
Geraniin Ameliorates Hypertensive Vascular Remodelling in a Diet-Induced Obese Animal Model through Antioxidant and Anti-Inflammatory Effects

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Geraniin Extraction and Purification
2.2. Animal Ethics and Housing Conditions
2.3. Diet Preparation, Composition, and Treatment
2.4. Blood Pressure Measurement
2.5. Blood Plasma and Tissue Collection
2.6. Isolation of Peripheral Blood Mononuclear Cells (PBMCs) from the Whole Blood Sample
2.7. Plasma Biochemical Assays
2.8. Vasomotor Assessment of the Thoracic Aorta
2.9. Histological Examination of the Thoracic Aorta
2.10. Protein Extraction and Gelatin Zymography
Western Blot of β-Actin Expression
2.11. Lucigenin-Chemiluminescence Assay
2.12. RNA Extraction and Real-Time PCR
2.13. Statistical Analysis
3. Results
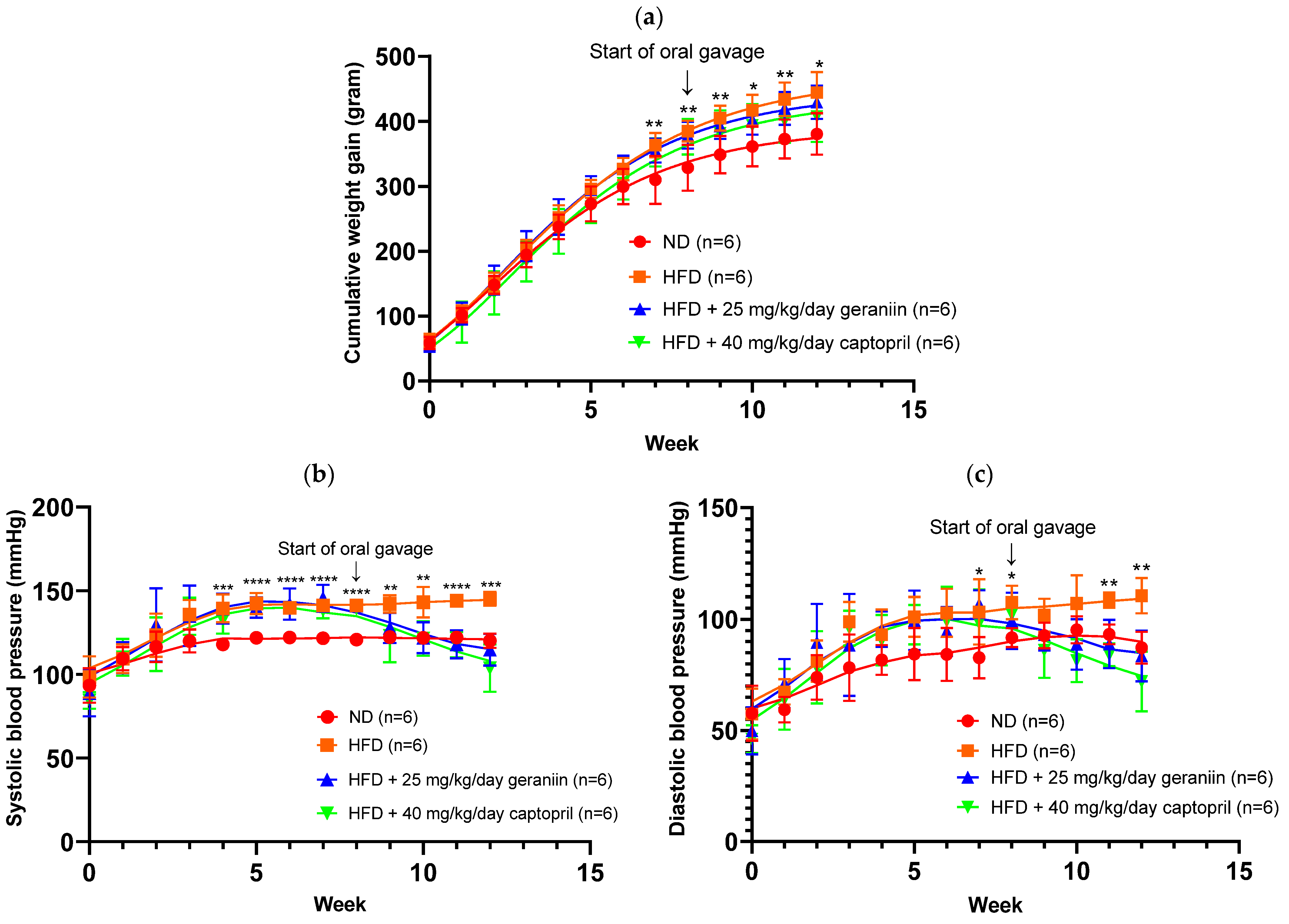
3.1. Geraniin Ameliorated Increase in SBP and DBP Induced by HFD
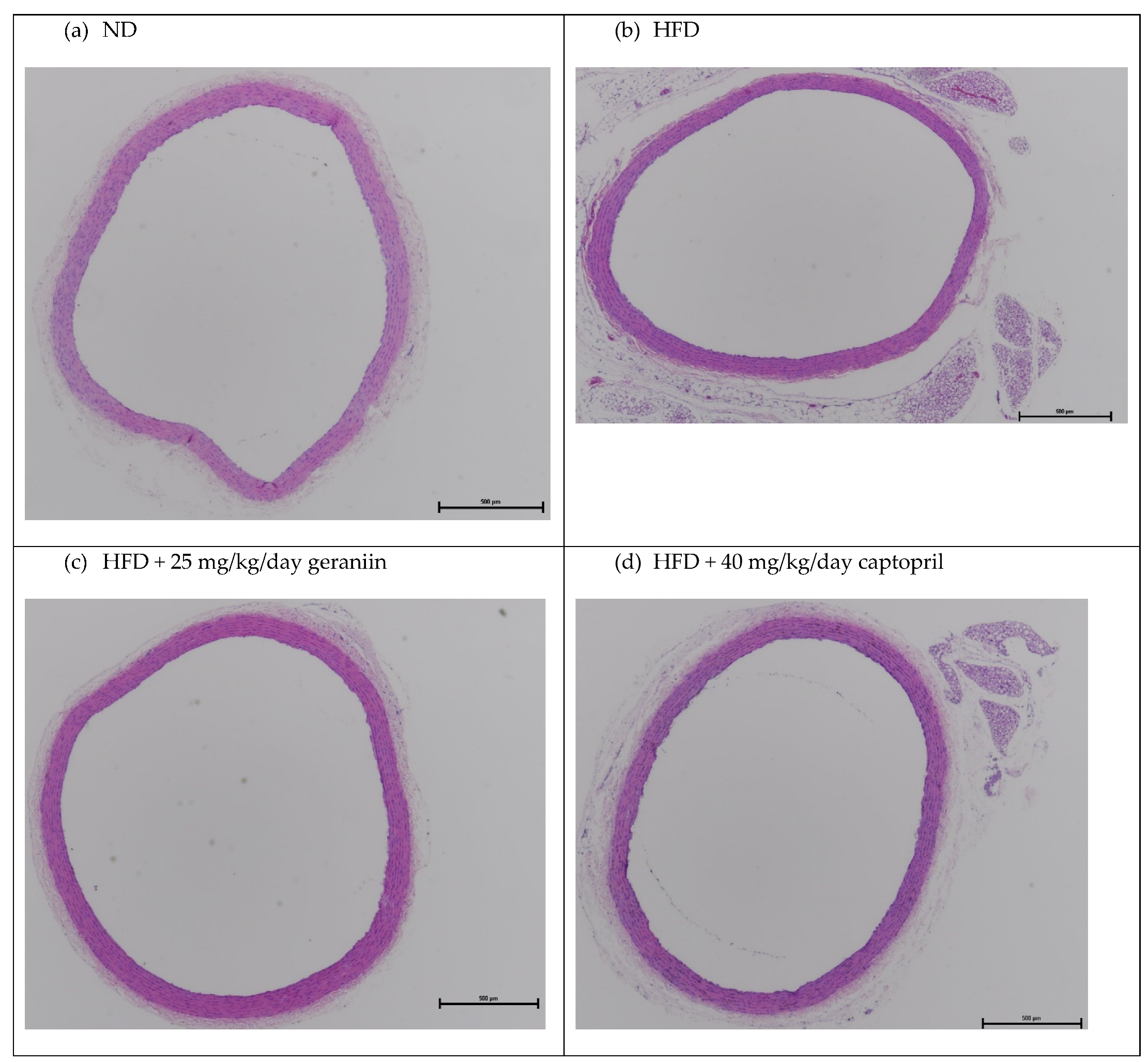
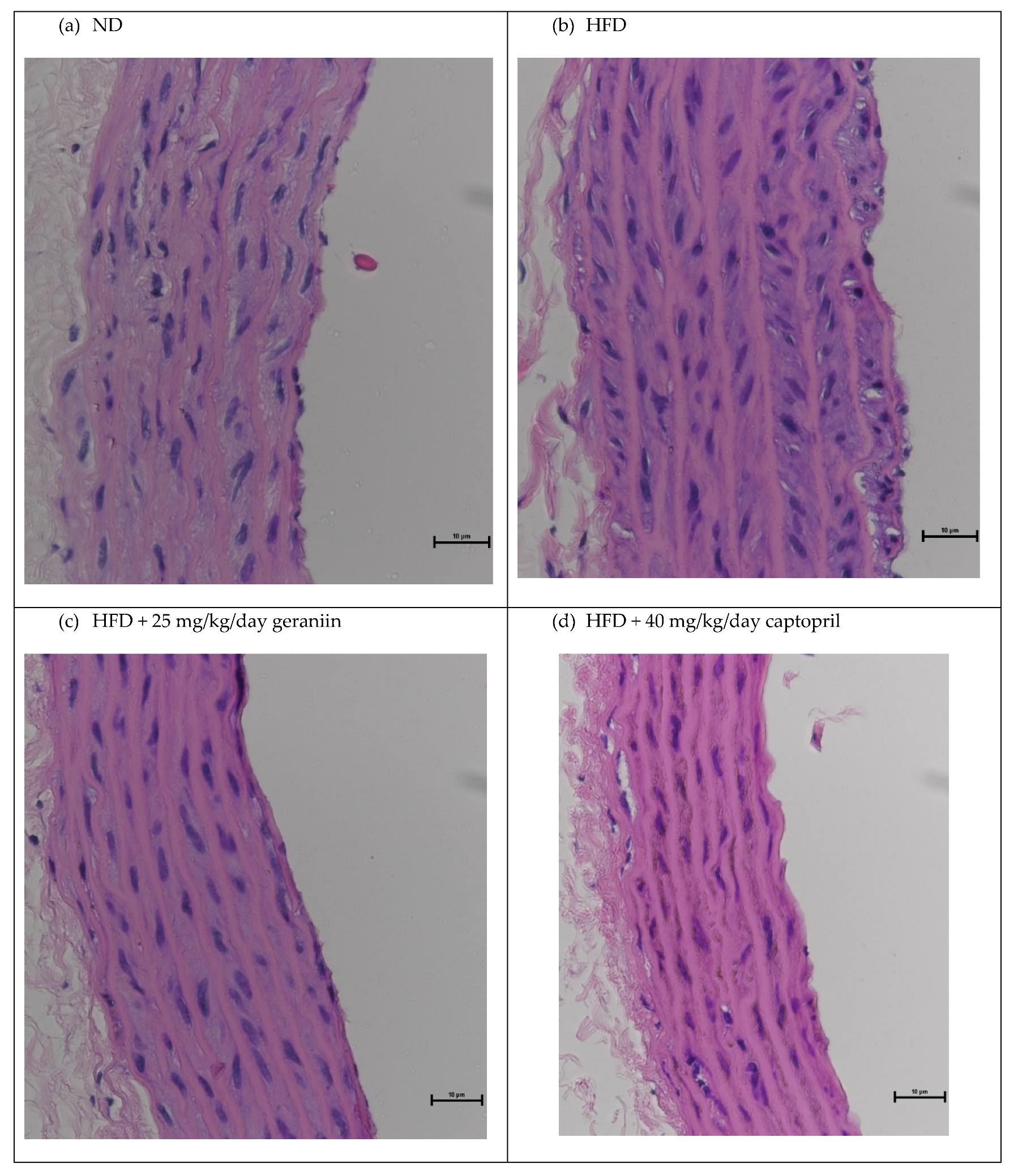
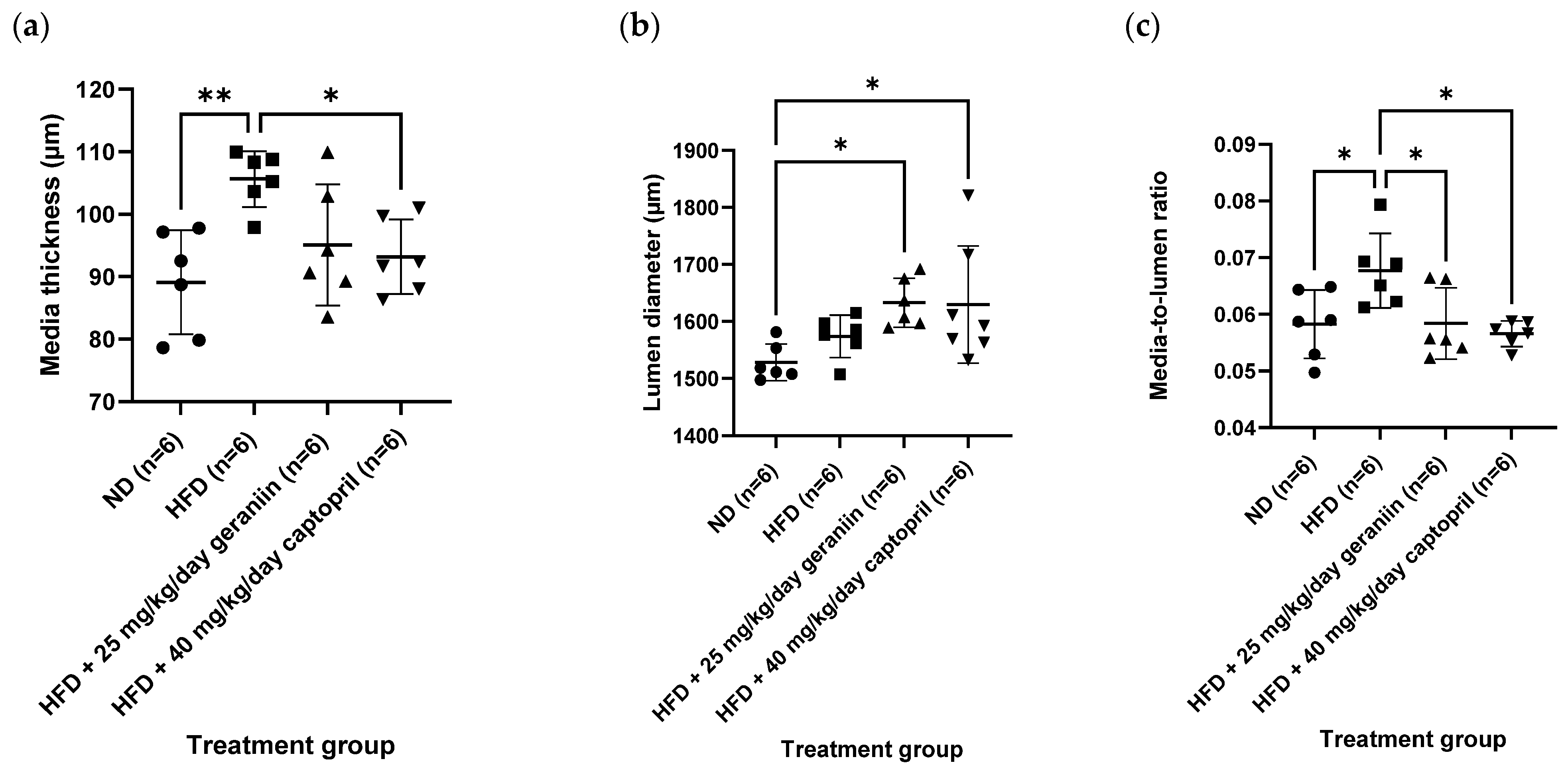
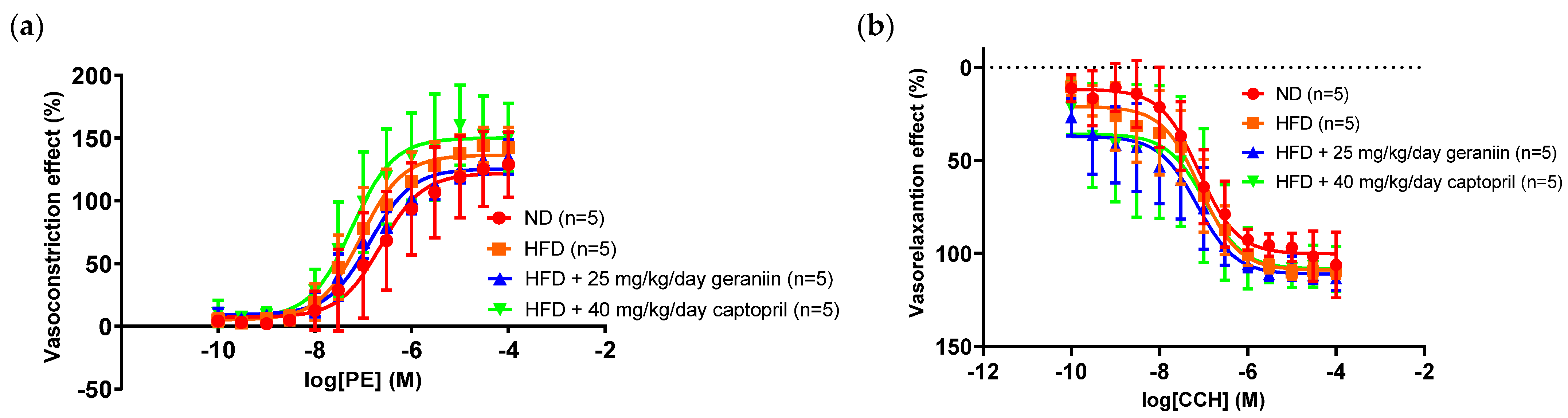
3.2. Geraniin Reversed HFD-Induced Vascular Remodelling
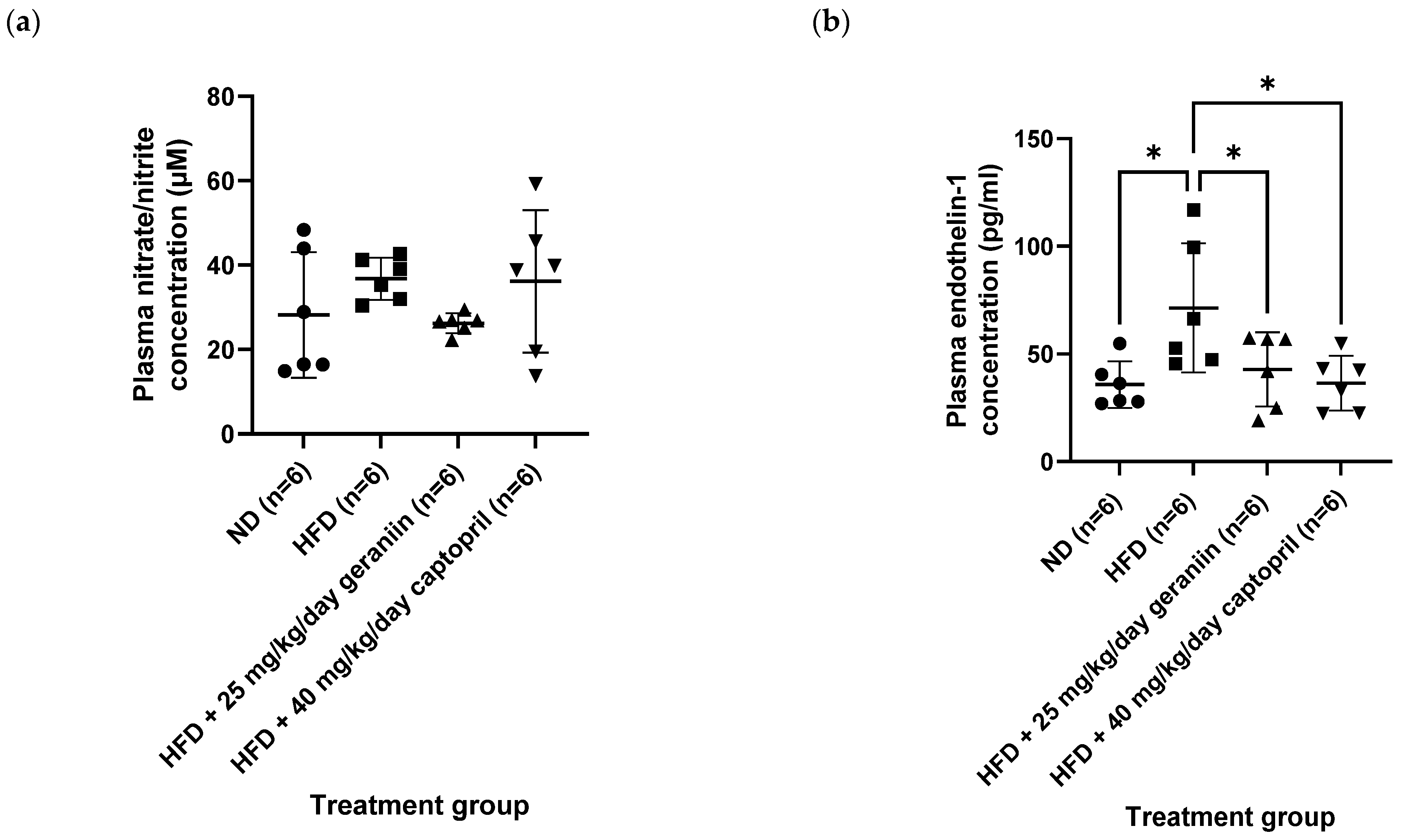
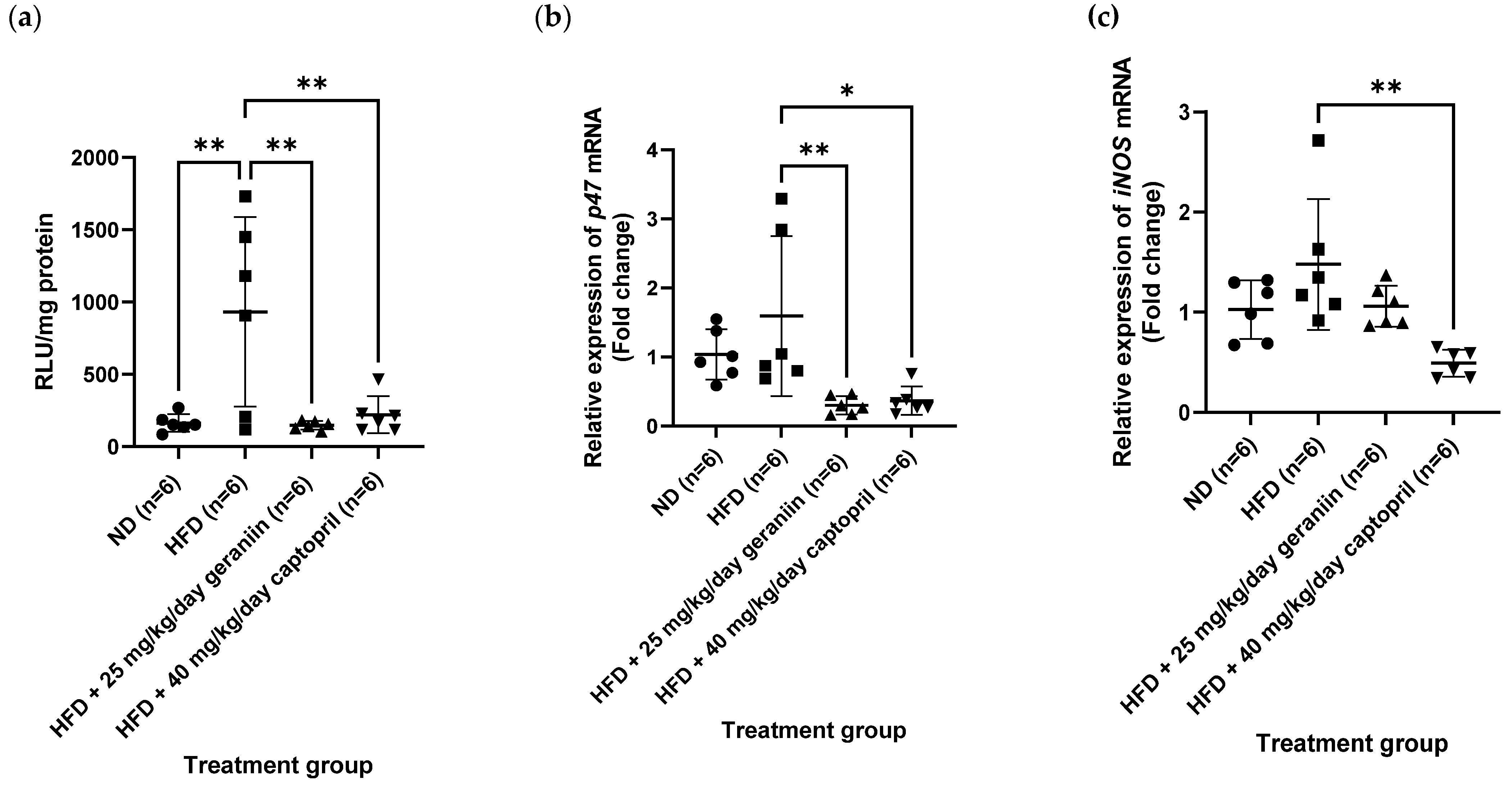
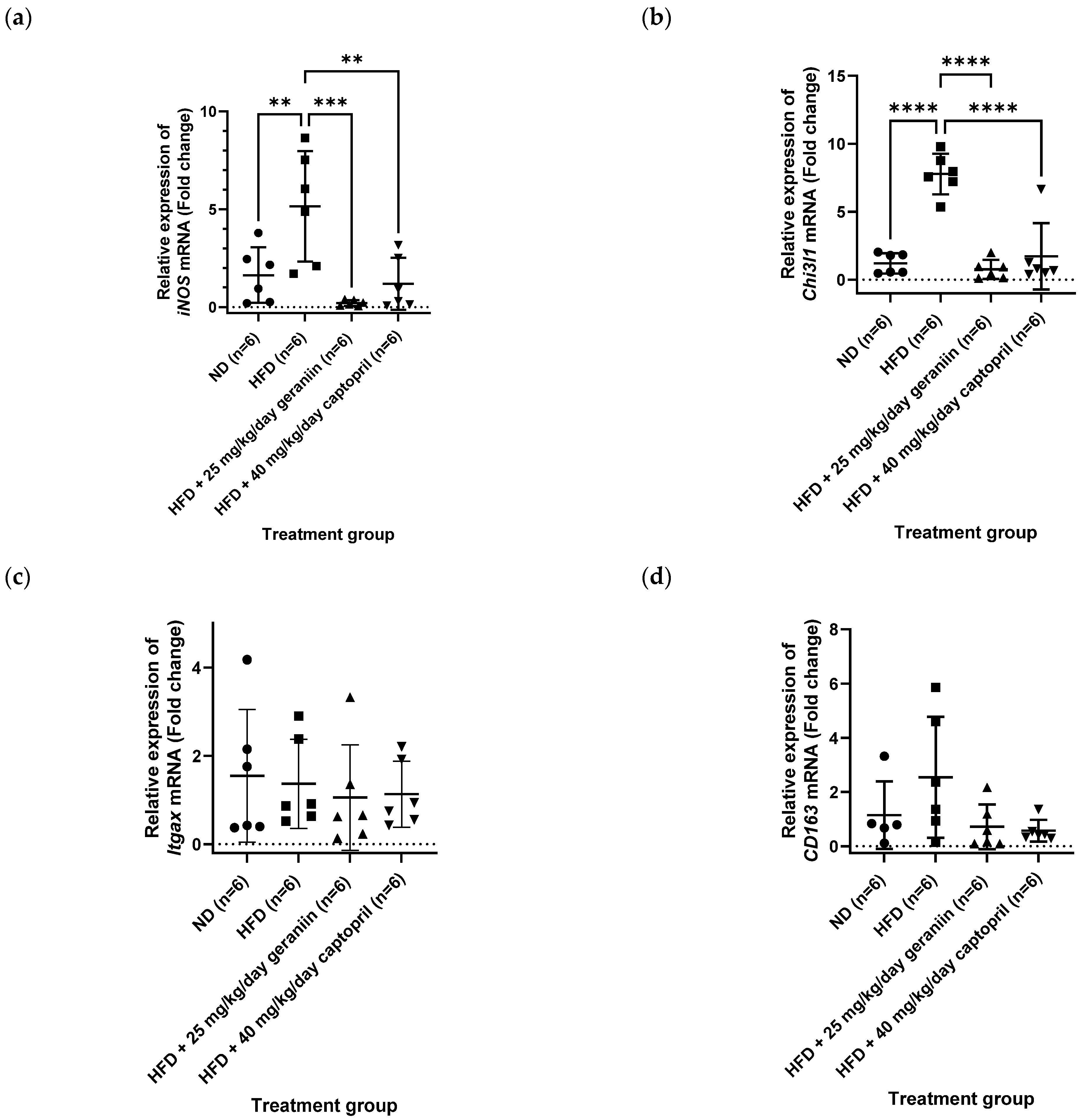
3.3. Geraniin Mitigated Excessive Superoxide Radical Production in the Thoracic Aorta and Systemic Inflammation Induced by HFD
4. Discussion
Study Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Cardiovascular Diseases (CVDs). Available online: https://www.who.int/news-room/fact-sheets/detail/cardiovascular-diseases-(cvds) (accessed on 1 April 2023).
- Jung, U.; Choi, M.-S. Obesity and Its Metabolic Complications: The Role of Adipokines and the Relationship between Obesity, Inflammation, Insulin Resistance, Dyslipidemia and Nonalcoholic Fatty Liver Disease. Int. J. Mol. Sci. 2014, 15, 6184–6223. [Google Scholar] [CrossRef]
- Garrison, R.J.; Kannel, W.B.; Stokes, J., 3rd; Castelli, W.P. Incidence and precursors of hypertension in young adults: The Framingham Offspring Study. Prev. Med. 1987, 16, 235–251. [Google Scholar] [CrossRef] [PubMed]
- Leggio, M.; Lombardi, M.; Caldarone, E.; Severi, P.; D’Emidio, S.; Armeni, M.; Bravi, V.; Bendini, M.G.; Mazza, A. The relationship between obesity and hypertension: An updated comprehensive overview on vicious twins. Hypertens. Res. 2017, 40, 947–963. [Google Scholar] [CrossRef] [PubMed]
- Dinh, Q.N.; Drummond, G.R.; Sobey, C.G.; Chrissobolis, S. Roles of inflammation, oxidative stress, and vascular dysfunction in hypertension. BioMed Res. Int. 2014, 2014, 406960. [Google Scholar] [CrossRef]
- Renna, N.F.; de Las Heras, N.; Miatello, R.M. Pathophysiology of vascular remodeling in hypertension. Int. J. Hypertens. 2013, 2013, 808353. [Google Scholar] [CrossRef] [PubMed]
- Ou, H.C.; Lee, W.J.; Lee, S.D.; Huang, C.Y.; Chiu, T.H.; Tsai, K.L.; Hsu, W.C.; Sheu, W.H. Ellagic acid protects endothelial cells from oxidized low-density lipoprotein-induced apoptosis by modulating the PI3K/Akt/eNOS pathway. Toxicol. Appl. Pharmacol. 2010, 248, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Chikezie, P.C.; Ojiako, O.A.; Ogbuji, A.C. Oxidative Stress in Diabetes Mellitus. Int. J. Biol. Chem. 2015, 9, 92–109. [Google Scholar] [CrossRef]
- Ghosh, A.; Gao, L.; Thakur, A.; Siu, P.M.; Lai, C.W.K. Role of free fatty acids in endothelial dysfunction. J. Biomed. Sci. 2017, 24, 50. [Google Scholar] [CrossRef]
- Mouton, A.J.; Li, X.; Hall, M.E.; Hall, J.E. Obesity, Hypertension, and Cardiac Dysfunction: Novel Roles of Immunometabolism in Macrophage Activation and Inflammation. Circ. Res. 2020, 126, 789–806. [Google Scholar] [CrossRef]
- Rudrapal, M.; Khairnar, S.J.; Khan, J.; Dukhyil, A.B.; Ansari, M.A.; Alomary, M.N.; Alshabrmi, F.M.; Palai, S.; Deb, P.K.; Devi, R. Dietary Polyphenols and Their Role in Oxidative Stress-Induced Human Diseases: Insights Into Protective Effects, Antioxidant Potentials and Mechanism(s) of Action. Front. Pharmacol. 2022, 13, 806470. [Google Scholar] [CrossRef]
- Cheng, H.S.; Ton, S.H.; Abdul Kadir, K. Ellagitannin geraniin: A review of the natural sources, biosynthesis, pharmacokinetics and biological effects. Phytochem. Rev. 2017, 16, 159–193. [Google Scholar] [CrossRef]
- Perera, A.; Appleton, D.; Ying, L.H.; Elendran, S.; Palanisamy, U.D. Large scale purification of geraniin from Nephelium lappaceum rind waste using reverse-phase chromatography. Sep. Purif. Technol. 2012, 98, 145–149. [Google Scholar] [CrossRef]
- Phang, S.C.W.; Palanisamy, U.D.; Kadir, K.A. Effects of geraniin (rambutan rind extract) on blood pressure and metabolic parameters in rats fed high-fat diet. J. Integr. Med. 2019, 17, 100–106. [Google Scholar] [CrossRef]
- Cheng, H.S.; Goh, B.H.; Phang, S.C.W.; Amanullah, M.M.; Ton, S.H.; Palanisamy, U.D.; Abdul Kadir, K.; Tan, J.B.L. Pleiotropic ameliorative effects of ellagitannin geraniin against metabolic syndrome induced by high-fat diet in rats. Nutrition 2020, 79–80, 110973. [Google Scholar] [CrossRef]
- Cheng, J.T.; Chang, S.S.; Hsu, F.L. Antihypertensive action of geraniin in rats. J. Pharm. Pharmacol. 1994, 46, 46–49. [Google Scholar] [CrossRef]
- Lin, S.Y.; Wang, C.C.; Lu, Y.L.; Wu, W.C.; Hou, W.C. Antioxidant, anti-semicarbazide-sensitive amine oxidase, and anti-hypertensive activities of geraniin isolated from Phyllanthus urinaria. Food Chem. Toxicol. 2008, 46, 2485–2492. [Google Scholar] [CrossRef]
- Chung, A.; Gurtu, S.; Chakravarthi, S.; Moorthy, M.; Palanisamy, U.D. Geraniin Protects High-Fat Diet-Induced Oxidative Stress in Sprague Dawley Rats. Front. Nutr. 2018, 5, 17. [Google Scholar] [CrossRef]
- de Kloet, A.D.; Krause, E.G.; Kim, D.H.; Sakai, R.R.; Seeley, R.J.; Woods, S.C. The effect of angiotensin-converting enzyme inhibition using captopril on energy balance and glucose homeostasis. Endocrinology 2009, 150, 4114–4123. [Google Scholar] [CrossRef] [PubMed]
- Miguel-Carrasco, J.L.; Zambrano, S.; Blanca, A.J.; Mate, A.; Vázquez, C.M. Captopril reduces cardiac inflammatory markers in spontaneously hypertensive rats by inactivation of NF-kB. J. Inflamm. 2010, 7, 21. [Google Scholar] [CrossRef]
- Davidson, E.P.; Coppey, L.J.; Dake, B.; Yorek, M.A. Effect of Treatment of Sprague Dawley Rats with AVE7688, Enalapril, or Candoxatril on Diet-Induced Obesity. J. Obes. 2011, 2011, 686952. [Google Scholar] [CrossRef]
- Lu, X.; Bean, J.S.; Kassab, G.S.; Rekhter, M.D. Protein kinase C inhibition ameliorates functional endothelial insulin resistance and vascular smooth muscle cell hypersensitivity to insulin in diabetic hypertensive rats. Cardiovasc. Diabetol. 2011, 10, 48. [Google Scholar] [CrossRef]
- Gohar, A.A.; Lahloub, M.E.; Niwa, M. Antibacterial polyphenol from Erodium glaucophyllum. Z. Nat. C J. Biosci. 2003, 58, 670–674. [Google Scholar] [CrossRef]
- Cheng, H.S.; Ton, S.H.; Phang, S.C.W.; Tan, J.B.L.; Abdul Kadir, K. Increased susceptibility of post-weaning rats on high-fat diet to metabolic syndrome. J. Adv. Res. 2017, 8, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Hoggatt, A.F.; Hoggatt, J.; Honerlaw, M.; Pelus, L.M. A spoonful of sugar helps the medicine go down: A novel technique to improve oral gavage in mice. J. Am. Assoc. Lab. Anim. Sci. 2010, 49, 329–334. [Google Scholar] [PubMed]
- Goshi, E.; Zhou, G.; He, Q. Nitric oxide detection methods in vitro and in vivo. Med. Gas Res. 2019, 9, 192–207. [Google Scholar] [CrossRef]
- Loong, B.J.; Tan, J.H.; Lim, K.H.; Mbaki, Y.; Ting, K.N. Contractile function of smooth muscle retained after overnight storage. Naunyn Schmiedeberg’s Arch. Pharmacol. 2015, 388, 1061–1067. [Google Scholar] [CrossRef]
- Su, J.; Xu, H.-T.; Yu, J.-J.; Gao, J.-L.; Lei, J.; Yin, Q.-S.; Li, B.; Pang, M.-X.; Su, M.-X.; Mi, W.-J.; et al. Luteolin Ameliorates Hypertensive Vascular Remodeling through Inhibiting the Proliferation and Migration of Vascular Smooth Muscle Cells. Evid. Based Complement. Altern. Med. 2015, 2015, 364876. [Google Scholar] [CrossRef] [PubMed]
- Guzik, T.J.; Channon, K.M. Measurement of vascular reactive oxygen species production by chemiluminescence. Methods Mol. Med. 2005, 108, 73–89. [Google Scholar] [CrossRef]
- Nakmareong, S.; Kukongviriyapan, U.; Pakdeechote, P.; Donpunha, W.; Kukongviriyapan, V.; Kongyingyoes, B.; Sompamit, K.; Phisalaphong, C. Antioxidant and vascular protective effects of curcumin and tetrahydrocurcumin in rats with L-NAME-induced hypertension. Naunyn Schmiedeberg’s Arch. Pharmacol. 2011, 383, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Guzmán, M.; Jiménez, R.; Sánchez, M.; Zarzuelo, M.J.; Galindo, P.; Quintela, A.M.; López-Sepúlveda, R.; Romero, M.; Tamargo, J.; Vargas, F.; et al. Epicatechin lowers blood pressure, restores endothelial function, and decreases oxidative stress and endothelin-1 and NADPH oxidase activity in DOCA-salt hypertension. Free Radic. Biol. Med. 2012, 52, 70–79. [Google Scholar] [CrossRef]
- Boonla, O.; Kukongviriyapan, U.; Pakdeechote, P.; Kukongviriyapan, V.; Pannangpetch, P.; Prachaney, P.; Greenwald, S.E. Curcumin improves endothelial dysfunction and vascular remodeling in 2K-1C hypertensive rats by raising nitric oxide availability and reducing oxidative stress. Nitric Oxide 2014, 42, 44–53. [Google Scholar] [CrossRef]
- Andriantsitohaina, R.; Auger, C.; Chataigneau, T.; Étienne-Selloum, N.; Li, H.; Martínez, M.C.; Schini-Kerth, V.B.; Laher, I. Molecular mechanisms of the cardiovascular protective effects of polyphenols. Br. J. Nutr. 2012, 108, 1532–1549. [Google Scholar] [CrossRef]
- Mendizábal, Y.; Llorens, S.; Nava, E. Hypertension in metabolic syndrome: Vascular pathophysiology. Int. J. Hypertens. 2013, 2013, 230868. [Google Scholar] [CrossRef] [PubMed]
- Munkong, N.; Hansakul, P.; Yoysungnoen, B.; Wongnoppavich, A.; Sireeratawong, S.; Kaendee, N.; Lerdvuthisopon, N. Vasoprotective effects of rice bran water extract on rats fed with high-fat diet. Asian Pac. J. Trop. Biomed. 2016, 6, 778–784. [Google Scholar] [CrossRef]
- Mahmoud, A.A.; Elshazly, S.M. Ursodeoxycholic acid ameliorates fructose-induced metabolic syndrome in rats. PLoS ONE 2014, 9, e106993. [Google Scholar] [CrossRef] [PubMed]
- Senaphan, K.; Sangartit, W.; Pakdeechote, P.; Kukongviriyapan, V.; Pannangpetch, P.; Thawornchinsombut, S.; Greenwald, S.E.; Kukongviriyapan, U. Rice bran protein hydrolysates reduce arterial stiffening, vascular remodeling and oxidative stress in rats fed a high-carbohydrate and high-fat diet. Eur. J. Nutr. 2018, 57, 219–230. [Google Scholar] [CrossRef]
- Kosmopoulos, M.; Drekolias, D.; Zavras, P.D.; Piperi, C.; Papavassiliou, A.G. Impact of advanced glycation end products (AGEs) signaling in coronary artery disease. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 611–619. [Google Scholar] [CrossRef]
- Kostov, K. The Causal Relationship between Endothelin-1 and Hypertension: Focusing on Endothelial Dysfunction, Arterial Stiffness, Vascular Remodeling, and Blood Pressure Regulation. Life 2021, 11, 986. [Google Scholar] [CrossRef]
- Ozkor, M.A.; Quyyumi, A.A. Endothelium-derived hyperpolarizing factor and vascular function. Cardiol. Res. Pract. 2011, 2011, 156146. [Google Scholar] [CrossRef]
- Zhang, J.X.; Yang, J.R.; Chen, G.X.; Tang, L.J.; Li, W.X.; Yang, H.; Kong, X. Sesamin ameliorates arterial dysfunction in spontaneously hypertensive rats via downregulation of NADPH oxidase subunits and upregulation of eNOS expression. Acta Pharmacol. Sin. 2013, 34, 912–920. [Google Scholar] [CrossRef]
- Berkban, T.; Boonprom, P.; Bunbupha, S.; Welbat, J.U.; Kukongviriyapan, U.; Kukongviriyapan, V.; Pakdeechote, P.; Prachaney, P. Ellagic Acid Prevents L-NAME-Induced Hypertension via Restoration of eNOS and p47phox Expression in Rats. Nutrients 2015, 7, 5265–5280. [Google Scholar] [CrossRef] [PubMed]
- Schramm, A.; Matusik, P.; Osmenda, G.; Guzik, T.J. Targeting NADPH oxidases in vascular pharmacology. Vasc. Pharmacol. 2012, 56, 216–231. [Google Scholar] [CrossRef] [PubMed]
- Seshiah, P.N.; Weber, D.S.; Rocic, P.; Valppu, L.; Taniyama, Y.; Griendling, K.K. Angiotensin II stimulation of NAD(P)H oxidase activity: Upstream mediators. Circ. Res. 2002, 91, 406–413. [Google Scholar] [CrossRef]
- Li, L.; Fink, G.D.; Watts, S.W.; Northcott, C.A.; Galligan, J.J.; Pagano, P.J.; Chen, A.F. Endothelin-1 Increases Vascular Superoxide via EndothelinA–NADPH Oxidase Pathway in Low-Renin Hypertension. Circulation 2003, 107, 1053–1058. [Google Scholar] [CrossRef] [PubMed]
- Basta, G.; Schmidt, A.M.; De Caterina, R. Advanced glycation end products and vascular inflammation: Implications for accelerated atherosclerosis in diabetes. Cardiovasc. Res. 2004, 63, 582–592. [Google Scholar] [CrossRef]
- Sánchez, M.; Galisteo, M.; Vera, R.; Villar, I.C.; Zarzuelo, A.; Tamargo, J.; Pérez-Vizcaíno, F.; Duarte, J. Quercetin downregulates NADPH oxidase, increases eNOS activity and prevents endothelial dysfunction in spontaneously hypertensive rats. J. Hypertens. 2006, 24, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Kuo, M.Y.; Ou, H.C.; Lee, W.J.; Kuo, W.W.; Hwang, L.L.; Song, T.Y.; Huang, C.Y.; Chiu, T.H.; Tsai, K.L.; Tsai, C.S.; et al. Ellagic acid inhibits oxidized low-density lipoprotein (OxLDL)-induced metalloproteinase (MMP) expression by modulating the protein kinase C-α/extracellular signal-regulated kinase/peroxisome proliferator-activated receptor γ/nuclear factor-κB (PKC-α/ERK/PPAR-γ/NF-κB) signaling pathway in endothelial cells. J. Agric. Food Chem. 2011, 59, 5100–5108. [Google Scholar] [CrossRef]
- Agita, A.; Alsagaff, M.T. Inflammation, Immunity, and Hypertension. Acta Med. Indones. 2017, 49, 158–165. [Google Scholar]
- Al Dubayee, M.S.; Alayed, H.; Almansour, R.; Alqaoud, N.; Alnamlah, R.; Obeid, D.; Alshahrani, A.; Zahra, M.M.; Nasr, A.; Al-Bawab, A.; et al. Differential Expression of Human Peripheral Mononuclear Cells Phenotype Markers in Type 2 Diabetic Patients and Type 2 Diabetic Patients on Metformin. Front. Endocrinol. 2018, 9, 537. [Google Scholar] [CrossRef]
- Harwani, S.C. Macrophages under pressure: The role of macrophage polarization in hypertension. Transl. Res. 2018, 191, 45–63. [Google Scholar] [CrossRef]
- Ji, Q.; Cheng, G.; Ma, N.; Huang, Y.; Lin, Y.; Zhou, Q.; Que, B.; Dong, J.; Zhou, Y.; Nie, S. Circulating Th1, Th2, and Th17 Levels in Hypertensive Patients. Dis. Markers 2017, 2017, 7146290. [Google Scholar] [CrossRef] [PubMed]
- Moorthy, M.; Wie, C.C.; Mariño, E.; Palanisamy, U.D. The Prebiotic Potential of Geraniin and Geraniin-Enriched Extract against High-Fat-Diet-Induced Metabolic Syndrome in Sprague Dawley Rats. Antioxidants 2022, 11, 632. [Google Scholar] [CrossRef] [PubMed]
- Xue, Q.; Yan, Y.; Zhang, R.; Xiong, H. Regulation of iNOS on Immune Cells and Its Role in Diseases. Int. J. Mol. Sci. 2018, 19, 3805. [Google Scholar] [CrossRef]
- Capone, M.; Maggi, L.; Santarlasci, V.; Rossi, M.C.; Mazzoni, A.; Montaini, G.; Cimaz, R.; Ramazzotti, M.; Piccinni, M.P.; Barra, G.; et al. Chitinase 3-like-1 is produced by human Th17 cells and correlates with the level of inflammation in juvenile idiopathic arthritis patients. Clin. Mol. Allergy 2016, 14, 16. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Su, Z.; Li, Y.; Zhang, X.; You, Q. Chitinase-3 like-protein-1 function and its role in diseases. Signal Transduct. Target. Ther. 2020, 5, 201. [Google Scholar] [CrossRef]
- Malinda, K.M.; Ponce, L.; Kleinman, H.K.; Shackelton, L.M.; Millis, A.J. Gp38k, a protein synthesized by vascular smooth muscle cells, stimulates directional migration of human umbilical vein endothelial cells. Exp. Cell. Res. 1999, 250, 168–173. [Google Scholar] [CrossRef]
- Junker, N.; Johansen, J.S.; Hansen, L.T.; Lund, E.L.; Kristjansen, P.E. Regulation of YKL-40 expression during genotoxic or microenvironmental stress in human glioblastoma cells. Cancer Sci. 2005, 96, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Michelsen, A.E.; Rathcke, C.N.; Skjelland, M.; Holm, S.; Ranheim, T.; Krohg-Sørensen, K.; Klingvall, M.F.; Brosstad, F.; Oie, E.; Vestergaard, H.; et al. Increased YKL-40 expression in patients with carotid atherosclerosis. Atherosclerosis 2010, 211, 589–595. [Google Scholar] [CrossRef]
- Ściborski, K.; Kuliczkowski, W.; Karolko, B.; Bednarczyk, D.; Protasiewicz, M.; Mysiak, A.; Negrusz-Kawecka, M. Plasma YKL-40 levels correlate with the severity of coronary atherosclerosis assessed with the SYNTAX score. Pol. Arch. Intern. Med. 2018, 128, 644–648. [Google Scholar] [CrossRef]
- Li, K.; Chen, Z.; Qin, Y.; Wei, Y.X. Plasm YKL-40 Levels Are Associated with Hypertension in Patients with Obstructive Sleep Apnea. BioMed Res. Int. 2019, 2019, 5193597. [Google Scholar] [CrossRef]
- Xu, T.; Zheng, X.; Wang, A.; Guo, Z.; Zhang, Y. Association of CHI3L1 gene variants with YKL-40 levels and hypertension incidence: A population-based nested case-control study in China. J. Cell. Mol. Med. 2021, 25, 919–924. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Iguchi, A.; Hatano, T. Identification of urinary and intestinal bacterial metabolites of ellagitannin geraniin in rats. J. Agric. Food Chem. 2008, 56, 393–400. [Google Scholar] [CrossRef]
- Ito, H. Metabolites of the ellagitannin geraniin and their antioxidant activities. Planta Med. 2011, 77, 1110–1115. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Zhang, L.; Yang, R.; Yang, Y.; Jin, H.; Zhang, X.; Hu, Q.; He, B.; Shen, Z.; Chen, P. Corilagin ameliorates atherosclerosis by regulating MMP-1, -2, and -9 expression in vitro and in vivo. Eur. J. Pharmacol. 2021, 906, 174200. [Google Scholar] [CrossRef]
- Chang, W.C.; Yu, Y.M.; Chiang, S.Y.; Tseng, C.Y. Ellagic acid suppresses oxidised low-density lipoprotein-induced aortic smooth muscle cell proliferation: Studies on the activation of extracellular signal-regulated kinase 1/2 and proliferating cell nuclear antigen expression. Br. J. Nutr. 2008, 99, 709–714. [Google Scholar] [CrossRef] [PubMed]
- Ou, T.T.; Lin, M.C.; Wu, C.H.; Lin, W.L.; Wang, C.J. Gallic acid attenuates oleic acid-induced proliferation of vascular smooth muscle cell through regulation of AMPK-eNOS-FAS signaling. Curr. Med. Chem. 2013, 20, 3944–3953. [Google Scholar] [CrossRef] [PubMed]
- Yılmaz, B.; Usta, C. Ellagic acid-induced endothelium-dependent and endothelium-independent vasorelaxation in rat thoracic aortic rings and the underlying mechanism. Phytother. Res. 2013, 27, 285–289. [Google Scholar] [CrossRef]
- de Oliveira, L.M.; de Oliveira, T.S.; da Costa, R.M.; de Souza Gil, E.; Costa, E.A.; Passaglia Rde, C.; Filgueira, F.P.; Ghedini, P.C. The vasorelaxant effect of gallic acid involves endothelium-dependent and -independent mechanisms. Vasc. Pharmacol. 2016, 81, 69–74. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physiological Parameters | Treatment Group | |||
|---|---|---|---|---|
| ND | HFD | HFD + 25 mg/kg/day Geraniin | HFD + 40 mg/kg/day Captopril | |
| Initial body weight (gram) | 58.5 ± 9.3 | 62.2 ± 10.1 | 56.8 ± 11.5 | 57.8 ± 7.7 |
| Final body weight (gram) | 380.6 ± 31.9 | 444.4 ± 31.3 * | 429.4 ± 25.7 | 407.4 ± 39.1 |
| rWAT to body weight ratio (%) | 2.54 ± 0.82 | 3.66 ± 0.42 * | 3.02 ± 0.66 | 2.73 ± 0.41 |
| Calorie intake (kcal/day) | 60.8 ± 4.3 | 63.0 ± 6.0 | 60.0 ± 6.0 | 54.7 ± 4.1 *, # |
| SBP (mmHg) | 120.2 ± 4.3 | 145.2 ± 4.1 *** | 114.8 ± 9.5 #### | 103.8 ± 14.1 *, #### |
| DBP (mmHg) | 87.2 ± 7.1 | 110.5 ± 7.8 ** | 83.5 ± 11.4 ## | 72.7 ± 14.1 #### |
| Pharmacological Parameters | Treatment Group | |||
|---|---|---|---|---|
| ND | HFD | HFD + 25 mg/kg/day Geraniin | HFD + 40 mg/kg/day Captopril | |
| Vasoconstriction Emax (%) | 125.1 ± 27.4 | 139.0 ± 13.2 | 127.8 ± 10.3 | 150.7 ± 31.5 |
| Vasorelaxation Emax (%) | 95.4 ± 4.5 | 108.9 ± 6.1 | 111.2 ± 4.1 | 108.3 ± 9.0 |
| PE pEC50 | 6.55 ± 0.60 | 6.95 ± 0.42 | 6.70 ± 0.26 | 7.21 ± 0.35 |
| CCH pEC50 | 7.34 ± 0.43 | 7.28 ± 0.77 | 7.27 ± 0.49 | 7.32 ± 0.86 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goh, B.H.; Cheng, H.S.; Alexandra, P.T.A/P.A.; Ting, K.-N.; Palanisamy, U.D.; Tan, J.B.L. Geraniin Ameliorates Hypertensive Vascular Remodelling in a Diet-Induced Obese Animal Model through Antioxidant and Anti-Inflammatory Effects. Nutrients 2023, 15, 2696. https://doi.org/10.3390/nu15122696
Goh BH, Cheng HS, Alexandra PTA/PA, Ting K-N, Palanisamy UD, Tan JBL. Geraniin Ameliorates Hypertensive Vascular Remodelling in a Diet-Induced Obese Animal Model through Antioxidant and Anti-Inflammatory Effects. Nutrients. 2023; 15(12):2696. https://doi.org/10.3390/nu15122696
Chicago/Turabian StyleGoh, Boon Hee, Hong Sheng Cheng, Pricilla Tracy A/P A. Alexandra, Kang-Nee Ting, Uma Devi Palanisamy, and Joash Ban Lee Tan. 2023. "Geraniin Ameliorates Hypertensive Vascular Remodelling in a Diet-Induced Obese Animal Model through Antioxidant and Anti-Inflammatory Effects" Nutrients 15, no. 12: 2696. https://doi.org/10.3390/nu15122696
APA StyleGoh, B. H., Cheng, H. S., Alexandra, P. T. A/P. A., Ting, K.-N., Palanisamy, U. D., & Tan, J. B. L. (2023). Geraniin Ameliorates Hypertensive Vascular Remodelling in a Diet-Induced Obese Animal Model through Antioxidant and Anti-Inflammatory Effects. Nutrients, 15(12), 2696. https://doi.org/10.3390/nu15122696