
Elucidation of the Anticancer Mechanism of Durian Fruit (Durio zibethinus) Pulp Extract in Human Leukemia (HL-60) Cancer Cells

 , , , , , ,
, , , , , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Fruit Extracts
2.2. Cell Culture HL-60
2.3. Cell Cycle Assay
2.4. Annexin V-FITC and PI Staining
2.5. DNA Fragmentation Assay
2.6. Comet Assay
2.7. Western Blotting Analysis of Apoptotic Protein Expression
2.8. Statistical Analysis
3. Results
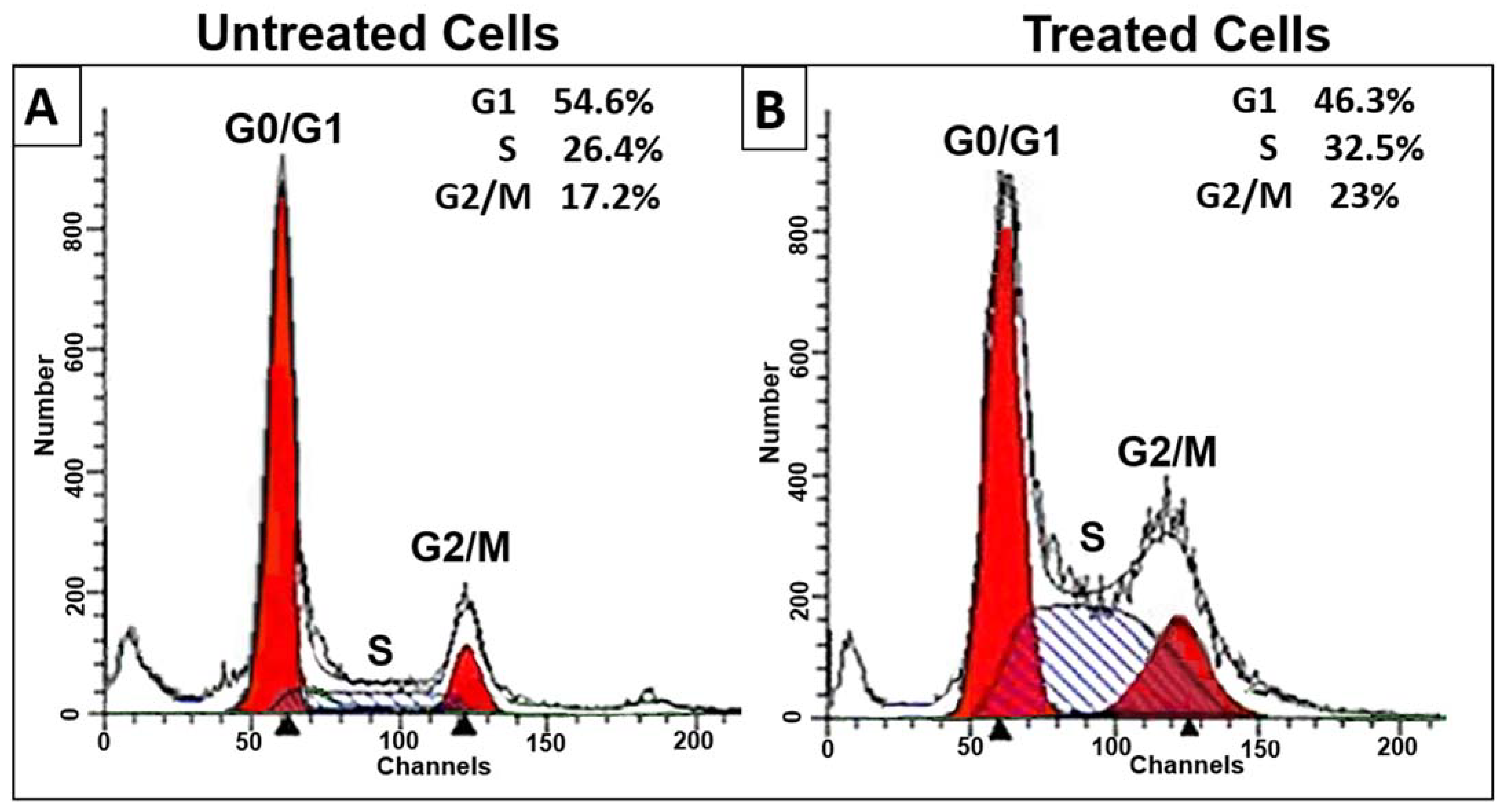
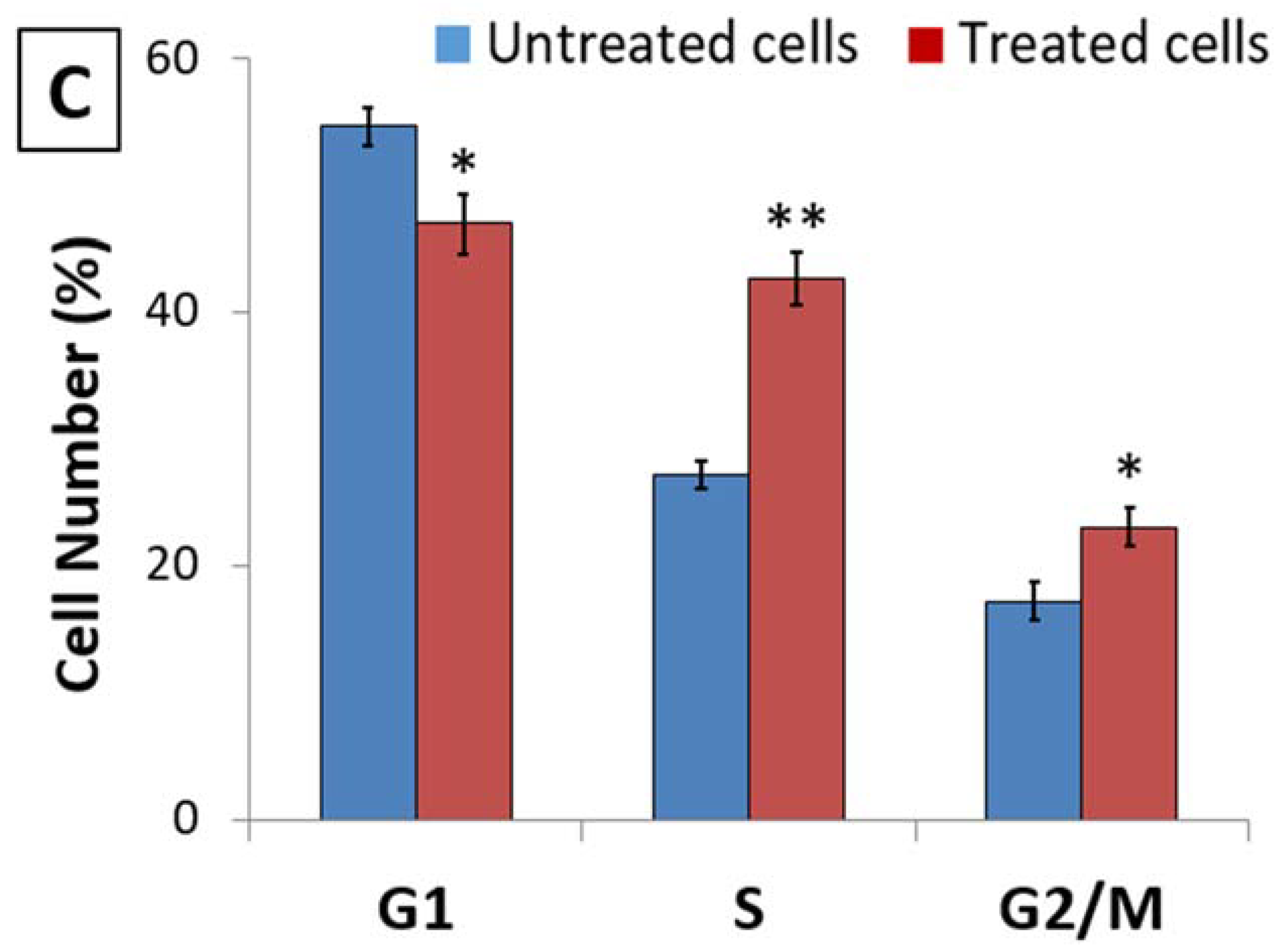
3.1. D. zibethinus Methanol Extracts Induced Cell Cycle Arrest in HL-60 Cells
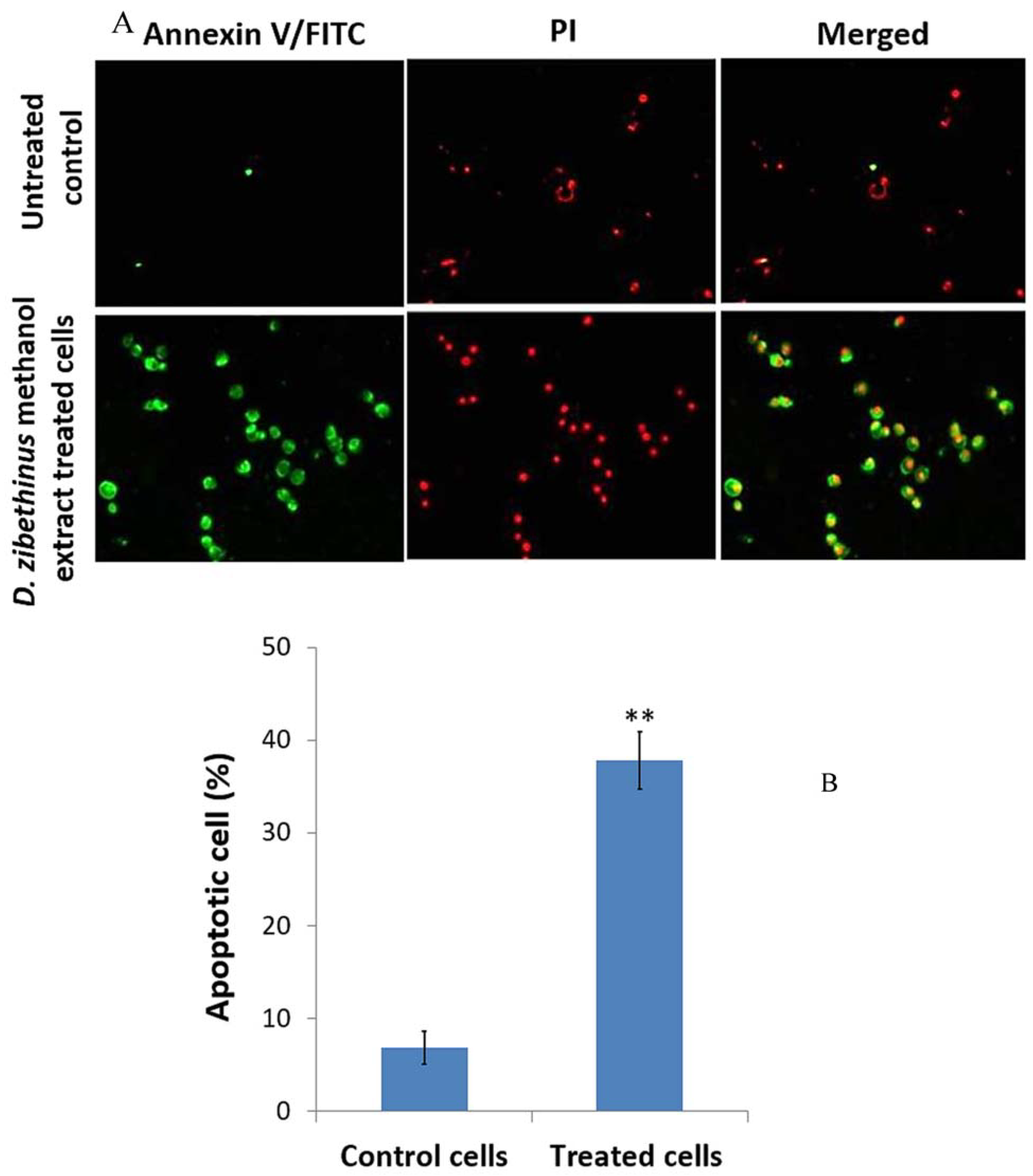
3.2. D. zibethinus Methanol Extracts Induced Apoptosis in HL-60 Cells
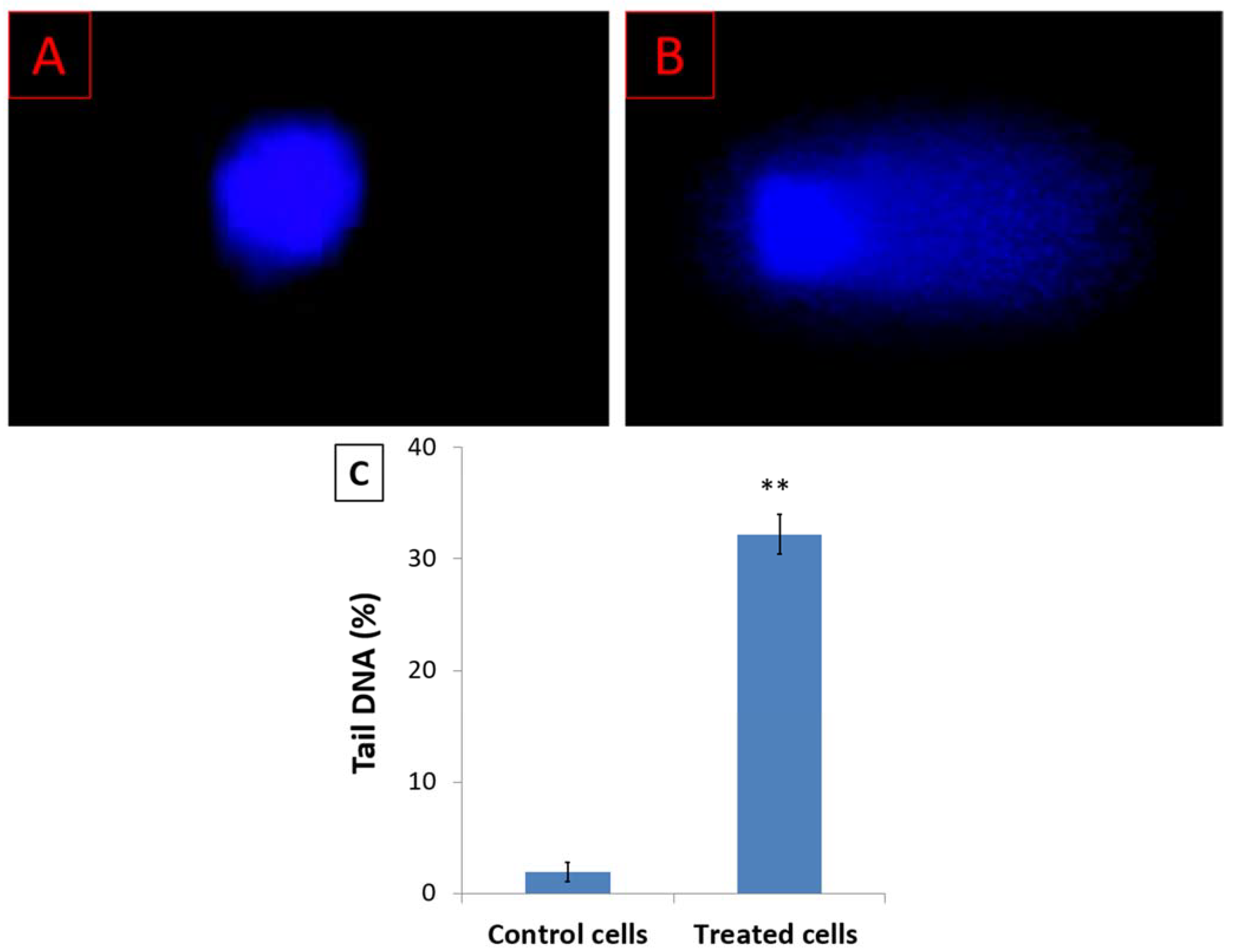
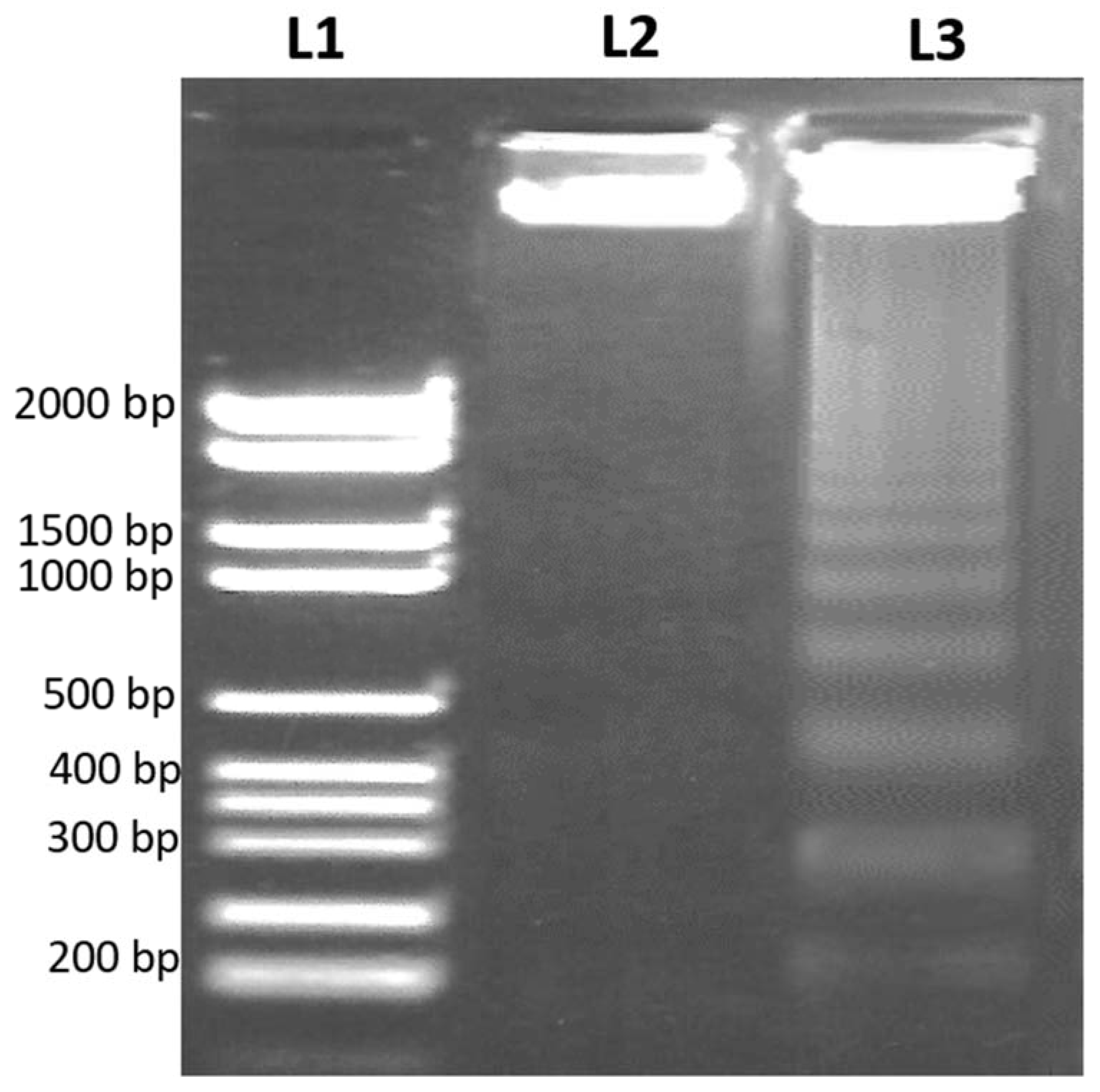
3.3. D. zibethinus Methanol Extracts Induced DNA Damage in HL-60 Cells
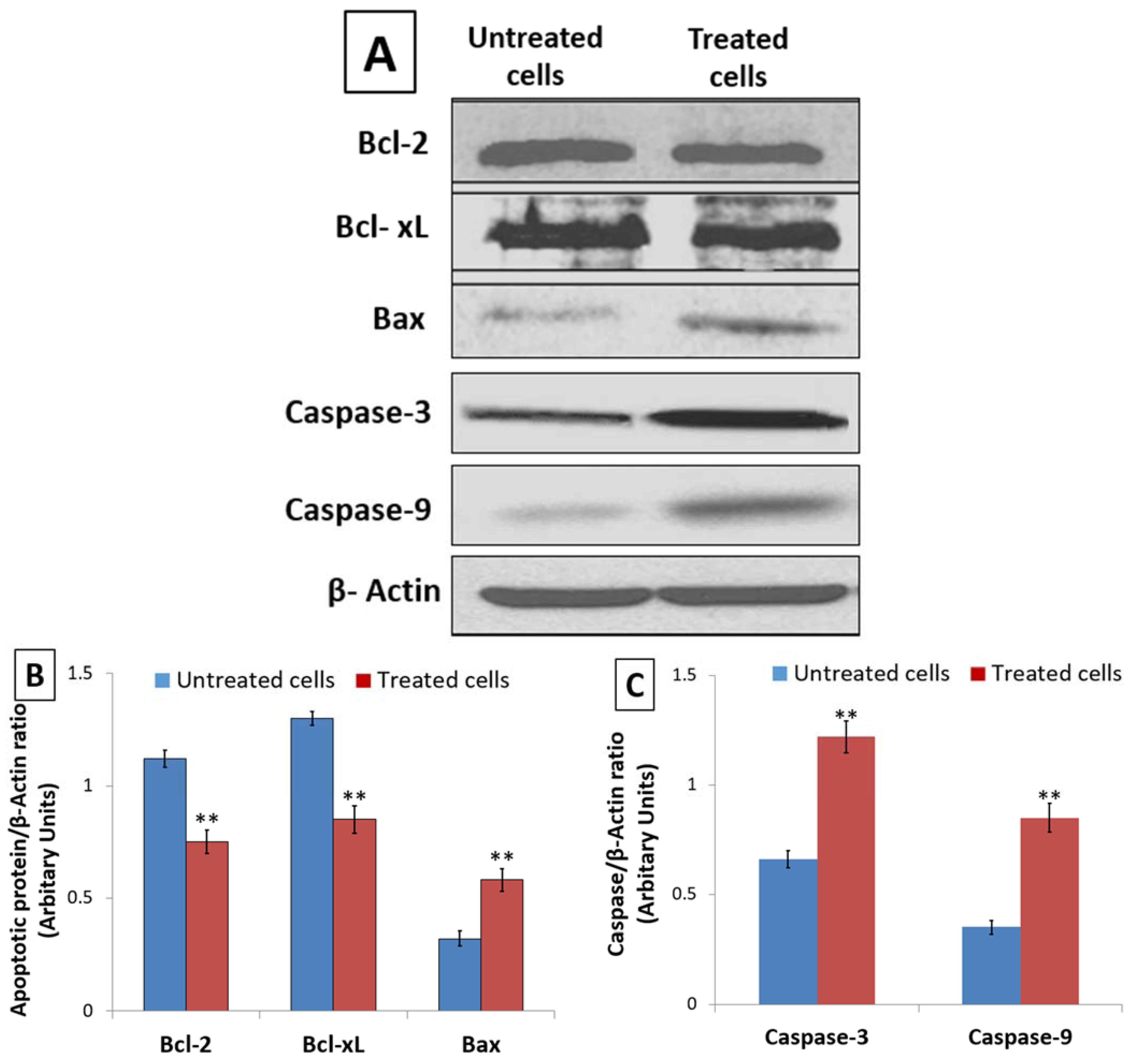
3.4. Effect of D. zibethinus Extract Treatment on Apoptotic Protein Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hemalatha, G.; Sivakumari, K.; Rajesh, S.; Shyamala Devi, K. Phytochemical profiling, anti-cancer and apoptotic activity of graviola (Annona muricata) fruit extract against human hepatocellular carcinoma(HepG-2) cells. Int. J. Zool. Appl. Biosci. 2020, 5, 32–47. [Google Scholar]
- Kainsa, S.; Kumar, P.; Rani, P. Medicinal Plants of Asian Origin Having Anticancer Potential: Short Review. Asian J. Biomed. Pharm. Sci. 2012, 2, 1–7. [Google Scholar]
- Devalaraja, S.; Jain, S.; Yadav, H. Exotic fruits as therapeutic complements for diabetes, obesity and metabolic syndrome. Food Res. Int. 2011, 44, 1856–1865. [Google Scholar] [CrossRef] [PubMed]
- Ho, L.-H.; Bhat, R. Exploring the potential nutraceutical values of durian (Durio zibethinus L.)—An exotic tropical fruit. Food Chem. 2015, 168, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Leontowicz, M.; Leontowicz, H.; Jastrzebski, Z.; Jesion, I.; Haruenkit, R.; Poovarodom, S.; Katrich, E.; Tashma, Z.; Drzewiecki, J.; Trakhtenberg, S.; et al. The nutritional and metabolic indices in rats fed cholesterol-containing diets supplemented with durian at different stages of ripening. Biofactors 2007, 29, 123–136. [Google Scholar] [CrossRef]
- Leontowicz, H.; Leontowicz, M.; Haruenkit, R.; Poovarodom, S.; Jastrzebski, Z.; Drzewiecki, J.; Ayala, A.L.; Jesion, I.; Trakhtenberg, S.; Gorinstein, S. Durian (Durio zibethinus Murr.) cultivars as nutritional supplementation to rat’s diets. Food Chem. Toxicol. 2008, 46, 581–589. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Maah, M.J.; Yusoff, I. Estimation of antioxidant phytochemicals in four different varieties of Durian (Durio zibethinus Murray) fruit. Middle-East J. Sci. Res. 2010, 6, 465–471. [Google Scholar]
- Haruenkit, R.; Poovarodom, S.; Leontowicz, H.; Leontowicz, M.; Sajewicz, M.; Kowalska, T.; Delgado-Licon, E.; Rocha-Guzmán, N.E.; Gallegos-Infante, J.-A.; Trakhtenberg, S.; et al. Comparative Study of Health Properties and Nutritional Value of Durian, Mangosteen, and Snake Fruit: Experiments In vitro and In vivo. J. Agric. Food Chem. 2007, 55, 5842–5849. [Google Scholar] [CrossRef]
- Haruenkit, R.; Poovarodom, S.; Vearasilp, S.; Namiesnik, J.; Sliwka-Kaszynska, M.; Park, Y.-S.; Heo, B.-G.; Cho, J.-Y.; Jang, H.G.; Gorinstein, S. Comparison of bioactive compounds, antioxidant and antiproliferative activities of Mon Thong durian during ripening. Food Chem. 2010, 118, 540–547. [Google Scholar] [CrossRef]
- Jayakumar, R.; Kanthimathi, M. Inhibitory effects of fruit extracts on nitric oxide-induced proliferation in MCF-7 cells. Food Chem. 2011, 126, 956–960. [Google Scholar] [CrossRef]
- Liu, X.; Ye, F.; Wu, J.; How, B.; Li, W.; Zhang, D.Y. Signaling Proteins and Pathways Affected by Flavonoids in Leukemia Cells. Nutr. Cancer 2015, 67, 238–249. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-W.; Chung, K.-S.; Seo, J.-H.; Yim, S.-V.; Park, H.-J.; Choi, J.-H. Sulfuretin from heartwood of Rhus verniciflua triggers apoptosis through activation of Fas, Caspase-8, and the mitochondrial death pathway in HL-60 human leukemia cells. J. Cell. Biochem. 2012, 113, 2835–2844. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhao, X.H.; Wang, Z.J. Flavones and flavonols exert cytotoxic effects on a human oesophageal adenocarcinoma cell line (OE33) by causing G2/M arrest and inducing apoptosis. Food Chem. Toxicol. 2008, 46, 2042–2053. [Google Scholar] [CrossRef] [PubMed]
- Sak, K.; Everaus, H. Established Human Cell Lines as Models to Study Anti-leukemic Effects of Flavonoids. Curr. Genom. 2017, 18, 3–26. [Google Scholar] [CrossRef] [PubMed]
- Mohamad Sitheek, A.; Sivakumari, K.; Rajesh, S.; Ashok, K. Anti-Proliferative Effect of Durian fruit (Durio zibenthinus) Against HL-60 cells and its phytochemical analysis. J. Adv. Sci. Res. 2020, 11 (Suppl. 5), 174–181. [Google Scholar]
- Rajesh, S.; Sivakumari, K.; Ashok, K.; Abitha, A.R. Anti-cancer activity of Cardiospermum halicacabum Linn. Leaf extracts against human breast cancer cell line (MCF-7). Eur. J. Biomed. Pharmaceu. Sci. 2016, 3, 213–222. [Google Scholar]
- Rieger, A.M.; Nelson, K.L.; Konowalchuk, J.D.; Barreda, D.R. Modified annexin V/propidium iodide apoptosis assay for ac-curate assessment of cell death. J. Vis. Exp. 2011, 50, 2597. [Google Scholar]
- Goud, T.S.; Upadhyay, R.C.; Kumar, A.; Karri, S.; Choudhary, R.; Ashraf, S.; Singh, S.V.; Kumar, O.S.; Kiranmai, C. Novel extraction of high quality genomic DNA from frozen bovine blood samples by using detergent method. Open Veter. J. 2018, 8, 415–422. [Google Scholar] [CrossRef]
- Singh, N.P.; McCoy, M.T.; Tice, R.R.; Schneider, E.L. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp. Cell Res. 1988, 175, 184–191. [Google Scholar] [CrossRef]
- Ravishankar, D.; Rajora, A.K.; Greco, F.; Osborn, H.M. Flavonoids as prospective compounds for anti-cancer therapy. Int. J. Biochem. Cell Biol. 2013, 45, 2821–2831. [Google Scholar] [CrossRef]
- Pilátová, M.; Stupáková, V.; Varinská, L.; Sarisský, M.; Mirossay, L.; Mirossay, A.; Gál, P.; Kraus, V.; Dianisková, K.; Mojzis, J. Effect of selected flavones on cancer and endothelial cells. Gen. Physiol. Biophys. 2010, 29, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Henamayee, S.; Banik, K.; Sailo, B.L.; Shabnam, B.; Harsha, C.; Srilakshmi, S.; Vgm, N.; Baek, S.H.; Ahn, K.S.; Kunnumakkara, A.B. Therapeutic Emergence of Rhein as a Potential Anticancer Drug: A Review of Its Molecular Targets and Anticancer Properties. Molecules 2020, 25, 2278. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.W.; Yang, Y.; Zhang, K.; Qiang, L.; Yang, L.; Yang, L.; Hu, Y.; Wang, X.T.; You, Q.D.; Guo, Q.L. Wogonin induced differentiation and G1 phase arrest of human U-937 leukemia cells via PKCdelta phosphorylation. Eur. J. Pharmacol. 2008, 591, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Poon, R.Y.C. Cell Cycle Control: A System of Interlinking Oscillators. Methods Mol. Biol. 2021, 2329, 1–18. [Google Scholar] [CrossRef]
- Barnum, K.J.; O’Connell, M.J. Cell cycle regulation by checkpoints. In Methods in Molecular Biology; Springer: Berlin/Heidelberg, Germany, 2014; pp. 29–40. [Google Scholar]
- Schwartz, G.K.; Shah, M.A. Targeting the Cell Cycle: A New Approach to Cancer Therapy. J. Clin. Oncol. 2005, 23, 9408–9421. [Google Scholar] [CrossRef]
- Rao, P.C.; Begum, S.; Sahai, M.; Sriram, D.S. Coptisine-induced cell cycle arrest at G2/M phase and reactive oxygen species-dependent mitochondria-mediated apoptosis in non-small-cell lung cancer A549 cells. Tumour Biol. 2017, 39, 1010428317694565. [Google Scholar] [CrossRef]
- Jovanović, M.; Tenji, D.; Nikolić, B.; Srdić-Rajić, T.; Svirčev, E.; Mitić-Ćulafić, D. In Vitro Study of Two Edible Polygonoideae Plants: Phenolic Profile, Cytotoxicity, and Modulation of Keap1-Nrf2 Gene Expression. Foods 2021, 10, 811. [Google Scholar] [CrossRef]
- Brown, M.J. Durio—A Bibliographic Review; Arora, R.K., Ramanatha Rao, V., Rao, A.N., Eds.; International Plant Genetic Resource Institute, IPGRI Office for South Asia: New Delhi, India, 1997; pp. 1–126. [Google Scholar]
- Sergedien, E.; Jönsson, K.; Szymusiak, H.; Tyrakowska, B.; Rietjens, I.; ČNas, N. Prooxidant toxicity of polyphenolic antioxidants to HL-60 cells: Description of quantitative structure-activity relationships. FEBS Lett. 1999, 462, 392–396. [Google Scholar] [CrossRef]
- Singh, M.; Singh, R.; Bhui, K.; Tyagi, S.; Mahmood, Z.; Shukla, Y. Tea polyphenols induce apoptosis through mitochondrial pathway and by inhibiting nuclear factor-kappaB and Akt activation in human cervical cancer cells. Oncol. Res. 2011, 19, 245–257. [Google Scholar] [CrossRef]
- Collins, J.A.; Schandl, C.A.; Young, K.K.; Vesely, J.; Willingham, M.C. Major DNA Fragmentation Is a Late Event in Apoptosis. J. Histochem. Cytochem. 1997, 45, 923–934. [Google Scholar] [CrossRef]
- Patel, A.; Soni, A.; Siddiqi, N.J.; Sharma, P. An insight into the anticancer mechanism of Tribulus terrestris extracts on human breast cancer cells. 3 Biotech 2019, 9, 58. [Google Scholar] [CrossRef] [PubMed]
- Sevrioukova, I.F. Apoptosis-Inducing Factor: Structure, Function, and Redox Regulation. Antioxid. Redox Signal. 2011, 14, 2545–2579. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdul Rahman, M.S.; Kanakarajan, S.; Selvaraj, R.; Kamalanathan, A.; Fatima, S.; Abudawood, M.; Siddiqi, N.J.; Alanazi, H.; Sharma, B.; de Lourdes Pereira, M. Elucidation of the Anticancer Mechanism of Durian Fruit (Durio zibethinus) Pulp Extract in Human Leukemia (HL-60) Cancer Cells. Nutrients 2023, 15, 2417. https://doi.org/10.3390/nu15102417
Abdul Rahman MS, Kanakarajan S, Selvaraj R, Kamalanathan A, Fatima S, Abudawood M, Siddiqi NJ, Alanazi H, Sharma B, de Lourdes Pereira M. Elucidation of the Anticancer Mechanism of Durian Fruit (Durio zibethinus) Pulp Extract in Human Leukemia (HL-60) Cancer Cells. Nutrients. 2023; 15(10):2417. https://doi.org/10.3390/nu15102417
Chicago/Turabian StyleAbdul Rahman, Mohamad Sitheek, Sivakumari Kanakarajan, Rajesh Selvaraj, Ashok Kamalanathan, Sabiha Fatima, Manal Abudawood, Nikhat J. Siddiqi, Humidah Alanazi, Bechan Sharma, and Maria de Lourdes Pereira. 2023. "Elucidation of the Anticancer Mechanism of Durian Fruit (Durio zibethinus) Pulp Extract in Human Leukemia (HL-60) Cancer Cells" Nutrients 15, no. 10: 2417. https://doi.org/10.3390/nu15102417
APA StyleAbdul Rahman, M. S., Kanakarajan, S., Selvaraj, R., Kamalanathan, A., Fatima, S., Abudawood, M., Siddiqi, N. J., Alanazi, H., Sharma, B., & de Lourdes Pereira, M. (2023). Elucidation of the Anticancer Mechanism of Durian Fruit (Durio zibethinus) Pulp Extract in Human Leukemia (HL-60) Cancer Cells. Nutrients, 15(10), 2417. https://doi.org/10.3390/nu15102417