8 to 16 mg/mL | [32] | |
| Ligilactobacillus salivarius ATCC 11741 | Apo-BLF | 37 °C, anaerobic | Growth inhibition 32 mg/mL | [32] |
| BLF hydrolysate | 37 °C, anaerobic | Growth inhibition 32 mg/mL | [32] | |
| Lacticaseibacillus rhamnosus ATCC 53103 | Apo-BLF | 37 °C, anaerobic | Growth inhibition 1 to 16 mg/mL | [32] |
| BLF hydrolysate | 37 °C, anaerobic | Growth inhibition 1 to 16 mg/mL | [32] | |
| Bifidobacterium longum ATCC 15707 | Apo-BLF | 37 °C, anaerobic | Growth inhibition 2 to 4 mg/mL | [32] |
| BLF hydrolysate | 37 °C, anaerobic | Growth inhibition 2 to 4 mg/mL | [32] | |
| Bifidobacterium lactis BCRC 17394 | Apo-BLF | 37 °C, anaerobic | Growth inhibition 2 to 8 mg/mL | [32] |
| BLF hydrolysate | 37 °C, anaerobic | Growth inhibition 2 to 8 mg/mL | [32] | |
| Bifidobacterium bifidum ATCC 15696 | BLF hydrolysate | 37 °C, anaerobic | Dose–response growth (0.01 to 1 mg/mL) | [45] |
| Bifidobacterium longum subsp. infantis ATCC 15697 | BLF hydrolysate | 37 °C, anaerobic | Dose–response growth (0.01 to 1 mg/mL) | [45] |
| Apo-BLF | 37 °C, aerobic, and anaerobic | Dose-dependent inhibition; MIC: 4 to 32 mg/mL | [46] | |
| Bifidobacterium breve ATCC 15700 | BLF hydrolysate | 37 °C, anaerobic | Dose–response growth (0.01 to 1 mg/mL) | [45] |
| Bifidobacterium bifidum ATCC 29521 | Apo-BLF | 37 °C, aerobic, and anaerobic | Dose-dependent inhibition (>0.25 mg/mL); MIC: 128 mg/mL | [46] |
| Limosilactobacillus reuteri ATCC 23272 | Apo-BLF | 37 °C, aerobic, and anaerobic | Dose-dependent inhibition (>0.25 mg/mL); MIC: 64 to 128 mg/mL | [46] |
| Loigolactobacillus coryniformis subsp. coryniformis ATCC 25602 | Apo-BLF | 37 °C, aerobic, and anaerobic | Dose-dependent inhibition (>0.25 mg/mL); MIC: 4 to 8 mg/mL | [46] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.-S.; Chen, P.-W. Featured Prebiotic Agent: The Roles and Mechanisms of Direct and Indirect Prebiotic Activities of Lactoferrin and Its Application in Disease Control. Nutrients 2023, 15, 2759. https://doi.org/10.3390/nu15122759
Liu Z-S, Chen P-W. Featured Prebiotic Agent: The Roles and Mechanisms of Direct and Indirect Prebiotic Activities of Lactoferrin and Its Application in Disease Control. Nutrients. 2023; 15(12):2759. https://doi.org/10.3390/nu15122759
Chicago/Turabian StyleLiu, Zhen-Shu, and Po-Wen Chen. 2023. "Featured Prebiotic Agent: The Roles and Mechanisms of Direct and Indirect Prebiotic Activities of Lactoferrin and Its Application in Disease Control" Nutrients 15, no. 12: 2759. https://doi.org/10.3390/nu15122759
APA StyleLiu, Z.-S., & Chen, P.-W. (2023). Featured Prebiotic Agent: The Roles and Mechanisms of Direct and Indirect Prebiotic Activities of Lactoferrin and Its Application in Disease Control. Nutrients, 15(12), 2759. https://doi.org/10.3390/nu15122759